C
ITRUSM
ITESThis page intentionally left blank
C ITRUS M ITES
Identifi cation, Bionomy and Control
Vincenzo Vacante
Professor of General and Applied Entomology
Department OASI, Mediterranean University
Reggio Calabria, Italy
CABI is a trading name of CAB International
CABI Head Offi ce CABI North American Offi ce
Nosworthy Way 875 Massachusetts Avenue
Wallingford 7th Floor
Oxfordshire OX10 8DE Cambridge, MA 02139
UK USA
Tel: +44 (0)1491 832111 Tel: +1 617 395 4056
Fax: +44 (0)1491 833508 Fax: +1 617 354 6875
E-mail: [email protected] E-mail: [email protected]
Website: www.cabi.org
© V. Vacante 2010. All rights reserved. No part of this publication may be reproduced in any form or by any means, electronically, mechanically, by photocopying, recording or otherwise, without the prior permission of the copyright owners.
A catalogue record for this book is available from the British Library, London, UK.
Library of Congress Cataloging-in-Publication Data Vacante, Vincenzo.
Citrus mites : identifi cation, bionomy and control / Vincenzo Vacante.
p. cm.
Includes bibliographical references and index.
ISBN 978-1-84593-498-9 (alk. paper)
1. Citrus—Diseases and pests. 2. Mites. I. Title SB608.C5V33 2010
634′.30496542–dc22
2009022588
ISBN-13: 978 1 84593 498 9
Typeset by AMA DataSet, Preston, UK.
Printed and bound in the UK by the MPG Books Group.
The paper used for the text pages in this book is FSC certifi ed.
The FSC (Forest Stewardship Council) is an international network to promote responsible management of the world’s forests.
In memory of Carlo Vidano, Master of Entomology and Ethics
This page intentionally left blank
vii
Contents
Preface x Acknowledgements xiv Part I Introduction
1 Citriculture and Injurious Mites 3
1.1 Introduction 3
1.2 Citriculture 3
1.2.1 Species and varieties cultivated 4
1.2.2 World production 7
1.2.3 Fruit exports 7
1.2.4 Juice production 8
1.3 Citrus Mites and Their Economic Importance 9
2 Introduction to Acari 11
2.1 Introduction 11
2.2 Morphology and Structure 11
2.2.1 Body division and external morphology 11
2.2.2 Gnathosoma 14
2.2.3 Idiosoma 16
2.2.4 Legs 16
2.3 Classifi cation 17
2.3.1 Higher classifi cation 17
2.3.2 Suborder Prostigmata 19
viii Contents
3 Methods and Techniques 20
3.1 Collecting 20
3.1.1 Collecting from plants 20
3.1.2 Collecting in substrate 21
3.2 Preserving 21
3.3 Preparing 22
3.3.1 Clearing/maceration 22
3.3.2 Temporary mounts 22
3.3.3 Permanent mounts 23
3.4 Rearing 24
4 Plant Damage 25
4.1 Feeding Mechanisms 25
4.2 Feeding Symptoms 27
4.3 Plant Damage 28
4.3.1 Local damage 28
4.3.2 General alterations 30
5 Control 31
5.1 Chemical Control 33
5.2 Side Effects of Chemicals 33
5.3 Biological Control 35
5.4 Integrated Pest Management 36
Part II Citrus Mites
6 Key to the Identifi cation of Families, Subfamilies, Tribes,
Genera and Species 41
7 Phytoptidae Murray 55
7.1 Introduction 55
7.2 Morphological Characteristics and Systematic Outline 55
7.3 Phytoptinae Murray 56
8 Eriophyidae Nalepa 58
8.1 Introduction 58
8.2 Morphological Characteristics and Systematic Outline 58
8.3 Eriophyinae Nalepa 61
8.3.1 Aceriini Amrine et Stasny 61
8.4 Cecidophyinae Keifer 67
8.4.1 Colomerini Newkirk et Keifer 67
8.5 Nothopodinae Keifer 68
8.5.1 Nothopodini Keifer 68
8.6 Phyllocoptinae Nalepa 70
8.6.1 Anthocoptini Amrine et Stasny 70
8.6.2 Calacarini Amrine et Stasny 84
8.6.3 Phyllocoptini Nalepa 87
Contents ix
9 Diptilomiopidae Keifer 101
9.1 Introduction 101
9.2 Morphological Characteristics and Systematic Outline 101
9.3 Diptilomiopinae Keifer 102
10 Tarsonemidae Canestrini et Fanzago 104
10.1 Introduction 104
10.2 Morphological Characteristics and Systematic Outline 104
10.3 Pseudotarsonemoidinae Lindquist 107
10.3.1 Pseudotarsonemoidini Lindquist 107
11 Tenuipalpidae Berlese 113
11.1 Introduction 113
11.2 Morphological Characteristics and Systematic Outline 113
11.3 Brevipalpinae Mitrofanov 117
11.4 Tenuipalpinae Mitrofanov 155
12 Tuckerellidae Baker et Pritchard 163
12.1 Introduction 163
12.2 Morphological Characteristics and Systematic Outline 163
12.3 Tuckerella Womersley 166
13 Tetranychidae Donnadieu 172
13.1 Introduction 172
13.2 Morphological Characteristics and Systematic Outline 172
13.3 Bryobiinae Berlese 177
13.3.1 Bryobiini Reck 177
13.3.2 Hystrichonychini Pritchard et Baker 183
13.3.3 Petrobiini Reck 188
13.4 Tetranychinae Berlese 194
13.4.1 Tenuipalpoidini Pritchard et Baker 194
13.4.2 Eurytetranychini Reck 195
13.4.3 Tetranychini Reck 219
14 Conclusions 298
14.1 Systematics 298
14.2 Bio-ecology 299
14.3 Pest Status 299
14.4 Natural Enemies 300
14.5 Means of Control 301
14.6 Horticultural Practices 302
14.7 Prevention 302
14.8 Integrated Pest Management 303
References 305 Index 371
x
An exhaustive treatment of injurious mites associated with economic plants gives rise to the necessity, according to the large number of species involved and their particular bionomics, of an organic presentation divided into systematic categories and individual crops. This would facilitate the presen- tation of the subject and avoids the consolidated and reprehensible practice, which tends to simplify, either for convenience or opportunism, the problem of mites injurious to citrus in regard to the small number of traditionally known species, legitimizing inappropriate phytoiatric choices on the basis of decisions based on macroscopic examination and/or the colour of an organism rather than on more scientifi c parameters (morphological, biological, etc.), to the economic, toxicological and ecological detriment of the entire fi eld and societies that directly or indirectly enter into a physical relationship with farming and/or its products.
The present work is the result of this refl ection and aims to assess the problem of injurious mites associated with citrus in the world. The work basically consists of a bibliographical research of the phytophagous mites recorded on citrus throughout the world, integrated with the author’s knowl- edge. Various researchers (Quayle, 1938; Bodenheimer, 1951; Ebeling, 1959;
Chapot and Delucchi, 1964; Talhouk, 1975; Jeppson, 1978, 1989; Smith and Peña, 2002) have briefl y dealt with the subject on both a regional and global scale. The list of species associated with citrus in southern California pub- lished by McGregor (1956) is well known, together with those of Muma (1975) for Florida, Vacante et al. (1989) for the Mediterranean area and Dhooria et al. (2005) for India. A more organic work was published by Jeppson et al. (1975), who outlined a global picture of the problem of mites injurious to different economic plants and illustrated with a wealth of details the case of citrus. Recently, Gerson (2003) presented a list of species known for citrus throughout the world. However, an updated work in the fi eld is required that is able suitably to guide personnel (researchers, technicians and managers
Preface
Preface xi
of phytopathological departments) interested in the problem of citrus mites.
In total 104 species ascribed to the Phytoptidae Murray, Eriophyidae Nalepa, Diptilomiopidae Keifer, Tarsonemidae Kramer, Tenuipalpidae Berlese, Tuck- erellidae Baker et Pritchard and Tetranychidae Donnadieu families were treated. All species quoted are formally phytophagous, although a number of them pose no problem to citrus groves (Bryobiinae, some Tuckerellidae and Tetranychidae and Eriophyidae) and were included in the list of injurious mites for completeness and out of respect for the bibliographical informa- tion. However, treatment of these latter species allows for the discrimination of mites that occasionally infest citrus from those that are potentially or real- istically harmful, assisting the work of phytosanitary services.
The book presents the fundamental elements of the external morphology of the mites with the aim of providing a tool that helps in their identifi cation, of the high systematics of the species dealt with and the means and methods of collecting mites from plants and substrate, together with their preserva- tion and preparation for study. The structure of the mouthparts of mites is briefl y dealt with to introduce the fundamental aspects of symptomatology and damage.
A key is presented for the identifi cation of the basic systematic catego- ries (families, subfamilies, tribes, genera) and species, brief information on the morphology and systematics of each category, the main elements of morphology and bionomics (geographical distribution, bio-ecology, natu- ral enemies, symptomatology and damage) of the different species. The morphological description of each species is mostly correlated by original black and white drawings produced at the time by the various authors and, where possible, information is given about web sites (Table 4.1) where the natural colours and features of leaving injurious mites and their damages on citrus can be observed.
Under no circumstances did the author discuss the merits of decisions of a taxonomical nature. Morphological descriptions of the various species were dealt with by the contributions of specialists; generally they are reported exactly and in some cases were simplifi ed and adapted to the needs of the text. The information on natural enemies mainly regards the species collected on citrus, or introduced into this crop from other regions and/or investigated for the control of injurious mites. Among these, Phytoseiidae mites represent the greatest number of species, according to the catalogue of de Moraes et al. (2004). This choice does not involve any position of the author on their taxonomic status but only the attempt towards a given mini- mum order to the discussion, resulting in the ideas of different specialists on the systematics of Phytoseiidae mites being an open problem. Symptoma- tology and damage are reported only in the case of suffi cient certainty, supported by appropriate bibliographic references. In any case, for each family, several tables are presented summarizing the pest status and geograph- ical distribution of each species.
For each species a paragraph on control examines the chemical and biological means and the control strategies (chemical, biological and inte- grated pest management) available in the different regions of the world.
xii Preface
The information given is taken from the offi cial bibliography and where possible has been integrated with the author’s experience. In this respect, the information on acaricides refers to their chemical groups in order to avoid giving information exceeded by the rapid evolution of the market and/or licence to use, commonly distinct by the withdrawal of various substances from the market and the entry of other substances. The suggested data do not have a general relevance, since the use of various acaricides is subject to the laws of individual countries. It is therefore the responsibility of the reader to verify their applicability in different regions.
The complexity of the subject and the considerable physical dimensions of the citrus crop, spread throughout the subtropical and tropical regions of the world, do not facilitate its treatment and make reference to ecological, horticultural and socio-economic aspects, which often differ from one another. In this context, environmental factors are very important and may directly infl uence the bio-ecology of the different species and require strate- gic choices, which may vary from one region to the next for the same species, as in the citrus rust mite, Phyllocoptruta oleivora (Ashmead), which is the most important phytophagous mite pest in the warm and humid areas of Florida, together with the Texas citrus mite, Eutetranychus banksi (McGregor), while the citrus bud mite, Aceria sheldoni (Ewing), is not an economic prob- lem; on the other hand, in the warm and arid areas of California, the citrus red mite, Panonychus citri (McGregor), is the most feared followed by A. shel- doni and Ph. oleivora and is an economic problem only in coastal areas (Childers et al., 1996). Similar situations have occurred in Mediterranean areas and in other humid regions of the world for the pink citrus rust mite, Aculops pelekassi (Keifer).
Of equal importance are the productive choices, which vary between regions and continents. Indeed, the control of pests of products destined for fresh consumption generally requires more interventions than that desig- nated for processing. With the former, cosmetic appearance is a priority, while with the latter, the development and abscission of the fruit are unaf- fected until 50–75% of the surface of the fruit is damaged (Allen and Stamper, 1979). As a result, production in Florida destined for the fresh market is treated with plant protection products three to four times a year, compared with up to two treatments for that designated for processing, with one petro- leum oil and one acaricide treatment (McCoy, 1985; Browning, 1992). From this point of view, Mediterranean citrus crops, fundamentally directed at sat- isfying the needs of a market orientated towards fresh consumption, is considerably different from that of the USA and is therefore different in terms of control strategies.
The FAO (FAOSTAT, 2008) estimated for citrus a disparity in yield values (hectogram/ha) between different regions of the world, linked to various environmental and socio-economic causes, including variable availability of technical means and appropriate knowledge and/or specializations in the various fi elds of agricultural production. From this point of view, the knowl- edge of biotic adversities of crops and fruits helps to optimize quality and improve yield. In India, more than 30% of citrus production is lost every year
Preface xiii
as a result of damage by insect and mite pests (Pruthi and Mani, 1945; Butani, 1979a, b).
The above-mentioned facts may infl uence the agrarian economy of a region and do not allow for generalizations. Indeed, control of Ph. oleivora in Florida requires an annual expenditure of between 75 and 100 million dollars (McCoy, 1996b), while it poses no problem in other important citrus areas such as Italy where it is not present. Equally important is the case of the red- dish black fl at mite, Brevipalpus phoenicis (Geijskes), responsible for the trans- mission in the American continent of ‘Lepra esplosiva’ or ‘Leprosis’ (Childers et al., 2001, 2003b), a feared viral disease (CiLV), prevention of which costs approximately 100 million dollars per year in Brazil alone (Rodrigues et al., 2003) but which is non-existent and does not spread from the mite in the Mediterranean region. In lemon groves in Italy and Spain, up to three acari- cide treatments are carried out annually with a cost per hectare of approxi- mately 450 per treatment. The control may have toxicological and ecological consequences, as in the case of the use of dithiocarbamates, aldicarb or other substances, whose social cost is diffi cult to quantify.
In conclusion, it is likely that in some cases the abnormality of the adaptations of one or more species may have escaped our attention, in which case we offer our apologies and willingness to compare with other authors involved in the subject who would like to contribute with their suggestions to improve this work.
Vincenzo Vacante Reggio Calabria, May 2009
xiv
I am particularly grateful to Professor Uri Gerson of the Hebrew University of Jerusalem, Rehovot (Israel), for his critical review of the text and kind sug- gestions. In addition, I would like to thank Professor Enrico De Lillo of the University of Bari (Italy), Professor Stefano Colazza of the University of Palermo (Italy), Professor Tetsuo Gotoh of Ibaraki University Ami, Ibaraki (Japan), Professor Michel Bertrand, Professor Serge Kreiter and Professor Alain Migeon of the University of Montpellier and of INRA (France), Profes- sor Carlos H.W. Flechtmann, of the University of São Paulo (Brazil), Profes- sor Carlo Duso of the University of Padova (Italy), Professor Eduard A.
Uckermann of ARC-Plant Protection Research Institute (South Africa), Pro- fessor Kazuhiro Tanaka, Editor-in-Chief of Acta Arachnologica (Japan), Doctor Kim Hogeland, Permissions Administrator, University of California Press (USA) and Evert E. Lindquist of Agriculture and Agri-Food, Canada, who have helped me by sending bibliographic material or for giving permis- sion to use several drawings.
I would also like to thank Professor Franco Zagari and Piero Donin of the OASI Department of the Mediterranean University of Reggio Calabria (Italy), and colleagues in the same department, who have been helpful fi nancially toward the editorial initiative.
Lastly, I especially thank Miss Sarah Hulbert of CABI Publishing for her availability and for the invaluable suggestions that led to the publication of this book.
Acknowledgements
I Introduction
Overview of the world’s citriculture, species and varieties cultivated,
●
and the importance of injurious mites.
Introduction to morphology and classifi cation of mites.
●
Methods and techniques for collecting, preserving, preparing and rearing
●
mites.
Feeding mechanisms, symptoms and plant damage.
●
Biological, chemical and integrated control.
●
This page intentionally left blank
© V. Vacante 2010. Citrus Mites: Identifi cation, Bionomy and Control
(V. Vacante) 3
Citriculture and Injurious Mites
1
1.1 INTRODUCTION
A brief overview of the fundamental aspects (harvested areas, cultivated spe- cies and varieties, yield, production quantity, economic importance, etc.) of global citriculture helps to illustrate its economic importance and yield data facilitating the presentation of limiting factors, including biotic adversity such as pathogens, insects and injurious mites.
1.2 CITRICULTURE
According to Webber (1967), ‘the various species of the genus Citrus are all believed to be native to the subtropical and tropical regions of Asia and the Malay Archipelago, and to have spread from there to other sections of the world. They have been cultivated from remote ages, and prototype forms of the most important species are not defi nitely known’. The har- vested area of citrus extends from 40° parallel north to 40° south (Chapot, 1975). It has been calculated that about 140 countries grow citrus and the FAO estimated a total harvested area of 8,322,605 ha for 2007 (FAOSTAT, 2008).
In the American continent, the main producing countries are Brazil (915,056 ha), Mexico (524,000 ha), USA (376,050) and Argentina (148,500 ha), where orange, lemon, lime, grapefruit and several mandarins are cultivated.
In Asia, China possesses the largest harvested area in the world (2,008,700 ha) with different varieties of citrus and particularly mandarins and pomelos, followed by India (690,100 ha), Iran (243,500 ha), Pakistan (192,700 ha), Thailand (97,600 ha), Iraq (68,750 ha) and Japan (64,730 ha), where orange, grapefruit, mandarin, lemon, lime and other citrus species are produced.
4 Chapter 1
In Africa, Nigeria has the largest citrus-growing area (732,000 ha), fol- lowed by Egypt (137,370 ha), Morocco (79,300 ha) and South Africa (69,980 ha), where orange, lemon, mandarin, grapefruit and lime are cultivated.
In the Mediterranean region, Spain is the leading producer (313,850 ha), followed by Italy (173,009 ha), Turkey (94,600 ha), Greece (57,250 ha) and Israel (18,965 ha). In this area, the most common citruses are orange, lemon, grapefruit and mandarin; other citrus fruits, such as lime, lemon, bergamot and bitter orange, have a localized distribution.
1.2.1 Species and Varieties Cultivated
The orange (Citrus sinensis (Linnaeus) Osbeck) is the most widespread spe- cies in the world, cultivated from the equator to the colder environments of the distribution area of citrus. The FAO estimated that in 2007, worldwide orange cultivation covered a total of 3,905,780 ha variously distributed in dif- ferent continents (FAOSTAT, 2008). In the same year, it was calculated that the total worldwide harvested area of lemon (Citrus limon (Linnaeus) Bur- man) and lime (Citrus aurantifolia (Christmann) Swingle) was 911,726 ha; the harvested area of lemon involves mainly the Mediterranean area, California and Argentina. Limes are common in tropical areas, where they effectively replace lemon and are concentrated in a number of countries such as Mexico (140,000 ha in total). The harvested area of grapefruit (Citrus paradisi Mac- fadyen) and pomelo (Citrus grandis (Linnaeus) Osbeck) amounted to 289,248 ha and is widespread in China, the USA, Cuba, Mexico, South Africa and other countries. The pomelo is present throughout tropical and subtropical Asia. Other citrus fruits have a localized distribution, like the Italian berga- mot (Citrus bergamia Risso) and citron (Citrus medica Linnaeus) in Italy and Puerto Rico, or bitter orange (Citrus aurantium Linnaeus) in Andalusia (Spain).
The harvested area of tangerines, mandarins (Citrus nobilis Lour.) and clemen- tines (Citrus reticulata Blanco) covers approximately 2,146,597 ha.
1.2.1.1 Orange
The FAO has estimated that in the American continent, Brazil possesses the largest harvested area of orange (799,356 ha) (FAOSTAT, 2008). In this coun- try, the most common varieties are Pera, Valencia and Natal, full of juice and principally used for the production of juice. Some cultivars of Navel (Bahia, Bahianinha) are destined for the fresh market. In addition, they are culti- vated fruit varieties with low acidity (Laranja lima, Piralima, Lima tardia).
Mexico occupies fourth place with 325,000 ha (mostly cultivar Valencia) and production is largely for fresh consumption and juices. The USA occupies fi fth place with a cultivated area of about 270,000 ha, most of which is used by the processing industry for juice production. Florida accounts for 82% of production, with a prevalence of the Valencia cultivar. In California, different cultivars of Navel and Valencia are used for fresh consumption. Argentina, where there is a tendency to replace the old varieties (Hamlin, Calderon, etc.)
Citriculture and Injurious Mites 5
with new cultivars of the Navel and Valencia group, occupies second posi- tion with a total harvested area of 59,000 ha.
As regards the Asian continent, India (cultivar Mosambi, etc.) has an orange-producing area of 440,000 ha and represents the second largest pro- ducing country in the world, followed by China with 385,500 ha in total, where the best oranges are grown in the central areas and in modern crops oranges of Navel type are grown with imported cultivars (Robertson, etc.).
Iran is the sixth largest producer with 150,000 ha, Pakistan is eighth with 135,000 ha and Indonesia 11th with 72,400 ha.
Among the African countries of the Mediterranean Basin, Egypt has an orange-growing area of 85,000 ha and constitutes the tenth largest producer in the world, followed by Morocco with 50,000 ha and South Africa with 40,000 ha, which has abandoned the production of old varieties of orange (Tomango, Hamlin, Premier) and where production is now oriented towards cultivars from international markets (Navel, etc.).
In the Mediterranean region, Spain has a growing area of 140,000 ha and is the sixth largest producer of oranges, largely Navel (cultivars Navelina, New Hall, Washington Navel, Navelate, Lanelate), whereas Italy has 105,334 ha, with production fundamentally characterized by a predominance of red pulp varieties such as the Tarocco (Tarocco nucellare, Scirè, Gallo, Tapi, Messina, Meli, Ippolito), the Moro and Sanguinelli. Other Navel orange types occupy niche positions. A secondary interest has other cultivar blondes with- out Navel, such as the Oval Calabrese and Belladonna. The Valencia orange is more important. Greece and Turkey both have growing areas of 40,000 ha, Tunisia has 12,500 ha, Lebanon 9700 ha, Portugal 7400 ha and Israel (cultivar Shamuti, etc.) 5540 ha (Damigella and Tribulato, 1980; Reforgiato Recupero and Russo, 2009).
1.2.1.2 Lemon and lime
According to FAO data, in 2007, the global harvested area of lemon and lime was approximately 911,726 ha (FAOSTAT, 2008).
In the USA, the cultivation of lemon (cultivars Lisbon, Eureka, Rosem- berg, Ross) covers 25,000 ha, mostly located in California. In Argentina 45,000 ha of lemons (cultivars Genoa, Villafranca) are cultivated, marketed as fresh products or processed, and placed in the global market, including Europe. In the Mediterranean region, the main producers of lemons are Spain (cultivars Fino, Verna, Real) with 46,500 ha, Italy (cultivars Femminello, Monachello, Interdonato, Lunario, Lemox) with 30,046 ha and Turkey (cultivars Kütdiken, Interdonato, Italyan Memeli, Lamas, Molla Mehmet, Kibris) with 20,000 ha.
Greece (cultivars Magalene, Karystos, Adamopoulos) has 10,000 ha, Israel 1735 ha and Portugal (cultivar Gallego) approximately 1000 ha (Calabrese and Crescimanno, 1980; Calabrese and Barone, 2009b).
As regards the limes, although FAO data are combined with lemon, it is possible to draw conclusions on the main producing countries. In the American continent, Mexico is the major lime-producing country (cultivar West Indian
6 Chapter 1
or Mexican or Key lime), with a cultivated area of 140,000 ha, followed by Brazil with 50,440 ha, partly cultivated with lemon (cultivar Femminello and other), Peru with 21,000 ha and the USA in South Florida (cultivars Persiana, Key, Tahiti, Bearss). In Asia, India (cultivars West Indian or Mexican or Key lime, Mitha Nimbu, Indiana or Palestine) is the largest producing country in the world with 230,000 ha, followed by China with 63,450 ha, partly culti- vated with lemon, Iran with 41,000 ha and Thailand with 26,700 ha. In the Mediterranean region, in Egypt (cultivars Key, Indiana or Palestine) about 15,000 ha of limes are grown. In Morocco 1100 ha of a lime acid (cultivar Limûn Boussera) are cultivated (Continella, 1980; Calabrese and Barone, 2009a).
1.2.1.3 Grapefruit and pomelo
FAO estimates indicate that in 2007 the worldwide harvested area of grape- fruit and pomelo was equal to 289,248 ha (FAOSTAT, 2008). In the American continent, the major producers of grapefruit are the USA (62,000 ha), Mexico (16,000 ha), Argentina (12,500 ha) and Cuba (17,000 ha). Florida produces about 78% of US grapefruit and a large share of production is destined for the fresh domestic market and exports, especially pink-pigmented fruits (Red Ruby, Burgundy, Star Ruby, Ray Ruby). In Mexico, the favourable climatic conditions give the pigmented varieties a fi ne organolectic quality and a sat- isfactory coloration that promote their export to Europe and Argentina. In South Africa, 14,000 ha of grapefruit are cultivated, widely bred and destined for export. In Asia, the main producing countries of grapefruit and pomelo are China (62,500 ha), the largest producer in the world of clear or pink fl esh pomelo (cultivars Mato, Banpeyu, Red Shaddock, Webber, Dirado Buntan), Thailand (12,000 ha) (cultivars Kao Panne, Kao Phuang, Thong Dee), Syria (10,000 ha), India (8,100 ha) and Bangladesh (6000 ha). In the Mediterranean region, the most important country for grapefruit production is Israel (5370 ha), where a gradual replacement has started of the Marsh variety, characterized by its clear fl esh, with the Star Ruby, marked by its more or less intense pink pigmentation (Continella, 1980; Calabrese, 2009).
1.2.1.4 Mandarin and mandarin-like
In 2007, the FAO estimated a total mandarin and mandarin-like cultivated area of 2,146,597 ha (FAOSTAT, 2008), characterized by different cultivars throughout the world, such as clementines in the Mediterranean, satsumas in Japan, hybrids in California, Ponkan in China and other countries of South-east Asia.
In the American continent, the main producing countries are Brazil (61,000 ha), Argentina (32,000 ha), Mexico (30,000 ha) and the USA (18,000 ha), where 72% of tangerines (mandarins and mandarin-like) are produced in Flor- ida. In California, excellent tangerines and tangelos are produced in the south- ern areas. In Brazil, the second most important species of citrus is the mandarin (C. reticulata), whose most common cultivar is the Cravo (Laranja Cravo).
Citriculture and Injurious Mites 7
In Asia, China (1,411,000 ha) is the largest producer in the world, and modern satsumas as well as Mediterranean mandarins and tangerines of the Dancy variety originate from Whenzhou. The Ponkan is common through- out South-east Asia. In Japan, 51,000 ha are cultivated, largely satsumas and mandarins, in Pakistan 50,000 ha, in Iran 45,000 ha and 37,200 ha in Thailand.
In Africa, the most important countries are Egypt (37,000 ha), Morocco (27,000 ha) and South Africa (5100 ha).
In the Mediterranean region, clementines represent about 70% of the production, whereas the cultivation of satsumas is declining and that of late- ripening hybrids (variety Fortune) has increased in recent years, especially in Spain. The main producing countries are Spain (125,000 ha), the second largest producer in the world, following by Italy (35,829 ha), Turkey (30,800 ha), Israel (5320 ha) and Portugal (4200 ha). In Spain, there is a preva- lence of clementines, which constitute 25% of all citrus production, and major cultivars (Marisol, Fina, Nules, Oroval, Hernandina) are selected. In Portu- gal, the Encore cultivar, used only in local markets, is well known. In Italy, there has been a strong decrease in the Havana mandarin as well as a crisis with the Tardivo of Ciaculli cultivar (Calabrese and Crescimanno, 1980; Cala- brese and Pensabene Bellavia, 2009).
1.2.2 World Production
The FAO estimated that in 2007 worldwide production of citrus amounted to 115,650,545 tonnes (t) in total (FAOSTAT, 2008). The ten largest producers are Brazil (20,682,309 t), China (19,617,100 t), the USA (10,017,000 t), India (6,286,000 t), Spain (5,703,600 t), Iran (3,739,000 t), Italy (3,579,782 t), Nigeria (3,325,000 t), Turkey (3,102,414 t) and Indonesia (2,600,000 t). The estimated yield (hectogram/ha) placed in descending order Indonesia (359,116), Turkey (327,950), the USA (266,374), Brazil (226,022), Italy (206,913), Spain (181,730), Iran (153,552), China (97,660), India (91,088) and Nigeria (45,423).
The disparity in the values of yield among different regions of the world is linked to various environmental and socio-economic causes, including the variable availability of technical means, and appropriate knowledge and/or specializations in the various fi elds of agricultural production. This directly affects the quantitative and qualitative standard and the export opportuni- ties and/or consumption of fresh products. From this point of view, knowl- edge of biotic adversities of crops and fruits helps to optimize quality and improve yield. In different countries, production is destined in varying degrees for fresh consumption on the local and national markets, and pro- cessed into juice also for export.
1.2.3 Fruit Exports
Citrus fruits are present in all world markets and are widely present in coun- tries with developed economies. A strong global demand feeds major exports
8 Chapter 1
from producing countries. In 2002, citrus fruit per capita consumption was calculated to be approximately 22 kg per capita per year (UNCTAD from FAO data).
The FAO estimate for 2005 showed that the export of citrus fruits in the world was equal to 12,088,535 t (10.45% of total) for a value of US$6,935,692,000.
The greatest exporter of fresh citrus in the world is Spain with 3,021,194 t, followed by South Africa with 2,041,225 t, the USA with 936,048 t, Turkey with 894,493 t, Argentina with 644,384 t, Morocco with 559,170 t and China with 450,553 t (FAOSTAT, 2008).
The global export of lemon and lime was calculated to be 2,143,935 t, with a value of US$1,220,184,000. Mexico is the largest exporter of limes with 387,196 t for a value of US$26,300,000, whereas Argentina is the largest exporter of lemons with 369,483 t for a value of US$151,830,000, followed by Spain, the third largest exporter of lemons with 362,577 t for a value of US$281,220,000, Turkey with 355,656 t for a value of US$169,395,000, South Africa with 135,014 t for a value of US$57,012,000, the USA with 111,408 t for a value of US$86,878,000 and Italy with 40,584 t for a value of US$31,553,000 (FAOSTAT, 2008).
As regards oranges, in 2005 worldwide exports amounted to 5,270,262 t for a value of US$2,589,246,000. The main exporting countries were South Africa with 1,235,027 t for a value of US$272,7764,000, Spain with 1,116,274 t for a value of US$863,876,000, Morocco with 256,160 t with a value of US$115,387,000, Egypt with 214,165 t for a value of US$74,914,000, Greece with 209,821 t for a value of US$100,326,000, Turkey with 193,538 t for a value of US$75,918,000, Argentina with 169,359 t for a value of US$46,503,000 and Italy with 100,742 t for a value of US$65,372,000. The USA and China occupy the ninth and tenth places with 583,471 and 55,867 t, respectively, and with a value equal to US$384,016,000 and US$18,730,000, respectively (FAOSTAT, 2008).
In 2005, the export of grapefruits and pomelos worldwide amounted to 1,354,128 t for a value of US$619,590,000. The main exporting countries were South Africa with 585,675 t for a value of US$98,649,000, the USA with 219,385 t for a value of US$154,995,000, Turkey with 98,962 t with a value of US$50,217,000, Israel with 78,068 t with a value of US$47,939,000 and Argen- tina with 34,103 t with a value of US$12,881,000 (FAOSTAT, 2008).
As regards tangerines, mandarins and clementines, total global exports for 2005 were estimated to be 3,320,210 t for a total value of US$2,506,672. The main exporting countries were Spain with 1,512,619 t for a value of US$1,475,896,000, China with 372,131 t for a value of US$10,522,000, Morocco with 302,452 t for a value of US$221,943,000, Turkey with 246,337 t for a value of US$109,312,000 and South Africa with 85,509 t with a value of US$54,621,000 (FAOSTAT, 2008).
1.2.4 Juice Production
Citrus fruit processing accounts for approximately one third of total citrus fruit production. More than 80% of it is orange processing, mostly for orange juice production. According to the FAO, in 2005 exports of citrus juice equalled
Citriculture and Injurious Mites 9
5,200,753 t for an export value of US$3,930,898,000. The most important pro- ducers are Brazil with 1,778,506 t for a value of US$1,111,305,000, primarily interested in the production of orange juice and exported totally; the USA with 432,771 t for a value of US$372,975,000, largely consisting of orange juice and grapefruit, with 90% consumed in the internal market; and Spain with 240,825 t for a value of US$168,925,000 and consisting largely of orange juice (FAOSTAT, 2008). In the USA, Florida contributes signifi cantly to the production of orange juice and is the major producer of grapefruit. Brazil controls the international market for orange juice (1,777,599 t) and Argentina that of lemon (50,703 t), leading both the international price of the product reference in the remaining countries of the world. Chinese production is now equal to about 6108 t, whereas Italian production (74,902 t) is generally char- acterized by the pigmented red variety, with a demand of juice in several foreign markets. Italian lemons are most sought after for their essences rather than for the juice. Equally important are the essential oils of mandarin, whereas there is less demand for the juices and essences of clementines.
1.3 CITRUS MITES AND THEIR ECONOMIC IMPORTANCE
In addition to pathological aspects related to viruses (Tristeza, Leprosis, etc.), fungi (Phoma tracheiphila (Petri) Kantachveli et Gikachvili, Phtophtora spp., etc.) and other injurious organisms, a variety of vertebrate and invertebrate animals commonly attack citruses in the different regions of the world, caus- ing serious damage to the crops and/or the harvest. In this context, arthro- pods are very important and among them the mites and insects (Quayle, 1938, 1941; Ebeling, 1950, 1959; Bodenheimer, 1951; Reuther et al., 1989; Bed- ford et al., 1998). Mites and insects possess both appendages articulate, but the fi rst are eight-legged and the last six-legged. Insects show a greater degree of ecological fi tness, deriving from the richness of the faunistic structure and from numerous evolutionary adaptations. Little is known about mites, at least in some countries, but they are no less important in the ecological and phytopathological context. The problem has been studied in the different cit- rus regions of the world (McGregor, 1956; Gerson, 1971; Rasmy et al., 1972;
Muma, 1975; Jeppson, 1978; Mijuskovic, 1973a, b; Mijuskovic and Tomasevic, 1975; Vacante and Nucifora, 1985; Garcia Marí et al., 1986; Vacante et al., 1989).
Nevertheless, with the exception of the contribution of Jeppson et al. (1975) on mites injurious to economic plants, including the citrus, and a recent pub- lication by Gerson (2003), there has not been a contribution that presents organic and up-to-date information on mites injurious to citrus in the world.
In marginal productive areas or in less developed areas, this want may sig- nifi cantly interfere with pest control and the prevention of the accidental introduction of pests from other countries of the world, with economic, eco- logical and toxicological disadvantages.
The complexity of the subject and the wide physical expanse of citrus crops, spread throughout the subtropical and tropical regions of the world, do not facilitate its treatment and make reference to ecological, horticultural
10 Chapter 1
and socio-economic aspects which often differ from one another. Environ- mental factors are very important and may directly infl uence the bio-ecology of the different species and require strategic choices, which may vary from one region to another for the same species, as in the case of the different behaviour of the citrus rust mite, Ph. oleivora, Texas citrus mite, E. banksi, and citrus bud mite, A. sheldoni in the warm and humid areas of Florida and in the warm and arid areas of California (Childers et al., 1996). In India, several factors contribute towards the decline in the yield of citrus trees, and the threat from different insects and mites has been considered one of the most important factors (Bindra, 1970); the losses related to injurious mites are quite substantial, especially during years when climatic conditions are more favourable for their development (Dhooria et al., 2005).
The productive choices, varying among regions and continents, infl u- ence mite control. Production destined for fresh markets generally requires more interventions than that destined for processing juices. In the former, cosmetic appearance represents a priority, whereas the latter permits greater levels of tolerance (Allen and Stamper, 1979). In Florida, production destined for the fresh market is treated with chemicals three to four times per year, whereas that intended for processing is treated only up to twice per year, with one petroleum oil and one acaricide treatment (McCoy, 1985; Browning, 1992). Mediterranean citrus crops, primarily destined for the fresh market, are considerably different from that of the USA.
These facts have serious economic importance and do not allow for generalizations, as in the case mentioned in the Preface, of the control of Ph.
oleivora in Florida, which requires an annual expenditure of US$75–100 mil- lion (McCoy, 1996a, b), whereas it poses no problem in other important citrus areas where it is not present. Similarly, the reddish black fl at mite, B. phoeni- cis, responsible for the transmission in the American continent of ‘Lepra esplosiva’ or ‘Leprosis’ (Childers et al., 2001b), a viral disease (CiLV), neces- sitates a prevention that costs approximately US$100 million per year in Brazil alone (Rodrigues et al., 2003), but is non-existent and does not spread from the mite in the Mediterranean region. In lemon groves in Italy and Spain, up to three acaricide treatments are carried out per year with a cost per hectare of approximately ?450 per treatment.
© V. Vacante 2010. Citrus Mites: Identifi cation, Bionomy and Control
(V. Vacante) 11
2 Introduction to Acari
2.1 INTRODUCTION
Acari are small animals, the adults of which range from 300 to 500 μm in body length, except for some eriophyoids that are approximately 100 μm long or certain ticks whose female measures about 30,000 μm. From a system- atic point of view, they represent a subclass of Arachnida, subphylum Cheli- cerata, and are distinguished from insects, with which they are often wrongly associated, for the lack of antennae, mandibles and maxillae and the presence in the adults of four pairs of legs, with the exception of eriophyoids, which possess two pairs of legs. Their biological cycle develops through the stages of egg, larva and nymph. The larva is six-legged and the nymph eight-legged.
At present, approximately 48,000 species have been described (Halliday et al., 1999), but it is believed that the group will grow to number over 1,000,000 species (Walter and Proctor, 1999). As regards ecological adaptation, Acari have colonized most of the available aquatic and terrestrial habitats, and pos- sess a capillary ability to exploit the trophic resources available thanks to different dietary regimes (phytophagous, parasitic, predaceous, mycopha- gous, saprophagous, coprophagous, necrophagous). Some species are pho- retic (Krantz, 1978; Lindquist, 1984; Walter and Proctor, 1999; Krantz, 2009b).
2.2 MORPHOLOGY AND STRUCTURE
2.2.1 Body Division and External Morphology
Acari have a globular or subglobular body, sometime fusiform or worm-like, pale in colour, rarely lively, with or without inconspicuous abdominal segmentation and are divided into two principal morphological regions,
12 Chapter 2
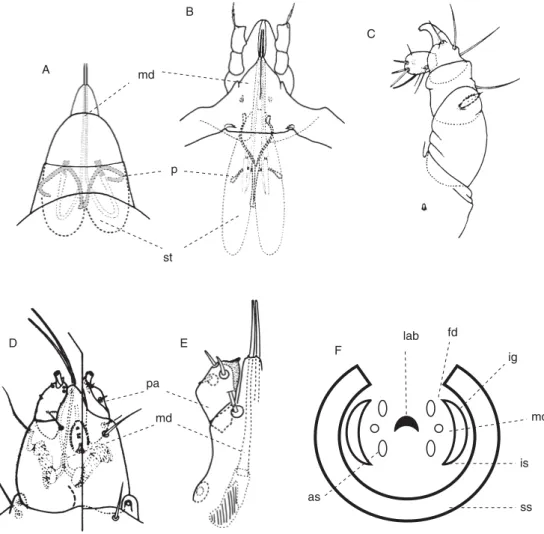
respectively called gnathosoma and idiosoma (Fig. 2.1). The limits between these regions of the body are not always well defi ned and are sometimes identifi ed by the existence of scarcely discernible sutures (circumcapitular, disjugal, sejugal, abjugal) (Krantz, 1978; Alberti and Coons, 1999).
Every group possesses a typical chaetotaxy, with a number of setae, characterized by shape, length, distance between their bases and function (Fig. 2.2).
ge
tr fe ti ta
Opisthosomal dorsum
IDIOSOMA
Prodorsum Gnathosoma
v2 sc1 sc2
c1 c2 c3
d1 d2
e1 e2
f1 f2
h1
Fig. 2.1. Tetranychus urticae Koch. Dorsal aspect of the female showing the different regions of the body (from Vacante and Nucifora, 1985, partially modifi ed); fe, femur; ge, genu;
ta, tarsus; ti, tibia; tr, trochanter. The setal notation is explained in the text on p. 176.
Introduction to Acari 13
ag
g1
g2
h3 ps2
ps1
B
C
D
E
F
G
H
I A
Fig. 2.2. (A) Ano-genital regions of female of Tetranychus urticae Koch (explained in the text) (from Vacante and Nucifora, 1985, partially modifi ed); (B) dorsal tactile seta of Bryobia praetiosa Koch; (C) duplex setae of tarsus IV of B. praetiosa (solenidion at left and tactile seta at right) (from Vacante, 1985, partially modifi ed); (D) solenidion and famulus of Thyreo phagus cooremani Fain; (E) supracoxal seta of Tyrophagus palmarum Oudemans (from Vacante, 1989); (F) trichobothrium of Humerobates rostrolamellatus Grandjean (from Vacante and Nucifora, 1985); (G) distal anasthomosis of peritreme of B. praetiosa; (H) distal anasthomosis of peritreme of Bryobia rubrioculus (Scheuten); (I) peritreme of Panonychus citri (McGregor) (from Vacante, 1985). The setal notation is explained in the text on p.176.
14 Chapter 2
Respiration may be cuticolar or tracheal, through paired stigmata or with peritremes of various shape and structure (Fig. 2.2). The tracheae may be placed dorsolaterally, ventrolaterally, anteriorly to the bases of chelicerae or near the base of the legs. Some characters (number, length, shape, location) of the stigmata are used in systematics.
The body presents a number of sensory organs, most of which are setae, primarily tactile or chemotactile (Fig. 2.2). The presence of ocelli and photo- sensitive organs is well known.
The presence of several cuticolar pores is attributable to that of an excre- tion organ. In addition, coxal glands are involved in the water balance and regulation of ion concentration (Krantz, 1978).
2.2.2 Gnathosoma
The gnathosoma or capitulum is the fi rst region of the body and derives from embrional cheliceral segment, and the derived biramous appendages of the second somite. From the fusion of the palpcoxae arises the subcapitulum, more or less conical and with a dorsal and longitudinal wrinkle hosting the chelicerae; it also possesses the preoral wrinkle, mouth and pharynx. The palps are located dorsolaterally. The dorsal anterior part of the propodosoma forms the prodorsum (Fig. 2.1).
Proterosoma Prosoma
Gnathosoma Precheliceral
I, II
III–IV V–VI VII–XVI
Prop Met
Podosoma
Hysterosoma Opisthosoma
Fig. 2.3. Diagram of primitive segmentation in the Acari (according to Coineau, 1974); prop, propodosoma; met, metapodosoma.
Introduction to Acari 15
The chelicerae are considered as derivative endopodal appendages of the fi rst somite and are placed dorsally compared with the opening mouth and commonly possess three segments called cheliceral base, digitus fi xus and digitus mobilis, respectively. The fi rst is basal and bears dorsally the digi- tus fi xus articulated with the distal digitus mobilis. The cheliceral bases may be partially or completely fused to each other and form, as in the case of tetrany- chids and other similar groups, the stylophore. The chelicerae may have a different morphology, as in the tetranychids that present the digitus mobilis modifi ed in a stylet fi t for piercing the cell wall (Fig. 2.4).
D
md
p
st A
B
C
E
ss is ig lab fd
md F
md pa
as
Fig. 2.4. Gnathosomal structures. (A) Dorsal view of gnathosoma of Tetranychus urticae Koch;
(B) dorsal view of gnathosoma and prodorsum (partially designed) of Brevipalpus phoenicis (Geijskes); (C) palp of Bryobia sp. (from Vacante and Nucifora, 1985); (D) gnathosoma with extended chelicerae of Tarsonemella africanus (Hirst); (E) apex of gnathosoma and palp of Asiocortarsonemus malayi Fain (from Lindquist, 1986); (F) diagram of transverse section of cheliceral and associated structures at level near apices of stylets (according to Lindquist, 1996);
as, auxiliary stylet; fd, fi xed digit; ig, infracapitular guide; is, infracapitular stylet; lab, labrum;
md, movable digit; p, peritreme; pa, palp; ss, stylet sheath; st, stylophore.
16 Chapter 2
The palps are postoral appendices, consisting of a variable number of articles (trochanter, femur, genu, tibia, tarsus and apotele), including from one or two to six (Fig. 2.4). The two coxae merge with each other and form the subcapitulum, which opens the mouth, bounded dorsally by the labrum and laterally by various structures (lateral labra, rutella, malae). The function of the palps is primarily related to the search for and handling of food, and usually the distal segment’s leading sensorial setae (mechanoreceptors, etc.), which acts as a support for the trophic activity and may provide the outlet of silk glands (Krantz, 1978, 2009a).
2.2.3 Idiosoma
The idiosoma is formed by the anterior propodosoma and posterior hys- terosoma. The propodosoma is anterior to the dorsal disjugal furrow, ante- rior to the ventral sejugal furrow, and includes the pairs of legs I and II. The hysterosoma is posterior to the dorsal disjugal furrow to ventral sejugal fur- row and includes legs III and IV (Fig. 2.3). The area bearing the legs derives from four embryonal somites and is also called podosoma; the part deriving from the fi rst two somites shelters the fi rst two pairs of legs and is the propo- dosoma, while the last part deriving from two other somites presents the other two pairs of legs and is called metapodosoma (Fig. 2.3) (Coineau, 1974;
Krantz, 1978; Alberti and Coons, 1999; Krantz, 2009a).
The part of the hysterosoma excluding legs III and IV constitutes the opisthosoma and derives, according to the systematic groups, from six to 13 embryonal somites and shelters the genital and anal openings (Fig. 2.3) (Coineau, 1974). Its shape is from ovoid to fusiform and presents a cuticle variously striate, with furrows or covered by plates or shields, of shape and size variable in the different systematic groups. Male and female genital openings open both anteriorly or posteriorly.
The sperm transfer may take place directly through the intrusion of the male copulatory organ or aedeagus (Fig. 2.5) in the female genital opening or in special extragenital structures located on the rear end of the idiosoma (Fig. 2.2). It is also known how indirect transfer of the sperm through the chelicerae of the male or the deposition of spermatophores on the substrate are subsequently recovered by the female through structures of the genital apparatus or epigynium. In various species, the female produces sexual pheromones (Krantz, 1978, 2009a).
2.2.4 Legs
The legs possess seven segments; coxa, trochanter, femur, genu, tibia, tarsus and pretarsus (Fig. 2.1). The coxae may be free or fused with podosoma ven- ter and the femur may be divided into basifemur and telofemur. The various segments present a precise number of setae, tactile or specialized (solenidia, eupathidia, famuli, microsetae, trichobothria), organized in whirls and set in
Introduction to Acari 17
dorsal, ventral and lateral positions. The distal end of each tarsus may bear the ambulatory appendages, consisting of a pair of lateral claws and a central empodium, with variations in different groups (Fig. 2.6). The pretarsus is also called ambulacrum when it possesses meolian elements (Krantz, 1978, 2009a).
2.3 CLASSIFICATION 2.3.1 Higher Classifi cation
The latest studies consider the Acari a subclass of the class Arachnida (Krantz, 1978; Johnston, 1982; Lindquist, 1984; Evans, 1992; Walter and Proctor, 1999;
Zhang, 2003; Lindquist et al., 2009). Recently, Zhang (2003) has partially mod- ifi ed the view of Johnston (1982) and Evans (1992) and has divided the sub- class into three superorders (Opilioacariformes, Parasitiformes and
A
B C
D
E
F
Fig. 2.5. Different shapes of aedeagus. (A) Petrobia tunisiae Manson; (B) Tetranychus urticae Koch; (C) Panonychus citri (McGregor) (from Vacante, 1985). Different shapes of receptaculum seminis; (D) Aplonobia histricina (Berlese); (E) Petrobia latens Müller (from Vacante, 1985); (F) Petrobia harti (Ewing) (from Meyer Smith, 1987).
18 Chapter 2
Acariformes) and seven orders. The superorder Opilioacariformes number the order Opilioacaridida, the superorder Parasitiformes, the orders Holothy- rida, Mesostigmata and Ixodida and the superorder Acariformes number the orders Prostigmata, Astigmata and Oribatida. Recently, Lindquist et al. (2009) recognized in the subclass only the superorders Parasitiformes and Acari- formes. The fi rst includes the order Opilioacaridida and the latter the orders Trombidiformes and Sacroptiformes. The species treated in this book belong to the suborder Prostigmata of the order Trombidiformes.
From a systematic point of view, the subclass Acari numbers currently 540 families, 124 superfamilies, 5500 genera and 1200 subgenera (Lindquist et al., 2009).
D
E
F A
B
C
Fig. 2.6. Different pretarsi. (A) Pretarsus I of female of Tyrophagus tropicus Robertson (from Vacante, 1989); (B) pretarsus I of female of Bryobia kissophila van Eyndhoven;
(C) pretarsus I of female of Panonychus citri (McGregor); (D) pretarsus I of female of Eotetranychus rubiphilus (Reck); (E) pretarsus I of female of Tetranychus urticae Koch (from Vacante, 1985, partially modifi ed); (F) empodium of Aculops pelekassi (Keifer) (from Keifer, 1959a, partially modifi ed).
Introduction to Acari 19
2.3.2 Suborder Prostigmata
The Prostigmata include about 17,170 described species, belonging to 1348 genera and 131 families (Walter and Proctor, 1999) characterized by different feeding habits (phytophagous, predaceous, mycophagous, etc.). The typical morphological character consists of the stigmata, set on the anterior margin of the prosoma or between the bases of chelicerae. Furthermore, the palp seg- ments may be blended or reduced. In different groups, the palptarsus may displace to the base of the tibia and form the ‘thumb claw complex’ (Fig. 2.4).
The digitus fi xus of the chelicerae may be missing or regressed and the digitus mobilis may be transformed into a stylet (Fig. 2.4). The cheliceral bases may form a stylophore or be fused to the subcapitulum (Fig. 2.4). The prosoma and opisthosoma often bear plates or dorsal shields (Zhang, 2003; Lindquist et al., 2009).
The species reported for citrus belong to the families Phytoptidae, Erio- phyidae, Diptilomiopidae, Tarsonemidae, Tenuipalpidae, Tuckerellidae and Tetranychidae.
© V. Vacante 2010. Citrus Mites: Identifi cation, Bionomy and Control
20 (V. Vacante)
3 Methods and Techniques
3.1 COLLECTING
In citrus crops, the Acari normally live on the plants, in the soil and/or in supports of a different nature and sometimes in all three habitats. The collect- ing method therefore varies depending on the behaviour of different species and the needs of the operator. Indeed, the needs of the control may be very different from those of a laboratory rearing or a systematic study. Regardless, identifi cation is desirable in order to collect the largest number of specimens and preferably in all biological stages, noting their colour, damage symp- toms and host plant, besides the date, location of collection and name of the collector. In this contribution, the techniques used for mites injurious to cul- tivated plants and citrus in particular are treated. However, several reasons may justify the examination of benefi cial mites and for this, information from Gerson et al. (2003) is applicable to the pest species.
3.1.1 Collecting from Plants
Various techniques allow the collection of immature and adult stages of Acari on plants (Jeppson et al., 1975; Upton, 1991; Amrine and Manson, 1996;
Perring et al., 1996; Walter and Krantz, 2009).
Many phytophagous or predaceous species living on the canopy can be collected by shaking the foliage with a stick or with hands on to a white or black support (tray, paper, etc.), depending on individual needs, and then using a needle or brush to collect them. Generally, the most common method involves keeping the manually collected plant material in sealed plastic or paper bags, storing it in the refrigerator at 10–15°C and examining it within a short time in the laboratory through a dissecting microscope. This avoids deterioration or death of specimens collected as a result of curing or predation.
Methods and Techniques 21
Working in this way, Acari are kept alive and can be used in the labora- tory for biological studies and various kinds of analyses (morphological, biochemical, etc.). The fi eld collection of Acari on the various organs of plants (leaves, branches, fruits, portions of bark) can also be carried out using pins mounted on sticks of various sizes, pipettes or fi ne hair brushes and using a hand lens, preserving them alive for a short time on empty vials or vials containing a preservative liquid such as 70% alcohol or Oudeman’s fl uid (see below). The method is not suitable for all systematic groups and is normally used for not very mobile species and those of middle or large size and still suffi ciently well known.
For highly mobile and medium- or large-sized Acari, an aspirator can be used, consisting of a glass vial, or one made from another transparent mate- rial, with a rubber bung and two tubes, one of which is used as a sucking tube, served behind a fi lter to avoid the Acari being sucked up by the oper- ator, and the other with a thin tip used for collecting the specimens from substrates.
The eriophyoids vagrant on leaves and fruits can be collected by pins and brushes or by pouring a syrup of sorbitol mixed with a 25% solution of isopropyl alcohol with a few iodine crystals on to the leaves and fruits, tak- ing care to collect everything in a small container and observe the material through a dissecting microscope. It is possible to wash the leaves and other parts of plants in hot water and separate the mites in the water by pouring the water through a sieve.
3.1.2 Collecting in Substrate
Mites on the soil surface can be collected with a hand-operated vacuum apparatus. The specimens collected on the fi lter can be observed directly with a hand lens or with a dissecting microscope, or you can pour the contents on a coloured mat, after washing with water or not. A Berlese–
Tullgren funnel can be used to collect mites from the ground vegetation and to extract living mites from soil and litter. A mass collection of mites may be conducted by placing attacked vegetation into a Berlese funnel. It can also be done by the canopy fogging method, involving the fumigation of vegetation or of part of the canopy with insecticides applied with fog (Krantz, 1978; McSorely and Walter, 1991; Upton, 1991; Walter and Krantz, 2009).
3.2 PRESERVING
The mites are usually kept in small vials or glass containers containing 70–80% alcohol with the addition of 5% glycerol to prevent alcohol evapora- tion (Evans et al., 1961). Oudeman’s fl uid (a mixture of 87 parts of 70%
alcohol, fi ve parts of glycerol and eight parts of glacial acetic acid) can be used as an alternative, which has the advantage in cases of long preservation
22 Chapter 3
of clarifying (in varying degrees according to the species) preserved speci- mens. Before conserving, it is necessary to immerse the mites in hot water, forcing them to extend their legs and other appendages, which facilitates their subsequent morphological examination; it is possible to fi x the mites in a solution of methanol and acetic acid in the quantity of two parts each and one part of distilled water, transferring them within a week to the preserva- tive medium (Evans, 1992; Saito and Osakabe, 1992).
3.3 PREPARING
3.3.1 Clearing/Maceration
The microscopic examination of the mites by optical phase contrast and interference system demands the specimens be cleared and mounted on slides. It is possible to use different media (Krantz, 1978; Walter and Krantz, 2009), including the very common lactophenol, produced by adding in sequence 50 parts lactid acid, 25 parts phenol crystal and 25 parts distilled water. To mitigate the aggressiveness of lactophenol on the most sensitive mites, a water solution of 50–95% lactic acid is used. In one way or another, depending on the size and nature of the species, it is possible to clear a spec- imen in a week at room temperature. Heating the mites in the clearing medium on a hot plate produces a rapid maceration within a period ranging from a few minutes to approximately 1 h. Cleared specimens must be washed with distilled water before mounting on slides.
3.3.2 Temporary Mounts
The mounting slide may be temporary or permanent and it is possible to consult various specialist contributions for this (Singer, 1967; Krantz, 1978;
Gutierrez, 1985a; Evans, 1992; Amrine and Manson, 1996; Walter and Krantz, 2009). Lactic acid can be used for temporary mounts, applying a drop at the centre of a glass slide and placing the mite at the centre with the help of an appropriately sized needle or spatula. It is possible to intervene with the same tools better to guide the specimen, paying attention to the position of the legs and avoiding them settling under the body. Subsequently, it is pos- sible to dispose above the mite a coverslip, of appropriate shape and size.
Again we must act carefully, ensuring the mite is at a suitable position; the outcome depends by the experience of the operator, the size of specimens and the slip covering the specimen. Some specialists use a concave glass slide, covering partially with the coverslip in order to guide the specimen in the study. This method is commonly applied in the study of particular groups (e.g. Oribatida).
Methods and Techniques 23
3.3.3 Permanent Mounts
Permanent mounts require the use of a slide, some drop of medium and a coverslip of different size and shape, depending on the various species and groups of mites.
Common media for permanent mounts are lactophenol media, resin- based media and Hoyer’s medium (Krantz, 1978; Saito et al., 1993; Upton, 1993; Amrine and Manson, 1996; Walter and Krantz, 2009). Among the lacto- phenol media, Heinze’s PVA is commonly employed, used as Hoyer’s medium and produced by mixing the following substances:
Polyvinyl alcohol 10 g
●
Distilled water 40–60 g
●
Lactic acid (85–92%) 35 ml
●
Phenol 1% aqueous solution 25 ml
●
Glycerol 10 ml
●
Chloral hydrate 100 g
●
Resin-based media include Canada Balsam and Euparal (and the opinion of acarologists on their use is divided), and these possess modest optical prop- erties, enabling them completely to macerate specimens and dehydrate those already treated.
Hoyer’s medium is widely used and basically derives from Berlese fl uid, based on the use of arabic gum and chloral hydrate, and can be used for clear, weakly sclerotized specimens; it also has good optical properties, even if it is not always considered a permanent medium. Since this is a toxic mixture, its use raises the need to take precautions, e.g. avoiding contact with skin and eyes. Moreover, it quickly degrades those specimens clarifi ed with lactic acid and not suffi ciently washed with distilled water. It is produced by mixing in sequence the following substances:
Distilled water 25 ml
●
Arabic gum 15 g
●
Chloral hydrate 100 g
●
Glycerine 10 ml
●
We must use arabic gum from the crystalline source, and not from powdered form, fi ltering the mixture several times.
The methodology of mounting the specimen in special positions was investigated in a number of cases, such as in tetranychid males, whose study requires that they be mounted sideways to see the male aedeagus on a lat- eral view. It is advisable to spread a small drop of Hoyer’s medium on the slide, placing the mite in the centre and laterally, and drying in a drying oven at 40°C for 3 h, or for a longer period at room temperature. Then, place a new drop of Hoyer’s medium on the specimen and on the layer of Hoyer’s medium that was semi-dry and the two layers should combine (Henderson, 2001).
24 Chapter 3
The permanent mounts require a drying process and hooping of the slides to identify the location of the mite in the slide. In particular, the use of water-soluble media demands that the specimen be fully dried in a hot oven at 40–50°C for 1–2 weeks. If the conservation environment of the dried slides does not possess a low humidity, the slides must be sealed around the cover- slip with a sealant. For this purpose, insulating paint is used (Glyceel, Euparal, Glyptal), ringing the coverslip with a small paintbrush and repeat- ing the coatings several times (Travis, 1968; Tribe, 1972; Fain, 1980).
3.4 REARING
Summarizing the information on the rearing methods of mites is not simple, since varying techniques according to the systematic groups and aims of study are employed. A comprehensive review appears in Krantz (1978) and, according to systematic groups, different contents can be used. For erio- phyoids refer to Oldfi eld and Perring (1996), for tarsonemids to Liang (1980) and Xu et al. (1994) and for tetranichids to Helle and Overmeer (1985a).
© V. Vacante 2010. Citrus Mites: Identifi cation, Bionomy and Control
(V. Vacante) 25
4 Plant Damage
Many morphological, physiological and behavioural adaptations for feeding on plants favour the Trombidiform mites. Thus, over 4000 species of Trombidi- formes belonging to about 300 genera are recorded for their obligate phytophagy (Amrine and Stasny, 1994; Bolland et al., 1998), including some important pests of economic plants. Mite pests can cause different degrees of serious damage to citrus trees and fruits according to various environmental factors. Thus, whereas LeClerg (1965) recorded a 2.5% loss in citrus fruit production related to damage by pest mites alone in the USA, more than 30% of production is damaged every year in India by insect and mite pests (Pruthi and Mani, 1945; Butani, 1979a, b).
Knowledge of feeding mechanisms and feeding symptoms helps in under- standing the nature and type of damage, to recognize the pest responsible and place the control in a context of rationality. Feeding symptoms are correlated to morphological and functional adaptations and the bio-ecology of the injurious species, aside from the attacked organ, the host plant and its phenology. The feeding activity of mites causes local damage and general alterations. In order to aid understanding of the symptoms and identifi cation of various species, a number of websites have been listed, which contain photographs of the most important species and their damage on citrus (Table 4.1).
4.1 FEEDING MECHANISMS
In the Tetranychoidea (Tetranychidae, Tenuipalpidae, Tuckerellidae) the sty- lophore, the chelicerae and the palps are involved in the feeding mechanisms (Fig. 2.4). The stylophore is mobile (retractable and extrudable) and lies on the dorso-medial surface of the infracapitulum; it consists of the bases of chelicerae consolidated between them, curves ventrally and is divided by a median and vertical septum and penetrated in two basal chelicerae, each of which hosts a stylet, attach