The fact that I am where I am today is a reflection of the guidance and support I have received over the past 15 years (and the years leading up to my writing that paper). I am very fortunate to have a mentor who cared so much about the success of the science and the student at the same time. Finally, I am grateful for the opportunities, facilities, and people outside the lab that I have worked with here at Vanderbilt.
These peptides are then digested into individual amino acids by exopeptidases within the parasite cytosol, where they are used for growth and maturation. As Hb is degraded to its simpler form, monomeric heme (Fe(II) protoporphyrin IX, Fe(II)-PPIX) is also released into the digestive food vacuole of the parasite.8, 9. Hemoglobin is degraded within the digestive food. parasite vacuole by a group of aspartic acid proteinases (plazepsins PfPM1, PfPM2, PfPM4, and PfHAP), cysteine proteinases (falcipains PfFP2, PfFP2', and PfFP3), and the metalloproteinase falcilysin.
Current malaria research uses two chemically and structurally identical crystal forms of the malaria pigment. From these observations, it was suggested that the persistence of Hz may either be an intrinsic property of the biomineral or.
Arginine
Extraction of biological material associated with crude Hz identified a range of host- and parasite-derived lipids, proteins, and nucleic acids.31 Analysis of the lipid fraction identified products of secondary lipid peroxidation, including and 15-. 75 and triggering changes in gene expression.76, 77 Given that Hz is clearly involved in the pathogenesis of malaria, the activity of this Hz-related lipid peroxidation prompts further investigation of putative roles in the immunomodulation that occurs during. It is well established that heme compounds are efficient mediators of non-enzymatic lipid oxidation, supporting the hypothesis that reactive lipid peroxidation products are formed during lipid oxidation by Hz.
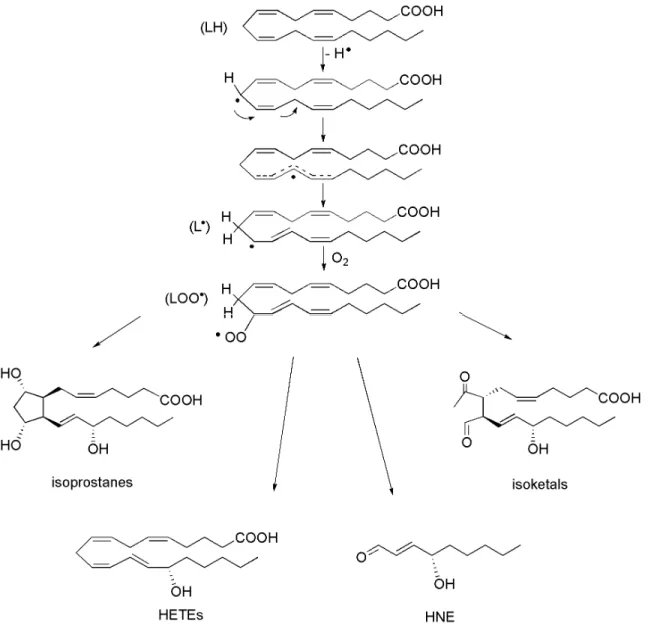
The mechanisms associated with lipid peroxidation include non-enzymatic or enzymatic reactions between molecular oxygen and a lipid molecule. These primary lipid peroxidation products are called N-HPETEs, where N identifies the carbon atom modified by the hydroperoxide group. Importantly, these intermediates lead to a diverse range of secondary lipid peroxidation species, including complex mixtures of regio- and strereo-isomers of isoprostanes, HETEs, HNE, and isoketals (IsoK), among others (Figure 12).
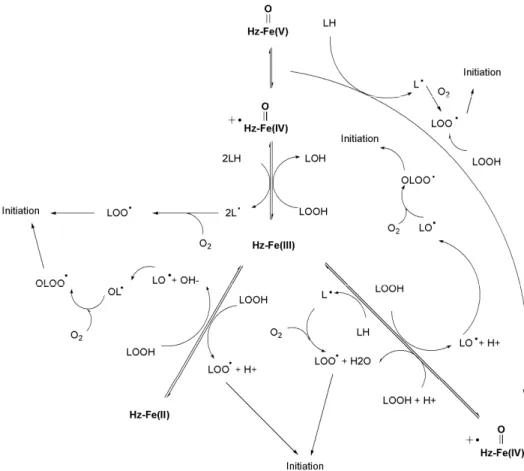
Free radical-mediated lipid peroxidation occurs through a mechanism consisting of three distinct steps: initiation, propagation, and termination (Figure 13). However, it is also well established that iron is intimately associated with lipid peroxidation.81 Iron and free heme (released during hemoglobin catabolism) mediate the generation of several redox-active species including peroxide and both superoxide and hydroxyl radicals.
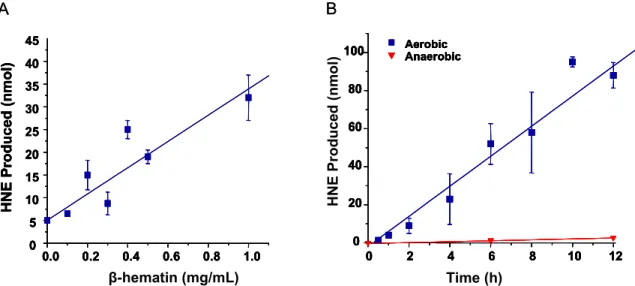
The purification was assessed by monitoring the absorption of the Soret heme band in the washes. The ability of BH to mediate HNE formation confirmed the putative reactivity of the dimeric iron core. Hydrophobic interactions suggest that BH-generated lipid peroxidation products are preferentially adsorbed to the biomineral surface.
To investigate the reactivity of BH in an appropriate model system, the ability of the biomineral to mediate peroxidation of lipids derived from erythrocyte membranes (i.e., ghost cells) was investigated. Analogous mechanisms describing the formation of HNE from arachidonic acid are proposed using 11- and 15-HPETE as precursors to 4-HPNE.93 Alternatively, the non-enzymatic formation of the PGH2 intermediate leads to the formation of several isoprostanes and isoketals. Inhibitory effect of the products generated from the interaction of BH with ghost cells.68 Cells were processed and analyzed as described in the experimental procedures.
Furthermore, biologically relevant levels of the individual lipid peroxidation products HNE and 15-HETE mimicked the dysfunctional response to hemozoin phagocytosis, suggesting the basis of Hz activity. The global effects of the individual components of Hz remain largely unexplored in the context of malaria; therefore, microarray technology was used to examine genes.
Comparative Analysis of the Gene Expression Response to HNE, BH, and Latex Beads Latex Beads
Consequently, Part II focuses on 15-HETE-induced steady-state gene expression changes in activated RAW 264.7 cells in the context of a nonspecific malaria toxin that may be involved in disease pathophysiology. Cytotoxicity profiles of RAW 264.7 cells treated with 15-HETE were determined by flow cytometry: LPS-stimulated cells were treated with increasing concentrations of 15-HETE for 24 h, and changes in viable, apoptotic, and they were dead. 15-HETE/L RBC.32 Statistically significant (p ≤ 0.025) changes in gene expression (fold change ≥1.8 relative to stimulated cells) were identified by microarray analysis.
Given that the aim of this study was to investigate possible changes in gene expression caused by 15-HETE during hemozoin phagocytosis, differentially expressed mRNAs were monitored under the same conditions by latex bead challenge (Figure 47). 15-HETE had a much greater effect on induction of gene expression than repression (293 transcripts vs. 100 transcripts). Venn diagrams show the intersection of genes transcriptionally altered by 40 μM 15-HETE with those altered by latex bead treatment and serum opsonized BH (0.1 mg/mL).
Ingenuity pathway analysis (IPA) was used to identify biological interaction networks associated with 15-HETE-mediated expression changes. Among the transcripts modulated by 15-HETE exposure, 263 were suitable for analysis based on IPA criteria, mapping to 11 relevant interaction networks. Transcripts altered ≥1.8-fold (p ≤ 0.025) in 15-HETE-treated RAW 264.7 cells (ie, focal genes) were overlaid on a global molecular network developed from information contained in the Ingenuity Knowledge Base ( IKB).
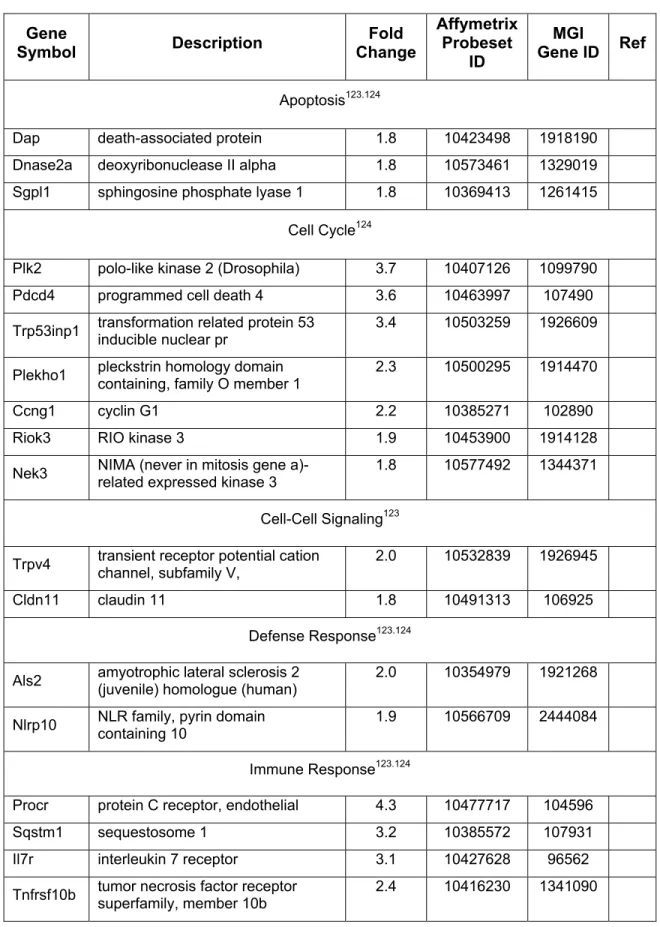
Networks reveal direct interactions between focal genes altered by 15-HET treatment and associated molecules within IKB. Comparison of the biological functions modulated by 15-HETE (Table 5) with those affected by BH and HNE shows that all three Hz constituents affect the genes 'Cell Cycle', 'Cell Morphology' and 'Cellular Assembly and Organization' after 24 hours affected. Consistent with the microarray results in terms of magnitude and direction of change, 15-HETE stimulated the expression of Arf3 (ADP-ribosylation factor 3), Cldn11 (claudin 11), Cxcl11 (chemokine (C-X-C motif) ligand 11), Mapk14 . mitogen-activated protein kinase 14), Prdx1 (peroxiredoxin 1), and Sdc1 (syndecan 1), and suppressed the expression of Egr1 (early growth response 1).
Changes in gene expression induced by 15-HETE in the current study were compared with two groups of transcripts. In the present study, 15-HETE induced the expression of several integrin signaling transcripts (e.g. Crkl, Rap2b, Arf3). In response to the large number of gene expression changes mediated by HNE, the global response to 15-HETE was examined in macrophage-like cells.
15-HETE can be incorporated into membrane lipids and alter vascular tone and EC permeability.116 Analyzes showed that the response to 15-HETE was mainly associated with altered expression of 'lipid metabolism' and 'small molecule biochemistry' genes. Although these results add insight and detail to the effect of 15-HETE on gene expression in macrophage-like cells, it should be noted that there are limitations to each model system.

Effects of Lipid Peroxidation Products on Matrix Metalloproteinase-9 Regulation in LPS stimulated RAW 264.7 Cells Regulation in LPS stimulated RAW 264.7 Cells
4-Hydroxynonenal Impairs LPS-Mediated Expression of Inducible Nitric Oxide Synthase
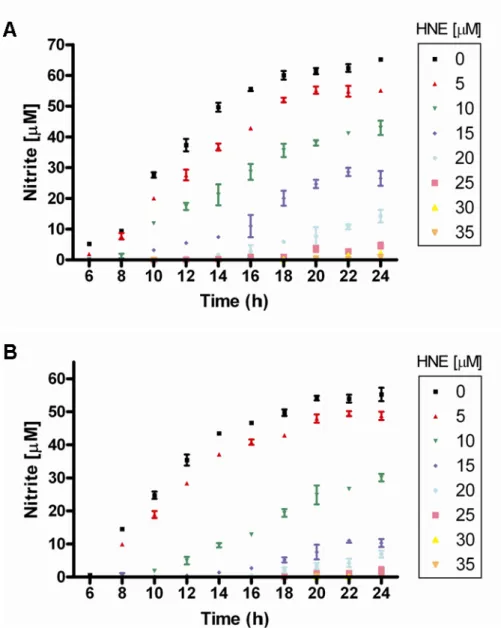
However, as shown in Chapter II, RAW 264.7 cells incubated with HNE are unable to generate NO. RAW 264.7 and J774 macrophage-like cells were treated with increasing concentrations of HNE (0–35 μM) and immediately stimulated with LPS to activate NF-κB signal transduction. Because IKK appears inactive in HNE-treated cells, a trend that has also been reported for cells exposed to acrolein185 and 15-deoxyprostaglandin J2113,186 which are structurally related to HNE, IKKα and IKKβ levels were assessed at 2 h after HNE treatment. by immunoblotting.
While IKKα and IKKβ levels appear stable, immunoblotting against reduced HNE-Michael adducts demonstrates HNE-modified protein in the molecular weight range of IKKα and IKKβ (Figure 66 A). Briefly, IKKα was purified from crude lysate by immunoprecipitation with an antibody against IKKα, and the IKKα eluent was examined for the presence of an HNE adduct by immunoblotting with an antibody against reduced HNE-Michael adducts. The results demonstrate that HNE targets IKKα in culture, but do not provide information on the site of HNE adduction.
RAW 264.7 cells were stimulated with LPS and treated with increasing concentrations of HNE (0–35 μM) (A) Immunoblots against HNE-modified protein, IKKα and IKKβ 2 hours after HNE treatment (B) Immunoblots against IKKα and IKKβ 12 hours after HNE treatment. Structurally, IKKα and IKKβ are very similar (Figure 68): each subunit contains an N-terminal kinase domain, a leucine zipper region, and a C-terminal helix-loop-helix domain.187 Previous studies with RAW 264.7 have shown that the activity involves both IKKα and - β are abolished in the presence of auranofine, a thiol-reactive metal compound. Consequently, Cys178 and Cys179 within the activation loops of IKKα and IKKβ, respectively, are potential targets of HNE adduction.
The ability of HNE to adduct the cysteine residues of both IKKα and IKK β peptides was examined by mass spectrometry. The fragmentation pattern of the molecular ion [M+2H]+2 1516.2 is shown and corresponds to a reduced Michael adduct on the cysteine residue. The fragmentation pattern of the molecular ion [M+2H]+2 1584.2 is shown and corresponds to a reduced Michael adduct on the cysteine residue.
Consistent with these results, HNE has been reported to differentially affect signal transduction.71 The data presented in this chapter provide insight into the biological activity of HNE and its ability to. Schematic showing the proposed mechanism of MMP9 regulation by HNE in RAW 264.7 macrophage-like cells stimulated with LPS. The data confirm that HNE inhibits murine macrophage NF-κB signaling at the level of IKK activity (Figure 73).
NONO
Selected genes associated with (A) specific genes or gene products correlated with malaria infection or (B) genes classified under specific. P; Adisa, A.; Dogovski, C.; Tilley, L., Food vacuole-associated lipid bodies and heterogeneous lipid environments in the malaria parasite Plasmodium falciparum. Schwarzer, E.; Turrini, F.; Ulliers, D.; Giribaldi, G.; Ginsburg, H.; Arese, P., Reduction of macrophage functions after ingestion of erythrocytes infected with Plasmodium falciparum or isolated malaria pigment.
Olliaro, P., The heme part of malaria pigment (β-hematin) mediates the inhibition of nitric oxide and tumor necrosis factor-α production by. Odeh, M., The role of tumor necrosis factor-alpha in the pathogenesis of complicated falciparum malaria. Prakash, D.; Fessel, C.; Jain, R.; Cazenave, P.-A.; Mishra, Gyan C.; Pied, S., Clusters of cytokines determine the severity of malaria in patients infected with Plasmodium falciparum from endemic areas of central India.
Schwarzer, E.; Alessio, M.; Ulliers, D.; Arese, P., Phagocytosis of the malarial pigment, hemozoin, impairs the expression of major histocompatibility complex class II antigen, CD54 and CD11c in human monocytes.