1
Four priority areas to advance invasion science in the face of rapid
1
environmental change
2
3
Anthony Ricciardi1, Josephine C. Iacarella2, David C. Aldridge3,4, Tim M. Blackburn5,6, James T.
4
Carlton7, Jane A. Catford8, Jaimie T.A. Dick9, Philip E. Hulme10, Jonathan M. Jeschke11,12,13, 5
Andrew M. Liebhold14,15, Julie L. Lockwood16, Hugh J. MacIsaac17, Laura A. Meyerson18, Petr 6
Pyšek19,20, David M. Richardson21, Gregory M. Ruiz22, Daniel Simberloff23, Montserrat Vilà24, 7
David A. Wardle25. 8
9
1Redpath Museum, McGill University, Montreal, Quebec, H3A 0C4, Canada 10
2Institute of Ocean Sciences, Fisheries and Oceans Canada, 9860 West Saanich Road, Sidney, British 11
Columbia, V8L 4B2, Canada 12
3Cambridge University, Department of Zoology, Pembroke Street, Cambridge, CB2 3QZ, UK 13
4BioRISC, St. Catharine’s College, Cambridge CB2 1RL, UK 14
5Centre for Biodiversity and Environment Research, Department of Genetics, Evolution and 15
Environment, University College London, Gower Street, London, WC1E 6BT, UK 16
6Institute of Zoology, Zoological Society of London, Regent’s Park, London, NW1 4RY, UK 17
7Maritime Studies Program, Williams College-Mystic Seaport, 75 Greenmanville, Mystic, CT 06355, 18
19 USA
8Department of Geography, King’s College London, 30 Aldwych, London, WC2B 4BG, UK 20
9Institute for Global Food Security, School of Biological Sciences, Queen’s University Belfast, Chlorine 21
Gardens, Belfast, BT9 5DL, UK 22
10Bio-Protection Research Centre, Lincoln University, PO Box 85840, Lincoln 7647, Canterbury, New 23
Zealand 24
11Leibniz-Institute of Freshwater Ecology and Inland Fisheries, Müggelseedamm 310, 12587 Berlin, 25
Germany 26
12Institute of Biology, Freie Universität Berlin, Königin-Luise-Str. 1-3, 14195, Berlin, Germany 27
13Berlin-Brandenburg Institute of Advanced Biodiversity Research, Königin-Luise-Str. 2-4, 14195 Berlin, 28
Germany 29
14US Forest Service Northern Research Station, 180 Canfield St., Morgantown, WV, USA 30
2
15Czech University of Life Sciences Prague, Faculty of Forestry and Wood Sciences, Praha 6 - Suchdol, 31
CZ 165 21, Czech Republic 32
16 Department of Ecology, Evolution and Natural Resources, Rutgers University, 14 College Farm Road, 33
New Brunswick, NJ 08901, USA 34
17Great Lakes Institute for Environmental Research, University of Windsor, Windsor, Ontario, N9B 3P4, 35
Canada 36
18Natural Resources Science, University of Rhode Island, 9 East Alumni Avenue, Woodward Hall 133, 37
Kingston, RI 02881, USA 38
19Institute of Botany, Czech Academy of Sciences, CZ-252 43, Průhonice, Czech Republic 39
20Department of Ecology, Faculty of Science, Charles University, Viničná 7, CZ-12844 Prague 2, Czech 40
Republic 41
21Centre for Invasion Biology, Department of Botany and Zoology, Stellenbosch University, Matieland 42
7602, South Africa 43
22Smithsonian Environmental Research Center, Edgewater, MD 21037, USA 44
23University of Tennessee, Department of Ecology and Evolutionary Biology, Knoxville, TN 37996, USA 45
24Estación Biológica de Doñana (EBD-CSIC), Avda. Américo Vespucio 26, Isla de la Cartuja, 41092 46
Sevilla, Spain 47
25Asian School of the Environment, Nanyang Technological University, 50 Nanyang Avenue, Singapore 48
49 50
Corresponding author: Anthony Ricciardi (email: [email protected] / Tel: 514-398- 51
4089) 52
53
Word count: 27,683 54
55
3
Abstract
56
Unprecedented rates of introduction and spread of non-native species pose burgeoning 57
challenges to biodiversity, natural resource management, regional economies, and human health.
58
Current biosecurity efforts are failing to keep pace with globalization, revealing critical gaps in 59
our understanding and response to invasions. Here, we identify four priority areas to advance 60
invasion science in the face of rapid global environmental change. First, invasion science should 61
strive to develop a more comprehensive framework for predicting how the behavior, abundance, 62
and interspecific interactions of non-native species vary in relation to conditions in receiving 63
environments and how these factors govern the ecological impacts of invasion. A second priority 64
is to understand the potential synergistic effects of multiple co-occurring stressors – particularly 65
involving climate change – on the establishment and impact of non-native species. Climate 66
adaptation and mitigation strategies will need to consider the possible consequences of 67
promoting non-native species, and appropriate management responses to non-native species will 68
need to be developed. The third priority is to address the taxonomic impediment. The ability to 69
detect and evaluate invasion risks is compromised by a growing deficit in taxonomic expertise, 70
which cannot be adequately compensated by new molecular technologies alone. Management of 71
biosecurity risks will become increasingly challenging unless academia, industry, and 72
governments train and employ new personnel in taxonomy and systematics. Fourth, we 73
recommend that internationally cooperative biosecurity strategies consider the bridgehead effects 74
of global dispersal networks, in which organisms tend to invade new regions from locations 75
where they have already established. Cooperation among countries to eradicate or control species 76
established in bridgehead regions should yield greater benefit than independent attempts by 77
individual countries to exclude these species from arriving and establishing.
78
4 79
Key words: biosecurity; climate change; ecological impact; invasive species; management; risk 80
assessment 81
82
Introduction
83
Invasion science – the systematic investigation of the causes and consequences of 84
biological invasions – is a rapidly evolving interdisciplinary field. Its explosive growth over the 85
past few decades mirrors societal concern over the upsurge in the global rate of invasions 86
(Seebens et al. 2017;Pyšek et al. 2020; Seebens et al. 2020) and reflects the fundamental and 87
applied importance of understanding how species spread into new regions, why some ecosystems 88
are more vulnerable to invasions, and what factors govern the impacts of non-native species. To 89
date, research addressing these issues has yielded valuable insights into the forces that structure 90
ecological communities, the relationship between diversity and stability, mechanisms of 91
adaptation and rapid evolution, causes of extinction and biotic homogenization, and the 92
connectedness between socioeconomic and ecological systems, among other phenomena 93
(Lockwood et al. 2013;Hui and Richardson 2019). More remains to be done to sharpen and 94
integrate these insights into predictive frameworks. In addition, pressure is increasing for 95
invasion science to adapt to emerging issues such as rapid advances in biotechnology, 96
accelerating global change, expanding transportation networks, abrupt landscape 97
transformations, and infectious disease emergence (Ricciardi et al. 2017; Nuñez et al. 2020).
98
Invasion science is a relatively young discipline (Ricciardi and MacIsaac 2008) that has 99
embraced diverse domains in ecology and cognate fields (e.g., population biology, biogeography, 100
evolutionary biology, paleoecology, physiology) and has formed linkages with disciplines related 101
5
to biosecurity – such as epidemiology, risk analysis, resource economics, and vector science 102
(Vaz et al. 2017). This multidisciplinary expansion reflects the increasing complexity of 103
biological invasions and their impacts (Richardson 2011; Pyšek et al. 2020).
104
Here, we consider how invasion science should adapt to the Anthropocene – an era of 105
burgeoning human influence, novel stressors, and rapid environmental change (Steffen et al.
106
2015; Waters et al. 2016). We are an international team of ecologists, with diverse and extensive 107
experience in biological invasions in many parts of the world. Our team gathered in September 108
2018 to consider emerging scientific, technological, and sociological issues which, if addressed, 109
should ensure that invasion science can more successfully contend with rapid global change.
110
Through consensus (see Supplemental Material), we arrived at four overarching issues, relevant 111
to a broad range of taxa, environments, and geographic regions, and which encompass some of 112
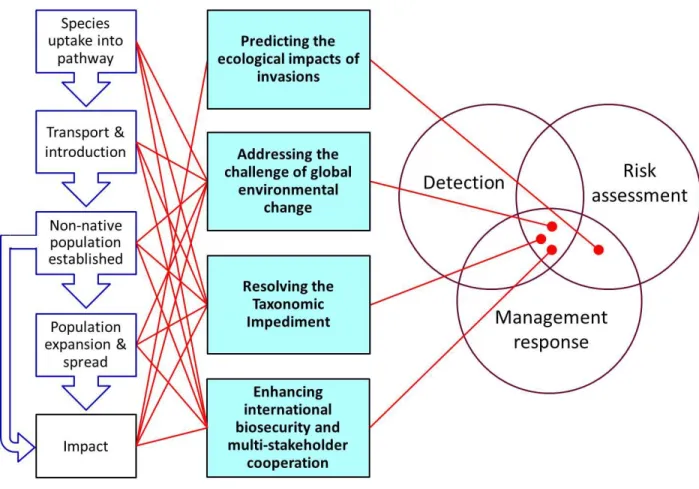
the most important challenges facing our field today (Figure 1).
113 114
1. Predicting ecological impacts of invasions under rapid environmental change
115
1.1. The need for greater predictive power: Major advances and ongoing challenges 116
1.1.1. Environmental context-dependency of impacts 117
While invasion science has made substantial progress in understanding how non-native 118
species arrive in new locations and establish self-sustaining populations (Catford et al. 2009;
119
Jeschke and Heger 2018), it has been less successful in forecasting when and where such species 120
will substantially affect their recipient environments (Ricciardi et al. 2013; Simberloff et al.
121
2013; Kumschick et al. 2015). Non-native species can affect ecological, economic, cultural, and 122
human health in diverse ways (Jeschke et al. 2014; Shackleton et al. 2018), but in this section we 123
focus on ecological impacts. Here, ‘impact’ is defined broadly as a measurable change to the 124
6
environment attributable directly or indirectly to the presence of a non-native species (Ricciardi 125
et al. 2013), and includes their effects on individual performance, population size and 126
composition of ecological communities of native species, which in some cases may be 127
irreversible (IUCN 2020).
128
Impact prediction is a long-standing, complex challenge. While rates of non-native 129
species introductions are increasing across regions (Seebens et al. 2017, 2020), impacts have 130
been recorded for only a small fraction of these species and the sites they invade (Ruiz et al.
131
1999; Ricciardi and Kipp 2008;Vilà et al. 2011; Hulme et al. 2013; Simberloff et al. 2013;
132
Evans et al. 2018b). It is generally assumed that most invasions have negligible environmental 133
consequences (Williamson and Fitter 1996), whereas a small proportion has significant and 134
sometimes enormous effects – an inverse magnitude-frequency distribution similar to that 135
associated with natural disasters (Ricciardi et al. 2011). However, uncertainty exists concerning 136
which cases truly reflect an absence of impact rather than a lack of study (Latombe et al. 2019).
137
Even well-known impacts exhibit substantial variation over time and space; invaders may remain 138
innocuous for years or even decades prior to becoming disruptive when, for example, 139
environmental change triggers a new impact (Crooks 2005; Coutts et al. 2018). The impacts of 140
any given invader can vary greatly among ecosystems (Strayer 2020) and across environmental 141
gradients within ecosystems (Kestrup and Ricciardi 2009; Stritar et al. 2010; Hulme et al. 2013;
142
Sapsford et al. 2020). Context-dependencies of invasion – that is, interactions among propagule 143
pressure, the traits of the invader, the composition of the recipient community, and the 144
physicochemical environment – have hardly been addressed by any formal body of theory, but 145
some overarching frameworks are now being explored (e.g., Cronin et al. 2015; Iacarella et al.
146
2015a; Dickey et al. 2020; Sapsford et al. 2020).
147
7
Coupled with the challenge of context-dependency is the sheer complexity of 148
mechanisms by which non-native species can interact with their environment (Ricciardi et al.
149
2013; Kumschick et al. 2015). Synergistic interactions, nonlinearities, time lags, threshold 150
effects, regime shifts, and indirect effects of non-native species are difficult to predict (Ricciardi 151
et al. 2013; Essl et al. 2015b; Kumschick et al. 2015; Aagaard and Lockwood 2016; Hui and 152
Richardson 2017; Strayer et al. 2017). Consequently, accurate risk assessment tools for sound 153
management decisions are still lacking.
154 155
1.1.2. Temporal variation and time lags of impacts 156
Factors affecting temporal variation in impact remain a major research gap, in large part 157
because of the vast majority of impact studies are conducted over very short time scales (Strayer 158
et al. 2006; Stricker et al. 2015). Time-since-invasion has been found to be an important correlate 159
of the ecological impacts of non-native species (Iacarella et al. 2015b; Evans et al. 2018a;
160
Zavorka et al. 2018), but time lags between establishment and peak impact have thus far evaded 161
prediction and are increasingly recognized as hindering risk assessment (e.g., Coutts et al. 2018).
162
Predictions of spatiotemporal variation in impact direction and magnitude could be improved 163
through experimental and theoretical investigations of the relationship between an invader’s per- 164
capita effect and its abundance (Yokomizo et al. 2009; Cronin et al. 2015; Sofaer et al. 2018;
165
Bradley et al. 2019; Strayer 2020). We must also consider the influence of spatial scale on per 166
capita effects or impacts measured in small plots and mesocosms; attempts to extrapolate these 167
effects up to landscape scales relevant to management (e.g., by calculating the product of the per 168
capita effect, local abundance, and range size of an invader) might not adequately capture 169
changes tobiodiversity, biotic interactions, and ecosystem function, and thus might 170
8
underestimate some large-scale consequences of invasion (Hawkins et al. 2015; Bernard-Verdier 171
and Hulme 2019; but see Dick et al. 2017b). Greater effort is required to test factors that mediate 172
indirect and multi-scale effects, particularly where an invader’s impact is transmitted across a 173
suite of interacting species (Feit et al. 2018).
174
Conservation interventions and ecosystem management must contend with significant 175
time lags between the onset of the environmental stressors and the expression of invader impacts, 176
and forecasting such phenomena is plagued by context dependencies and non-linearities (Essl et 177
al. 2015b, c; Coutts et al. 2018). An understudied issue is how to recognize and manage the 178
interactive and cumulative effects of time lags in ecological responses to invasion. Delayed 179
biodiversity responses (e.g., dominance shifts, species turnover, metapopulation dynamics, 180
extinction debt) to anthropogenic stressors such as invasion can lead to abrupt shifts in 181
ecosystem functioning (Essl et al. 2015b) and underestimation of rates of contemporary 182
biodiversity change (Essl et al. 2015c). Given the management implications of this phenomenon, 183
ecological responses to compounded and cumulative stressors are becoming an increasing focus 184
of theory, experiments, and time series analyses (Foster et al. 2016; Candolin et al. 2018;
185
Kleinman et al. 2019; Shinoda and Akasaka 2020).
186 187
1.1.3. Impacts on ecosystem processes 188
Demand is growing for reliable assessments and predictions of the ecosystem-level 189
impacts of non-native species, especially those impacts that affect the provision of ecosystem 190
services in rapidly changing environments (Vilà and Hulme 2017). This need reflects the larger 191
challenge of understanding how ecosystem function is altered by the combined effects of species 192
gains (invasion, range expansion) and losses (extinction, range contraction), which are 193
9
simultaneously consequences and drivers of global change. With few exceptions (e.g., Mascaro 194
et al. 2012; Kuebbing et al. 2015), work on how these two forces affect ecosystem functioning 195
has developed largely in isolation (Wardle et al. 2011). Owing to this disconnect, ecologists are 196
unable to predict over the coming decades the net ecosystem consequence of these two opposing 197
forces – specifically, whether or not species that are gained at local scales through invasion will 198
affect ecosystem process rates in a comparable way to those native species that are lost.
199
Moreover, despite the many ecosystem impacts revealed thus far (Ehrenfeld 2010; Vilà et al.
200
2011; Simberloff et al. 2013), few types of ecosystems and invaders have been studied relative to 201
those that exist (Crystal-Ornelas and Lockwood 2020). It is likely that an enormous number of 202
non-native species have affected individual performance, population sizes, and community 203
structure, though direct and indirect effects on native species (e.g. via competition, herbivory, 204
predation, hybridization, and as diseases or their vectors), or by changing the physical, chemical 205
or structural characteristics of the environment (Blackburn et al. 2014; IUCN 2020), in ways that 206
have not been documented (Carlton 2009; Simberloff 2011). Ecosystem-level impacts must 207
remain a major focus, with researchers taking advantage of available technological tools (e.g., 208
Asner et al. 2008). Further, research on how biodiversity loss affects ecosystem functioning must 209
be evaluated alongside effects of non-native species additions, to better understand how human- 210
driven species change will affect ecosystem processes across scales. For example, given that 211
community composition can influence biosphere-atmosphere exchange of greenhouse gases 212
(Metcalfe et al. 2011), how non-native species influence processes that underpin this exchange 213
relative to native species extirpations can have significant, currently unrecognized consequences 214
for climate change.
215 216
10 1.2. New and future challenges
217
1.2.1. Impacts of interventions for restoring ecosystem function 218
Co-occurring environmental stressors are increasing pressures to use non-native species 219
for restoring ecosystem functions eroded by native species loss (Mascaro et al. 2012; Castro- 220
Díez et al. 2019). The notion of restoring ecosystems that have lost important species by 221
substituting non-native species to perform key functions traces back at least to the 1980s 222
(Atkinson 1988) and has seen growing interest in recent years (Seddon et al. 2014a; Galetti et al.
223
2017; Pires 2017). Of particular interest are proposals and ongoing projects to establish species 224
to replace seed dispersers of plant species that have lost their ancestral native mutualisms 225
(Seddon et al. 2014a; Galetti et al. 2017), and large herbivores and carnivores to fulfill lost 226
trophic linkages (Svenning et al. 2016). These efforts are often listed under the rubric of 227
rewilding (Lorimer et al. 2015; Svenning et al. 2016). Calls for active rewilding to restore 228
ecological processes (Perino et al. 2019) have primarily focused on the reintroduction of native 229
species, but some practitioners have advocated a ‘flexible’ approach to restoration that entails 230
using non-native species (Ewel and Putz 2004; but see Sotka and Byers 2019) as well as the 231
reintroduction of species into parts of their native range from which they have been absent for 232
various lengths of time.
233
As with translocation to accommodate climate change (see section 2.2.3), proposals for 234
translocations to restore ecosystem functions (e.g., IUCN 2013; Aslan et al. 2014) have been the 235
subject of substantial discussion of potential risks and benefits (Nogués-Bravo et al. 2016;
236
Rubenstein and Rubenstein 2016; Fernández et al. 2017; Pettorelli et al. 2018; Perino et al.
237
2019). Lunt et al. (2013) have compared possible risks and benefits of translocations to restore 238
ecosystem functions and translocations to address climate change, pointing to the possibility of 239
11
addressing both goals simultaneously. To employ proposed decision tools and adhere to the 240
International Union for Conservation of Nature (IUCN) guidelines, both advocates and critics 241
increasingly agree that progress is required on more accurate risk assessments and on 242
characterization, categorization, and quantification of the environmental impacts of 243
translocations (Jeschke et al. 2014), as has occurred with the EICAT framework (Blackburn et al.
244
2014; Hawkins et al. 2015; Evans et al. 2016), which has been adopted as an IUCN standard 245
(IUCN 2020), and similarly for socioeconomic impacts, as has begun under the SEICAT 246
framework (Bacher et al. 2018).
247
Conversely, other efforts to conserve native species or restore ecosystems involve non- 248
native species eradication. Such interventionsshould be preceeded by a predictive risk 249
assessment of the indirect effects of invader removal (Bergstrom et al. 2009; Caut et al. 2009;
250
Ruscoe et al. 2011;Lindenmayer et al. 2017) and the legacy effects of invasion (Corbin and 251
D’Antonio 2012; Grove et al. 2015; Reynolds et al. 2017; Pickett et al. 2019). Eradication has 252
had demonstrable benefits to biodiversity (Baider and Florens 2011; Monks et al. 2014; Jones et 253
al. 2016), but targeting the removal of a single invasive species within an ecosystem that 254
contains several non-native species can be counterproductive. A predictive framework must 255
consider the topology of species interactions, both trophic and non-trophic, to determine when 256
single-species management may lead to unintended consequences (Glen et al. 2013; Ballari et al.
257
2016; Hui and Richardson 2019).
258 259
1.2.2. Burgeoning novel organisms 260
Escalating risks are associated with the intentional and unintentional release of novel 261
organisms (those with no analogue in the natural environment) through biotechnological 262
12
advances that create transgenic or genetically engineered organisms. For example, some 263
proposals for rewilding entail de-extinction – i.e., creation of various sorts of proxies of extinct 264
species for release to the wild. Versions of de-extinction are expected to become increasingly 265
feasible (Stokstad 2015; Shapiro 2017). The process involves either backbreeding (Stokstad 266
2015) or the reconstruction of the genome of an extinct species from recovered strands of DNA, 267
which can then be used either to modify or to replace the genome of a suitable living relative or 268
to genetically engineer embryos that can be implanted in a compatible host. Some 269
conservationists will advocate for such proxy species to be reintroduced to a suitable former 270
geographic environment (Seddon et al. 2014b), and perceived ecosystem management benefits 271
may arise from doing so (Church 2013). Environmental differences between contemporary and 272
historic habitats (Peers et al. 2016) might encourage further genetic manipulation to create better 273
adapted species. Depending on the length of time the proxy species has been extinct and the 274
method used to produce the proxy, introducing such entities to the wild is tantamount to 275
introducing a non-native species (IUCN 2013; IUCN/SSC 2016; Genovesi and Simberloff 2020), 276
an action that in the absence of predictive knowledge increases the likelihood of unintended 277
ecological consequences.
278
Advances in biotechnology will also facilitate the creation of self-replicating synthetic 279
cells designed for novel tasks such as contaminant remediation, carbon sequestration, and the 280
production of biofuels (Menetrez 2012; Azad et al. 2014; Singh et al. 2016; Dvorak et al. 2017).
281
As synthetic and transgenic organisms will contain combinations of ecological traits that are 282
unlikely to be encountered naturally, recipient communities will be evolutionarily naïve to these 283
organisms and could be predisposed to being altered by them (Saul and Jeschke 2015). Such 284
impacts could be subtle but far-reaching, as has been demonstrated for macroscopic transgenic 285
13
species (Post and Parry 2011; Vacher et al. 2011; Oke et al. 2013). Among the larger risks is the 286
capacity for such organisms to evolve in the wild and to exchange genes with other organisms 287
(Dana et al. 2012). Given the exponential growth of molecular technology, the rate of 288
development of such organisms could outpace progress in developing effective risk assessments 289
of their ecological effects. This issue emphasizes a need for greater integration of evolutionary 290
and microbial biology into invasion science, and for developing impact theory and risk 291
assessment methods that explicitly consider evolutionary change in both the invader and 292
interacting species.
293 294
1.3. The way forward: a theoretical framework and tools for impact management 295
1.3.1. Developing and expanding a theoretical framework of impact 296
To meet societal demands, invasion science must continue to build a body of theory for 297
understanding and predicting impacts from the level of populations to ecosystems (Ricciardi et 298
al. 2013; Blackburn et al. 2014; Bacher et al. 2018). Progress toward this goal requires that 299
hypotheses explicitly integrate abiotic and biotic context-dependencies, including biotic and 300
abiotic drivers of spatiotemporal variation in impact. This integration parallels and perhaps can 301
be informed by studies of how species loss affects ecosystem functioning in different 302
environmental contexts (Ratcliffe et al. 2017; Baert et al. 2018; Kardol et al. 2018). One example 303
of an integrative hypothesis is Environmental Matching (Ricciardi et al. 2013), which posits that 304
the per capita effects of an invader vary along environmental gradients such that they are 305
maximal where abiotic conditions more closely match the physiological optimum of the invader 306
(Kestrup and Ricciardi 2009; Iacarella et al. 2015a; Iacarella and Ricciardi 2015).
307
14
A second example that integrates context-dependence is the Ecological (or Functional) 308
Distinctiveness Hypothesis (Diamond and Case 1986; Vitousek 1990; Ricciardi and Atkinson 309
2004), which predicts that impact is most severe in communities missing species functionally 310
similar to the invader. This hypothesis is derived from two observed patterns with strong 311
empirical support. One such pattern is that a community’s lack of eco-evolutionary experience, 312
or ecological naïveté, determines its vulnerability to non-native consumers, parasites, pathogens, 313
and competitors (Sih et al. 2010; Saul and Jeschke 2015; Davis et al. 2019; Nunes et al. 2019;
314
Anton et al. 2020). The second empirically supported pattern is that the largest community-level 315
and ecosystem-level impacts are generated by invaders that use key resources differently or more 316
efficiently than natives do and that can alter disturbance regimes, habitat structure, or food web 317
configurations (Vitousek 1990; Funk and Vitousek 2007; Morrison and Hay 2011). Given that 318
more closely related species tend to be ecologically similar (Burns and Strauss 2011), it follows 319
that phylogenetic distance, or simple taxonomic relatedness, is a proxy for functional 320
distinctiveness. Thus, an allied hypothesis predicts that invaders representing novel taxa, once 321
established in the community, are more likely to affect native populations negatively than 322
invaders that are taxonomically similar to natives in the recipient community (Ricciardi and 323
Atkinson 2004; Strauss et al. 2006; Davis et al. 2019). Despite longstanding recognition of eco- 324
evolutionary experience as a driver of impact, most risk assessments do not consider 325
evolutionary context. The consequences of the contemporary evolution of non-native species 326
(e.g., Bertelsmeier and Keller 2018), and the effects of invaders on the evolution of native 327
species, are underexploited but promising areas of research (Saul and Jeschke 2015; van Kleunen 328
et al. 2018) that point to the importance of integrating evolutionary biology in ways that enhance 329
the predictive power of invasion science.
330
15
Several distinct, and over a dozen overlapping, hypotheses explain invader impact 331
(Ricciardi et al. 2013), and additional hypotheses addressing invasion establishment success 332
could potentially be extended to understanding impact (Catford et al. 2009; Jeschke and Heger 333
2018). These hypotheses could be organized into a coherent body of impact theory by 334
eliminating redundancies and identifying commonalities (e.g., through consensus mapping of 335
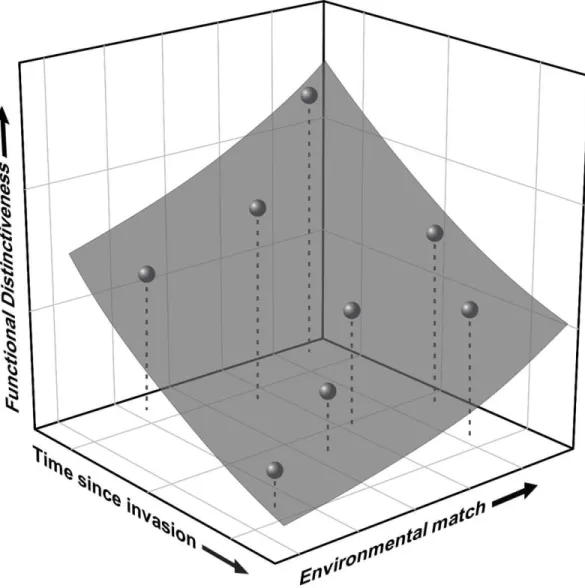
hypothesis networks; Enders et al. 2020). We can envision a general predictive framework built 336
upon multiple axes that consider, among other things, 1) abiotic and biotic environmental 337
context; 2) functional distinctiveness between native and non-native species; and 3) time-since- 338
invasion (Figure 2). The generality of hypotheses needs to be tested within various ecological 339
and evolutionary contexts using, for example, spatially distributed experiments such as those 340
employed to examine plant responses to nutrient enrichment and exclosure of mammalian 341
herbivores (Borer et al. 2014). Experimental and survey designs that incorporate eco- 342
evolutionary context have rarely been applied to the study of non-native species (but see Wardle 343
et al. 2001; Colautti et al. 2014; Grimm et al. 2020). To address this gap, we advocate 344
comparisons of conspecific populations across invaded and native ranges, recognizing that 345
invasions and impact outcomes are population-level phenomena. Such experiments could be 346
coordinated by collaborative global networks (Packer et al. 2017), which are a potentially 347
powerful approach to understand the factors that govern large-scale variation in invader impact 348
across climatic gradients, disturbance gradients, biogeographic realms, and boundaries of 349
evolutionary significance.
350
Moreover, scientists would profit by looking to other areas of ecology and evolution, 351
disease biology, and the social sciences, for theory that could potentially explain many 352
components of impact and seeking to integrate these approaches into invasion science. Several 353
16
classical ecological hypotheses, metrics, and concepts that have been tested in various contexts 354
relevant to invasions (e.g., theories addressing biological control, island biogeography, metabolic 355
scaling, resource utilization, competition) have arguably been underexploited by invasion 356
scientists.Experimental approaches that have sought to incorporate principles of trophic ecology 357
have revealed important patterns (Dick et al. 2017a, b; Cuthbert et al. 2018, 2020). For example, 358
prey switching (frequency-dependent predation) is a classical concept that has until recently been 359
virtually ignored by invasion science (Cuthbert et al. 2018, 2019). In recent years, the classical 360
functional response – the relationship between per capita consumption and resource density 361
(Solomon 1949; Holling 1959) – has been adapted and applied to forecasting and explaining 362
non-native species impacts through multispecies comparisons (Dick et al. 2017a, b; Dickey et al.
363
2018; Faria et al. 2019). The rationale for exploring these experimental approaches is that 364
invasion success and impact are often mediated by resource acquisition, a concept at the 365
foundation of many hypotheses in invasion science (Catford et al. 2009; Ricciardi et al. 2013;
366
Jeschke and Heger 2018) and that is relevant for both animals and plants (Rossiter-Racher et al.
367
2009; Ehrenfeld 2010). Indeed, several high-impact invaders have been found to be more 368
efficient at using limiting resources than their native and non-invasive counterparts (Rehage et al.
369
2005; Funk and Vitousek 2007; Morrison and Hay 2011; Dick et al. 2017a; DeRoy et al. 2020).
370
Broadening analyses to a more comprehensive community context could also help predict 371
impacts in different environmental contexts (Smith-Ramesh 2017). An underexploited approach 372
is to treat invaded communities as complex adaptive networks (Lurgi et al. 2014; Valdovinos et 373
al. 2018; Hui and Richardson 2019). Predictive information could potentially be gained from 374
modeling the dynamic responses of an ecological network, after developing appropriate metrics 375
17
of interaction strength, and thus identify resident species that are either facilitated or suppressed 376
by the invasion (Hui and Richardson 2019).
377 378
1.3.2. Toward more comprehensive quantifications of invader impact 379
There is growing interest in quantifying impacts beyond traditional ecological and 380
economic measures by using an ecosystem services framework that can capture information on 381
provisioning (e.g., food, timber, fuel), regulating (e.g., climate, floods, nutrient cycling) and 382
cultural services (Perrings 2010; Simberloff et al. 2013). For example, in highly-degraded 383
ecosystems some established non-native species may offer beneficial services to some 384
stakeholders (McLaughlan and Aldridge 2013), although any benefits of local cultivation of such 385
species must be weighed carefully against risks of further spread. Such accounting would also 386
need to consider negative impacts, which are diverse and substantive, on ecosystem services 387
(e.g., Walsh et al. 2016; Vilà and Hulme 2017; Milanović et al. 2020). However, at present we 388
know remarkably little about how even the most high-profile non-native species affect ecosystem 389
services (Vilà et al. 2010; McLaughlan et al. 2014), a problem related to the challenges of 390
evaluating ecosystem-level impacts (Simberloff 2011; Ricciardi et al. 2013). More reliable 391
quantification of potential ecosystem services of invasive species, coupled with a deeper 392
understanding of context-dependencies, would allow a more informed and comprehensive 393
impact assessment. To this end, the Millennium Ecosystem Assessment and, more recently, the 394
Intergovernmental Platform on Biodiversity and Ecosystem Services (IPBES), which have 395
examined how humans have altered ecosystems and these alterations have affected ecosystem 396
services and human well-being (Millennium Ecosystem Assessment 2005; Díaz et al. 2019), 397
could provide a suitable framework for developing protocols for risk assessment, perhaps 398
18
informed by the EICAT and SEICAT classification schemes (Hawkins et al. 2015; Bacher et al.
399
2018).
400
Comprehensive impact quantification is challenged by knowledge gaps that may render 401
risk assessments incomplete or misleading (Kumschick et al. 2015). One major gap is predictive 402
knowledge of the role of species traits, combinations of traits, and trait-environment interactions 403
in impacts, particularly at the ecosystem level. It is not clear under what situations the same 404
species traits that confer an ecosystem service can also damage an existing ecosystem service 405
(Vilà and Hulme 2017) or contribute to an ‘ecosystem disservice’ – properties or functions that 406
are disadvantageous to humans (Milanović et al. 2020). Another major context-dependency that 407
could distort risk assessment of a given invader is the presence of other invaders. Predictions, as 408
well as post-hoc assessments, are potentially hampered by synergistic or antagonistic interactions 409
between invaders, including those that can contribute to invasional meltdown – in which one 410
invader facilitates another, leading to compounded impacts and potentially self-reinforcing 411
effects (Simberloff and Von Holle 1999; Ricciardi 2001; Green et al. 2011). Disentangling the 412
influence of various species involved in meltdowns requires detailed experimental planning (e.g., 413
Braga et al. 2020), whereas invader interactions in multiple invaded ecosystems are generally 414
poorly studied (Kuebbing et al. 2013). It therefore seems likely that most synergistic effects go 415
unrecognized. Even where interactive effects do not occur, the cumulative effects of burgeoning 416
numbers of low-impact invaders on ecosystems have been virtually ignored. Approaches toward 417
quantifying and assessing the effects of multiple environmental stressors (Boyd et al. 2018;
418
Hodgson and Halpern 2018; Hodgson et al. 2019) could potentially be adapted for multiple 419
invading species and, furthermore, might be enhanced by efforts to collate experimentally- 420
validated invader interactions within global databases.
421
19 422
2. Addressing the challenge of global environmental change in invasion science
423
The second overarching issue is how invasion science can adapt to the onslaught of 424
global environmental changes presently altering the rates, dynamics, and impacts of invasions 425
through myriad drivers including climate change, overharvesting, extinction, pollution, 426
landscape transformation, and shifting trade patterns. Ecosystems are likely to become more 427
susceptible to invasions as these drivers degrade and modify food webs. For some native species, 428
global changes create physiologically intolerable or suboptimal conditions that lower relative 429
fitness (Catford et al. 2020) or provoke range shifts, further altering community composition and 430
susceptibility to invader impacts (Gallardo and Aldridge 2013; Wallingford et al. 2020).
431
Environmental change often affects native and non-native species differentially, modifying their 432
interactions and selection pressures through shifting abiotic and biotic ecosystem conditions 433
(Xiao et al. 2016; Meyerson et al. 2020; Stern and Lee 2020). This issue is well recognized and 434
has been widely investigated for several years, yet the need for research and management 435
solutions through the lens of invasion science is ongoing and increasing. Invasion science must 436
continue to develop an understanding of key issues regarding global environmental change 437
including interactions between invasions and other environmental stressors, climate adaptation 438
and mitigation strategies, and evaluating and managing species range shifts and translocations. In 439
this section, we primarily focus on climate change (Figure 3) but note that many other forms of 440
human-induced environmental change facilitate invasions and the relative dominance of non- 441
native species (Catford et al. 2014; Seabloom et al. 2015; Liu et al. 2017; Essl et al. 2019).
442 443
2.1. Ecological synergies between invasions and climate change 444
20 2.1.1. Non-native species performance
445
Species distributions worldwide are mostly determined by climate, tectonic movements, 446
and orographic barriers (Ficetola et al. 2017). Climate change will therefore have a major impact 447
on species range and distributions irrespective of whether species are native or non-native to a 448
particular region. However, differences in the magnitude of potential range shifts predicted for 449
non-native and native species will be determined by differences in their biology, such as 450
physiological tolerances and dispersal potential (Essl et al. 2019). The last decade has 451
accordingly seen major efforts to investigate the role of climate change in the introduction, 452
establishment, spread, and impact of non-native species (Hulme 2017).
453
Various meta-analyses have shown that non-native species often outperform and adjust 454
better than native species to a rapidly changing climate (Sorte et al. 2013; Oduor et al. 2016; Liu 455
et al. 2017). For example, hotter, drier environmental conditions enable non-native Asian tiger 456
mosquitoes to outcompete native tree-hole mosquitoes in the United States (Smith et al. 2015), 457
Eastern mosquitofish (Gambusia holbrooki) persist more successfully than native fish species in 458
France (Cucherousset et al. 2007), and non-native Monterey pine (Pinus radiata) to grow faster 459
than native conifers in Spain (Godoy et al. 2011). Warmer temperatures in freshwater 460
ecosystems will favor non-native species as these frequently have a greater heat tolerance than 461
related native species (Bates et al. 2013); similarly, in the Mediterranean Sea, increases in 462
temperature have facilitated the establishment of non-native tropical species (Raitsos et al. 2010).
463
A key element of climate change is an increase in the frequency and magnitude of 464
extreme climatic events, which can have greater effects on invasion than changes in average 465
conditions (Sheppard et al. 2012). Strong winds, floods, large waves, and storm surges can 466
transport organisms into new regions (Diez et al. 2012), as discussed below. Critically, extreme 467
21
climatic events like heat waves, fires, severe storms, droughts, and floods act as major 468
disturbances and will invariably destroy and damage resident native biota, reducing the uptake of 469
resources, and can also increase resource supply (Catford and Jones 2019). Such disturbances are 470
known to facilitate invasion (Davis et al. 2000), because many invasive species can take 471
advantage of fluctuations in resource availability caused by disturbances (Catford et al. 2012;
472
Singh et al. 2018). For example, European Bromus grasses that are highly invasive in North 473
America can exploit available soil moisture more efficiently and thus recover more rapidly than 474
native vegetation after drought (Harris 1967), enabling them to invade areas formerly dominated 475
by native woody species (Kane et al. 2011). Similarly, a non-native freshwater phytoplankton 476
species was able to invade and establish in a reservoir following the combined disturbance events 477
of macrophyte removal and extreme drought (Crossetti et al. 2019).
478 479
2.1.2. Non-native species range shifts 480
Shifts in temperature and rainfall patterns attributed to climate change can increase the 481
probability of establishment of non-native species, which were previously constrained by climate 482
(Walther et al. 2009; Hulme 2017) or climate-mediated interactions with native biota (Catford et 483
al. 2020). Increasing evidence indicates that non-native species tend to respond faster than native 484
species to climate change, with spread rates an order of magnitude higher than the velocity of 485
climate change (Hulme 2012). For example, non-native plants have expanded upwards in the 486
European Alps twice as fast as native species in response to warming (Dainese et al. 2017).
487
Nevertheless, climate change can lead to both increases (Kriticos et al. 2003; Barbet-Massin et 488
al. 2013; Gilioli et al. 2014) and declines (Bradley et al. 2009; Bellard et al. 2013; Xu et al. 2014) 489
in the geographical range of non-native species. A general finding is that, as a result of climate 490
22
change, the distribution range of non-native invertebrates and pathogens will expand, but range 491
contractions are mostly expected for non-native plants and vertebrates (Bellard et al. 2018). For 492
example, by the end of this century the suitable area worldwide for the red imported fire ant 493
(Solenopsis invicta) is predicted to be 21% greater (Morrison et al. 2014), whereas for the velvet 494
tree (Miconia calvescens) it is predicted that suitable habitat will be reduced in both its native 495
and introduced ranges (González-Muñoz et al. 2015). However, trends may differ between 496
terrestrial and aquatic environments. For instance, the warming of North American lakes is likely 497
to increase thermal suitability for southern species of fishes that could expand their distribution 498
poleward into non-native regions, potentially as far as the Arctic (Sharma et al. 2007; Della 499
Venezia et al. 2018).
500
Besides overall change in temperature and precipitation, extreme climatic events can also 501
help spread non-native species by overcoming dispersal barriers (Diez et al. 2012). For instance, 502
hurricanes promoted dispersal of non-native cactus moth (Cactoblastis cactorum) across the 503
Caribbean and into Mexico where it threatens native Opuntia species (Andraca-Gómez et al.
504
2015). Hurricane frequency was also positively correlated with the expansion of the non-native 505
grass Phragmites australis across wetlands along the Gulf Coasts of the USA (Bhattarai and 506
Cronin 2014). Likewise, flood events can increase pool connectivity and provide non-native 507
freshwater species access to newly inundated areas (Vilizzi et al. 2014). For example, floods 508
enabled the escape of cultured black carp (Mylopharyngodon piceus) in the Missouri River, US 509
(Nico et al. 2005), and tilapia cichlids in southeast Asia (Canonico et al. 2005) and have 510
facilitated the spread of zebra mussels (Dreissena polymorpha) in the Mississippi River 511
catchment (Tucker 1996). Nevertheless, the natural variability of climate makes it difficult to 512
23
attach high levels of confidence to some of the predicted changes, particularly those associated 513
with extreme weather events (Bellard et al. 2013).
514 515
2.1.3. Novel interactions and per capita impacts 516
Climate change will, in many cases, increase the introduction rate, establishment 517
probability, and spread rate of non-native species (Bellard et al. 2013), while simultaneously 518
facilitating extensive range shifts of native species (Inderjit et al. 2017; Pecl et al. 2017; Essl et 519
al. 2019), leading to novel ecological interactions and increased impacts. Range shifts are 520
expected to contribute to widespread biotic homogenization (where more species are shared 521
among communities) in some regions and the formation of novel communities in others (García- 522
Molinos et al. 2015). Diverse novel biotic interactions and assemblages will arise from divergent 523
responses of species and populations to climate change (Blois et al. 2013; Pecl et al. 2017). As 524
discussed previously, new biotic interactions often result in high impacts when resident species 525
have not co-evolved with newly arrived species (Ricciardi and Atkinson 2004; Cox and Lima 526
2006; Saul and Jeschke 2015). In some cases, range shifts of native species can cause impacts 527
similar to those involving non-native species (Sorte et al. 2013; Inderjit et al. 2017), although 528
impacts will be tempered by the eco-evolutionary experience of the resident species (sensu Saul 529
and Jeschke 2015). Few studies have addressed range shifts of native and non-native species as a 530
joint issue (Gallardo and Aldridge 2013; Sorte et al. 2013; Dainese et al. 2017; Inderjit et al.
531
2017; Singh et al. 2018).
532
While many studies have linked climate change to the spread of invasive species 533
(detailed above), the role of environmental factors in determining ecological impacts is 534
understudied (Dickey et al. 2020). Climatic conditions that shift towards the physiological 535
24
optimum of a non-native species could promote increased feeding rates, growth, or reproduction 536
that amplifies its competitive or predatory effects (Hellmann et al. 2008; Iacarella et al. 2015a).
537
For example, an invasive bryozoan is expected to have enhanced growth rates at warmer 538
temperatures in the Northwest Atlantic, with greater modeled impacts on kelp beds under future 539
climate conditions (Denley et al. 2019). Similarly, higher growth rates enable an invasive plant 540
to outcompete a native plant in China along higher latitudes in the field and at warmer 541
experimental temperatures (Wu et al. 2017). Predation rates of non-native species may also 542
increase when warming temperatures are within the physiological optima of the invader 543
(Iacarella et al. 2015a). For instance, the predatory response of an invasive freshwater amphipod 544
increases when exposed to elevated temperatures and infected by a common parasite (Laverty et 545
al. 2017). Given that non-native species are expected often to outperform native species in 546
response to environmental change, as discussed above, their competitive and predatory impacts 547
will likely also increase under these circumstances. A method has recently been developed that 548
incorporates the per capita and abundance effects of non-native species under altered variables 549
such as temperature, oxygen, salinity, and indeed any other variable in isolation or combination 550
(Dickey et al. 2020). This predictive method crucially also factors in the climate response of the 551
affected species (e.g., native prey), such that overall impact is holistically predictable. This 552
method is in its infancy and ground-truthing is now limited only by data (Dickey et al. 2020).
553 554
2.1.4. Changes to ecosystem services and human well-being 555
Research on the interaction between invasions and global environmental change is 556
essential to identify effects on ecosystem services and human well-being (Dukes and Mooney 557
1999; Walther et al. 2009; Pecl et al. 2017; Vilà and Hulme 2017). Although tools such as 558
25
SEICAT (Bacher et al. 2018) and INSEAT (‘INvasive Species Effects Assessment Tool’;
559
Martinez‐Cillero et al. 2019) have been developed to classify non-native species within a 560
framework of ecosystem services and human well-being, these tools rely on expert elicitation as 561
there are still surprisingly few quantitative data on the ecosystem services effects of even the 562
most prolific invasive species. This is, in part, owing to the context-dependent impacts of 563
invaders (see section 1) and because environmental change can alter the balance of positive and 564
negative effects (McLaughlan et al. 2014). For instance, disturbed river banks and roadsides in 565
Africa favor proliferation of the invasive tree, Prosopis juliflora (Shiferaw et al. 2019), which 566
increases local income from wood sales but reduces habitat suitable for livestock and results in 567
lower income from cattle sales (Linders et al. 2020). The predicted future effect of interactions 568
among climate, socioeconomic factors, and invasions on plant biodiversity hotspots constitutes 569
the greatest threat in emerging economies located in megadiverse regions of the Southern 570
Hemisphere (Seebens et al. 2015). Invasions and climate change also pose a combined threat to 571
native species in protected areas and thus seriously compromise conservation of biodiversity and 572
ecosystem services (Gallardo et al. 2017; Iacarella et al. 2020). Interactions between invasions 573
and climate change will also affect human health; for instance, climate change models predict an 574
increase in the life-cycle completion rate and extended periods suitable for development of the 575
invasive mosquito Aedes aegypti, a vector of arboviruses including dengue, zika, and yellow 576
fever, resulting in accelerated invasion in North America and China (Iwamura et al. 2020).
577
To investigate the effects of invasions on ecosystem services and human well-being, 578
models should integrate interactions among several components of global change, not only 579
climate change (Walther et al. 2009). Furthermore, studies should also explore these interactions 580
in productive systems such as managed forests, agriculture, and aquaculture (Thomson et al.
581
26
2010; Ziska and Dukes 2014; Liebhold et al. 2017). A major concern for these resource sectors is 582
that drought, warming, and elevated CO2 will affect the performance of non-native species (i.e., 583
pests, pathogens, and weeds) in complex and currently unpredictable ways. Research on their 584
impacts requires, for example, quantifying not only how altered environmental conditions 585
change weed and crop performance in isolation, but the magnitude of weed-crop competition on 586
crop damage (Ramesh et al. 2017).
587 588
2.2. Human responses to climate change that favor non-native species 589
2.2.1. Changes to invasion pathways 590
Global change is also altering invasion risk by promoting new commercial trading routes 591
and corridors. Shifting global economic forces (e.g., tariffs, manufacturing trends, recession, 592
regional conflicts, climatic disasters) determine trade volume and thus the frequency with which 593
aircraft or oceanic vessels travel between airports or seaports (Seebens et al. 2015). Such shifts 594
drive temporal rates of species introduction and the range of taxa that invade (Levine and 595
D’Antonio 2003; Hulme 2015; Bertelsmeier et al. 2018). For example, commercial shipping at 596
polar latitudes of North America and Eurasia is either planned or already occurring, providing 597
novel opportunities for introducing non-native species to Arctic waters (Miller and Ruiz 2014;
598
Chan et al. 2019). The Southern Ocean is likewise becoming increasingly vulnerable to species 599
introductions owing to increased propagule pressure from vessel traffic and reduced physical and 600
physiological barriers (Aronson et al. 2015; Hughes and Ashton 2017; Smith et al. 2017;
601
McCarthy et al. 2019; Cárdenas et al. 2020). Such human responses to climate change (Figure 3) 602
are altering the origins, taxonomic identity, and rate of introduction of non-native species in 603
27
terrestrial, freshwater, and marine habitats worldwide (Seebens et al. 2015; Early et al. 2016;
604
Della Venezia et al. 2018).
605 606
2.2.2. Climate adaptation: planting non-native species and adding infrastructure 607
As governments increasingly develop adaptive strategies to address climate change, 608
many of these strategies are likely to entail using non-native species. Proposed interventions 609
include initiatives to develop agricultural or aquacultural enterprises to deliver carbon-neutral 610
energy sources (e.g., macroalgae and plants for biofuels) using known invasive non-native 611
species (Barney and DiTomaso 2008). Pressure is also increasing to develop new varieties of 612
pasture species that can better cope with changing climates, such as drought-tolerant and disease- 613
resistant species, many of which are non-native in the countries in which they are sold and 614
planted (Driscoll et al. 2014). Increased development of green roofs, vertical gardens, and water- 615
saving horticulture to mitigate effects of climate change (Perini and Rosasco 2016) carry the risk 616
of introducing non-native species by promoting drought-tolerant plants or breeding drought- 617
resistant varieties, cultivars, or hybrids. Similarly, many large-scale tree-planting programs have 618
not led to the replenishment of degraded forests with native tree species, but rather to 619
afforestation of non-forest land, including biodiverse grsslands, with monocultures of non-native 620
trees. Such efforts include massive tree-planting campaigns using non-native trees with the aim 621
of mitigating the impacts of climate change and for other poverty alleviation (Brundu et al.
622
2020). Such plantings might not help offset greenhouse gas emissions as much as expected, 623
owing to unforeseen fluxes and complex system dynamics (Covey et al. 2012; Luyssaert et al.
624
2018; Popkin 2019). Indeed, inappropriate afforestation, especially in naturally treeless areas, 625
can have serious consequences for sustainable development, biodiversity conservation, and 626
28
ecosystem functioning (reviewed in Brundu et al. 2020). Furthermore, many species used in such 627
programs are highly invasive, which means that their impacts extend beyond areas identified for 628
afforestation (Brundu and Richardson 2016; Brundu et al. 2020).
629
Besides directly introducing species to sustain economic activities or to mitigate 630
emissions, governments at all levels are responding to environmental change by developing new 631
infrastructure. Strategies to combat sea-level rise have largely been addressed through 632
engineered solutions (armoring, raising road-beds, flood control structures). Each of these 633
adaptation strategies presents an opportunity for existing non-native species to expand their 634
range or impact and can create new suitable habitat for non-native species that arrive via ballast, 635
hull-fouling, or the marine aquarium trade (Bulleri and Chapman 2010). Offshore wind farms 636
also provide novel fouling habitats and ‘stepping stones’ for invasions (Adams et al. 2014; De 637
Mesel et al. 2015). Similarly, frequent droughts lead to efforts to provide secure water sources to 638
urban populations, including construction of dams, canals, and other water-diverting mechanisms 639
that can spread non-native species (Strayer 2010; Zhan et al. 2015; Gallardo and Aldridge 2018).
640
However, infrastructure developments can be designed to reduce their suitability as novel 641
habitats or invasion routes for invasions by non-native species, by minimizing environmental 642
disturbances or emulating natural habitats (Dafforn et al. 2015).
643 644
2.2.3. Species translocations for conservation 645
Conservation scientists have introduced species to locations outside their native range for 646
three main reasons: (1) to avoid extinction caused by an introduced species, often an introduced 647
predator; (2) to restore an ecological function (as detailed in section 1.2.1); or (3) to allow 648
species' ranges to keep up with climate change (Corlett 2016). Introductions to accommodate 649