Around the world, efforts are underway to find the petroleum replacement sources to overcome the depletion of fossil fuels and to reduce greenhouse gas emissions. In this respect, the replacement of petrochemicals with renewable alternatives is one of the main challenges of our day. Hexanoic acid can be widely used as a precursor for biofuel and for the production of chain-based chemicals such as adipic acid, hexane, amino-caproic acid and so on.
To produce specific carbon chain fatty acids, the β-oxidation pathway of Escherichia coli was developed. In this study, a maximum of 210 mg/L hexanoic acid was produced by the engineered strain (MGREBA2) from 1 g/L oleic acid in 48 hours of biotransformation. Based on these results, it is suggested that various oils as raw materials will be directly applied for the production of short-chain fatty acids, especially hexanoic acid.
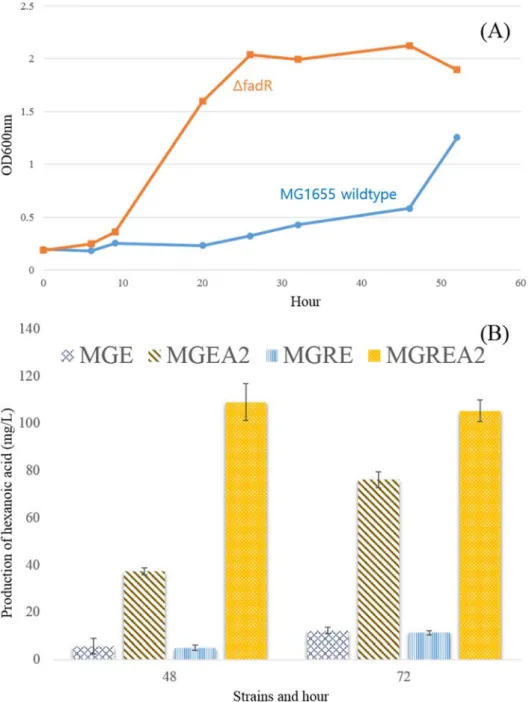
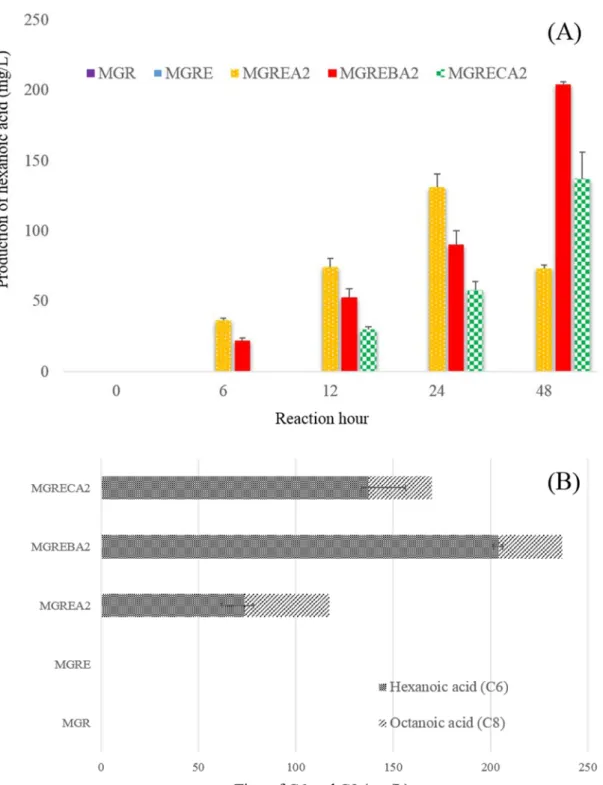
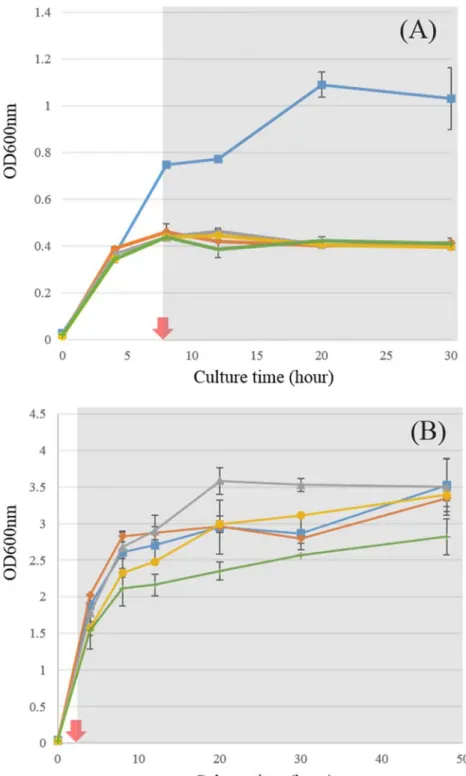
Growth curve between wild-type strain and fadR knockout strain (A), biotransformation to hexanoic acid from oleic acid in LB broth (B). Hexanoic acid titer of supernatant and culture medium (A), production of hexanoic acid by biotransformation in LB broth (B).
Introduction
- Hexanoic acid
- Fatty acids metabolism in E. coli
- Beta-oxidation pathway
- Thioesterase specificity in E. coli
- Modification of beta-oxidation in E. coli
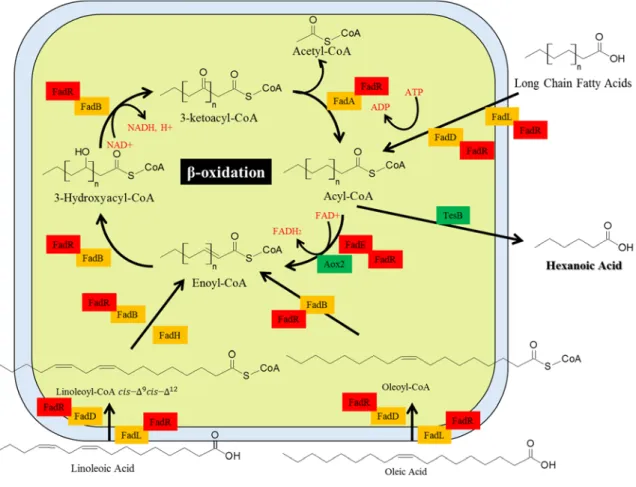
The initial phase of β-oxidation is mediated by acyl-CoA synthetase (FadD) encoded by the fadD gene. And then this acyl-CoA is converted to enoyl-CoA, which oxidation is mediated by acyl-CoA dehydrogenase (FadE) encoded by the fadE gene. Therefore, β-oxidation engineering is an important part of the production of specific chain fatty acids if the acyl-CoA dehydrogenase substrate specificity is modulated16.
The next step is conversion of enoyl-CoA to 3-ketoacyl-CoA via 3-hydroxyacyl-CoA mediated by FadB. Finally, 3-keto acyl-CoA is converted to acyl-CoA by cutting out acetyl-CoA and replacing new coenzyme A mediated by FadA (Fig. 1) 17. FAD can be reduced to FADH₂ mediated by acyl-CoA dehydrogenase and acyl-CoA oxidase in the mitochondria and the peroxisome, respectively.
Acyl-CoA oxidases are directly on oxygen to generate hydrogen peroxide in order to convert FAD from FADH (Fig. 2) 18b. The yeast saccharomyces cerevisiae (S. cerevisiae) has the only acyl-CoA oxidase such as POX1, which has substrate specificity for short- to long-chain substrate specificity. FADH2, which is mediated by acyl-CoA dehydrogenase with conversion to enoyl-CoA form, is converted to FAD by complexⅡ in the electron transport chain, acyl-CoA oxidases are directly in oxygen to generate hydrogen peroxide in order to convert to FAD from FADH.
ThioesteraseⅠ, encoded by the tesA gene, cleaves fatty acyl-acyl carrier protein (ACP), whereas ThioesteraseⅡ(TesB), encoded by the tesB gene, has specificity for the acyl-CoA form23. ThioesteraseIII, which was identified relatively recently, was found to have the substrate specificity for long-chain acyl-CoA-derived unsaturated fatty acid 24 . Among these three thioesterases, ThioesteraseⅡ has the broadest substrate specificity of acyl-CoA chain length.
ThioesteraseⅠ is specific for the chain length of C12 to 18 acyl-CoA ester, while thioesteraseⅡ has specificity for the chain length of C6 to C18 as the broad chain length23. Therefore, in this study, we chose TesB to produce hexanoic acid because it has C6-acyl-CoA specificity.
Materials and methods
- E. coli strains and growth condition
- Construction of plasmids and strains
- Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) analysis
- Growth study
- Biotransformation in LB broth
- Whole cell biotransformation in M9 broth
- Analysis of biotransformation products
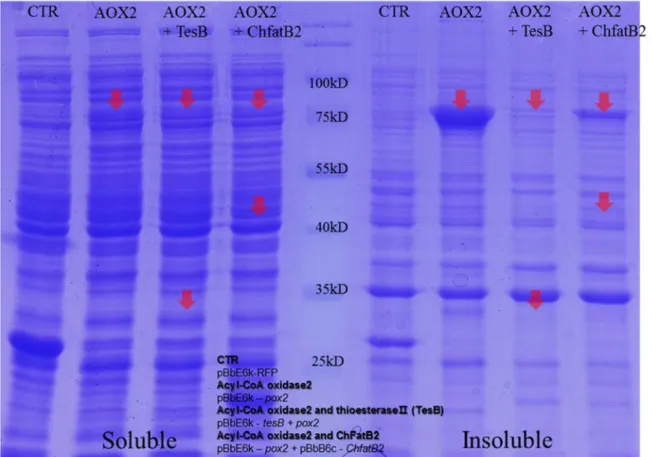
20111 pBbE6k-Aox2 pBbE6k with the POX2 gene derived Y.Lipolytica This study pBbE6k-TesB/Aox2 pBbE6k with the tesB gene with POX2 gene This study pBbB6c-ChfatB2 pBbB6c with the fatB2 gene derived Cuphea hookeriana This study Table 1. Overnight culture medium as LB broth at 37 ℃ was inoculated into 50 ml fresh LB broth at 1/100 diluted ratio with appropriate antibiotics. And then the harvested cell was resuspended in lysis buffer (50 Mm Tris-HCl pH 8.0) and sonicated using Vitra Cell Sonicator (SONICS, USA).
After separation of supernatant and pellet by centrifugation at 13,000 rpm, and mixed with loading buffer. Overnight culture medium as LB broth at 37°C was separately inoculated into 50 ml of fresh LB broth and M9 medium containing 0.1% oleic acid, 0.5% TWEEN80 and 100 mM potassium phosphate buffer in a 1/100 diluted ratio with appropriate antibiotics. The absorbance for cell growth was monitored with Libra S22 Spectrophotometer (Biochrom, UK) at 600 nm.
After preparation of the initial culture, each engineered strain and control cell were inoculated into 50 ml of fresh LB broth in 250 ml flask. One milliliter samples were collected at different time points and 72 hours after oleic acid injection) for GC/FID analysis. When an OD600 of 0.7 was approached, 0.1 mM IPTG was added for induction of protein expression.
After washing steps, these cells were injected into the transformation medium, which is M9 medium containing 0.1% oleic acid, 0.5% TWEEN80, 100 mM potassium phosphate buffer, and the appropriate antibiotic as 50 μg/mL kanamycin or 30 μg/mL mL of chloramphenicol. The collected solution was vortexed for 10 seconds and 500 μL was transferred to a labeled glass gas chromatography (GC) vial.
Results
- Expression of acyl-CoA oxidase2 and thioesterases
- Growth of the engineered strains
- Comparison between fadE knockout and fadR/fadE double knockout strain
- Biotransformation to hexanoic acid
Except for strain MGR as a control, all strains showed lack of growth in M9 medium with oleic acid (Figure 4A). For the formation of hexanoic acid from long-chain fatty acids, biotransformation on LB slurry with 0.1%. Prior to biotransformation, because growth on LB medium could affect product composition such as hexanoic acid, pure LB was used for 48 h cultivation for all strains in this study without the addition of a carbon source.
The results showed that MGREA2 produced more hexanoic acid than MGEA2 after 48 h of reaction (Fig. 5B). MGREA2 was converted to approx. 110 mg/L hexanoic acid from 1 g/L oleic acid, whereas MGEBA2 showed approx. 80 mg/L hexanoic acid converted during 72 hours. Before biotransformation, the hexanoic acid titer between supernatant and whole medium was checked to measure product secretion out of the cell.
This experiment was performed via whole-cell biotransformation using the MGREA2 strain on M9 medium with 0.1% oleic acid. To measure the secretion of hexanoic acid to the medium from the cell, an identical sample, which reacted by biotransformation for 48 h, was prepared and then divided into supernatant and complete culture medium. MGREA2-containing sole of Aox2 showed a production of approximately 75 mg/L hexanoic acid (Fig. 7A), and also an octanoic acid titer of approximately 45 mg/L (Fig. 7B).
MGRECA2 produced approximately 140 mg/l hexanoic acid and also formed approximately 30 mg/l octanoic acid. In the case of MGREBA2, the hexanoic acid was produced to the highest titer (Fig. 7A). The MGREBA2 sample as the highest titer was measured for hexanoic acid production for unit time.
As a result, the production of hexanoic acid was gradually increased for 48 h (Figure 7A). Finally, over 200 mg/L of hexanoic acid with a fraction of octanoic acid was produced after 48 h reaction in E. Under biotransformation conditions in LB, the supernatant and the whole culture medium were measured by the hexanoic acid titer using the strain MGEA2 (A ).
The titer of hexanoic acid from biotransformation in M9 with the sole carbon source of 0.1% oleic acid was measured per unit time.
Discussion
- Effect of hexanoic acid production for growth
- Effect of fadR knockout on hexanoic acid production
- Effect of thioesterase Ⅱ on hexanoic acid production
- Future research plan
By biotransformation through LB broth, the titer of hexanoic acid was measured between MGREA2 and MGEA2 strain. Accordingly, fadR deleted MGREA2 strain showed higher titer of hexanoic acid (Fig. 5B), and this is considered to be caused by major repressor removal of β-oxidation. Therefore, a greater amount of hexanoic acid was produced by enzyme reaction inside cell because fadR knockout strain could take up more oleic acid than non-fadR deleted strain32.
To find a suitable thioesterase for hexanoic acid production, TesB and ChfatB2 were separately expressed with Aox2, and then the titer of hexanoic acid production was measured by GC/FID. As a result, the best hexanoic acid strain was MGREBA2, which contained Aox2 with TesB (Figs. 6, 7), it is believed that the production of hexanoic acid is related to the substrate specificity of these enzymes. For this reason, it is assumed that the input oleic acid in the β-oxidation cycle is shortened to hexanoyl-CoA after 6 cycles, and then this limited production of Aox2 is converted to hexanoic acid by overexpressed TesB inside the cell.
The MGREBA2 strain showed increasing hexanoic acid production titer over time ( Fig. 6B ). Furthermore, MGREA2 strain showed decreased hexanoic acid titer after 48 h in LB score (Fig 6B), M9 score was also observed to decrease hexanoic acid titer after 24 h (Fig 7A). In this case, it is assumed that hexanoic acid is shortened to butanoic acid and acetyl-CoA to β-oxidation without overexpressed TesB, because Aox2 has very little substrate specificity for C6.
Therefore, in the condition of C6-acyl-CoA presence inside the cell, this hexanoyl-CoA could react with TesB than Aox2, this assumption is supported by the results of MGRCA2 strain (Fig. 7A). MGRCA2 containing ChfatB2, which has substrate specificity for C8 to C10, is slowly produced to hexanoic acid as a result on M9. 21. oleic acid in β-oxidation could be catalyzed by ChfatB2 to octanoic acid, it is also reacted by Aox2 inside the cell.
It is assumed that the cause of slow production in MGRCA2 strain occurred at conversion from C8 to C6 in β-oxidation because ChfatB2 competes with Aox2 at that moment. In this study, all fadE knockout strains showed growth deficiencies on the sole carbon source of oleic acid.
Conclusions
Cho, H.; Cronan, J., Escherichia coli thioesterase I, molecular cloning and sequencing of the structural gene and identification as a periplasmic enzyme. J.; Steinbuchel, A., Fatty acid synthesis in Escherichia coli and its applications for the production of fatty acid-based biofuels. a) Francoise Le, B.; Jean, D., Interaction between peroxisomes and mitochondria in fatty acid metabolism. Wang, H.; Le Dall, M.-T.; Waché, Y.; Laroche, C.; Belin, J.-M.; Nicaud, J.-M., Cloning, sequencing and characterization of five genes encoding acyl-CoA oxidase isozymes in the yeast Yarrowia lipolytica.
Zheng, Z.; Gong, Q.; Liu, T.; Deng, Y.; Chen, J.-C.; Chen, G.-Q., Thioesterase II of Escherichia coli plays an important role in the production of 3-hydroxydecanoic acid. Nie, L.; Ren, Y.; Schulz, H., Identification and Characterization of Escherichia coli Thioesterase III Functioning in Fatty Acid β-Oxidation†.