BEHAVIORAL ECOLOGY OF NEOCHLAMISUS LEAF BEETLES:
THE ROLE OF FECAL CASES AND ASSOCIATED TRAITS By
Christopher G. Brown
Dissertation
Submitted to the Faculty of the Graduate School of Vanderbilt University
in partial fulfillment of the requirements for the degree of
DOCTOR OF PHILOSOPHY in
Biological Sciences May, 2009 Nashville, Tennessee
Approved:
Professor Daniel J. Funk Professor Dave McCauley
Professor Patrick Abbot Professor Manuel Leal Professor Jim Fordyce
This work is due to my parents, Glen and Sylvia Brown, whose loving care brought me to the top of the hill
and
to my beloved wife, Jennell Talley,
whose patience and (mostly) gentle encouragement pushed me to the other side of it.
ACKNOWLEDGEMENTS
They say it takes a village to raise a child and it probably takes that much or more to instill a doctorate. This work was supported by the financial help of a Student
Research Grant from the Animal Behaviour Society, an Exploration Grant from the Explorers Club, and a Dissertation Enhancement Grant from Vanderbilt University, as well as NSF IOB 0616135.
I appreciate all the support I received from my major advisor, Daniel J. Funk, who allowed me to work on his beetle babies and yet not on his main passion, their process of speciation. I am also grateful for the other members of my committee, Dave McCauley, Patrick Abbot, Jim Fordyce, and Manuel Leal who not only gave me wings to try new directions, but boundaries that kept me grounded in possibilities. Through them I have learned what it means to be a scientist.
Thanks to Roz Johnson, who always had an answer or knew where to find one.
I will forever remember the people that I met during this journey. Thanks to my cohort: Dr. Jennifer "Flowergirl" Ellis and Natasha Sherman, we did it together. Thanks to the hall: Dr. David Wills, Dr. and Mrs. Mark Chapman, Vikram Chakre, Jesse
Wenzler, Eric Janson, Dan Erickson, Officer Mark Mandel, Ryan Waples, and John
"Nutmeg" Gibbons. Thanks to those not on the hall, but who might have wished they were: Jenifer Ferguson, Robin Bairley, Elizabeth Thatcher, and Corey Snelson. And of course, I owe a great debt to the rambling gamblers: Scott P. Egan and Dan Duran. These people taught and retaught me how to laugh, to exercise, how to ignore with conviction, to speak up when it may or may not be necessary, to catch beetles with cheese wrappers,
to listen to birds, to listen to rap, to eat more healthily, the rules of roller derby, how to care passionately about subjects that no one else in the room has even heard of, about the theory of expanding Earth, to live life to the fullest, how to walk a 10-minute mile, and to appreciate every possibility, especially if there is evidence.
Thanks also goes to many friends and family, most of which I knew before I came to Vanderbilt, others I've met since then. I would like to thank my original family: Glen, Sylvia, and Matt Brown and Doris Peterson, my extended family: Mr. And Mrs. Talley, Houston, and Samantha. Thanks to friends from home: the Hutchisons and the Winters, and friends from abroad: the Vanhooses, Lovelesses, Campbells, RJ, and Derek. I also want to remember Gould and Ouida Brown and Clarence "CC" Peterson, who told me repeatedly that education was the most important gift I could ever give myself.
But above them all, I thank Jennell Talley, who stood beside me when I couldn’t stand up, who listened to me when I should have shut up, and who loves me no matter what. Without her here, none of this would have happened. Everything I am is for and because of her. She constantly reminds me what it means to be human and that simply being human is not good enough. I love you.
TABLE OF CONTENTS
Page
DEDICATION ... ii
ACKNOWLEDGEMENTS ... iii
LIST OF TABLES ... x
LIST OF FIGURES ... xi
Chapter I. INTRODUCTION ...1
There's no place like home ...1
It's a fecal matter ...7
A special case ...10
In the beginning ...16
Follow the directions...25
What's next ...29
References ...31
II. ASPECTS OF THE NATURAL HISTORY OF NEOCHLAMISUS (COLEOPTERA: CHRYSOMELIDAE): FECAL CASE-ASSOCIATED LIFE HISTORY AND BEHAVIOR, WITH A METHOD FOR STUDYING INSECT CONSTRUCTIONS ...41
Abstract ...41
Introduction ...42
Materials and Methods………...47
Study animals ...47
Oviposition and egg case construction ...48
Larval biology and a method for studying case construction ...48
Pupation and adult emergence ...51
Additional observations ...51
Results and Discussion ...53
Oviposition and egg case construction ...53
Larval biology ...58
Pupation and adult emergence ...67
Adult biology and phenology...71
Comparisons with other camptosomates ...71
Acknowledgments...78
References ...78
III. TESTING THE EFFECTS OF FECAL CASES ON SURVIVAL AND GROWTH RATE OF LARVAL NEOCHLAMISUS PLATANI IN FIELD CONDITIONS ...84
Abstract ...84
Introduction ...85
Methods...90
Test animals ...90
Host plants ...91
Predator exclusion treatments ...91
Experimental setup...92
Data collection and statistics ...92
Results and Discussion ...94
Effect of treatments on survival ...94
Rate of development in field conditions ...98
Larval wandering ...100
Potential factors contributing to mortality ...101
Possible competitors ...105
Conclusion ...106
Acknowledgements ...108
References ...108
IV. ANTIPREDATORY PROPERTIES OF A BIZARRE ANIMAL ARCHITECTURE: HOW COMPLEX FAECAL CASES THWART ARTHROPOD ATTACK ...112
Abstract ...112
Introduction ...113
Methods...118
Study Animals ...118
Experimental Treatments ...120
Experimental Design and Protocols ...121
Repeated observation trials ...121
Attic/no-attic repeated observation choice trials...123
Continuous observation trials ...123
Statistical analyses ...123
Results ...129
Repeated Observation Trials ...129
General findings ...129
N. bimaculatus ...130
N. platani...130
Attic/No-Attic Repeated Observation Choice Trials ...133
Continuous Observation Trials ...133
Discussion ...136
Faecal Cases ...137
External Trichomes ...138
Trichome Attic ...140
Defensive Behaviours Associated with Cases ...141
Parasitoids ...141
Faecal Case Evolution...143
Acknowledgements ...146
References ...146
V. UNORTHODOX TASTES: ACTIVE APPROPRIATION OF AN ANTI-HERBIVORE DEFENSE BY TRICHOME-SEEKING NEOCHLAMISUS LEAF BEETLES...153
Abstract ...153
Introduction ...154
Methods and Results ...159
Study animals ...159
Study plants ...159
Statistics ...160
Experimental leaf treatments ...160
No-choice feeding tests ...161
Repeated observation assays ...162
Field observations ...169
Behavioral observations ...170
Effects of trichomes on development and case strength ...174
Discussion ...177
General findings ...177
Possible benefits: indirect ...178
Possible benefits: direct ...181
Conclusion ...183
Acknowledgments...184
References ...184
VI. HOW MOIST SHOULD FECES BE? ECOLOGICAL TRADEOFFS IN A PHENOTYPICALLY PLASTIC ANIMAL ARCHITECTURE ...190
Abstract ...190
Introduction ...191
Methods...195
Study organisms and test conditions ...195
Statistical analyses ...196
Effects of case on surviving submersion ...196
Effects of case on pupal mass change as a function of RH ...197
Effects of case on survival and adult emergence as a function of RH ...198
Effects of RH on case strength...198
Effects of RH on predator handling time ...201
Results ...201
Effects of case on surviving submersion ...201
Effects of case on pupal mass changes as a function of RH ...202
Effects of case on survival and adult emergence as a function of RH ...202
Effects of RH on case strength...205
Effects of RH on predator handling time ...205
Discussion ...205
General findings ...207
Factors influencing case plasticity and tradeoffs ...210
Conclusion ...214
Acknowledgments...215
References ...216
VI. THE ROLE OF FECAL CASES, PLANT HAIRS, AND MICROHABITAT SELECTION IN THE BEHAVIORAL THERMOREGULATION OF NEOCHLAMISUS LEAF BEETLES ...220
Abstract ...220
Introduction ...221
Methods...225
Study organisms ...225
Effect of pupal cases, trichomes, and leaf position in the field ...225
Effect of pupal cases on time to reach equilibrium temperature ...228
Microhabitat selection of N. platani larvae ...228
Statistical analysis ...229
Results ...231
Effect of pupal cases, trichomes, and leaf position in the field ...231
Effect of pupal cases on time to reach equilibrium temperature ...234
Microhabitat selection of N. platani larvae ...234
Discussion ...240
General findings ...240
Fecal case evolution and temperature control ...242
Potential trade-offs of casebearing in relation to temperature ...244
Conclusion ...246
Acknowledgments...247
References ...248
VI. DEATH BY MOTH: INCORPORATION OF NEOCHLAMISUS LEAF BEETLE FECAL CASES INTO SILK BAGS OF THE BAGWORM THYRIDOPTERYX
EPHEMERAEFORMIS. A BUILDING BUILT WITH BUILDINGS ...253
Abstract ...253
Introduction ...254
Materials and Methods ...256
Results and Discussion ...258
General findings ...258
Implications of case incoporation ...261
Some thoughts on bag construction ...265
Conclusion ...266
Acknowledgments...266
References ...267
IX. SUMMARY AND RELATED PROJECTS ... 270
General Findings ...270
Related Projects ...276
Faecal case architecture in the gibbosus species group of Neochlamisus Karren, 1972 (Coleoptera: Chrysomelidae: Cryptocephalinae: Chlamisini) ...277
Building gnomon ...279
Unanswered questions ...282
Conclusion ...285
References ...286
LIST OF TABLES
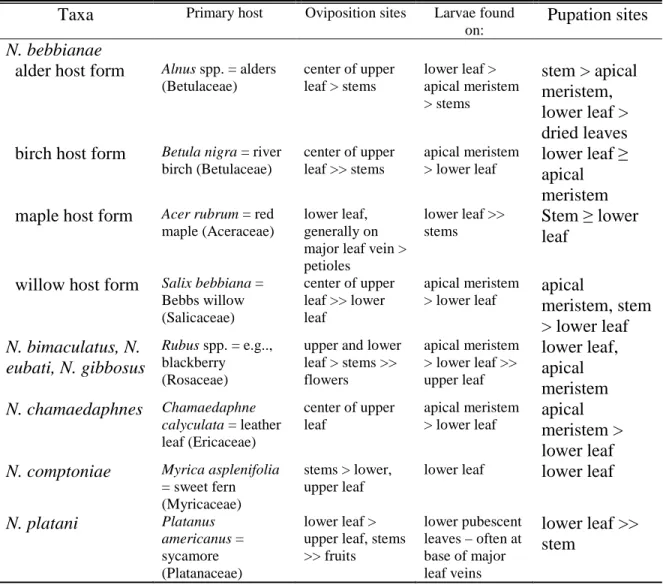
Table Page 2-1. Host plants and relative frequency of plant part usage by highly studied
Neochlamisus taxa ...46
3-1. Survival time...97
4-1. Percentages of immatures that experienced varying levels of threat (investigation, attack, death) as a function of beetle species, case-associated trait, predator, and life history stage ...124
4-2. Mean threat level experienced by immatures as a function of beetle species, case- associated trait, predator, and life history stage ...125
4-3. Timing between stages of predation threat as a function of beetle species, case- associated trait, predator, and life history stage ...126
4-4. Number of analyses (from Tables 1-3) in which a positive versus negative effect on resistance to predation was observed for each case-associated trait as function of predator, beetle species, and life history stage ...132
4-5. Summary of behaviours performed in the continuous observation assay ...135
7-1. Results of thermocouples inserted into various treatments in the field ...232
8-1 Details of bagworm bags from sites that contained Neochlamisus cases. ...257
LIST OF FIGURES
Figures Page
1-1. History of fecal retention in the subfamilies of Chrysomelidae. ...12
1-2. Relationships among camptosomate groups...22
2-1. Photographs and diagrammatic illustrations depicting external and internal aspects of Neochlamisus cases across life history stages ...44
2-2. Method for studying case construction ...50
2-3. Oviposition and egg case construction ...52
2-4. Aspects of the case-associated biology of Neochlamisus ...55
2-5. Neochlamisus larval development ...59
2-6. Temporal patterns of case construction ...65
2-7. The ‘trichome attic’ ...68
2-8. Temporal aspects of Neochlamisus life history ...72
3-1. Experimental treatments and field setup ...86
3-2. Diagrammatic representation of the field setup as shown perpendicular to the line of pools and treeline ...89
3-3. Experimental design ...93
3-4. Survival time...96
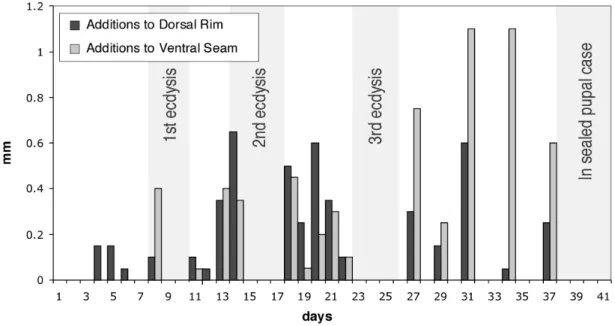
3.5. Number of observations per day in which fecal cases were observed to be "fixed" to the substrate as if the larva was undergoing ecdysis ...99
4-1. Typical appearance of N. platani larvae ...114
4-2. Basic structure of the experimental design for the repeated observation trials ...122
4-3. Ethograms of N. platani larval behaviour and cricket predatory activity from the 1-hr continuous observation trials ...134
5-1. Photograhic depictions of N. platani larvae, trichome structure, and leaf
treatments ...157
5-2. Results of feeding and location preference tests ...163
5-3. Time course of expression of preference for pubescent cuttings in choice tests 1 and 2 ...165
5-4. Time course of expression of preference for entire pubescent leaves despite confounding of treatment with leaf order on test branches in choice test 5 ...168
5-5. Distribution of the trichome density of leaves on which eggs and larvae were collected in the field ...171
5-6. Diagrammatic representation of larva clipping trichomes ...173
5-7. Results of N. platani larvae reared on pubescent and denuded leaves ...175
6-1. Photographs and cut-away diagrams of fecal cases containing a larva, a pupa, and an emerging adult ...194
6-2. Diagram of the sleigh-based crank and pulley system used to standardize the manner in which the penetrometer contacted the case and measured its strength ...200
6-3. Survival and floating in the submersion experiments ...203
6-4. Effects of RH treatment on pupal mass and survival ...204
6-5. Effects of RH treatment on pupal case strength ...206
6-6. An ecological tradeoff ...209
7-1. Diagrammatic representation of the six thermocouple treatments ...227
7-2 Effects of six treatments on temperature when taken every ten minutes in field conditions ...230
7-3. Effect of fecal cases on internal temperatures (average of ten-minute measurements every hour) ...233
7-4. Time taken to reach environmental temperature as measured by two thermocouple treatments ...235
7-5. Average temperature of fecal cases of live larvae through time ...236
7-6. Differences in larval temperature compared to the adjacent leaf surface and the opposite leaf surface ...238 7-7. The proportion of actions larvae undertook throughout the day according to
location within the plant...239 8-1. Thyridopteryx ephemeraeformis bags containing egg cases of Neochlamisus platani ...259 8-2. A Thyridopteryx ephemeraeformis bag containing a larval case and living larva of Neochlamisus platani ...260 8-3. Diagrammatic representation of how an N. platani egg case is attached by T.
ephemeraeformis to its bag using several strands of silk ...262 9-1. Relationships among investigated factors ...272 9-2. Proposed logarithmic spiral pattern of fecal cases ...281
CHAPTER I
INTRODUCTION
There's no place like home…
Many species of animals gather materials and attach and piece them together in particular ways to construct buildings. From protozoa to primates, animals across the kingdom are known to build cases, homes, traps, signals, and innumerable other kinds of animal architecture. Mike Hansell defines animal architecture as a behavior applied to materials to make a structure (Hansell 2005, 2007). The materials used are almost as diverse as the taxa that build with them and include mud, plant parts, silk, mucus, wax, paper, and even bubbles (von Frisch 1974, Hansell 1984, 2005). Some architectures are very intricate, e.g., weaverbirds delicately weave blades of grass together to form a cradle for their eggs (Collias and Collias 1984); others are amazingly durable, e.g., some potter wasp nests are thought to remain on the cliff face for thousands of years (Hansell 2007);
others are quite large, e.g., 30 m “mima prairies” in South Africa built by termites and mole rats (Lovegrove 1993). Beavers offer a well-known and illustrative example of the intricacy, efficiency, and art of animal architecture. Beavers build dams out of sticks and mud to stop the flow of streams, which in turn create artificial ponds (Zurowski 1992).
These ponds provide the beaver with ample food supply (bark) that is protected and easily accessible (underwater) in places where the surface freezes. Beavers also build an intricate lodge whose inner chamber is above the water's surface, but whose entrance is below, thus protecting the inside from terrestrial predators. The lodge is tightly
constructed on most sides and is therefore waterproof and maintains warmth, but has loose construction on top for airflow, a perfect home for raising a family.
A given structure often allows its builder to cope with external conditions without major alterations to physiology or requiring many new genetic adaptations (Dawson et al.
1988, Danks 2002, Hansell 2005). For example, termites are physiologically sensitive to changes in humidity, yet some species often forage long distances from their nests across inhabitable terrain. They maintain optimal conditions by building long tunnels of mud, feces, and carton (i.e., chewed plant material), which allow them to forage away from home without distinct morphological characteristics to live in different conditions (von Frisch 1974). Many caddisfly larvae can be keyed out based on case structure when the larvae themselves are relatively indistinguishable (Stuart and Currie 2001). This might also be why insects (which have highly adaptable cuticles) are so morphologically diverse, whereas there are relatively few body types in spiders (which have highly adaptable web arrangements).
Materials are either collected, secreted, or both (Hansell 2005). Collected materials include, but are by no means limited to, sticks, leaves, mud, stones, shells, the building materials of other organisms, and other organisms themselves. This latter group is interesting because sometimes the material also benefits from the construction, as in sea anemones on hermit crab shells that benefit from the crab's mobility by filtering diverse areas for food. Other times they probably do not, as in leaves used by weaver ants (although living leaves apparently do a good job of removing carbon dioxide and adding oxygen to the tightly woven nest chamber [Gould and Gould 2007]). Secreted materials used in animal architecture include silk (spiders), saliva (swallows), wax (bees), “glue”
(sticklebacks), and feces (see below). By secreting materials, animals can avoid any collecting costs incurred when gathering objects in the environment, but may suffer a cost of nutrients or energy when making the materials. Secreted materials, moreover, are consistent and uniform, although some processing is usually still needed, and allow the associated building behaviors to be simpler and more repetitive (Hansell 2005). Some materials are secreted for one purpose and collected for another. For example, down feathers, used to insulate adult ducks from weather and water, are also used to line nests (Hansell 2000). The palisade moth plucks out its own wing scales, used in flying and for coloration, to build a protective structure around its eggs (Gould and Gould 2007).
Although builders span most animal groups, the major taxa with constructing behaviors can be found in the arthropods and birds, and to a lesser extent the mammals (mostly rodents and humans). Despite their diversity and multiple evolutionary origins, Hansell (2005) divides animal-built structures into three main categories based on their function: food-gathering, communication, and shelter. I add locomotion to these
categories, since it is a much different purpose, even though it is much rarer.
Architectures are used to increase food availability in a number of ways, from traps to storage. Spiders and antlions are well-known trap makers, but another example would be caddisfly larvae that spin silk nets to filter the water current. One species of caddisfly builds a box with an intake tube and an outflow tube with a silk net spun across the middle (Hansell 2005). It sits tucked into a protected side chamber and collects food trapped in the net. In order to "sneak up on" prey, some insect predators cover themselves in debris from their habitats, such as leaf hairs or dead ants (Eisner et al. 1978, Brandt and Mahsberg 2002). These covers masquerade the predator as a common, less dangerous
component of the environment: a veritable "wolf-in-sheep's clothing" (Eisner et al. 1978), thus allowing them closer access to food sources. Once built, food stores and some leaf- rolls and mines make food provisions easier to access and can increase food quality.
Callosobruchus maculatus, a beetle that develops entirely in a single bean, builds walls within its chamber to reduce intraspecific competition so that multiple individuals can successfully emerge from one bean (Mano and Toquenaga 2008). And of course, many animals (mostly birds and primates) create tools, also considered animal architecture, which are used in food gathering and preparation (Hansell 2007).
Animal constructions are also used for communication, both between and within species. Male bowerbirds build intricate nest-like structures that are not used for rearing offspring, but rather serve as sexual signals by which females judge male quality
(Diamond 1986). Male mole crickets of Gryllotalpa australis dig a chamber with two cones curving to the surface that amplify their mating songs (Daws et al. 1996). Some species of orb-weaver spiders add zig-zag style patterns to their webs (stabilimenta) that may serve as visual cues to lure prey or alert flying birds that might otherwise damage the web to their presence (Eberhard 1990).
Although fewer in number and not considered as a major category by Hansell (2005, 2007), some animal built structures are used for locomotion and dispersal.
Individual caterpillars, mites, and spiderlings release silk strands, which catch the wind, and soar from one place to another, a method of movement called "ballooning" (Bell et al. 2005). Some caterpillars and spiders also release "draglines." These silken ropes slow their descent when falling and allow them to find their original location more easily.
More unusually, the violet snail Janthina floats on the water's surface by means of a raft of bubbles made by the snail's mucus (Hansell 1984).
By far the most common use of animal architecture is for shelter. Shelters provide a barrier between the builder and the immediate environment around it. Such homes include nests, burrows, mines, lodges, mounds, and condominiums. They serve to reduce predation by reducing detection (primary defense) and successful attack (secondary defense), and to reduce any harmful effects of abiotic conditions, such as temperature and humidity fluctuations (Hansell 1984, 2005, Danks 2002). Usually, shelters are built by parents for the protection of their young, and are not used by the adults after the breeding season. For instance, many birds and wasps build large shelters for eggs, but not winter shelters for themselves (Gould and Gould 2007). Sometimes though, especially in the holometabolous insects, juveniles build their own home. These range from relatively simple tunnels to intricate leaf rolls and shelters built by some caterpillars. Also common are larvae that build shelters around their bodies by sowing sticks, leaves, sand grains, and other objects together with silk to form a kind of tube or sack. These include the cases of caddisfly larvae (Trichoptera) and bags of the Psychidae (Lepidoptera). Not only do these structures provide protection from various environmental threats, they also allow the added bonus of motility, an unusual feature when considering most animal
architectures.
Numerous studies have elucidated the adaptive benefit of animal-built constructions, but building behaviors can also incur costs, including energy loss and increased conspicuousness. Gathering or secreting, processing, and building a structure takes energy and nutrients that might otherwise be used for reaching a larger size or
greater fecundity (Stevens et al. 1999, Otto 2000, 2000, Venner et al. 2003, McKie 2004).
Some builders even construct a new nest every year. Alternatively, building may allow builders to spend less energy than other organisms, for example, by using traps to aid hunting. They may also be able to save more energy in shelters while resting than animals that must maintain stricter vigilance or regulate their own homeostasis. Another cost, however, is that builders are often more conspicuous than non-builders. Much
architecture is large and immobile and thus can be easily found by predators and parasitoids, especially while in the act of building (Murakami 1999, Müller and Hilker 1999, Gould and Gould 2007). Since shelters often allow many conspecific individuals to live in a small confined place, such as bee hives and mole rat burrows, these species can also suffer higher rates of fungal, bacterial, and viral infection and more parasites than if they were living individually (e.g., Kopachena et al. 2007). Many birds (Hansell 2000) and mammals including badgers (Butler and Roper 1996), rats (Roper et al. 2002), and bats (Reckardt and Kerth 2007) avoid using the same roosting area for too long a time in order to avoid build-up of ectoparasites. This might be why some builders sacrifice the energy to build a new shelter every year. Despite these trade-offs, building behaviors have evolved numerous times across the animal kingdom.
In fact, animal architecture may even promote speciation in some instances (Gould and Gould 2007). Niche availability might expand with a change in building materials. Hansell (2007) tells a nice story of how architecture may increase diversity using a hypothetical wasp species that normally builds in clay. One population might begin adding plant materials, which allows building nests that are of lighter weight. Less heavy nests can be hung in trees rather than the previously required cliff face, thus
increasing nesting opportunities and reducing competition and predation. Concordantly, new designs may evolve incrementally over time as populations are geographically separated. Building behaviors can be genetically simple (Dawson et al. 1988, Bult and Lynch 1997) and organisms can adapt to a different environments through changes in building behaviors without drastic changes in morphology. When genetic blueprints for separate building designs cannot be mixed effectively, there may be reproductive isolation via lower hybrid fitness, since hybrids do not build a functional structure (e.g., Dawson et al. 1988). This could eventually produce separate species. Populations of a building species can become specialized to use certain materials, such as leaves of a certain plant species when making a leaf-roll, and become less efficient on materials used by related populations. Social insects, in which multiple individuals contribute to one design, have to work together using the same "blueprint." Therefore, combining different genotypes reduces their ability to build an effective structure (Gould and Gould 2007, Hansell 2007). Reproductive isolation is especially likely when the structure is used as a mating signal, as in bowerbirds, or as a mating site, as in swallows, since females from different groups may choose males based on their building behaviors.
Architectures can change the environment, and thus selective pressures, for non- building organisms too, a type of niche construction (Hansell 1993). Beavers change the landscape dramatically (e.g., Rosell et al. 2005), but other such "ecosystem engineers"
can be a little subtler. Old organ pipe wasp nests are used by cliff swallows to attach their nests (Hansell 2007). Cultivated fungi that are reared in an insect's structure can evolve in conjunction with their host insect (Farrell et al. 2001). Large structures often become home to a variety of other plants and animals. Several birds "squat" in termite nests and
social spider webs (Brightsmith 2000, 2004, Kesler and Haig 2005) and may require unknown specializations to do so. Leaf shelters are known to induce dramatic changes in diversity within and around a host plant by attracting, and increasing niches for, other species (Ohgushi 2005). Therefore animal-built structures can greatly impact the morphology, behaviors, and ecology of the builder and other non-related organisms.
Architectures too, uniquely supply quantifiable physical evidence of behaviors that persists across species bifurcations and geological time, thus allowing us to map many subtle behavioral traits in phylogenetic studies that would otherwise be lost to time (e.g., Winkler and Sheldon 1993).
It's a fecal matter…
One of the most intriguing building media, at least by human standards, is feces.
Yet many organisms (mostly insects) have evolved amazing ways to deal effectively with their wastes (Weiss 2006). Feces in insects are used as an aggregation attractant (Suzuki 1985, Weaver et al. 1989), a way to find mates (Fettköther et al. 2000), or the way back home (Miller and Koehler 2000). Feces may also reduce competition and often act as an oviposition deterrent (Anderson et al. 1993, Anbutsu and Togashi 2002, Borg-Karlson et al. 2006, Addesso 2007). Leaf-cutter ants and fungivorous termites use it as fertilizer in their fungus gardens (Hansell 2005). Waste products are also the main link in aphid-ant mutualisms (Stadler and Dixon 2005). Fecal material may contain toxins that reduce predation. For instance, Eloria noyesi moths excrete an unusual feeding-deterrent known as cocaine when feeding on Erythroxylum coca (Blum et al. 1981). And other insects use
waste products as a building material, as seen in thrips, termites, spittlebugs, lepidopterans, and beetles (Weiss 2006).
Most animals simply discard their waste products. It is often considered unhealthy to remain around fecal material since it can be infected with fungi and bacteria and may aid the transmission of parasites whose cycles include internal and external stages (Collison et al. 2004). However, Weiss (2003) found that keeping frass around did not affect survival and says that no studies have found a negative effect of living with frass, at least in insects (Weiss 2006). In fact, some species are known to incorporate
antimicrobial properties in their excrement (Müller and Hilker 2004). Leaf-cutter ants are known to add specific antifungal properties in the "manure" used in their fungal gardens to prevent unwanted fungus from growing, but without affecting the desired "crop"
species (Poulsen et al. 2005). Some shelter-dwelling insects whose frass accumulates in their shelters (e.g., tent caterpillars and some leaf-rollers) may emit antifungal chemicals to prevent pathogen growth (Weiss 2006). Perhaps insects are physiologically less likely to suffer from the negative issues of feces and this may explain why building with waste products is more prevalent in arthropods than vertebrates. This tendency may be even more likely in phytophagous insects. Excrement from a leaf-feeder may simply be less likely to become infectious than a meat-eater's, because of the chemical nature of their food source or differences in internal parasite quantity or life cycles. Phytophagous insects often sequester toxins and antimicrobial chemicals from their host plants and naturally such substances can easily be incorporated into feces. Insects, however, do face the danger of attack by predators and parasitoids, which can use feces as a kairomone to locate their hosts (Müller and Hilker 1999, Schaffner and Müller 2001, Weiss 2003).
As a building material, feces share the beneficial attributes of other secreted materials, mainly reduced collecting costs, uniform building blocks, and malleability.
However, feces have one major advantage in that they are produced by almost all organisms as waste material. Unless the builder adds chemicals or other objects to the feces, there is no additional cost to the organism's metabolism (e.g., Olmstead and Denno 1992). Fecal material can be easily made of materials of different strengths and densities, such as digested and undigested plant material (Chaboo et al. 2008, Chapter V). Such composite materials (like reinforced concrete) are often better at resisting breaking under tension and compression (Hansell 2000). Feces may also exhibit the same thixotropic properties as mud, a material used by many builders. Using feces as a building medium is also beneficial since it instantaneously camouflages. It is a very earthy, natural color and blends in well with most habitats. The puriri moth caterpillar Aenetus virescens covers its tunnel entrance with silk and feces, making it barely distinguishable from the surrounding tree trunk (Buffalo Museum of Science website). Of course, fecal coverings also let a builder masquerade as feces. Even parasitoids that use feces to locate their host, may not expect to find their host under the feces itself.
The production of feces as a building material is dual-purpose. One is digestive, i.e., to acquire as many nutrients as possible and remove wastes effectively. The other is for building, i.e., to make a consistent building material that will hold together and serve whatever functions are intended (e.g., maintain humidity, prevent predation, etc). These requirements could select for specific types of feeding or differences in digestion and may induce tradeoffs. That is, the most nutritive food may not be the best for making buildings and thus a healthy individual may sacrifice structural integrity. Alternatively,
producing feces that can best be used as a building material may enable less nutrient absorption, and thus more time feeding.
A special case…
As in several categories, the most prevalent users of feces might be the beetles.
Tenebrionids use it as an aggregation attractant (Suzuki 1985, Weaver et al. 1989), weevils (Addesso et al. 2007) and long-horned beetles (Anbutsu and Togashi 2002) use it as an oviposition deterrent, and, as stated above, a seed beetle uses feces to build a wall to prevent competition within a bean (Mano and Toquenaga 2008). Feces in most beetle architectures are used for protection (Gross 1993, Weiss 2006). Cerambycid larval tunnels often get packed with feces and sometimes the larva is surrounded by it when pupating (Rodd 1951). Many create a thick "frass plug" that seals off the pupal chamber, which could prevent other organisms from accessing them (although the parasitoid Megalyra fasciipennis locates these plugs and oviposits through them [Rodd 1951]). A fecal covering is similarly used to protect ladybird beetle eggs (Arakaki 1988).
Feces used as shelter may be most common in the leaf beetles (Chrysomelidae) (Olmstead 1994, Müller and Hilker 2004). An estimated twenty percent of leaf beetle species (~7,000) cover their eggs or larvae in waste products (Vencl and Morton 1999, Vencl et al. 1999). These fecal coverings take a variety of shapes and sizes and serve to protect young from predators (Gross 1993, Weiss 2006) or desiccation (as suggested by Karren 1964, Damman and Cappucino 1991, Olmstead 1994, Nahrung and Marohasy 1997). Species of both Donaciinae and Criocerinae subfamilies are known to deposit substances, such as “glue” or "slime", on egg batches, which may attach the eggs to the
substrate, protect them from parasitoids, or contain beneficial symbionts (Schmitt 1988, Müller and Hilker 2004). Almost every other subfamily within Chrysomelidae contains species that are known to cover their eggs in feces. This trait may have evolved only once shortly after their split from their closest common ancestor and been lost in most species, or multiple times across taxa (Fig. 1). The prevalence of fecal use in so many leaf beetle subfamilies (~9/11) may indicate that this trait, or at least the propensity to evolve this trait, is ancestral in this family. Fecal retention may have evolved multiple times in association with the evolution from an ancestor that used concealed habitats to a derived state of using more open ones, such as leaf surfaces, (at least in larvae) (Vencl and
Morton 1999, Chaboo 2007, JB Karren personal communication). Indeed, lineages of leaf beetles often described as basal have subterranean, aquatic, mining, or galling larvae and do not use feces in an architectural sense.
Toxins in the feces of some galeuricines protect nearby larvae, although the larvae do not carry them (Müller and Hilker 2004). Some criocerinae carry loose, wet feces on
their dorsum as larvae and possess a dorsal anus to aid this behavior. Species in two genera of Alticinae also cover their eggs and/or larvae in wet fecal material (Evans et al.
2000, Jolivet and Verma 2002), but it is unknown if they share the same morphological feature. Loose, wet fecal coverings in alticines are known to protect larvae from ant attack (Di Giusto et al. 2001), but the defensive mechanism may be reliant on chemicals
derived from the host plant rather than the feces itself (Morton and Vencl 1998, Vencl and Morton 1998). One chrysomeline larva builds a tubular shelter out of feces that is
attached to the host plant (Jolivet and Verma 2002). The larva typically remains
Figure 1-1. History of fecal retention in the subfamilies of Chrysomelidae. Relationships of the subfamilies are redrawn from phylogenies in the publications listed above each tree: A) From Farrell and Sequiera 2004, B) from Duckett et al. 2004, C) from Gomez-Zurita et al. 2007, D) from Hunt et al. 2007. Use of fecal material from Isono 1988, Schmitt 1988, Olmstead 1994, Vencl and Morton 1999, Jolivet and Verma 2002, and Müller and Hilker 2004. Although the relationships among the groups vary, each indicates that covering the eggs in waste material is prevalent throughout the leaf beetles and may have evolved once, possibly basally to the family.
Fecal coverings in larvae, however, may have evolved four separate times: twice as a wet mass over the dorsum (Criocerinae and Alticinae), once as a fecal shield (Cassidinae), and once as a fecal case (Camptosomata: Lamprosomatinae and Crpytocephalinae). ? next to Eumolpinae egg coverings indicate that I do not know where the species that possess the trait appear on the phylogeny.
facedown inside the tube. This is a remarkable self-supporting structure and apparently unique to this group.
The most diverse fecal structures of the chrysomelids are the shields and thatches of the subfamily Cassidinae sensu lato (Fig. 1). Larvae in this group often retain both shed exuviae and feces on projections from the abdomen known as urogomphi or furca.
These "shields" often take the form of unstructured masses (Chaboo 2007), but can be woven into basket-like shapes that nearly cover the entire larval body (Eisner and Eisner 2000). Feces are applied to the growing shield with a telescopic and quite maneuverable anus. The fecal shields on the tip of the abdomen are often shifted and oriented in the direction of disturbance, which is detected by tactile stimulation of lateral setae and projections (Eisner et al. 1967, McBride et al. 2000) or visually (Chaboo 2002). Shields of a variety of species have been tested for a protective function and are known to be used as a physical barrier, both by blocking attack and thrashing attackers (Eisner et al.
1967, Olmstead and Denno 1993, Müller 2002), and as a chemical deterrent (Vencl et al.
1999, Vencl et al. 2005). Cassidines can rebuild their shields if removed, and repair damaged shields. Fecal shields are probably a synapomorphy of this group and Chaboo (2007) claimed they evolved two times from a basal leaf-mining hispinoid-like ancestor (Fig. 1).
The most architecturally complex fecal structure in leaf beetles, however, would have to be the fecal cases of the group Camptosomata, consisting of the subfamilies Lamprosomatinae and Cryptocephalinae (formerly Chlamisinae, Clytrinae, and
Cryptocephalinae) sensu lato. Fecal cases are bell-shaped and completely surround eggs and almost completely surround larvae (there is an opening at one end through which the
head and legs are extended for walking and feeding). They take a variety of shapes and may include mineral and plant-derived components (Erber 1988, Chaboo et al. 2008).
Unlike fecal shields, these structures are not maneuverable and are not rebuilt if removed (Donisthorpe 1902, Briggs 1905, Müller and Hilker 2004, Brown and Funk 2005,
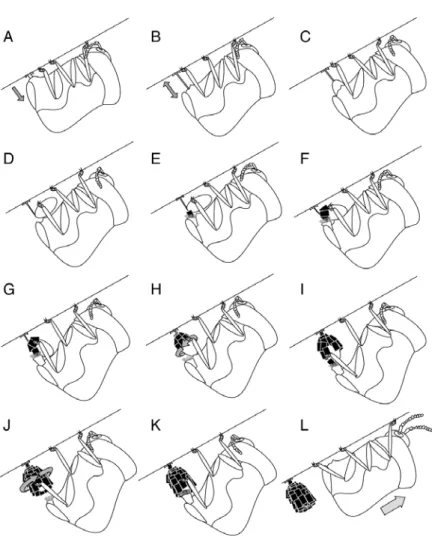
Chapter II), but they can be repaired. Fecal cases begin as an egg covering that ovipositing females build out of many individual fecal plates around each egg.
Immediately after producing an egg, the female grasps it in her hind legs and attaches fecal plates with abdominal appendages, building walls around it and eventually creating a complete case (Fig. 1). The larva later breaks open the egg case roof, but it does not discard the egg case. Instead it turns it over and carries it on its back throughout the juvenile stages, adding to it in rows rather than plates via an anus that is positioned near the opening (Erber 1988, Brown and Funk 2005, Chapter II). Eventually the case is rounded out and the opening is sealed with more fecal material. Pupation then takes place inside the sealed pupal case. Adults emerge by cutting a circular line around the apex of the case and removing a "pupal cap" to escape.
With the exception of burrowers, animal architects often lack building-specific morphologies (Hansell 2005, 2007). Interestingly, like some other chrysomelids (in which species possess a dorsal anus or urogomphi), the Camptosomata possess several morphological synapomorphies associated with building and maintaining their fecal structures (Chaboo et al. 2008). (I am not sure what makes the fecal-retaining lifestyle special in this way, but it may relate to the intimacy between the builder, secretion of materials, and the building itself.) Adult females in the Camptosomata possess a complex structure of scleritized plates and muscles inside the abdomen with which the fecal plates
are pressed and extruded, called the kotpresse, and beautifully illustrated by Erber (1968).
Crytocephalines, but not lamprosomatines, also have a dip or fovea on the ventral side of one posterior segment of the abdomen that is surrounded by brush-like setae, into which the egg is placed while the egg case is built. Hairs on another extendible segment may serve to indicate to the female when the egg case wall has been extended far enough and construction of the egg case roof (or sealing of the egg case) can begin (Brown and Funk 2005, Chapter II). Larvae are U-shaped and bent such that the anus is near the mouth, granting larvae easy access to their building materials. Larval camptosomates also have longer legs, which must be extended below and beyond the case when walking, and fewer setae, spiracles, and less scleritization than related beetles (Moldenke 1970, LeSage 1982, Root and Messina 1983), perhaps relying more on the case for protection from predators or to regulate homeostasis than their own physiology. Although they have fewer setae, they do possess many "spiny or rounded protuberances" that could aid attachment of the larvae to their cases (Root and Messina 1983). It seems plausible that adults and larvae have a specialized internal organs that create and store the right mixture of feces, plant material, and chemicals for fecal construction. Female Neochlamisus have
proportionately longer abdomens than males (Adams and Funk 1997), which in many insects reflects selection for increased fecundity, but in camptosomates may also be involved in fecal plate formation and storage of materials. Larvae may possess special glands that produce some kind of adhesive secretion to mix in with their feces that help attach new rows to old rows (Erber 1988). Contrastingly, fecal coverings also offer a place where larvae of some leaf beetles are known to store toxic chemicals, without developing special glands for this purpose or requiring a wounding event for their
production (Hilker 1992). In fact, adult camptosomates have a reduced set of defensive glands compared to some leaf beetles (Deroe and Pasteels 1982). If this is also true of their larvae, it may be evidence that the casebearers are indeed relying on their cases for storage of chemicals and protection from predators. The chemical make-up of any of the camptosomate fecal cases is, however, unknown.
In the beginning…
Exploring the many examples of extant insects that use feces could shed light on how other insects may have evolved to use feces as building media. Egg coverings and temporary larval coverings exist in several insect taxa, including grasshoppers (Gangwere 1993), hymenoptera (Seymour 1974), lepidoptera (Bernays and Woods 2000), and many different families of beetles (Weiss 2006), indicating that this behavior has evolved multiple times. Manduca sexta larvae are known to excrete feces and cover their backs with waste when parasitoids are nearby, indicating a protective function (Bernays and Woods 2000). Seymour (1974) found that sawfly larvae cover their venters with wet fecal material when the temperature is raised above a certain threshold and other measures fail to lower internal temperature.
More specifically, carrying a fecal case is an ancient trait whose evolution has not been well explored. Leaf beetle larvae possessing fully developed fecal cases are known from Dominican amber (Poinar 1999, Grimaldi and Engel 2005), a rare example of behavior in fossils, and may be as old as 45 million years (Chaboo, Engel, and Chamorro- Lacayo in prep). Despite similarities to other leaf beetle fecal structures, especially the fecal thatches of Hemisphaerota, which completely cover the larva's body, fecal cases are
probably not derived from them (Fig. 1). Instead, the wet coverings in Criocerinae and Alticinae, the fecal shields of the Cassidinae, and the fecal cases of the Camptosomata probably each evolved independently from the other, although they could have all been derived from an egg-covering ancestor. The morphological adaptations within each group do not suggest similar evolutionary trajectories. For instance, it is unlikely that a given lineage that developed a dorsal anus to aid covering itself in loose material would later evolve a curved abdomen with the anus next to the mouth to build a hardened case. By most accounts, the Camptosomata are monophyletic (Fig. 1), and it is likely that fecal cases evolved only once.
It seems that no species in the Camptosomata have lost the case-carrying trait, whereas some cassidine species no longer carry a shield (Chaboo 2007). (Although Erber [1988] states that species of Labidostomis and Temnodachrys may cover eggs only in
"anal secretion" with no fecal component, this seems inaccurate [Wasowska 2007], especially since larvae of Labidostomis humeralis still carry a normal-looking fecal case [Erber 1988].)
More intriguingly, how did a larval case evolve out of a fecal egg covering in the Camptosomata, since other species possess larval coverings with no egg covering or egg coverings that are not incorporated into larval coverings? Often, architectural behaviors have no extant intermediates, such as in palisades moths and whorl moth caterpillars, in which only one species of the genus expresses the building behavior. Fortunately, a potential evolutionary pathway for fecal cases is apparent in other leaf beetles. One cassidine species, Chelymorpha cassidea, lays its eggs atop long stalks, as do many camptosomates. The larvae then feed below these stalks in groups until they are able to
build their shields (Olmstead 1994). Illustrating an even more likely evolutionary
pathway, larval Lilioceris subpolita apparently carry their eggshells for the first day after hatching. Only later do they discard the eggshell and begin covering their backs with feces (Schmitt 1988). Mothers of this species apparently do not coat the eggs with feces, but it is easy to see how similar behaviors in a group that had already evolved egg coverings could be taken up and extended by larvae to become the fecal cases of the Camptosomata.
Although not numerically diverse by leaf beetle standards, the Camptosomata (~4000 species) (Chaboo et al. 2008) can be found in ecologically diverse habitats (Erber 1988, Jolivet and Verma 2002) and there are interesting differences and similarities of fecal-related traits within and among the tribes of this group. Based on examples of cases in the literature, the fecal cases within the subfamily Cryptocephalinae are quite similar in appearance, with the main exception of a few species within the Chlamisini (although fecal addition by larvae may vary and some of the components of cases differ across tribes) (see Fig. 17 in Erber 1988). Unusually, Coleothorpa dominicana franscicana, a clytrine, builds a cone surrounding the plant stem on which it lays its eggs (Slosser 2003).
The cone opens toward the base of the branch and may protect the eggs from passersby, much like the plastic cones that my grandfather put around his birdfeeder to keep the squirrels out of the birdseed. I am not aware of any other camptosomate building structures separate from the case. See Erber (1988) and Chaboo et al. (2008) for more comparisons of case components throughout the group.
Within the Chlamisini, however, the shape and development of fecal cases can be dramatically different. For instance, Neochlamisus (studied in this dissertation) and
Exema produce fecal cases similar to other camptosomates: i.e., an egg case composed of many individual plates that becomes incorporated into a relatively simple smooth tube (i.e., the larval case) with fecal additions in two planes, represented by the dorsal rim and the ventral suture (Erber 1988, Brown and Funk 2005, Chapter II). Brown (1943) and Karren (1972) consider fecal cases to be too similar among North American chlamisines to offer anything of value as characters. However, species from two related South
American genera, Fulcidax and Chlamisus, construct cases much differently than what is known for Neochlamisus. For instance, neither genus seems to employ a ventral suture to add width to the case. Fulcidax monstrosa, the largest extant chlamisine, does not wrap its eggs in plates of fecal material, but rather covers them in one continuous strip, forming a spherically-shaped, rather than bell-shaped, case (Flinte and Macêdo 2004b).
The larvae add to this case in large bands that extend out from the previous material, forming several flanges that spiral in different directions. The maternal portion is never dissected, as in Neochlamisus, and remains as a distinct ball on the back of the larval case. The finished case is quite irregular in shape, rather than a smooth tube. The life cycle of Chlamisus minax is nicely illustrated by Reu and Del-Claro (2005). The eggs of this species are wrapped in plates, but the egg case still takes the form of a round ball.
This maternal construction is again not altered, but larvae add rows of increasing size in one direction from it, forming a long cone. Prior to pupation, a much wider section is added to the bottom of this larval cone, and the previous sections are cut off and discarded. I have never seen another example of parts of a larval case deliberately removed by larvae, although Donisthorpe (1902) says that the maternal portion of the larval case of one clytrine breaks off and the larva seals the remaining hole.
The adults of the Chlamisini share the same “warty” appearance unique to the tribe, and many authors consider the tribe monophyletic (Karren 1972, LeSage 1984, Reid 1990), perhaps indicating that changes in building behaviors may be freer to diversify than adult morphology. There is no phylogeny of the genera within this tribe, but it might be assumed that the style of case building in Fulcidax and Chlamisus is extremely derived from the basic pattern seen elsewhere. Considering only the structurally "simpler" method of egg wrapping seen in Fulcidax, this trait may seem basal, but considering the shared traits of Neochlamisus and Exema cases with the rest of the camptosomates, this is phylogenetically implausible. This means that Fulcidax evolutionarily lost the fecal plates made by ovipositing females. The larval cases of some species of the phylogenetically ambiguous camptosomate subfamily Lamprosomatinae appear tube-like, as in most Cryptocephalinae (Kasap and Crowson 1976), while others appear more like Fulcidax cases with larval additions added to two lateral flanges (Erber 1988).
The chlamisines, and probably the majority of the camptosomates, are mostly found in the neotropics and may have originated in South America, although they are now found in all zoogeographical areas (Karren 1970). Most camptosomate adults are herbivorous, as are most chrysomelids, but the majority of larval camptosomates are saprophagous or fungivorous, which is quite unusual (Riley 1874, Donisthorpe 1902, LeSage 1984b, Erber 1988, Jolivet and Verma 2002). Many of the Cryptocephalini live in the leaf litter and apparently cannot climb plants (LeSage 1984b, 1985, 1986), whereas most chlamisines are intimately tied to and dependent on their host species (Brown 1943, LeSage 1984a, Funk 1998, Flinte et al. 2003). Many species of clytrines are
myrmecophiles, meaning they live inside ant nests as larvae, and some are even willing and able to eat dead arthropods, possibly including their insect hosts (LeSage and Steifel 1996). Some authors have proposed that these egg cases are attractive to ants, which induced to bring them into the nest, either through chemical manipulation or by mimicking normal foods or material for nest construction (Donisthorpe 1902, Erber 1988). Results from choice tests are mixed (LeSage and Stiefel 1996, Stiefel and Margolies 1998), but fecal cases would most assuredly help the beetle survive the trip, thus allowing it access to a new niche. Adults apparently drop eggs near ant nest openings and some adult clytrines may even mimic the ants themselves (Selman 1962), perhaps to avoid being eaten by their hosts as they leave the nest (Donisthorpe 1902, Erber 1988). The ground-dwelling taxa have straight tarsal claws and often retreat inside their cases when disturbed. The case falls to one side, but the larva's flattened head blocks the entrance (Donisthorpe 1902, Lawson 1976, Erber 1988). In Chlamisini however, falling from the host plant could be fatal, so larvae have recurved claws that dig into leaf tissue and they hold their cases down to the surface when provoked, covering the entire body, legs included. (Karren 1964, Wallace 1970, LeSage 1984a, Neal 1989, Brown and Funk 2005, Chapter II).
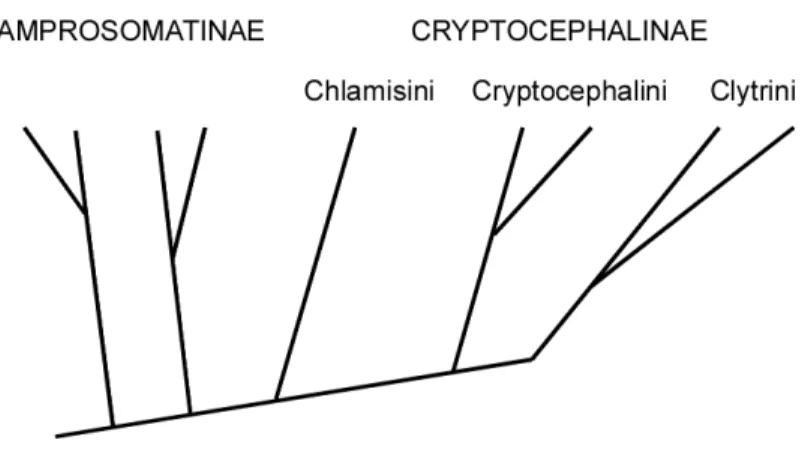
Kasap and Crowson (1976) created a phylogenetic dendrogram of the Camptosomata using 19 morphological and ecological characters, although LeSage (1985) found fault in one of their trait delineations. They conclude that the subfamilies Lamprosomatinae and Sphaerocharinae (now included within Lamprosomatinae) are basal to the group, with Chlamisini branching off next, and Cryptocephalini and Clytrini splitting last (Fig. 2). This tree might indicate that wrapping eggs in individual plates and
Figure 1-2. Relationships among camptosomate groups. Cladogram redrawn from phylogenetic dendrogram based on 18 morphological characters and 1 ecological
character (association with ants) of larvae and adults of the Camptosomate group in Kasap and Crowson 1976.
the tube-like larval case structure are derived; unfortunately no case-associated
morphological or behavioral traits were included in their diagram. If true, it would also suggest that living in ant nests is derived, perhaps meaning that this lifestyle is facilitated by the fecal case, rather than vice versa. Of course a phylogeny of the Camptosomata using molecular data from many species within each tribe, especially those whose cases are described in detail, would be most helpful.
Knowledge of the evolution of other building components also begs for more information. Firstly, new material not only needs to stick to itself as it is added, but each row of newly added fecal material must stick to the previously applied one. This is true for adult egg case building and larval case building. The attachment point between the rows that larvae add is a particularly weak one and a source of anisotropic failure (personal observations, Hansell 2005), but the horizontally and vertically overlapping addition of plates by females to the egg case does not appear to suffer the same affect.
What building traits have evolved to accommodate these factors? Secondly, larvae of the casebearer Neochlamisus emit some kind of liquid from their mouths (Brown and Funk 2005, Chapter II) (perhaps a hypopharnyx secretion) and/or anus (Briggs 1905) when disturbed. This substance then hardens into a dark caramel-colored solid, similar to the color of finished cases. If camptosomate larvae incorporate a fixing agent into their feces before applying new layers (as suggested by Riley 1874, Donisthorpe 1902, Briggs 1905, Erber 1988, Brown and Funk 2005), this substance may be derived from a similar
secretion. It would be interesting to know the make-up of this substance and to determine if it is incorporated into new layers of the case (representing a predaptation if it originated for defensive purposes), if it is incorporated into the maternally formed plates (indicating
an anatomically different origin since adults do not use their mouthparts to build egg cases), and if it is host-plant derived. Erber (1988) says both larvae and adults possess rectal glands with which they create an adhesive secretion used in case-building,
indicating potentially similar anatomy for this "glue", but a different one for the defensive spitting.
The egg stalk found in some casebearers (including species of Clytrini and
Lamprosomatini, Exema and Neochlamisus) is also intriguing. Similar strands holding up stenogastrine wasp nests, lacewing eggs, and many spider egg sacs are considered as defensive, preventing attack from ants (Hansell 2005), but Neochlamisus stalks become twisted as the mother adds rows of plates, sometimes lowering the egg case close to the leaf surface, perhaps exposing them to ants. Perhaps the egg stalk simply serves to keep larvae tied to their host plants, since leaving them would be fatal. It would be interesting to see if the presence of egg stalks in the Camptosomata is correlated with the evolution of herbivory or host plant specificity, assuming it has evolved multiple times.
Alternatively, it could be basal to the casebearers and lost in nonherbivorous groups.
Although not necessarily affecting fitness, egg stalks may explain different egg case structures among camptosomates. For example, perhaps the egg stalk provides stability, allowing Neochlamisus to build a flat egg case roof, while other species close the egg case in a more gradual way (see Lawson 1976, Erber 1988).
There is also no fungal growth on Neochlamisus fecal cases found in the wild, whereas those in the lab can become covered in tiny white mycelia (personal
observation). Although antifungal substances may be inherent in chrysomelids or derived from the host plant, camptosomates may have evolved the production of special
chemicals or even symbiotic associations for this purpose. They may have also needed to develop different mechanisms to incorporate previously evolved antimicrobial chemicals into their fecal material, such as glands in the digestive tract.
Follow the directions…
Two recent books on animal architecture, Animal Architecture: Building and the Evolution of Intelligence (Gould and Gould 2007) and Built by Animals (Hansell 2007), speculate on the cognitive capacity required to build some of the intricate structures found in nature. Although not a focus of my studies, a comparison with Camptosomates might prove fruitful. Gould and Gould state, "when it comes to building, solitary insects provide textbook examples of innate control." (p. 74). In other words, most examples from insects show that building behaviors are hardwired. This is true even in insects that are known to possess learning and processing abilities during other activities. For
example, hunting wasps follow strict rules when constructing burrows and nests (Gould and Gould 2007), but finding their nests and provisioning their young requires memory (Tinbergen 1951). Hansell (2005, 2007) says that building behaviors are selected to be few in number, stereotyped and repetitive. Indeed, weaver ants build elaborate nests in a very variable environment (i.e., the tree canopy), but may only express 29 individual behaviors to do so (Wilson 1980). Caddisfly larvae can build intricate and precisely round cylinders by using one simple rule: Add grains of sand of size similar to previous grains to the least extended section of the case, using body size (or head size) as a template for case diameter (Hansell 2005, 2007). Most of the building processes in Camptosomata are probably heavily regimented in a similar way. In fact, Briggs (1905)
replaced the fecal case of a fourth instar Neochlamisus larva with a hand-made case of paraffin wax. The larva continued to add to its case, including cutting and adding more material to the ventral suture. This indicates that any encapsulating case-like structure induces accurate building behaviors. They are probably cued to add material to any edge of material and are obviously not concerned with the content of the current case. Further, when the case is removed, larvae will sometimes begin coating the substrate in a flat layer of fecal material (personal observation). Obviously larvae are not concerned about the shape or location of this pre-built fecal edge. This may explain why larvae are incapable of building a case de novo. The rule "add material to material" requires a starting point.
For many builders, the starting point is the most difficult since there is no template and behaviors must be more flexible given variations in habitat (e.g., spider webs across bushes and weaver bird nests in trees) (Hansell 2005). For camptosomates, this may not be a problem. The egg serves as a starting point and template for females and the egg case is the starting point for larvae. Larvae then probably use their own bodies as a template for case size and roundness. Once, I disturbed an ovipositing female after she had begun building an egg case. The base and about half of the walls were completed. She walked away from her egg, turned around, and came back to it. When her head reached the egg, she ate the entire egg, leaving just the egg case. (Many insects will devour their own eggs if conditions are not appropriate for their development and the resources are better spent elsewhere. And an unwrapped Neochlamisus egg will not survive.) She then walked over the half-finished, empty egg case, grabbed it in her hind legs in the typical egg-wrapping position, and promptly closed off the egg case by
building a roof. She walked away from what looked like a normal, completely closed egg case, only it was half as tall as it should be and it was empty. This observation indicates that females are stimulated by the presence of the egg case to finish wrapping an egg, but they use the egg itself to determine when to finish the egg case. The egg is the template for the egg case. Since the case was empty, she could no longer feel the top of the egg, and so moved to the next step in the sequence: begin closing the case.
Building a fecal case probably requires different cues for different purposes.
Transitions between only adding length (as done in the beginning instars) to
predominantly adding width (in the last instar) (Chapter II) may be directly cued by genetic signals and/or to larval size, which is also dependent on feeding. Chapman (1998) states that some insects perform ecdysis based on gut size. In Neochlamisus, the cue for ecdysis would also need to cue attaching the case to the leaf, which takes place prior to ecdysis. Neal (1989) found that when females are held upside-down, they produce egg cases that are significantly wider and shorter than in the field. This result could be a difference between field and laboratory conditions since the proper controls were not done, or could be due to differences in gravitational cues. Although spiders seem to rely on gravity when building their webs, it is not apparently necessary for proper web building (Witt et al. 1977). In Neochlamisus' case, orientation may more likely affect a female's ability to hold onto the egg while wrapping it properly and this gravitational requirement might explain some of the interspecific variation in case size since some species lay more eggs on the top of the leaf, whereas others lay more eggs on the bottoms (e.g., N. platani, which lays most of its eggs on the abaxial side of leaves, produces a more elongate egg case than related species) (Brown and Funk 2005, Chaboo et al. 2008).
Therefore camptosomate case construction might be quite innate, yet variation may reflect environmental factors.
Larval case construction actually seems to involve two distinct developmental steps. During the first few instars, the case is mostly lengthened, but during the last instar it is greatly widened into a barrel-shape in preparation for pupation (Brown and Funk 2005). Although it may maintain a general, low-angle logarithmically spiraling shape, which in other species (e.g., snails and helicopsychid caddisflies) allows volume to increase with minimal use of material and without excessive surface area, pupal cases seem to be the product of two separate logarithmic trajectories (as found in some brachiopods, Aldridge 1999) (A. Aldridge personal communication).
There are several impressive traits in Neochlamisus building that highlight the sensitivity and acclimatization needed to build such a delicate structure. Female
Neochlamisus use their tarsal claws to hold onto the egg case, but never touch the more sensitive egg, which might be damaged by them (Brown and Funk 2005, Chapter II).
After watching several oviposition events, I have never seen a female accidentally dislodge one of the fecal plates. This maneuverability requires quick coordination and is based only on touch, since the female is blind to the entire process, as it is facing away from the egg. Fecal cases also require two distinct behaviors that must be regularly performed: 1) attaching horizontal additions to the dorsal rim and 2) opening the ventral suture along a previously determined suture and filling in the opening with vertical additions (Brown and Funk 2005, Chapter II). The second aspect adds a new dimension not found in the simpler uniform tubes of caddisflies. Once completed, the case wall is uniformly thick across its circumference, except for a longitudinal indention at the ventral
suture. This represents a thinner point in the case wall, which may make cutting through the case for further expansion easier. The fact that the case wall is otherwise of uniform thickness is amazing considering the builder does it all from an internal position. Perhaps larvae also add another coat of feces to the entire internal wall of the case, explaining its smooth internal surface (Chaboo et al. 2008). Since the case is constantly being
expanded, it is unknown how often such a procedure would be necessary or what triggers it.
Neochlamisus larvae are capable of repairing damage to their cases (Briggs, 1905, Erber 1988, Brown and Funk 2005, Chapter II, see also Eisner and Eisner 2000 for repair in a cassidine), whereas repairing a structure is not guaranteed in other animal builders.
Gould and Gould (2007) relate an example of a wasp that builds a large funnel out of smoothened mud to protect its egg chamber from parasitoids. Once one section of the funnel is created, the wasp will never return to it, even if it is partially damaged, or if the damage negates the function of the funnel. So it seems that like other architects,
casebearers may be relying on a simple set of rules to produce their elaborate structures, but unlike some architects, they may possess multiple blueprints and are willing to alter their plans when necessary.
What's next…
My thesis focuses on the unique architectures of the Camptosomata by studying species in the genus Neochlamisus. My studies explore the role that fecal cases play in the interactions of larvae and pupae with their biotic and abiotic environments. Chapter II provides a detailed account of the formation of the case and its role in the life history of