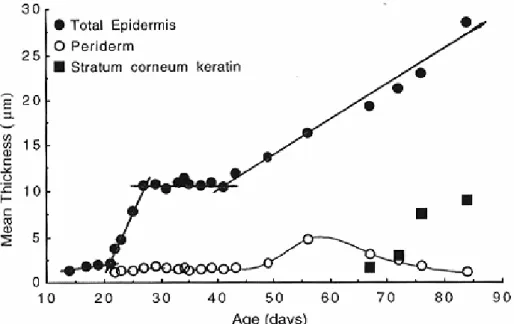
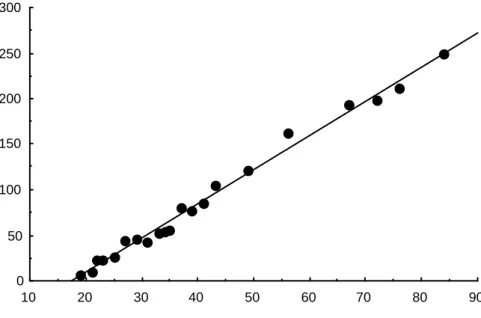
Mean thickness of the periderm, the keratin of the stratum corneum and the total epidermis in the skin of Crocodylus porosus embryos, from 14 days to 1 day after hatching. Stylized representation of the four collagen patterns identified in the skin of juvenile/subadult Crocodylus porosus. The growth of the skin as a whole (up to about 250 µm at hatching) is linear.
The skin continues to grow linearly after hatching, with generalized thickening of the epidermis and dermis. Certainly, skin buyers and tanners were largely ignorant of the morphological characteristics of crocodile skins. Crocodiles were caught with a harpoon (see Walsh 1987), pulled alongside a boat and shot into the back of the skull platform.
Variation in skin pattern
Attempts were initially made to take skin samples from immobilized crocodiles (using “Flaxedil”), but this proved inappropriate due to the nervous reaction of the animals when the skin was cut. All crocodiles were shot into the back of the skull platform at close range using a 0.22 inch (short caliber) rifle. Skin samples (3-5 mm x 1-2 mm x 3-4 mm) were taken from different parts of the body of farm and wild animals of different sizes (e.g. chin, midabdomen, pelvic abdomen, thoracic abdomen, caudal abdomen, ventral tail , dorsolateral tail, flank, occiput, nuchal translucency, posterior nuclei, crosswise, dorsal pulse, ventral pulse).
Because the value of crocodile skins is significantly reduced if the belly area has even the tiniest hole, sampling was limited to areas away from the valuable part of the skin.
Level of heritability and effect of incubation temperature on skin pattern
Even with the use of local anesthetic along with the immobilizing agent, specimens were generally of little value for histological purposes. In crocodiles, the nerve response remains after death (a typical reptilian trait due to their anaerobic metabolism). The spinal cord was then transected and the animal was left to bleed before being treated.
Adults were placed in 20 identical pens together, allowing offspring to be assigned and compared to known parents.
Industry perceptions
Results
Embryological development of skin
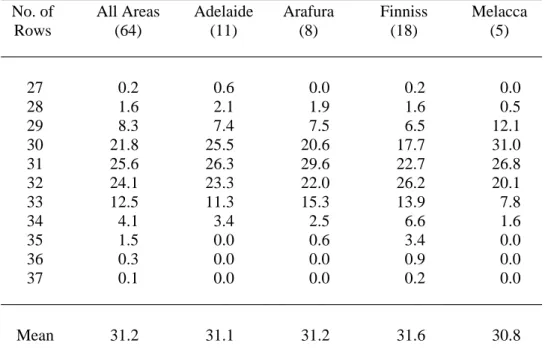
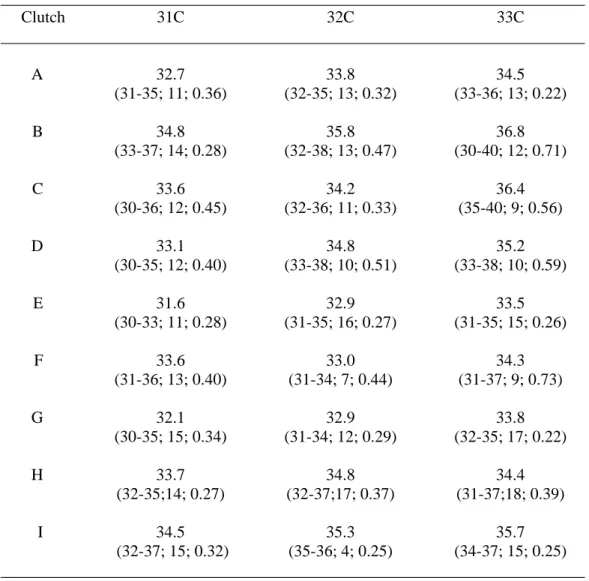
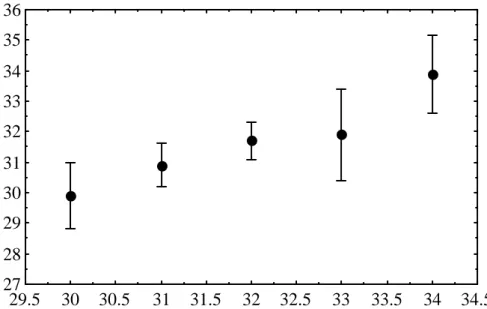
Distribution (%) of the number of scale rows for Crocodylus porosus juveniles from 64 flocks, in areas from which 5 or more claws were represented. The difference between the highest and lowest degree row is calculated within 85 clutches of wild Crocodylus porosus eggs. Relationship between mean number of scale rows and constant incubation temperature for hatchlings from 8 clutches of Crocodylus porosus eggs.
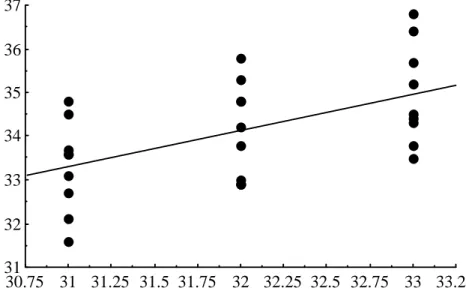
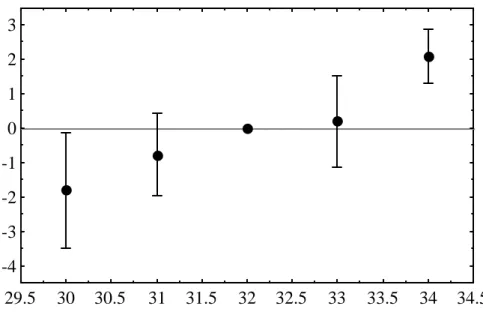
Middle scale bar counts 9 clutches of Crocodylus porosus eggs incubated at constant 31, 32 and 33C. The numbers in parentheses are the range, sample size, and standard error, respectively. However, since the mean number of scale rows varies among flocks incubated at the same temperature, this regression relationship can only be applied to data for those particular flocks. To overcome significant herd effects, 32C was chosen as the standard incubation temperature and the mean number of scale rows at this temperature was treated as zero.
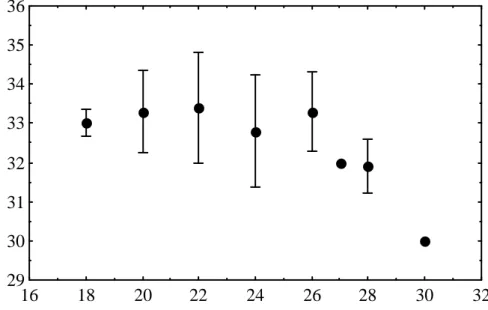
Mean number of scale bars for clutches incubated at 32 C for 13 days, 'pulsed' at different temperatures for the equivalent of 20 days of development at 30 C and then returned to 32 C to complete incubation. Thus, the stage at which high incubation temperatures affect the number of scale types occurs between 26 and 30 days. In two breeding seasons (1997/98 and 1998/99), we checked the contribution of the parents to the number of lines on the scale of the offspring; there was not enough data for the 1996/97 season.
Average number of rows of hatchlings in relation to their father's, in the nesting season 1997/98. Average number of rows of hatchlings in relation to their father's, in the nesting season 1998/99. Data on offspring of known males (and females) over three nesting seasons (Table 4) indicate the degree of variation in mean scale row counts from year to year at 32C incubation.
Average number of rows on the progeny scale of adult males of the species Crocodylus porosus for which two or more broods were produced in three breeding seasons (1996/97 to 1998/99).
Farmed crocodiles
As previously described for hatchlings, the keratin of the interscalar region is composed of alpha-keratin, while the scale itself is of harder beta-keratin (Figure 12). The superficial dermis of the lateral and dorsal areas of the skin contained many melanophores and free melanin granules (Figure 13). Note the different staining of the scalar (transparent) and interscalar (dark) keratin, the extensive pigment of the loose.
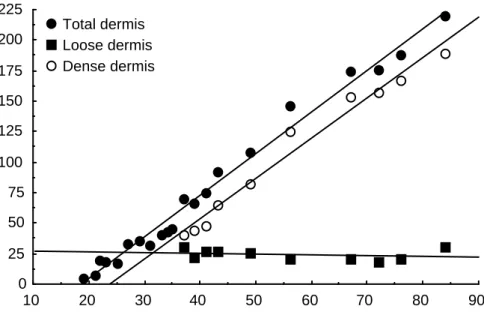
In both brooding and large crocodiles there is an extensive arborizing network immediately below the epidermis, in the same zone as the melanophores. Note the large dark-staining melanophores and the diffuse light brown appearance of the extensive iridocyte processes in the loose dermis. It was the arrangement of the dense dermal collagen that varied between sites, and the dense dermis could be divided into four 'architectural' patterns (shown diagrammatically in Figure 14).
The first (pattern I) is seen in the dermis of the wrist, ankle and dorsolateral tail skin. In the vertical plane, primary beams run in the direction of the axis. The row width is greatest in the deep layers and smallest in the superficial layers.
The arrangement of collagen fibers is clearer in the ventral skin samples than in the dorsal skin samples. There is an irregular arrangement of collagen bundles, which are both thick and thin, and have no specific orientation on the skin surface.
Wild versus farmed crocodiles
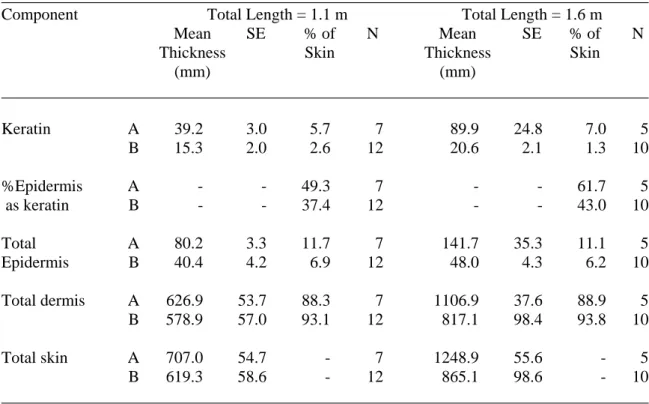
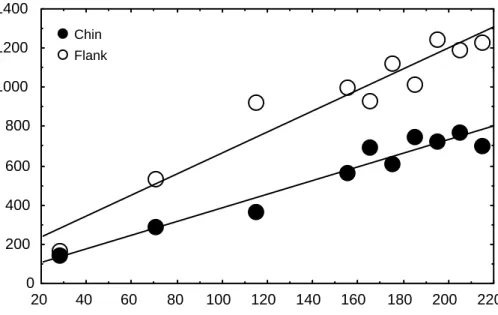
Although some thick bundles are present, the collagen fibers do not form thick rows as in sample I. For example, skin from the chin of wild crocodiles was 33% thicker than skin from farmed animals. This difference was due to an increase in the thickness of the dermis (41%) and a decrease in the thickness of the epidermis (including keratin; -35%).
The keratin layer in the epidermis of wild crocodile skin was also slightly thinner (9%) than in cultured specimens. In farmed crocodiles with a total length of less than 1.5 m, the dermis comprised about 74.8% of the thickness of the skin, which is lower than in larger crocodiles. The average proportion of keratin in the epidermis was similar (about 32.5%) in all sizes of farmed crocodiles.
Thus, the thickness of keratin (31.7 µm), epidermis (including keratin; 96.2 µm), and dermis (902 µm), although variable in individuals taller than 1.5 m, did not change significantly with ventral wrist skin size . The wrist skin of wild crocodiles also had a thinner epidermis (48% and 49% thinner for the dorsal and ventral wrist, respectively). In this case, however, the dermis of wild crocodiles was less thick than that of farmed crocodiles, resulting in an overall thinner skin [this was more pronounced in the dorsal pulse (21%) than in the ventral pulse (2%)].
Industry perception
This greatly affects the visual appearance of the abdominal skin and limits where a skin can be used. Double scaling appears to reflect changes in shell growth such that the inner portion becomes higher and defined from the lower portion of the shell. Some product manufacturers have indicated that skins with more scale rows are more desirable, but the limited number of them currently on the market does not justify a premium price.
For example, a row of scales in the center of the skin may divide into two rows as it progresses to the flanks. Manufacturers concerned with products of the highest quality (and value) agreed that if a regular source of hides with a high number of scale rows and a regular scale pattern were available, such hides could fetch a higher price. Wild hides were considered higher quality by some hide buyers because of their "texture."
A large tannery in Singapore, with a very high turnover of skins, did not observe a difference between wild and farmed skins in terms of skin thickness, but could tell by the 'feel' of the skin whether it was farmed or wild is. On the other hand, an Australian and Japanese buyer perceived wild skins as thicker than farm skins. The ability to distinguish between wild and farm hides (by whatever means), and to identify the origin of hides (farm or locality), was linked to experience in the industry.
A number of variables, including habitat type for wild pelts and farming practices for farmed pelts were identified as important factors, although reasons for differences were not known. For example, in Japan, a trading house will buy raw hides from a farmer, and sell to the tanner, who will sell to the product manufacturer, who will sell to a retailer.
Discussion
Crocodile farms in the Northern Territory mainly rely on wild harvesting of eggs for hatching stock. In both, the skin thickens with increasing size, but there is great variability in skin thickness in individuals of similar size, and there were significant differences in the dermis in different parts of the body. It is these differences in the structure of the collagen in the dermis that may explain why buyers sometimes consider wild skins to be of higher quality than farmed skins.
Removal of the keratin layer is one of the first steps in the tanning process, and the epidermis contributes little to the overall thickness of the final tanned skin. Fat can also be deposited in the epidermis, but it is difficult to identify this histologically and we were unable to do so. It is more likely that changes in the dermis would be more significant in determining the "quality" of a skin, and may explain why some industry players noted that the skin of wild animals is thicker.
Despite the four identified skin structure patterns, the patterns of collagen in the dermis may be influenced by the conditions under which captive crocodiles are reared. The longitudinal collagen bundles in the dermis of wild crocodiles tended to be narrower and more compact compared to farm animals, which had a loose architecture. Again, the histological techniques used here failed to determine whether fat levels in the dermis differed significantly.
The distribution of keratin throughout the scales, especially in the interscalar area, makes the skin flexible. This is a clear effect of breeding on the structure and quality of the leather produced.
Implications
Recommendations