This paper is submitted in partial fulfillment of the requirements for the title Master of Medical Sciences (Biochemistry and Molecular Biology). I would also like to thank all the members of the Department of Biochemistry and Molecular Biology at UAE University who have helped me during my studies and research. My special thanks go to the College of Medicine and Health Sciences and the United Arab Emirates University for providing us with grants to conduct our study.
Introduction
Diabetes Mellitus
Epidemiology of DM
However, in type 2 diabetes, there is no specific diagnostic test to identify it, apart from the exclusion of the other causes. The classification of patients with diabetes mellitus into subgroups based on disease progression and risk of complications is being investigated (Ahlqvist et al., 2018; Pearson, 2019).
DM Classification and Diagnosis
- Type 1 Diabetes Mellitus
- Type 2 Diabetes Mellitus (T2DM)
- Islet Morphology
- Characteristics of Human Β- Cells in T1DM and T2DM
However, IA-2 autoantibodies or ZNT8 autoantibodies are rarely detected first (Thomaidou et al., 2018; DiMeglio et al., 2018). In a study performed on T1DM mice, clearance of senescent β-cells reduced the incidence of diabetes (Thompson et al., 2019). In another study of human T1DM, loss of the β-cell markers insulin, proinsulin, and amylin preceded β-cell death (Damond et al., 2020).
Management of Diabetes Mellitus
- Lifestyle Modification
- Pharmacotherapy
Metformin can reduce weight and HbA1c when combined with lifestyle interventions (Meneghini et al., 2011; Domecq et al., 2015). It also promotes pancreatic beta cell function unrelated to insulin resistance (Lyu et al., 2017). Exenatide is injected twice daily one hour before meals and liraglutide once daily (Diego et al., 2006).
Peptides as Therapeutic Agents
Furthermore, several clinical studies indicate that the use of GLP-1 receptor agonists lowers systolic blood pressure and exerts cardiovascular protective effects (Fonseca et al., 2014; Russell-Jones et al., 2009; Marre et al., 2009). Exenatide and liraglutide are the two pharmaceutical forms of GLP-1 receptor agonists available on the market. However, recent changes in drug delivery systems, new formulations and other areas involved have made it possible to overcome these disadvantages and make use of their advantages such as high potency and selectivity of their targets as well as the low toxicity, low accumulation in tissues and rapid clearance, which seen in several natural peptide drugs such as insulin (Anderson & Delgado, 2008; Barabási et al., 2011).
Lipocalin-2
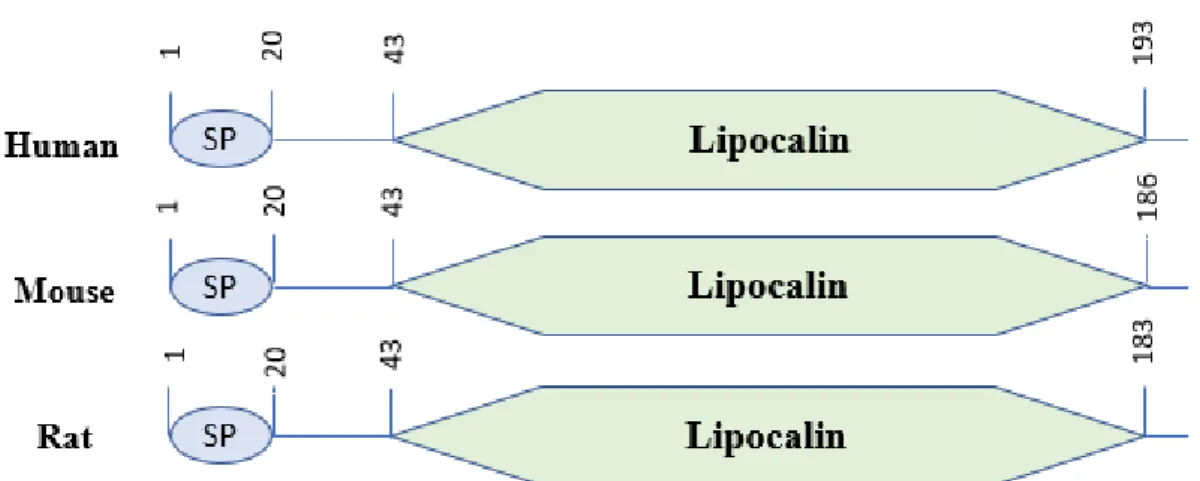
- Structure
- Tissue Distribution of Lipocalin 2
- Inductors of Lipocalin-2 Expression
- Inhibitor of Lipocalin-2 Expression
- Controversial Factors in LCN2 Expression
- Lipocalin-2, Obesity and Insulin Resistance
- LCN2 Gene and Diabetes
LCN2 occurs in different molecular forms as a monomer (25-kDa), a disulfide-linked homodimer (46-kDa), and a disulfide-linked heterodimer with human neutrophil gelatinase B (135-kDa) (Kjeldsen et al., 1993). In the embryo, LCN2 is expressed in hypertrophic and perihypertrophic regions of developing cartilage (Owen et al., 2008). A study by Yan et al., showed that white adipose tissue (WAT) was the main source of LCN2 expression and reported its absence in brown adipose tissue (BAT) in male wild-type mice.
Transplantation studies in humans and mice show that LCN2 expression originates from the damaged nephrons (Ashraf et al., 2016). In addition, chronic kidney disease (CKD) is also associated with a marked increase in LCN2 levels in tissues and body fluids (blood and urine) (Stenvinkel, 2010; Viau et al., 2010). Whereas once the lesions had healed, LCN2 expression naturally decreased, suggesting that LCN2 expression is regulated by the disease process (Mallbris et al., 2002).
In addition, hypoxia induced LCN2 mRNA expression and protein levels in HL-1 cells (Xiong et al., 2016). Another study examining reactive astrocytosis also confirmed that LPS treatment (100 ng/ml) markedly increased LCN2 expression (Lee et al., 2009). Another study also revealed a significant increase in LCN2 expression and secretion in adipose tissue of obese (ob/ob) mice compared to lean mice (Yan et al., 2007).
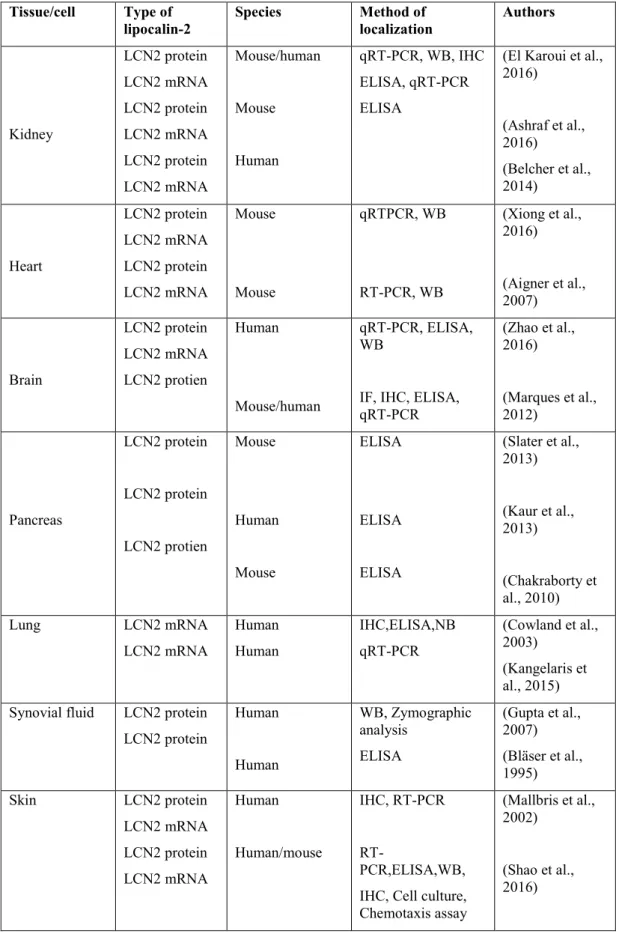
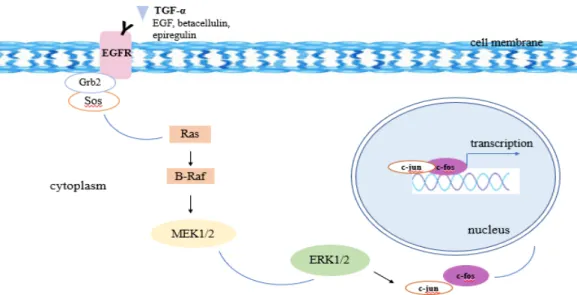
The activation of C/EBPβ is responsible for the transcriptional activation of LCN2 expression induced by AGE (Chung et al., 2013). Insulin can also stimulate LCN2 expression through PI3K and MAPK signaling pathways (Tan et al., 2009). Increased expression of LCN2 was detected in hepatocytes, biliary epithelial cells and proximal tubular epithelial cells of the kidney (Adler et al., 2010).
Aims, Hypothesis and Objectives
Hypothesis
Aim
Objectives
Materials and Methods
- Animal Model
- Establishing the Experimental Diabetes Model
- Experimental Design
- Blood/Tissue Collection and Tissue Processing
- Immunofluorescence Staining of Paraffin Sections
- Immunoelectron Microscopy
- Markers of Oxidative Stress
- Biochemical and Peptide Hormone Analysis
- Biochemical Analysis Using Cobas®
- Peptide Hormone Analysis Using Magpix®
- Cell culture, Treatment, and Insulin Secretion
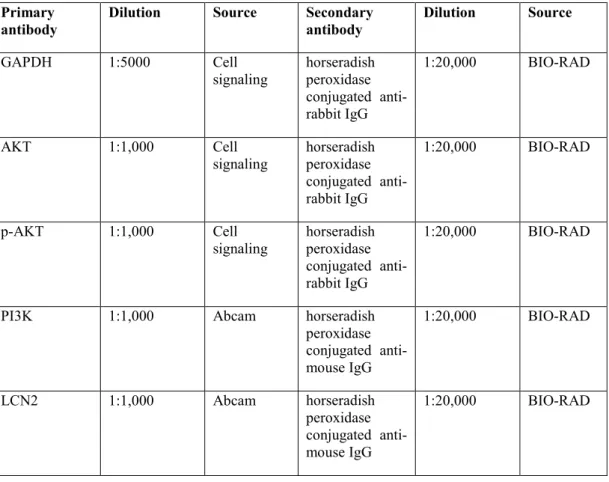
- SDS-PAGE and Western Blot Analysis
- Statistical Analysis
Tissues were then incubated on a rotator for one hour at room temperature in a ratio of 1:1 LR White resin and 95% ethanol. The next morning the samples were incubated with 100% fresh LR White for two hours at room temperature and the samples were embedded in pure resin in gelatin capsules. For polymerization, gelatin capsules were placed in a container under UV lamp (360-365 nm) in a room for 24-36 hours at room temperature.
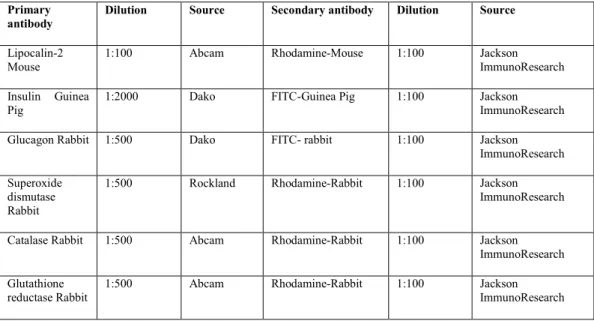
Blocking buffer was then added to the tissue sections for 45 min at room temperature to avoid any nonspecific binding. The next day, the slides were washed with PBS 3 times for 5 min each, and then the secondary antibody was added and incubated for 1 h at room temperature (FITC-guinea pig, rhodamine-rabbit/mouse, and FITC-rabbit). Grids containing pancreatic sections were washed with deionized water and then the grids were incubated with 10% H2O2 in water for 10 min.
Then the grids were washed with wash buffer for 5 minutes, then the tissues were blocked with wash buffer containing 20% normal goat serum (NGS), then the primary antibody (mouse anti-lipocalin-2) was added and incubated overnight at 4°C . The next day, grids were kept at room temperature for one hour, washed 3 times with PBS and incubated with blocking buffer for 20 minutes at room temperature. Then, secondary antibodies (goat anti-mouse IgG coated with 10 nm gold particles) were added and incubated for 2 hours at room temperature.
Grids were washed again with PBS for 3 times, and the above steps were repeated for the same grids using a different primary antibody (rabbit anti-insulin) and incubated at room temperature for 2 h.
Results
- Metabolic Parameters
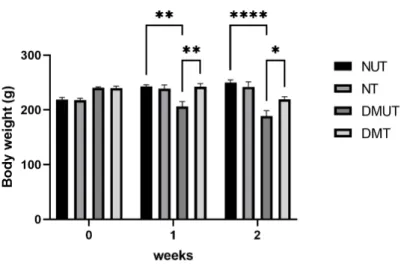
- Body Weight
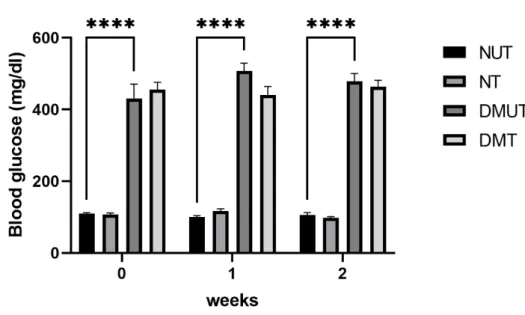
- Glucose Level
- IPGTT
- Insulin Release
- Immunohistochemical Localization of LCN2
- Co-localization Analysis of LCN2 with Insulin in Pancreas
- Co-localization Analysis of LCN2 with Glucagon in Pancreas
- Electron Microscopy
- Markers of Oxidative Stress
- Effects of LCN2 on Biochemical Parameters
- Liver Function Test
- Kidney Function Test
- Lipid Profile
- Peptide Hormones Analysis
- Insulin Signaling Pathway
This study demonstrates that LCN2 is co-localized with insulin in the pancreatic islets of Langerhans as shown in Figure 10. However, a significant increase in the antioxidant enzymes SOD and GSH reductase was found in serum of the LCN2-treated STZ-induced diabetic animals compared to the diabetes control. Note the significant elevation of AST and ALT levels in diabetic animals and the significant effect of LCN2 in lowering both enzyme levels in diabetic animals.
On the other hand, there is no noticeable difference in creatinine levels in all test animals, with or without the treatment. In animals with diabetes, a significant reduction in insulin levels was observed compared to the normal control group. LCN2 caused a significant increase in insulin levels in diabetic animals, while it had no effect in non-diabetic animals.
LCN2 also caused a significant increase in the levels of Amylin in diabetic treated groups compared to diabetic control animals. LCN2 effects on leptin levels have been shown in Figure (25), where the induction of DM caused marked reduction in leptin levels, but a small improvement in leptin levels with LCN2 treatment was detected in diabetic animals. DM induction caused a significant increase in the levels of IL-6 and MCP1, and the treatment with LCN2 resulted in a significant decrease in both the levels of IL-6 and MCP1.
Note that LCN2 caused a significant increase in the levels of Amylin and insulin in the diabetes-treated group compared to the diabetes-control group.
Discussion
- Metabolic Parameters
- Body Weight
- Glucose Levels
- Intraperitoneal Glucose Tolerance Test (IPGTT)
- Effects of LCN2 on Insulin Secretion In Vitro
- Immunohistochemical Localization of LCN2
- Distribution of Lipocalin-2 (LCN2) in Cytoplasmic Organelles
- Effects of LCN2 on Antioxidant Enzymes Activities in Serum
- Effect of LCN2 on Biochemical Parameters
- Liver and Kidney Function
- Lipid Profile
- Effects of LCN2 Treatment on Peptide Hormones of Normal
- Effects of LCN2 Treatment on Inflammatory Markers of Normal
- Insulin Signaling Pathway
Plasma glucose levels before glucose administration were markedly higher in the diabetic group compared to control groups. The number of islet cells containing LCN2 was significantly lower in the pancreatic islet of diabetic rats compared with normal rats. There was no evidence of co-localization of LCN2 with glucagon in pancreatic islet alpha cells in the normal rats.
Several sizes of IgG-conjugated gold particles were evaluated to examine the degree of co-localization of LCN2 and insulin in cytoplasmic organelles of beta cells. However, a significant increase in the antioxidant enzymes SOD and GSH-reductase was found in the serum and pancreatic tissue of STZ-induced LCN2-treated diabetic animals compared to the diabetic control, indicating enhanced ROS scavenging ability. Induction of DM caused a significant increase in serum urea and urea nitrogen levels.
Amylin is co-produced with insulin in the pancreatic β-cells, the effects of LCN2 on amylin had the same tendency as its effect on insulin. As a result, the hypoglycemic effects of LCN2 may be exerted by improving leptin levels as the data showed in the diabetic and normal rats. As a result, the possible anti-inflammatory role of LCN2 was investigated in the diabetic model used.
LCN2 treatment had a marked effect on IL-6 where it caused a significant reduction in serum IL-6 levels.
Conclusion
Limitations
Advancement in the Field
Progressive loss of β-cell function leads to impaired glucose tolerance in first-degree relatives of subjects with type 2 diabetes. Role of α-cell dysregulation in fasting and postprandial hyperglycemia in type 2 diabetes and therapeutic implications. . Human pancreatic islet transcriptome: Expression of candidate genes for type 1 diabetes and influence of pro-inflammatory cytokines.
Systolic blood pressure reductions with liraglutide in patients with type 2 diabetes: Insights from a patient-level pooled analysis of six randomized clinical trials. Predictors and longitudinal changes in insulin sensitivity and secretion preceding the onset of type 2 diabetes. Effects of dipeptidyl peptidase-4 inhibitors on beta-cell function and insulin resistance in type 2 diabetes: Meta-analysis of controlled trials of random.
Different gene expression pathways in islets from individuals with short- and long-term type 1 diabetes. Fifty years of pancreatic island pathology in human type 1 diabetes: insights gained and progress made. Diagnosis of type 2 diabetes mellitus and associations with cardiovascular and mortality risks: findings from the Swedish National Diabetes Registry.
The natural history of insulin secretory dysfunction and insulin resistance in the pathogenesis of type 2 diabetes mellitus.