Flow cytometric quantification of STING-NP cellular uptake and qPCR analysis of STING-NP-treated cell subsets. STING-NP demonstrates similar activity in human tissues ..91 4.1 Potential targets for synergistic combination therapy with STING-NP.
Cancer Immunity and the State of Patient Care
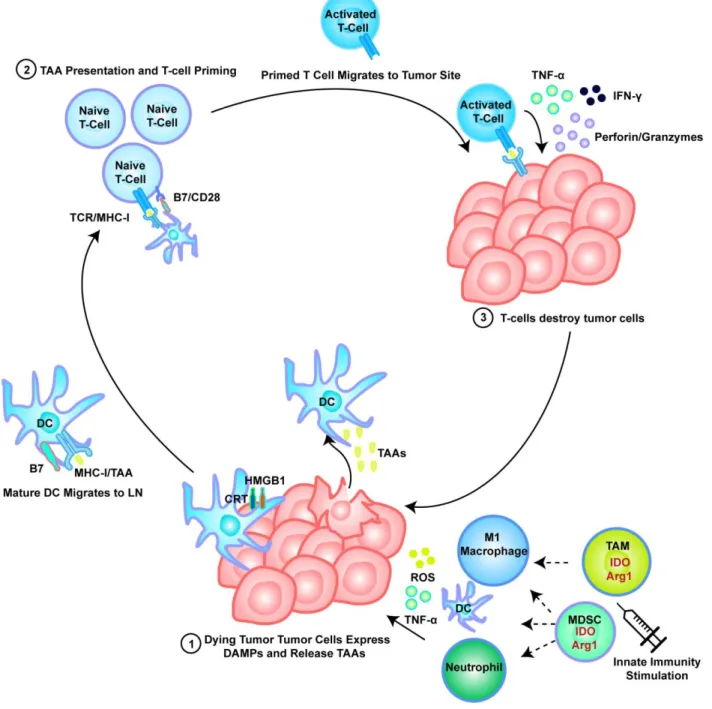
The remaining 42% of patients are thought to show poor sensitivity to checkpoint blockade and exhibit low TIL numbers, indicating a phenotype in which the immune system has failed to mount an antigen-specific T-cell response of any appreciable magnitude. Second, treatments will need to prime de novo T cell responses in the significant proportion of patients who do not achieve endogenous antitumor T cell mobilization.
The Immunosuppressive Tumor Microenvironment Inhibits Tumoricidal
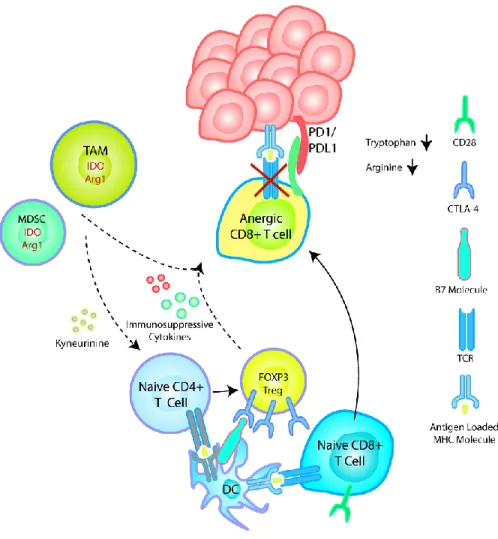
First, it is necessary to continue developing strategies that abrogate a broader set of immunosuppressive mechanisms in the TME to fully unleash the potential of antitumor T cells. Expression of IDO and Arg1 depletes tryptophan and arginine in the TME, leading to loss of CD3ζ expression and loss of TCR-dependent signal transduction.
Innate Immunity and Priming of Adaptive Immunity
High levels of immunosuppressive cytokines such as IL-10 found in the TME suppress maturation of DCs, decrease their expression of MHC and co-stimulatory molecules, inhibit their T-cell priming ability.30-31 The tumor immunosuppressive program disrupts the steady state state balance of GS production and systemic maturation, and lower numbers of both GSs in the periphery and in the LNs are typically found in tumor-bearing patients compared to healthy cohorts.32 As expected, this decrease in immune supportive DCs is typically accompanied with an increase in immature and monocytic DC progenitors throughout the host which then perpetuates an immunosuppressive feedback loop by expressing molecules such as IL-10, IDO, Arg-1 and various checkpoint molecules.33 In addition to suppressing T-cell function in the TME, this effect is particularly deleterious in the LN, in which immunosuppressive DCs and DC progenitors fail to generate effector cytotoxic T lymphocytes (CTLs) while actively promoting the expansion of Tregs.34. This provides further motivation for the development of strategies to reprogram myeloid cells in the TME to a proinflammatory phenotype.
Exploiting the cGAS/STING Pathway in Cancer Immunotherapy
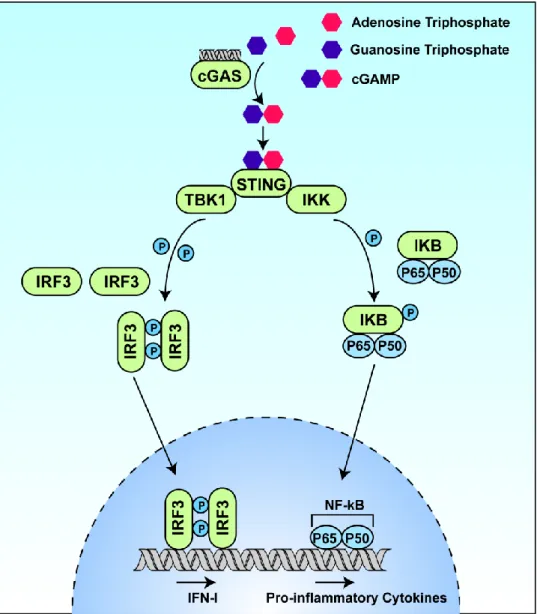
In connection with endogenous cancer detection, cGAMP is produced intracellularly for STING recognition of cGAS upon detection of dsDNA in the cytosol. These strategies represent critical advances in STING-dependent cancer immunotherapy, but nevertheless only mitigate the delivery challenges associated with CDN delivery.
Developing Nanoparticle Delivery Platforms to Improve CDN Delivery
Achieving efficient delivery of nanoparticles to the TDLN involves exploiting differences in vascular and lymphatic transport in the TME. Vascular defects in the vicinity of the TME present an opportunity to enhance nanoparticle accumulation at tumor sites through dilation.
Aims and Scope
In addition to tumor rejection, treatment produces long-lasting, systemic antitumor immunity, as evidenced by elicited resistance to rechallenge with subsequent inoculation of tumor cells. Special emphasis is placed on characterizing the effect of STING-NP on the tumor microenvironment.
Introduction
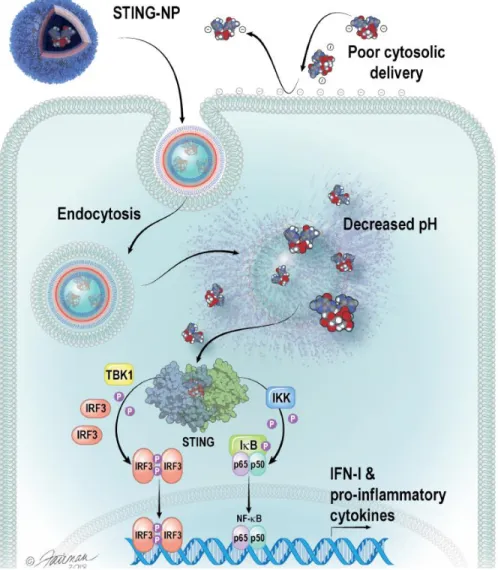
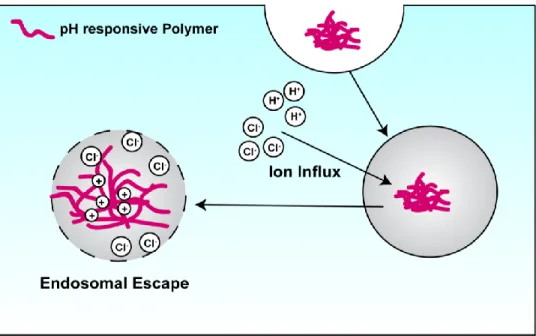
Specifically, STING-NPs consist of a hydrophilic poly(ethylene glycol) (PEG) chemical block commonly used to inhibit protein absorption and particle opsonization68 linked to a hydrophobic and pH-responsive poly[(2-diethylaminoethyl methacrylate)-co-(butyl) methacrylate)-co-(pyridyl disulfide ethyl methacrylate)] (DBP) block copolymer that was optimized for disruption of the endosomal membrane. Where free cGAMP molecules are repelled by the negatively charged cell membrane, STING NPs are endocytosed.
Design, Synthesis, and Optimization of STING-NPs
STING-NPs comprise cGAMP molecules encapsulated by pH-responsive and endosomolytic PEG-DBP polymers self-assembled into a PEGylated vesicular structure. STING-NPs comprise PEG-DBP polymer chains, which contain a pH-responsive (DEAEMA), membrane-destabilizing (DEAEMA + BMA) and cross-linked (PDSMA) hydrophobic block.
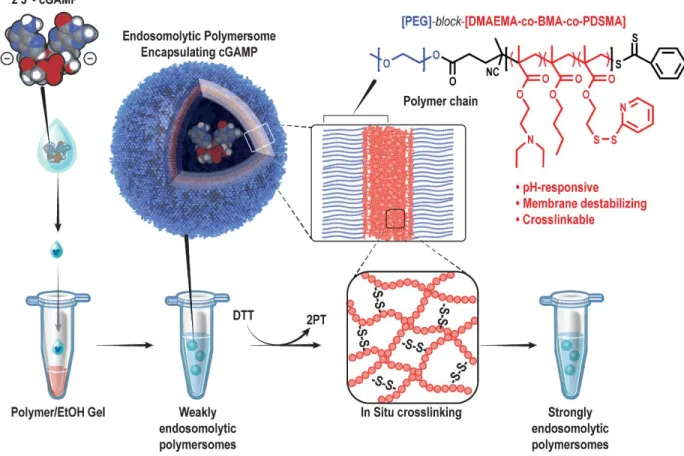
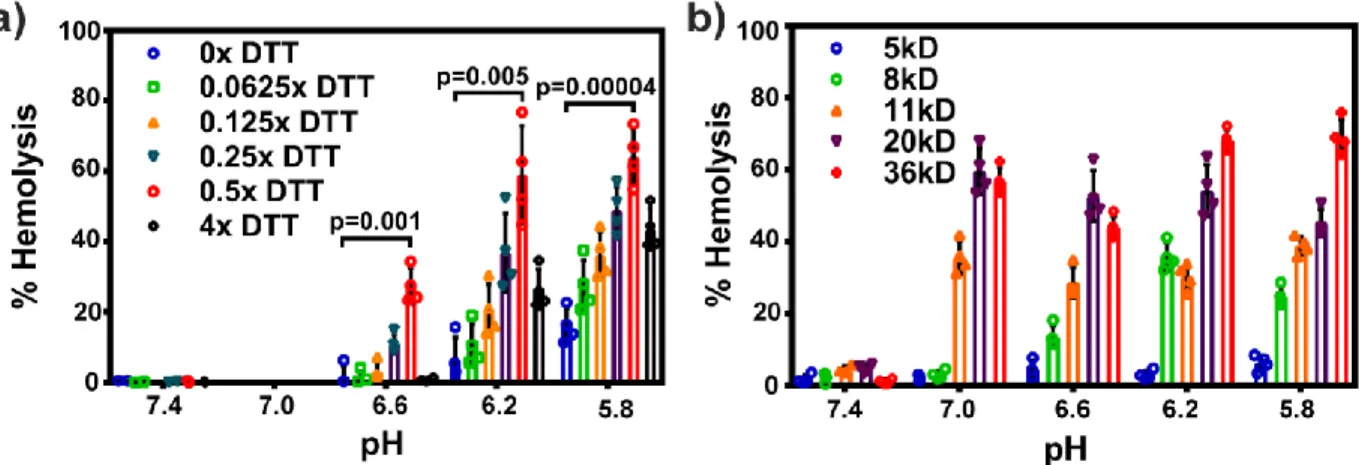
In Situ Crosslinking is Critical to Achieving Endosomolytic Potency and
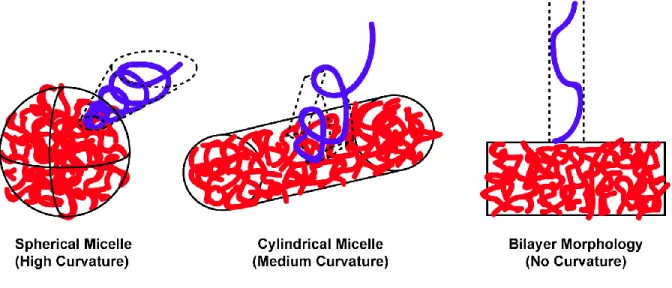
Similarly, we assume that the density of the hydrophobic material is not affected by its self-assembling morphology. A second reason why higher molecular weight polymer chains prefer spherical and cylindrical structures relates to the interfacial area of the particles and the degree of elongation found in the hydrophobic core blocks.
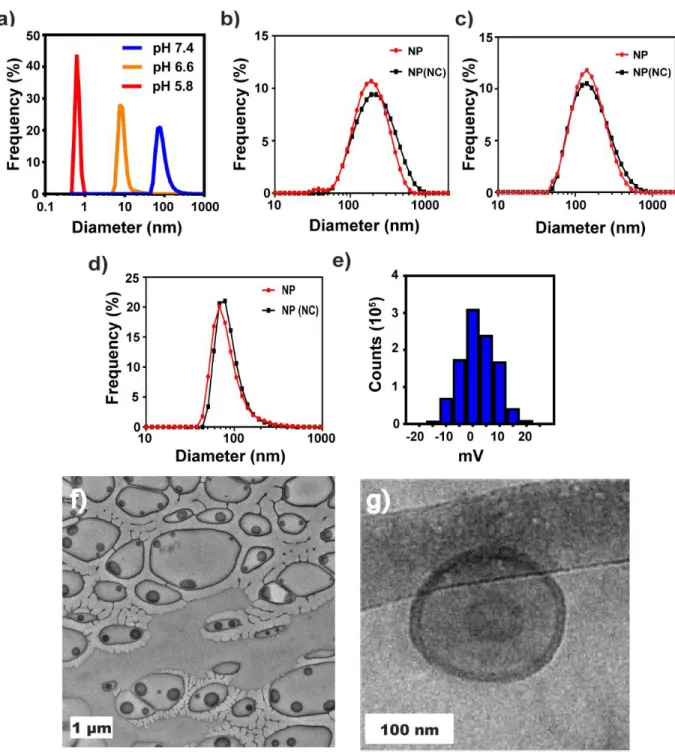
Evaluation of the Direct Hydration Method for cGAMP Encapsulation
Where Ve is the encapsulated volume, R is the radius of the repeating sphere and T is the thickness of the vesicle bilayer. Where MT represents the total mass of the polymer in the system, WT represents the weight fraction of the polymer in nonsolvent at the point of vesicle formation, and ρpol and ρsolvent represent the density of the polymer and nonsolvent, respectively. Thus, we expect some contribution to the measured encapsulation efficiency in STING NPs due to loading of cGAMP into the hydrophobic phase of the material.
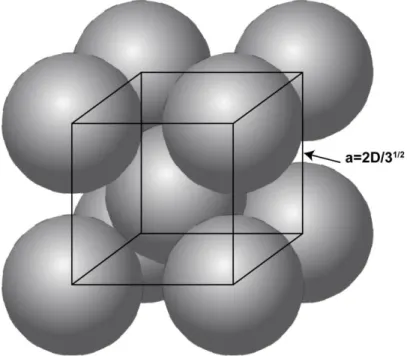
In Vitro Validation of STING-NP Activity
Quantification of IFN-I via ELISA in DC 2.4-treated dendritic cells revealed similar trends to those described above, with STING-NP acting as a much more potent activator of IFN-I production than free cGAMP (Figure 2.11 a). Consistent with data in cell lines, STING-NPs were more potent inducers of DC maturation marker expression than was free cGAMP. These data demonstrate that STING-NPs induce desirable phenotypic changes in cell types that are implicated in the failure of antitumor immunity in the TME.
Conclusion
This motivated further testing of STING-NP in the most representative immature primary DCs and immunosuppressive macrophages: two cell types that we expect are particularly important in the TME. STING-NPs induced repolarization to a pro-inflammatory phenotype in macrophages and maturation in DCs, suggesting that STING-NPs have the potential to reprogram immunosuppressive elements in the TME that can inhibit antitumor immunity as well as jump start the of DC-mediated antigen presentation. for priming an antigen-specific T cell response. Collectively, these data indicate that STING-NPs are a promising immunotherapeutic for the treatment of tumors, which is explored in more detail in the next chapter.
Materials and Methods
The solvent was removed by rotary evaporation to yield a gum residue which was dissolved in 15 mL of dichloromethane (DCM). 1.6 mL, 20 mmol) was added and the solvent was removed by rotary evaporation. After stirring for 2 min, anhydrous tert -butyl hydroperoxide (5.5 M in decane, 0.55 mL, 3 mmol) was added and the reaction mixture was stirred for an additional 30 min at room temperature. The reaction mixture was stirred for 10 min at room temperature and quenched with pyridine (10 mL).
Introduction
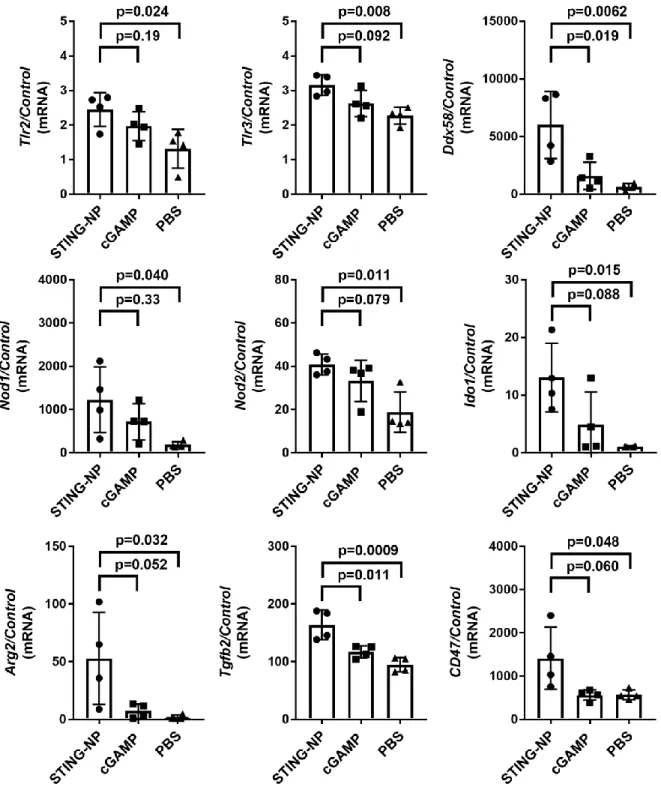
No return to baseline levels was seen in the investigated time frame for STING-NP treated tumors. In arrayed genes, we observed a consistent 5-10-fold increase in gene expression in mice treated with STING-NP versus cGAMP, with the exception of Cxcl1, a neutrophil chemokine, (35-fold) and Ifna2 (20-fold) . This was further confirmed by unsupervised hierarchical clustering of genes with significantly different expression levels relative to vehicle control, which revealed similar gene clusters between cGAMP and STING-NP treated tumors (Figure 3.3c).
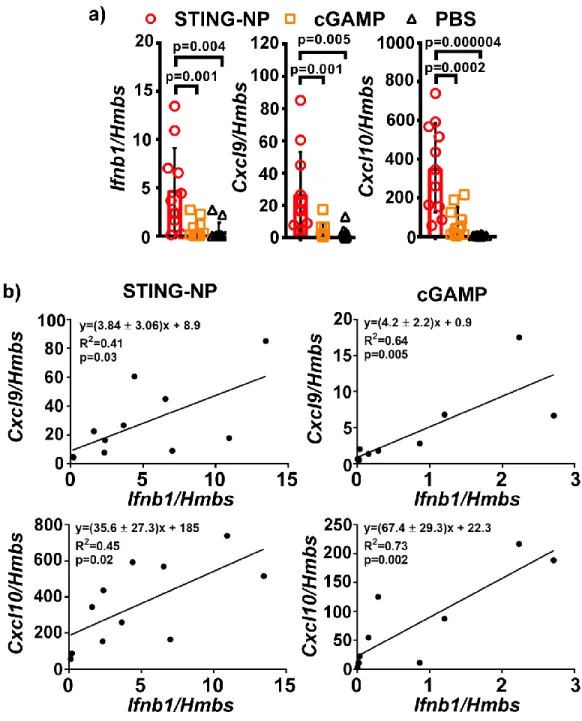
Nanoparticle Formulation Increases Cellular Uptake of CDNs
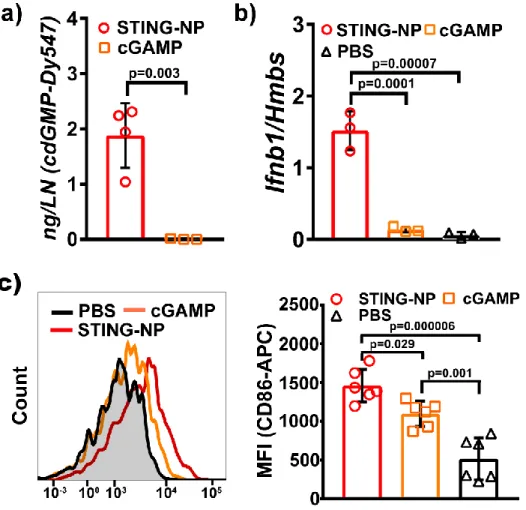
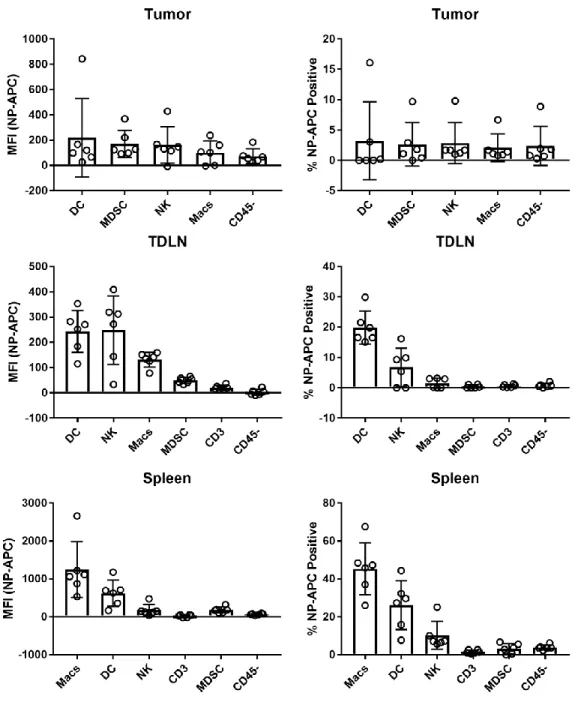
Among these cells, the highest levels of cdGMP-Dy547 were detected in NK cells, DCs, and macrophages, which we considered to be the most active consumers of STING-NPs in the TME. DCs (DC2.4), macrophages (RAW264.7), splenic NK cells, and B16.F10 melanoma cells were cultured in vitro and incubated with STING-NPs for 4 h. Collectively, these data suggest that macrophages and DCs are the primary immunocellular targets of STING-NPs.

STING-NPs Traffic to the Tumor Draining Lymph Node, Facilitating
TDLNs were harvested 2 h post-injection for qPCR analysis, indicating that increased lymphatic trafficking of STING-NPs was associated with increased expression of Ifnb1 as expected. Effects of STING-NP treatment could be seen in the TDLN as far as 48 h from the time of injection, as flow cytometric analysis of DCs harvested from the TDLN showed significantly more surface expression of the costimulatory marker CD86, which is both critical for T-cell priming and a marker of a mature DC phenotype in what is typically an immunosuppressed environment (Figure 3.5c). Intratumoral administration of STING-NP is not associated with the onset of long-term toxicological side effects.
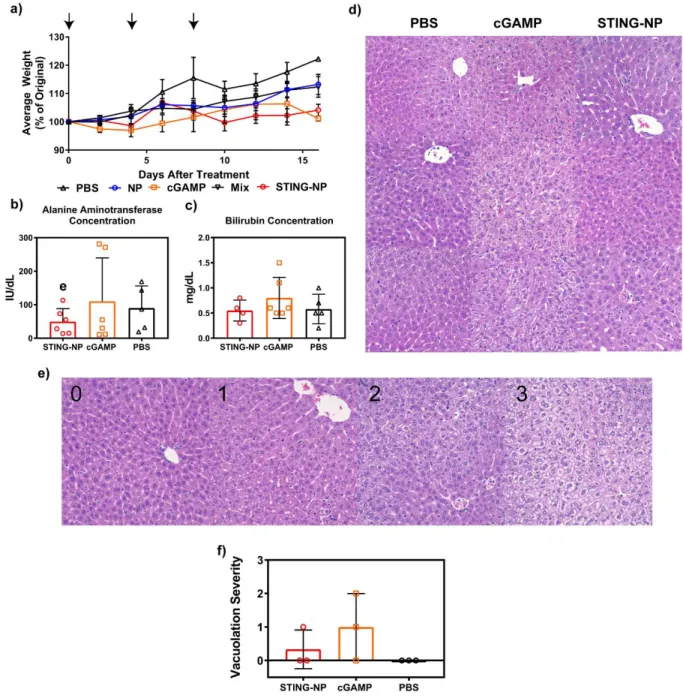
Intratumoral Administration of STING-NP is Not Associated With Onset
Potential long-term toxicological effects of STING-NPs were further investigated by blood chemistry and histological analysis focusing particularly on pathology of the liver, the organ through which the majority of STING-NP clearance is expected to occur. Nevertheless, these data again show no clear evidence of STING-NP-driven liver toxicity (Figure 3.6b). Strikingly, hepatocellular vacuolation was also observed in cGAMP-treated mice, indicating that induction of liver pathology was not a unique consequence of STING-NP treatment.
STING-NPs Treatment Drives an Influx of Activated T-cells and Myeloid
Tumors were isolated and characterized via flow cytometry 2 days after treatment. a) STING-NP treatment induces particularly high magnitudes of neutrophil infiltration in the TME after IT treatment. Higher numbers of m-MDSCs and g-MDSCs are also found in the TME after treatment, to a lesser extent. Both CD4 + and CD8 + T cells are able to produce more TNF-α and IFN-γ after STING-NP treatment.
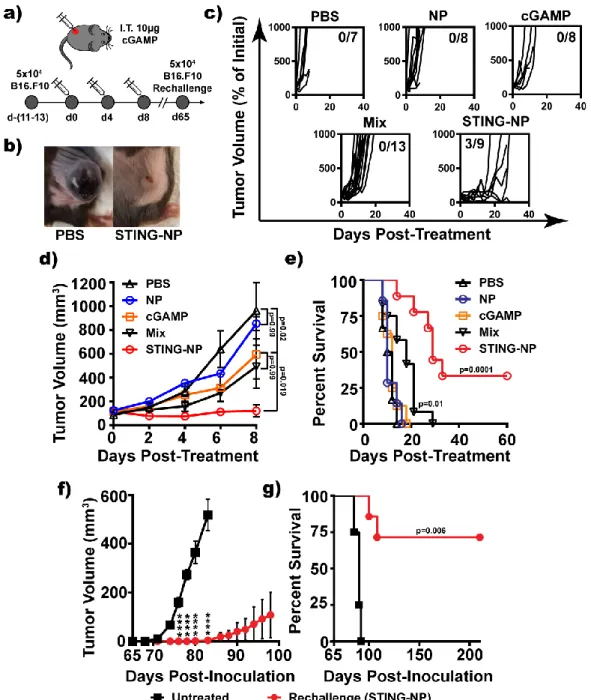
In Situ Vaccination with STING-NPs Generates Sustained and Systemic
Furthermore, these data further demonstrate that the PEG2k-DBP4,5k polymers are largely biologically inactive in the STING-NP formulation and that the therapeutic efficacy is primarily an effect of the improved cGAMP delivery efficiency in the STING-NP formulation. STING-NP treatment significantly delayed the growth of untreated contralateral tumors relative to PBS (DTPBS, ~1.9d, DTSTING-NP ~3.0d), although to a lesser extent than the treated tumor (DTPBS ~1.7d, DTSTING-NP ~ 5.3d). These data are consistent with the priming of circulating antitumor T cells by IT administration of STING-NPs, although the induced immune response does not appear to be able to reject preformed distal tumors without local administration of STING-NPs.
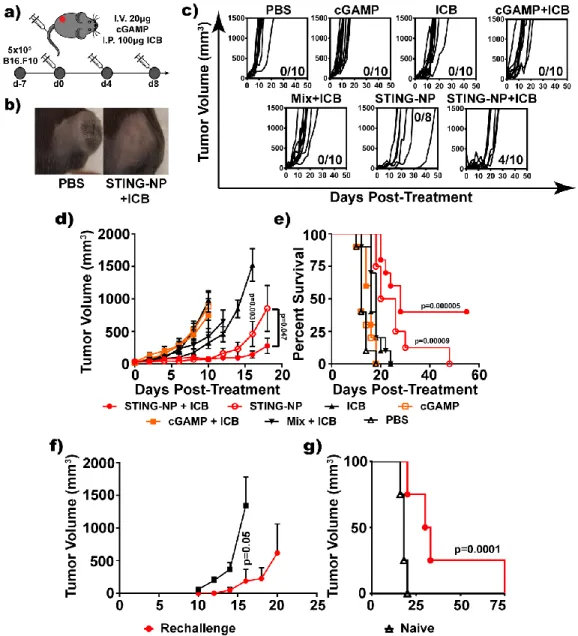
STING-NPs are Effective Antitumor Immunotherapeutics when
Intravenous administration of STING-NP inhibits tumor growth and enhances the efficacy of ICB. a) Scheme of systemic treatment. A unique route of IV administration, however, was the occurrence of neutrophilic infiltration in the liver after STING-NP treatment. Although this is sometimes interpreted as a symptom of hepatocellular injury, due to the established ability of cells exposed to STING-NPs to produce a large amount of the neutrophil chemokine Cxcl1.
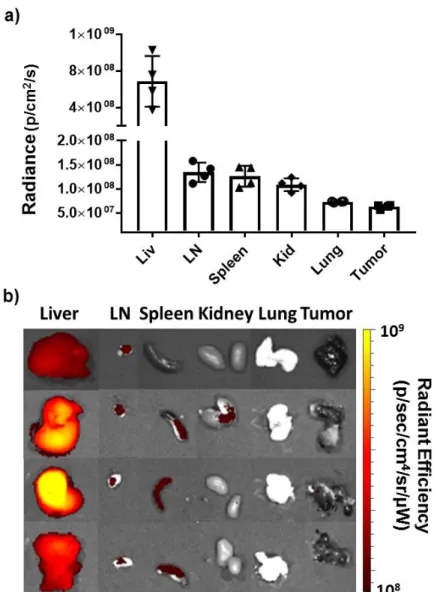
Evaluation of Biodistribution Following Systemic STING-NP
NP accumulation in the liver and kidney compartments is likely a simple consequence of particle uptake and clearance. The apparent lack of particle accumulation in the tumor suggests that IV STING-NP administration does not directly activate STING in the TME. As expected, accumulation of STING-NPs in the TDLN is accompanied by upregulation of Ifnb1 and Cxcl10.
Cellular Uptake of STING-NPs in the Tumor and Lymphoid Organs
Flow cytometric analysis of STING-NP accumulation in cells in tumor and lymphoid organs. Analogous signaling may be insufficient without the accumulation of large amounts of STING-NPs in the TME in case of IV administration. Instead, most STING-NPs may accumulate in the TDLN through uptake by circulating DC and DC precursors, which then migrate to the TDLN after STING-induced maturation.
STING-NPs Stimulate Pro-Inflammatory Signaling in Resected Human
Consistent with findings in mouse models, STING-NPs showed superior immunostimulatory activity, increasing the expression of Ifnb1 (48-352-fold) and Tnf (4-5-fold) and Cxcl10 (15-23-fold), a chemokine that correlates with T cell infiltration in human metastatic melanoma. These data are relatively preliminary, as the ex vivo experimental design abrogates any effects of STING-NP treatment to mediate cell recruitment, whether tumoricidal or proinflammatory, and does not monitor tumor growth rates after treatment. Nevertheless, these data suggest that STING-NPs may exhibit similar antitumor efficacy in human melanoma.
Methods
When they reached ~100 mm, tumors were injected intratumorally with 100 μL of PBS containing 10 μg of cGAMP in the appropriate formulation. Mice bearing subcutaneous tumors were intratumorally injected with PBS, cGAMP, or STING-NP, corresponding to a 10 μg dose of cGAMP. When tumors reached a size of ~100 mm3, STING-NPs were injected into the tail vein.

STING-NPs Improve the Delivery of cGAMP to STING, Activating
STING-NPs enhance cGAMP delivery to STING, activating myeloid and T cells for a tumoricidal effect. As a single agent, locally administered STING-NPs can generate robust and complete responses, inducing systemic antitumor immunity that can protect against tumor challenge. Importantly, STING NPs can be administered via both IT and IV routes, either as monotherapy or in combination with ICB for therapeutic benefit, potentially opening new clinical opportunities for exploiting STING agonists.
Suggestions for Future Work
Taken together, these data may serve as a basis for potential clinical investigation of STING-NPs. Another potential area of investigation involves exploiting the ability of STING-NPs to encapsulate a variety of cargo. Finally, the efficacy of STING-NPs can be improved through simple co-administration of receptor-blocking antibodies.
Sequential investigation of the effect of cellular depletion on the antitumor efficacy of STING-NPs will reveal the critical cellular players in tumor rejection. The susceptibility of STING-NPs to RES scavenging suggests that translational potential can be significantly enhanced by modification of the nanoparticle corona.
Nanoparticle Size Distribution Used for Theoretical Encapsulation Efficiency
Synthetic Scheme for PDSMA
Synthetic Scheme for PEG-DB and PEG-DBP Block Copolymers
Synthetic Scheme for PEG-CTA synthesis
Synthetic Scheme for 2’3’-cGAMP
LCMS Characterization of 2’3’-cGAMP
Comparison of Biological Activity of Synthesized and Commercially Available
Nanostring analysis of upregulated genes following IT STING-NP or
Synthetic scheme for fluorescent polymer labeling
Gating scheme for flow cytometric analysis of immune cell populations in the TME