Cecidochares connexa (MACQUART) (DIPTERA:
TEPHRITIDAE) SEBAGAI AGENS HAYATI
GULMA KIRINYUH
MURNI INDARWATMI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
INFORMASI
Dengan ini saya menyatakan bahwa tesis yang berjudul : “Biologi dan Kisaran Inang Lalat Puru Cecidochares connexa (Macquart) (Diptera: Tephritidae) sebagai Agens Hayati Gulma Kirinyuh” adalah benar merupakan hasil karya saya sendiri dan belum pernah diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Semua sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, November 2006
MURNI INDARWATMI. Biologi dan Kisaran Inang Lalat Puru Cecidochares
connexa (Macquart) (Diptera: Tephritidae) sebagai Agens Hayati Gulma
Kirinyuh. Dibimbing oleh UTOMO KARTOSUWONDO dan PUDJIANTO. Lalat puru, Cecidochares connexa (Macquart) (Diptera: Tephritidae), adalah spesies eksotik yang berperan penting sebagai agens hayati gulma kirinyuh atau Chromolaena odorata (L.) King & Robinson (Asteraceae), yang diintroduksi dari Colombia tahun 1993. Sesudah dilepaskan di lapangan tahun 1995, aspek biologi lalat ini di lapangan belum banyak dievaluasi. Tujuan penelitian adalah 1) mempelajari biologi lalat C. connexa di lapangan, 2) mempelajari perkembangan dan jumlah instar larva, dan 3) mempelajari kisaran inang pada tiga tanaman famili Asteraceae. Biologi lalat puru yang diteliti meliputi perkembangan dan siklus hidup, serta lama hidup dan keperidian. Pendugaan instar larva dilakukan dengan mengukur mandibel, sklerit hipofaring dan volume larva. Hasil pengukuran tersebut disajikan dalam distribusi frekuensi, dan setiap puncak grafik mewakili satu instar. Penelitian kisaran inang dilakukan dengan mengamati tingkat infestasi dan oviposisi, perkembangan dan kemampuan hidup larva, serta perkembangan puru pada tanaman uji. Hasil penelitian menunjukkan bahwa telurdiletakkan berkelompok, dalam pucuk terminal maupun lateral yang daunnya belum membuka. Larva masuk ke dalam jaringan batang, membentuk puru batang, dan tumbuh sampai menjadi pupa. Puru mulai terlihat secara visual dua minggu setelah oviposisi. Dalam satu puru terdapat 1-12 larva. Larva instar akhir membuat jendela puru yang kelak menjadi jalan keluarnya imago. Dalam satu puru berisi 1-7 pupa. Selama fase pradewasa, lalat puru hidup di dalam puru, sedangkan imagonya hidup bebas. Siklus hidup lalat puru diselesaikan dalam 56-77 hari. Tidak ada korelasi antara ukuran puru dengan jumlah larva. Lama hidup imago betina 8-17 hari dan imago jantan 7-11 hari. Keperidian lalat ini adalah 140-329 butir telur/betina. Mortalitas lalat puru banyak terjadi pada stadia larva, dan disebabkan antara lain oleh musuh alami terutama predator. Hasil penelitian pendugaan instar menunjukkan bahwa dalam perkembangan, larva C. connexa
MURNI INDARWATMI. Biology and Host Range of Gall Fly, Cecidochares
connexa (Macquart) (Diptera: Tephritidae) as a Biological Control Agent of
Siam Weed. Advised by UTOMO KARTOSUWONDO and PUDJIANTO. Gall fly, Cecidochares connexa (Macquart) (Diptera: Tephritidae) is an important natural enemy of siam weed Chromolaena odorata (L.) King & Robinson (Asteraceae), that was introduced to Indonesia from Colombia in 1993. After being release in 1995, the biological aspects of the fly have not been evaluated yet. This research was conducted to study: 1) the biology of C.
connexa in the field, 2) the development and number of larval instars, and 3) the
host range of the gall fly. Biological parameters observed were larval development, life cycle, longevity of adults and fecundity of the females. Larval instars were determined by observing morphological characters of mandible, sclerite hypopharyng and larval size (volume). The fly host range was evaluated by observing fly infestation and oviposition, larval development and survival, as well as gall formation on three different plant species that were closely related to siam weed (Asteraceae). Results of the study indicated that the eggs were laid in clusters in tissue of terminal or axillary bud. After hatching, larvae bored deeper into the bud and lived inside the gall that was formed on the infested bud until pupation. Gall were visible about two week after oviposition. One gall contained 1 to 13 larvae. There was no correlation between the gall size and number of larvae living in the gall. The gall fly pupate inside the gall. The adults emerged from the pupae and leaved the gall through out gall windows that were prepared and made by the last instar larvae before pupation. The life cycle of the gall fly was completed in 56-77 days. In the study was found that the highest mortality occurred when the fly was in the larval stage, and the natural enemies, especially predators, were the most important mortality factors. Longevity of the fly was about 8-17 days and 7-11 days for female and male, respectively. The fecundity of the fly was 140-329 eggs/female. Based on the size of the mandible and sclerite hypopharyng, there was indication that C.
connexa larva had four instars to complete its development. The characters of
mandible and hypopharyng were good indicators to determine larval development stage of C. connexa, but not the larval volume. Mandible character was found to be the best indicator to determine the development stage of C.
connexa. The shape and size of mandible was different among different instars
of C. connexa larvae. Form the host range test, it was known that C. connexa
© Hak cipta milik Institut Pertanian Bogor, tahun 2006
Hak cipta dilindungi
Dilarang mengutip dan memperbanyak tanpa izin tertulis dari Institut Pertanian Bogor, sebagian atau seluruhnya dalam bentuk apapun,
Cecidochares connexa (MACQUART) (DIPTERA:
TEPHRITIDAE) SEBAGAI AGENS HAYATI
GULMA KIRINYUH
MURNI INDARWATMI
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Entomologi/Fitopatologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
(Macquart) (Diptera: Tephritidae) sebagai Agens Hayati Gulma Kirinyuh
Nama : Murni Indarwatmi NRP : A451030011
Program Studi : Entomologi/Fitopatologi
Disetujui, Komisi Pembimbing
Prof. Dr. Ir. Utomo Kartosuwondo, MS Dr. Ir. Pudjianto, M.Si Ketua Anggota
Diketahui,
Ketua Program Studi Dekan Sekolah Pascasarjana Entomologi/Fitopatologi
Dr. Ir. Sri Hendrastuti Hidayat, M.Sc Prof. Dr. Ir. Khairil Anwar Notodiputro, MS
مﻴﺣرﻟاﻦﻣﺣرﻟاﷲامﺴﺒ
Syukur alhamdulillahirabbil’alamin penulis panjatkan ke hadirat Allah SWT, atas rahmat dan karunia yang diberikan sehingga penulis dapat menyelesaikan penelitian dan penulisan tesis yang berjudul “ Biologi dan Kisaran Inang Lalat Puru Cecidochares connexa (Macquart) (Diptera: Tephritidae) sebagai Agens Hayati Gulma Kirinyuh”. Tesis ini merupakan salah satu syarat untuk memperoleh gelar Magister Sains pada Sekolah Pascasarjana, Institut Pertanian Bogor.
Pada kesempatan ini penulis mengucapkan terima kasih kepada komisi pembimbing Prof. Dr. Ir. Utomo Kartosuwondo, MS sebagai ketua dan Dr. Ir. Pudjianto, M.Si sebagai anggota, atas pengarahan, motivasi, dan bimbingan dengan penuh keikhlasan mulai penyusunan usulan penelitian, pelaksanaan penelitian dan penulisan tesis.
Kepada Kepala Batan dan Kepala Pusdiklat Batan, disampaikan terima kasih sebesar-besarnya atas kesempatan dan beasiswa yang diberikan kepada penulis untuk melanjutkan studi ke Program Pascasarjana IPB. Ucapan terima kasih juga penulis sampaikan kepada Rektor IPB, Dekan Sekolah Pascasarjana IPB dan seluruh Staf Pengajar Program studi Entomologi/Fitopatologi, yang telah memberi kesempatan kepada penulis untuk menyelesaikan studi pada Program Studi Entomologi/Fitopatologi. Penulis mengucapkan terima kasih kepada Dirjen Pendidikan Tinggi Departemen Pendidikan Nasional yang telah memberikan dana penelitian ini melalui program Hibah Tim Pasca Sarjana – DIKTI tahun 2003-2006.
Penulis mengucapkan terima kasih yang setulus-tulusnya kepada Ayahanda Suhirman (alm) dan Ibunda Darinah serta seluruh keluarga atas doa dan dukungannya. Tak lupa pula penulis berterima kasih kepada suamiku tercinta Bambang Sutoyo dan anak-anak tersayang Muhammad Farhan dan Nabila Nursya’bani atas doa dan kasih sayang serta pengorbanan yang diberikan agar penulis dapat menyelesaikan studi.
Kepada Akhmad Rizali, SP, M.Si, Hasmiandy Hamid, SP, M.Si dan Ropiudin, STP, penulis berterima kasih sebesar-besarnya atas bimbingan dan sarannya dalam pengolahan data maupun penulisan. Terima kasih juga penulis sampaikan kepada rekan-rekan tim Hibah Pascasarjana, anggota Laboratorium Bioekologi Parasitoid dan Predator Departemen Proteksi Tanaman IPB, serta semua pihak atas dukungan dan bantuannya selama dan sesudah penelitian. Semoga Allah SWT memberikan balasan amal baik mereka dengan pahala tak terhingga. Akhirnya, semoga tesis ini dapat berguna bagi kita semua. Amin.
Bogor, November 2006
Murni Indarwatmi
Penulis dilahirkan di Purwokerto, Jawa Tengah pada tanggal 30 November 1969, sebagai anak pertama dari tiga bersaudara dari ayah bernama Suhirman (alm) dan Ibu bernama Darinah.
Penulis menamatkan Sekolah Lanjutan Tingkat Atas pada SMA Negeri I Purwokerto pada tahun 1987. Pada tahun yang sama, penulis melanjutkan pendidikan pada Fakultas Biologi, Universitas Gadjah Mada (UGM) Yogyakarta. Penulis memilih Jurusan Biologi Lingkungan dan berhasil meraih gelar Sarjana Biologi pada tahun 1993.
Sejak tahun 1994, penulis bekerja sebagai Staf Peneliti di Pusat Aplikasi Teknologi Isotop dan Radiasi Badan Tenaga Nuklir Nasional (PATIR-BATAN) Jakarta. Selanjutnya, penulis mendapat kesempatan untuk melanjutkan pendidikan Ke Program Magister Sains tahun 2003 pada Sekolah Pascasarjana, Institut Pertanian Bogor, Program Studi Entomologi/Fitopatologi dengan bantuan biaya dari Pusat Pendidikan dan Latihan Badan Tenaga Nuklir Nasional (PUDIKLAT-BATAN).
Halaman
Lalat Puru C. connexa sebagai Agens Hayati Gulma Kirinyuh... 4
Kisaran Inang Lalat Puru C. connexa... 6
Interaksi Serangga-Tanaman dalam Pembentukan Puru... 6
Indikator Instar Larva... 7
Spesies Gulma Eksotik Invasif Kirinyuh ... 8
BAHAN DAN METODE ... 11
Waktu dan Tempat Penelitian ... 11
Metode Penelitian ... 11
Penyediaan Koloni Lalat Puru C. connexa untuk Penelitian Lapangan ... 11
Pengamatan Biologi Lalat Puru C. connexa di Lapangan ... 12
Pengamatan Perkembangan dan Siklus Hidup Lalat Puru C. connexa di Lapangan... 12
Pendugaan Instar Larva Lalat Puru C. connexa dengan Mengukur Mandibel dan Sklerit Hipofaring ... 12
Pendugaan Instar Larva C. connexa dengan Mengukur Volume Larva ... 14
Pengamatan Keterkaitan antara Perkembangan Puru dengan perkembangan Larva... 16
Pengamatan Lama Hidup dan Keperidian Lalat Puru C. connexa di Lapangan... 17
Biologi Lalat Puru C. connexa di Lapangan ... 20
Perkembangan dan Siklus Hidup Lalat Puru C. connexa di Lapangan ... 20
Instar Larva Lalat Puru C. connexa... 26
Keterkaitan antara Perkembangan Puru dengan Perkembangan Larva... 32
Lama Hidup dan Keperidian Lalat Puru C. connexa di Lapangan ... 34
Kisaran Inang Lalat Puru C. connexa... 38
KESIMPULAN DAN SARAN... 44
Kesimpulan ... 44
Saran... 45
DAFTAR PUSTAKA ... 46
Halaman 1 Tiga spesies tanaman yang digunakan sebagai tanaman uji dan
kontrol pada uji kisaran inang ... 18 2 Perkembangan pradewasa lalat puru C. connexa... 21 3 Mortalitas lalat puru C. connexa pada masing-masing fase
perkembangan di lapangan... 23 4 Parasitoid dan predator yang menyerang lalat C. connexa pada
masing-masing stadia ... 24 5 Rata-rata jumlah larva/puru berumur 1-5 minggu sesudah oviposisi ... 25 6 Ukuran mandibel dan hipofaring larva lalat puru C. connexa pada
setiap instar... 28 7 Parameter kehidupan imago lalat puru C. connexa di lapangan ... 35 8 Perkembangan lalat puru C. connexa pada beberapa tanaman uji ... 38 9 Tingkat infestasi dan jumlah telur C. connexa pada tiga tanaman
Halaman 1 Mandibel dan sklerit hipofaring larva lalat puru C. connexa yang
telah dilakukan digitasi dengan program tpsdig ... 14 2 Pengukuran volume larva: (a) larva C. connexa yang telah
didigitasi dengan program tpsdig, (b) posisi titik-titik digitasi, dan (c) hasil digitasi dirotasikan ke sumbu X sehingga diperoleh
setengah volume larva ... 15 3 Siklus hidup lalat puru C. connexa: (a) telur, (b) larva, (c) pupa,
dan (d) imago ... 20 4 Sintasan lalat puru C. connexa di lapangan ... 22 5 Larva lalat puru C. connexa yang terserang musuh alami: (a) larva
yang terserang parasitoid famili Braconidae dan (b) puru koyak
dan larva dimangsa oleh predator (B) ... 26 6 Distribusi frekuensi ukuran larva C. connexa: (a) panjang
mandibel, (b) lebar mandibel, (c) panjang hipofaring, dan (d) lebar
hipofaring ... 27 7 Perkembangan mandibel dan sklerit hipofaring larva C. connexa
pada masing-masing instar (perbesaran 100 x)... 29 8 Distribusi frekuensi volume larva C. connexa... 30 9 Perkembangan instar larva C. connexa per minggu pengamatan ... 31 10 Perkembangan diameter dan panjang puru batang C. odorata per
minggu pengamatan ... 33 11 Korelasi antara diameter dan panjang puru dengan jumlah larva ... 34 12 Rata-rata jumlah telur harian lalat puru C. connexa di lapangan... 35 13 Tiga tanaman uji dan kontrol yang digunakan dalam uji kisaran
inang: (a) kirinyuh (kontrol), (b) babadotan, (c) daun tanah, (d)
babanjaran ... 39 14 Jumlah larva/pucuk lalat puru C. connexa pada tiga tanaman uji
dan kontrol ... 41 15 Perkembangan diameter dan panjang puru kirinyuh (kontrol) dan
Latar Belakang
Lalat puru Cecidochares connexa (Macquart) (Diptera: Tephritidae) adalah lalat pembentuk puru batang (stem gallers) pada tanaman inang gulma kirinyuh atau Chromolaena odorata (L.) King & Robinson (Asteraceae) (McFadyen et al. 2003). Lalat puru meletakkan telur pada permukaan pucuk terminal maupun lateral yang daunnya belum membuka. Telur menetas menjadi larva, kemudian larva masuk ke dalam jaringan batang. Puru mulai terlihat secara visual dua minggu setelah oviposisi. Adanya larva dalam puru batang dapat menghambat pertumbuhan batang, produksi biji, dan mengurangi karbohidrat dalam penyimpanan untuk cadangan makanan (Erasmus et al. 1992). Jika terdapat dalam jumlah banyak, puru dapat mematikan tanaman inang (Ehler et al. 1984 dalam McFadyen et al. 2003).
Muniappan dan Bamba (2002) menyebutkan bahwa ada 13 spesies dari genus Cecidochares dan semua berasal dari daerah tropis di Amerika. Semua spesies dari genus ini diketahui membentuk puru batang atau puru bunga atau memakan bunga pada tanaman inang Asteraceae. Sebagian besar spesies pembentuk puru adalah sangat spesifik inang dan kadang-kadang hanya menyerang satu jenis tanaman. Oleh karena itu, lalat puru ini sangat menguntungkan untuk digunakan sebagai agens hayati gulma kirinyuh.
seperti karet, kelapa, kelapa sawit, kakao, tebu, kapas, dan sengon (SEAWIC 1991) serta dapat menurunkan hasil panen (Setiadi 1989; Syamsudin et al. 1993; Tjitrosemito 1998). Kirinyuh juga mendominasi suatu habitat dan mendesak padang penggembalaan, seperti di hutan lindung Pananjung Jawa Barat, dan Taman Nasional Baluran Jawa Timur, sehingga satwa yang dilindungi kekurangan hijauan (Tjitrosemito 1998).
Pelepasan lalat puru C. connexa telah dilakukan pada tahun 1995 di Parung Panjang, Jawa Barat, dan kemudian di beberapa daerah lainnya. Walaupun telah tersebar dan mapan, khususnya di Jawa Barat (Chenon et al. 2002), namun C. connexa belum mampu menekan populasi kirinyuh. Evaluasi terhadap biologi lalat puru di lapangan sesudah pelepasan termasuk penelitian perkembangan dan jumlah instar larva C. connexa belum banyak dilakukan. Salah satu kendala untuk mengamati perkembangan dan jumlah instar adalah karena sepanjang stadia larva sampai pupa lalat C. connexa hidup di dalam puru batang sehingga sulit diamati pergantian kulit dan perkembangan instarnya.
Masalah inang alternatif bagi lalat puru C. connexa juga perlu diteliti lebih lanjut. Hal ini penting bagi kelangsungan hidup lalat puru di lapangan karena pada musim kemarau kirinyuh mengering dan mati sehingga populasi lalat puru
C. connexa menurun. Pada musim hujan, kirinyuh akan tumbuh dengan cepat
dan lalat puru C. connexa tidak dapat mengejar pertumbuhan kirinyuh. Hal ini dapat diatasi apabila lalat puru C. connexa mempunyai inang alternatif untuk bertahan hidup pada saat tidak ada gulma kirinyuh.
Berdasarkan permasalahan di atas dan pentingnya lalat puru C. connexa
Tujuan Penelitian
Secara umum tujuan penelitian adalah untuk mempelajari biologi dan kisaran inang C. connexa di lapangan sesudah pelepasan. Secara khusus penelitian ini bertujuan: 1) mempelajari biologi dan siklus hidup lalat puru
C. connexa di lapangan, 2) mempelajari perkembangan dan jumlah instar larva
C. connexa, dan 3) mempelajari kisaran inang lalat puru C. connexa pada tiga
tanaman famili Asteraceae yang berpotensi sebagai inang alternatif.
Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan informasi dasar perkembangan instar larva, kemampuan hidup lalat puru C. connexa yang sesungguhnya di lapangan, dan potensi untuk memperluas inang serta kemampuan hidup pada inang alternatif. Kemampuan hidup di lapangan dan adanya inang alternatif merupakan landasan penting dalam usaha konservasi
TINJAUAN PUSTAKA
Lalat Puru C. connexa sebagai Agens Hayati Gulma Kirinyuh Lalat puru C. connexa diketahui dengan sinonim Urophora connexa
Macquart tahun 1848, Trypeta nigerrima Loew tahun 1862, dan Oedaspis
leucotricha Schiner 1868. Spesimen lalat ini disimpan dalam koleksi di U.S.
National Museum (USNM), Washington DC. Dr. G. Steyskal dari USNM berdasarkan spesimen dari Bolivia dan Trinidad memberi nama
Procecidochares connexa. Dr. Allen Norrbom, seorang ahli spesialis
Tephritidae di USNM mengkonfirmasi identifikasi tersebut. Dr. Norrbom tidak menerima nama P. connexa seperti yang telah digunakan pada literatur sebelumnya dan memberi nama C. connexa pada tahun 1992 (Munniappan & Bamba 2002).
Imago berwarna hitam, sayap transparan dengan strip hitam berselang seling. Pada bagian posterior masing-masing tergit juga terdapat strip hitam berselang seling dengan putih perak. Imago betina berukuran panjang 6,9 mm dan lebar 2 mm dengan rentang sayap 11,2 mm, sedangkan jantannya berukuran panjang 5,6 mm dan lebar 1,8 mm dengan rentang sayap 10 mm. Telur berbentuk lonjong dengan ujung meruncing, dan berwarna putih berukuran panjang 0,7 mm dan diameter 0,2 mm. Larva berbentuk oval silindris, berwarna putih transparan dan menjadi putih krem pada perkembangan selanjutnya. Larva instar akhir berukuran panjang 4,2 mm dan lebar 2,1 mm. Pupa berwarna putih kekuningan pada waktu muda dan berubah menjadi coklat, berukuran panjang 4,2 mm dan lebar 2,1 mm (Widayanti et al. 1999).
jendela puru. Lama stadia pupa adalah 15-25 hari. Lalat dewasa aktif antara pukul 08.00 – 14.00. Oviposisi biasanya terjadi antara pukul 10.00 – 14.00. Setiap betina mampu meletakkan 50-70 telur sepanjang hidupnya. Lama hidup lalat dewasa adalah 5-11 hari (McFadyen et al. 2003). Siklus hidup lalat C.
connexa 47-73 hari dengan rata-rata 60 hari (Chenon et al. 2002).
Lalat puru C. connexa mempunyai sifat-sifat biologis yang sangat dekat dengan Procecidochares alani Steyskal dan Procecidochares utilis Stone.
P. alani dan P. utilis telah digunakan sebagai agens hayati gulma Ageratina
(Eupatorium) adenophora (Sprengel) dan A. riparia (Regel) di banyak negara
(Sipayung & Chenon 1995). Berdasarkan keberhasilan pengendalian kedua gulma tersebut oleh lalat puru yang serupa dengan C. connexa yaitu P. utilis
dan P. alani (Julien & Griffiths 1998), C. connexa diusulkan sebagai agens
hayati gulma kirinyuh (Cock 1984).
Lalat puru C. connexa telah digunakan sebagai agens hayati gulma kirinyuh di beberapa negara. Introduksi pertama dilakukan di Indonesia dan Filipina pada tahun 1993. Lalat ini kemudian diintroduksi ke Papua New Guinea dan Palau pada tahun 1996, serta ke Guam pada tahun 1998 (Muniappan 2002). Introduksi lalat ini di Guam adalah hasil perbanyakan koloni C. connexa
dari Indonesia (Muniappan & Bamba 2002). Sekarang lalat ini sudah mapan di Palau, Papua New Guinea, Guam, dan Thailand (McFadyen et al. 2003).
Di Indonesia, lalat puru diintroduksi dari Colombia pada tahun 1993 oleh Balai Penelitian Kelapa Sawit (BPKS) Marihat, Sumatera Utara. Uji kekhususan inang dilakukan pada tahun 1994. Pelepasan lalat puru C. connexa
Kisaran Inang Lalat Puru C. connexa
Dalam genus Cecidochares terdapat 13 spesies yang semua berasal dari Amerika tropis. Semua spesies dari genus ini diketahui membentuk puru batang atau puru bunga atau pemakan bunga pada tanaman inang Asteraceae. Sebagian besar spesies pembentuk puru adalah sangat spesifik inang, kadang-kadang hanya menyerang satu jenis tanaman (Muniappan & Bamba 2002). Tanaman inang yang tercatat pada label spesimen lalat puru C. connexa di USNM adalah
C. odorata di Argentina Utara (puru batang), di Trinidad (puru bunga), dan
Chromolaena sp. di Panama (puru batang) (Muniappan & Bamba 2002).
Zachariades et al. (1998) melaporkan bahwa lalat C. connexa yang dikoleksi dari tumbuhan kirinyuh di Indonesia tidak dapat berkembang biak di Afrika Selatan. Hal ini berarti lalat C. connexa yang diuji kekhususan inang di Indonesia adalah biotipe yang sangat spesifik inang. Namun demikian, pada tumbuhan babadotan (A. conyzoides) dan daun tanah (A. inulifolium) lalat
C. connexa mampu meletakkan telur walaupun larva tidak berkembang lebih
lanjut sehingga tidak dapat menyelesaikan siklus hidupnya (Sipayung & Chenon 1995)
Interaksi Serangga-Tanaman dalam Pembentukan Puru
Puru (gall) adalah penyimpangan pola pertumbuhan abnormal yang dihasilkan oleh reaksi spesifik terhadap keberadaan dan aktivitas organisme asing (Anonim 2006). Puru berkembang dalam berbagai bagian tanaman seperti daun, batang, akar, buah dan bunga (Jolivet 1998). Setiap serangga membentuk hanya satu bagian spesifik dari tanaman (Anonim 2006). Bentuk, ukuran dan warna puru umumnya bervariasi (Jolivet 1998).
Pada puru yang disebabkan oleh serangga, puru terbentuk ketika tanaman bereaksi terhadap berbagai stimulus seperti cairan yang diinjeksi oleh imago pada saat meletakkan telur atau adanya larva serangga yang mengeluarkan sekresi atau saliva pada jaringan tanaman (Drees 2006). Jolivet (1998) juga mengemukakan bahwa umumnya puru belum tumbuh sebelum telur menetas menjadi larva dan mulai aktif makan jaringan tanaman. Pertumbuhan puru berhenti ketika larva menjadi pupa. Interaksi serangga dan tanaman menghasilkan tipe puru berbeda. Puru biasanya berbentuk bola, membengkak atau melengkung.
Indikator Instar Larva
Pendugaan instar larva untuk larva yang mudah diamati secara langsung biasanya dilakukan dengan pengamatan langsung secara visual pergantian kulit yang terjadi. Untuk serangga yang tidak dapat diamati secara langsung, pengukuran dilakukan terhadap struktur yang dapat dipercaya untuk identifikasi, antara lain struktur tersklerotisasi seperti mandibel dan kapsul kepala.
Beberapa contoh pendugaan instar yang pernah dilakukan misalnya, Rocha et al. (2004) yang mengukur kait mandibel dan dimensi tubuh untuk mengamati jumlah instar pada Fopius arisanus (Hymenoptera: Braconidae). Godin et al. (2002) mengukur lebar kapsul kepala Acrobasis vaccinii
(Lepidoptera: Pyralidae). Alencar et al. (2001) mengukur panjang kapsul kepala lateral dan lebar apodema kepala Simulium pervlafum (Diptera: Simuliidae). Lebar kapsul kepala sering digunakan untuk menentukan instar berbagai serangga ordo Lepidoptera (Godin et al. 2002).
Jumlah instar larva pada Diptera berkisar 4-9, pada umumnya 4 instar dan kadang-kadang mereduksi menjadi 3 (Foote 1991). Chenon et al. (2002) melaporkan bahwa terdapat 3 instar pada perkembangan lalat puru C. connexa.
Spesies Gulma Eksotik Invasif Kirinyuh
Kirinyuh adalah tumbuhan perdu dengan tinggi 1,5-2 m dan kadang-kadang mencapai 6-7 m apabila terdapat pohon-pohon yang menopangnya. Tumbuhan bersifat herba pada waktu masih muda, kemudian berkayu dan bercabang-cabang banyak. Batang hijau, berbentuk silindris dan sedikit berbulu. Daun berhadapan, berbentuk bulat telur dengan ujung runcing, bergerigi kasar atau hampir rata dan permukaannya berbulu halus (Tjitrosoedirdjo 1989).
Bunga kirinyuh tersusun dalam tipe malai rata, terdiri atas 25-30 kepala, masing-masing kepala mempunyai 30-36, bunga bertangkai 1-2 cm. Kelopak 5, bunga putih keunguan, dan sedikit berbau. Mahkota bunga seperti genta, berlobi 5, masing-masing lobi berbentuk segitiga. Putik berbelah 2 dan panjang. Buah bersudut, berukuran panjang 5 mm coklat atau hitam dengan rambut-rambut pendek pada sudut-sudutnya.
Kirinyuh berkembang biak dengan bijinya (Tjitrosoedirdjo 1989). Pada tingkat kepadatan yang tinggi, seperti di Pantai Gading, tumbuhan ini dapat menghasilkan sekitar 109 biji/ha. Pelepasan buah sangat memerlukan kondisi cuaca yang kering dan berangin. Penyebaran buah secara khas dilakukan oleh angin dan mungkin juga oleh binatang (Binggeli 1997). Tumbuhan ini termasuk dalam suku Eupatoria sub famili Lactucoideae yang sebagian besar anggotanya merupakan gulma penting.
kirinyuh pertama kali dilaporkan pada tahun 1934 dari koleksi herbarium di Lubuk Pakam, Sumatera Utara oleh van Meer Mohr dan saat ini masih berada di Herbarium Bogoriense, Bogor. Saat ini, penyebaran kirinyuh meliputi seluruh wilayah Indonesia mulai dari Sumatera, Jawa, Nusa Tenggara Timur, dan beberapa daerah lainnya (Tjitrosemito 1999).
Kirinyuh merupakan gulma penting pada sistem produksi pertanian, tanaman budidaya, dan hutan tanaman industri jati karena dapat berkompetisi secara kuat dengan tanaman budidaya (Setiadi 1989; Syamsudin et al. 1993; Tjitrosemito 1998). Sipayung dan Chenon (1995) melaporkan bahwa keberadaan kirinyuh pada tanaman perkebunan (kakao, karet, kelapa sawit) dapat menyebabkan kehilangan hasil terutama di pulau Jawa dan Kalimantan. Di daerah pengembalaan hutan lindung Pananjung, Jawa Barat, dan Taman Nasional Baluran, keberadaan kirinyuh dapat mengurangi hamparan padang pengembalaan banteng (Bos javanicus) dan rusa (Muntiacus muncak). Selain itu, kirinyuh juga dapat menimbulkan keracunan pada hewan-hewan ternak yang memakannya karena kandungan nitrat yang sangat tinggi terutama pada tunas-tunas muda yang tumbuh kembali sesudah pemangkasan (Torres & Paller 1989). Di Afrika bagian barat, tumbuhan ini mampu menekan regenerasi spesies pohon pada daerah yang mengalami suksesi, sedangkan di Afrika bagian selatan, mengurangi keanekaragaman spesies dan merupakan ancaman pada daerah tepi hutan (Binggeli 1997).
Waktu dan Tempat Penelitian
Penelitian merupakan penelitian lapangan yang dilaksanakan pada bulan April 2005 – Februari 2006. Penelitian biologi lapangan dilaksanakan di salah satu lahan di Kampus IPB, Darmaga Bogor, sedangkan penelitian kisaran inang dilakukan di lahan sekitar gedung Departemen Proteksi Tanaman IPB Bogor. Pengamatan biologi secara umum dilakukan di Laboratorium Bioekologi Parasitoid dan Predator, Departemen Proteksi Tanaman, IPB Bogor.
Metode Penelitian
Penyediaan Koloni Lalat Puru C. connexa untuk Penelitian Lapangan
Lalat yang digunakan untuk penelitian ini berasal dari Desa Setu, Kecamatan Jasinga, Bogor. Lalat dari lapangan diperoleh dengan mengumpulkan puru yang sudah berjendela dan di dalamnya berisi larva instar akhir atau pupa. Puru berjendela tersebut kemudian dimasukkan dalam kurungan pemeliharaan di laboratorium. Setelah muncul, lalat diberi pakan berupa madu 10% yang dioleskan pada kapas dan digantung di dalam kurungan pemeliharaan. Selanjutnya lalat siap digunakan untuk penelitian.
Sepuluh hari setelah pelepasan lalat, kurungan dibuka kembali agar kirinyuh dapat tumbuh normal dan percobaan berjalan secara alami.
Pengamatan Biologi Lalat Puru C. connexa di Lapangan
Pengamatan Perkembangan dan Siklus Hidup Lalat Puru C. connexa
di Lapangan. Pengambilan contoh untuk pengamatan telur dilakukan pada hari ke-4, 6, 8, dan 9 sebanyak 5 pucuk/kurungan. Pengambilan contoh untuk pengamatan larva dan pupa selanjutnya dilakukan setiap minggu dari umur 1 sampai 12 minggu sebanyak 5 puru/kurungan. Jadi, jumlah total pengambilan pucuk atau puru dalam satu kali pengamatan adalah 45. Contoh pucuk atau puru dari lapangan dibawa ke laboratorium dan dibedah di bawah mikroskop, kemudian diamati letak dan jumlah telur, larva atau pupa yang ada di dalamnya. Diamati pula morfologi dan jumlah individu lalat per pucuk. Selanjutnya diambil sebanyak 20 telur dan 20 pupa untuk diukur volumenya. Larva yang ditemukan disimpan di dalam freezer untuk pengamatan pendugaan instar. Setelah puru berjendela, 5 puru dikurung dengan kurungan kasa berukuran panjang ± 20 cm dengan diameter ± 7 cm agar imago yang muncul tidak terbang keluar untuk pengamatan nisbah kelamin. Diamati pula waktu dan jumlah imago yang muncul. Pucuk atau puru yang rusak oleh serangan musuh alami yang berada di dalam kurungan percobaan yang sudah dibuka juga diamati. Data perkembangan dan siklus hidup lalat dilaporkan secara deskriptif dan dalam bentuk tabel serta grafik.
diletakkan di atas gelas obyek dan ditetesi larutan Hoyer, diatur sampai mendapatkan posisi yang sesuai, kemudian ditutup dengan gelas penutup. Preparat kemudian dikeringkan dengan hot plate selama beberapa hari (Borror
et al. 1992). Mandibel dan sklerit hipofaring terlihat berwarna gelap.
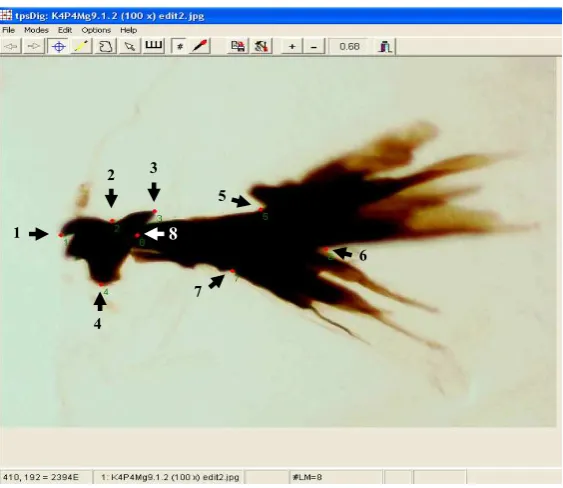
Preparat mandibel dan hipofaring sebanyak 529 diamati dibawah mikroskop binokuler Olympus BX 51 dan difoto menggunakan kamera digital mikroskop Olympus DP 11 dengan perbesaran 100 x. Foto ditransfer ke komputer, kemudian dilakukan digitasi menggunakan program tpsdig (Bennet & Hoffmann 1998). Digitasi dilakukan terhadap bagian mandibel dan sklerit hipofaring yang keberadaannya konsisten sepanjang stadia larva yaitu mandibel, yang diukur dari ujung kait ke pangkal mandibel (jarak 1-2-3) selanjutnya disebut panjang mandibel, mandibel jarak 3-4 selanjutnya disebut lebar mandibel, sklerit hipofaring-tentorofaring jarak 6-8 selanjutnya disebut panjang hipofaring dan sklerit hipofaring-tentorofaring jarak 5-7 selanjutnya disebut lebar hipofaring (Gambar 1).
Pengukuran nilai konversi dilakukan untuk mendapatkan nilai konversi dari nilai vektor ke ukuran obyek sesungguhnya. Skala mikrometer difoto dengan semua perbesaran yang ada menggunakan kamera digital mikroskop Olympus DP 11 pada mikroskop binokuler Olympus BX 51 dan SZ 11. Hasil foto skala mikrometer ditransfer ke komputer dan dilakukan digitasi menggunakan program tpsdig (Bennet & Hoffmann 1998). Hasil digitasi berupa nilai vektor, selanjutnya dimasukkan ke dalam persamaan berikut untuk mendapatkan ukuran sesungguhnya.
Dv = ((X1−X2)2 +(Y1−Y2)2) [1] dimana Dv adalah jarak vektor; X1,X2,Y1, dan Y2 adalah absis dan ordinat
titik-titik yang diukur (titik-titik 1 dan titik-titik 2)
Ds (mm) = Dvx/Dpx [2]
dimana Ds (mm) adalah jarak sesungguhnya obyek yang diukur; Dvjarak vektor;
1
2 3
4
5
6
7 8
Gambar 1 Mandibel dan sklerit hipofaring larva lalat puru C. connexa yang telah dilakukan digitasi dengan program tpsdig
satuannya (mm); x adalah perbesaran (magnifikasi), Dv dan Dp diukur pada perbesaran yang sama.
Pendugaan Instar Larva Lalat Puru C. connexa dengan Mengukur Volume Larva. Untuk mengetahui ukuran larva, lebih tepat dilakukan dengan mengukur volume larva daripada panjang dan lebar larva. Hal ini disesuaikan dengan perkembangan bentuk larva. Larva yang baru keluar dari telur berbentuk oval silindris, dalam perkembangan selanjutnya panjang larva hanya sedikit pertambahannya, sementara volumenya bertambah dengan pesat.
terhadap titik-titik yang akan diukur yaitu seperempat permukaan larva pada daerah yang cembung (Gambar 2A). Larva C. connexa berbentuk oval dan diasumsikan simetris atas- bawah sehingga hasil digitasi seperempat larva ini (Gambar 2B) kemudian dirotasikan ke sumbu X untuk memperoleh setengah volume larva (Gambar 2C). Larva C. connexa juga simetris kiri dan kanan, sehingga untuk mendapatkan volume larva penuh, volume setengah larva hasil rotasi kemudian dikalikan dua.
0 200 400 600 800 1000 1200 1400
(c) (c) (b)
X
Hasil digitasi dengan tpsdig dibuka dalam program Microsoft Excell sehingga diperoleh ordinat dan absis titik-titik tersebut. Titik-titik koordinat ini kemudian dibuat grafik polinomial sehingga diperoleh persamaan kuadrat y = ax2 + bx + c dan nilai a,b, dan c diketahui. Kemudian, persamaan kuadrat y dikuadratkan menjadi y = (ax2 + bx + c)2 dan dimasukkan persamaan untuk mendapatkan setengah volume larva yaitu:
(
)
Volume yang diperoleh dimasukkan ke persamaan volume (persamaan 3 dan 4) untuk mendapatkan volume yang sebenarnya.
V (mm3) = Vvx/Vpx [3]
dimana V (mm3) adalah volume sesungguhnya obyek yang diukur; Vv adalah
volume berdasarkan skala mikrometer yang telah diketahui satuannya (mm3); x adalah perbesaran (magnifikasi); Vv dan Vp diukur pada perbesaran yang sama.
V (µl) = V (mm3) [4]
dimana V (µl) adalah volume sesungguhnya dalam µl.
Data hasil pengukuran mandibel, hipofaring, dan volume larva ditampilkan dalam bentuk histogram distribusi frekuensi dalam selang kelas tertentu sehingga diperoleh pengelompokan ukuran dengan puncak–puncak yang nyata dan terpisah satu dengan lainnya yang menandakan pergantian instar. Distribusi frekuensi hasil pengukuran diasumsikan terdistribusi normal dan membentuk puncak-puncak, setiap puncak mewakili satu instar (Godin et al. 2002).
jumlah larva dianalisis dengan analisis korelasi menggunakan program Statistica
for Windows 6.0 (StatSoft 1995).
Pengamatan Lama Hidup dan Keperidian Lalat Puru C. connexa di Lapangan. Tumbuhan kirinyuh di lapangan dibersihkan dari semua serangga yang tidak diinginkan, kemudian ditutup dengan kurungan mika berdiameter 50 cm dan tinggi 70 cm yang diberi jendela dari kain kasa. Kirinyuh yang digunakan adalah tanaman yang mempunyai 7-20 pucuk tanpa telur lalat C.
connexa sebelumnya. Bagian dasar kurungan dialasi dengan tatakan pot dan
diolesi vaselin untuk menghindari datangnya semut yang dapat menyerang lalat. Satu pasang imago dilepaskan ke dalam kurungan tersebut pada pagi hari, pukul 07.00-08.00. Setelah 24 jam, lalat dikeluarkan dari kurungan dan dipindahkan ke tanaman kirinyuh baru. Lama hidup imago diamati dengan cara mencatat setiap hari lalat jantan atau betina yang mati.
Pengamatan produksi telur harian dilakukan dengan mengambil pucuk-pucuk yang sudah diinfestasi telur setiap hari, dibawa ke laboratorium untuk diamati di bawah mikroskop dan dihitung jumlah telurnya. Prosedur ini dilakukan terus menerus sampai imago mati. Keperidian dihitung dengan menjumlahkan semua telur yang dihasilkan oleh satu imago betina sejak imago tersebut muncul sampai mati. Periode sejak imago betina muncul hingga pertama kali menghasilkan telur disebut periode praoviposisi. Periode pasca oviposisi adalah periode sejak imago betina tidak lagi mengeluarkan telur hingga imago betina tersebut mati. Penelitian dilakukan dengan 3 ulangan/kurungan jadi total 27 ulangan. Data keperidian dilaporkan secara deskriptif dan dalam bentuk tabel serta grafik jumlah telur harian.
Uji Kisaran Inang Lalat Puru C. connexa
conyzoides), daun tanah (Austroeupatorium inulifolium) dan babanjaran
(Clibadium surinamense) ditetapkan sebagai tanaman uji serta tanaman
kirinyuh sebagai kontrol (Tabel 1). Tanaman kirinyuh dan babadotan diperoleh dari sekitar lahan percobaan di kampus IPB Darmaga, daun tanah diperoleh dari Gunung Bunder, dan babanjaran diperoleh dari Desa Setu Kecamatan Jasinga, Bogor. Identifikasi tanaman uji dilakukan di Biotrop, Bogor.
Tanaman kirinyuh, daun tanah dan babanjaran diperbanyak dengan stek batang, sedangkan babadotan ditanam dari anakan yang diperoleh dari lapangan. Masing-masing tanaman ditanam dalam polybag berdiameter 35 cm. Setelah tumbuh, tanaman dipangkas untuk mendapatkan tajuk dengan tunas yang banyak. Sebelum perlakuan semua tanaman uji dibersihkan dari serangga dan organisme lain.
Masing-masing tanaman uji dan kontrol yang telah bersih dari serangga dimasukkan dalam kurungan (satu kurungan satu jenis tanaman uji). Untuk satu kali perlakuan digunakan masing-masing 50 pucuk tanaman. Semua pucuk ditandai untuk memudahkan penghitungan dan pengamatan selanjutnya. Kemudian, 15 pasang imago C. connexa dilepaskan ke dalam masing-masing kurungan. Setelah 72 jam, lalat dikeluarkan kembali dari kurungan. Percobaan ini diulang 3 kali.
Pengamatan tingkat infestasi lalat C. connexa dilakukan dengan menghitung jumlah pucuk terinfestasi per total pucuk. Pucuk terinfestasi ditandai dengan adanya bintik coklat pada jaringan pucuk yang daunnya belum
Tabel 1 Tiga spesies tanaman yang digunakan sebagai tanaman uji dan kontrol pada uji kisaran inang
1 Chromolaena odorata (Kontrol) Kirinyuh Asteraceae Gulma
2 Ageratum conyzoides Babadotan Asteraceae Gulma
3 Austroeupatorium inulifolium Daun tanah Asteraceae Gulma
membuka. Data tingkat infestasi dianalisis sidik ragam, dan jika berbeda nyata diuji lebih lanjut dengan uji BNT menggunakan program Statistix for Windows Release 8 (Statistix 2003).
Pengamatan jumlah telur per pucuk dilakukan dengan mengambil contoh pucuk sebanyak tiga pucuk dari setiap tanaman yang diuji dan dibedah di bawah mikroskop. Data jumlah telur/pucuk dianalisis sidik ragam, dan jika berbeda nyata diuji lebih lanjut dengan uji BNT menggunakan program Statistix for
Windows Release 8 (Statistix 2003).
Pengamatan perkembangan larva dan kemampuan hidup lalat pada tanaman inang yang diuji diamati dengan mengambil tiga contoh pucuk atau puru dari setiap tanaman uji per minggu selama 12 minggu, kemudian dibedah di bawah mikroskop dan diamati jumlah telur, larva, atau pupa yang ada. Data hasil pengamatan ditampilkan dalam bentuk grafik.
Biologi Lalat Puru C. connexa di Lapangan
Perkembangan dan Siklus Hidup Lalat Puru C. connexa di Lapangan
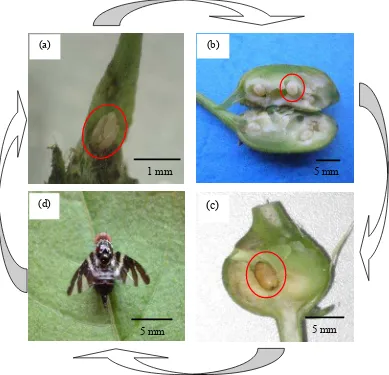
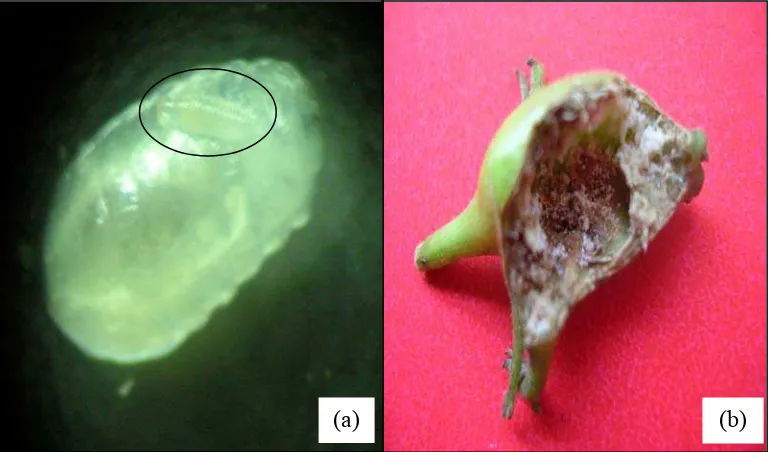
Berdasarkan pengamatan terhadap pucuk kirinyuh di lapangan diketahui bahwa telur lalat puru C. connexa diletakkan secara berkelompok dan ditancapkan pada permukaan pucuk terminal maupun lateral yang daunnya belum membuka. Telur berwarna putih, berbentuk lonjong dengan ujung agak meruncing (Gambar 3). Jumlah telur per pucuk berkisar 1 - 14 butir (Tabel 2).
Imago
(a) (b)
(c) (d)
1 mm 5 mm
5 mm 5 mm
Setelah telur menetas, larvanya masuk ke dalam jaringan batang dan hidup di dalam puru batang sampai menjadi pupa. Larva C. connexa tidak bertungkai (apoda). Skeleton kepala tidak ada (acepala). Segmen tubuh berjumlah 12 dengan barisan papilae melingkar dan antar segmen membentuk lekukan (Gambar 3). Dalam satu puru terdapat 1-12 larva dan masing-masing larva menempati satu ruang tersendiri (Tabel 2). Sebelum menjadi pupa, larva instar akhir membuat jendela puru yang jumlahnya satu jendela/larva.
Lalat puru C. connexa berpupa di dalam puru sampai berubah menjadi imago yang keluar melalui jendela puru. Pupa berbentuk oval silindris, berwarna putih kecoklatan pada saat masih muda dan menjadi coklat tua dengan ujung hitam pada saat pupa hampir muncul menjadi imago (Gambar 3). Pupa tipe coarctata. Jumlah pupa per puru adalah 1-7 (Tabel 2).
Imago lalat puru C. connexa berwarna hitam, mata berwarna merah, sayap transparan dengan gambaran pita hitam berselang-seling dengan bagian yang transparan (banded wings). Pada toraks dan abdomen terdapat pola pita berselang seling antara hitam dan putih. Imago jantan dan betina mudah dibedakan, yakni adanya ovipositor pada imago betina. Selain itu, ukuran betina lebih besar daripada yang jantan (Gambar 3).
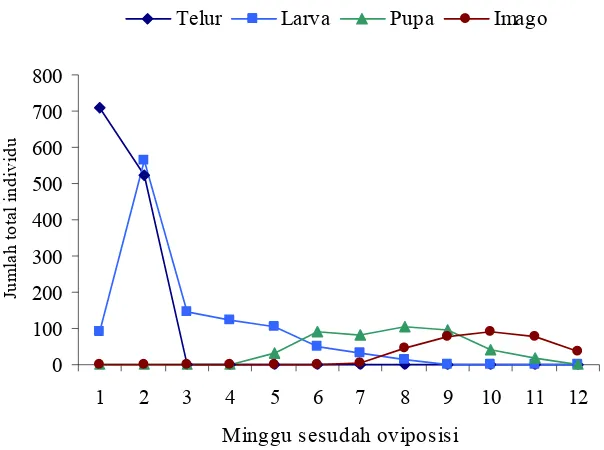
Hasil pengamatan perkembangan dan siklus hidup lalat C. connexa di lapangan dari telur hingga imago disajikan pada Gambar 4. Telur C. connexa
di lapangan hanya ditemukan pada pengamatan hari ke-4, 6, dan 8 setelah oviposisi. Larva mulai ditemukan pada minggu pertama dan mencapai puncaknya pada minggu ke-2 setelah oviposisi. Jumlah larva menurun tajam
Tabel 2 Perkembangan pradewasa lalat puru C. connexa
0
Gambar 4 Sintasan lalat puru C. connexa di lapangan
pada minggu ke-3, kemudian makin menurun dan semua larva telah menjadi pupa pada minggu ke-9 setelah oviposisi. Pupa ditemukan mulai minggu ke-5 sampai minggu ke-10 setelah oviposisi. Imago mulai ditemukan pada minggu ke-7, dan pada minggu ke-12 setelah oviposisi seluruh pupa telah menjadi imago.
Pengamatan nisbah kelamin lalat C. connexa juga dilakukan bersamaan dengan pengamatan perkembangan dan siklus hidup. Berdasarkan pengamatan terhadap 334 imago, perbandingan imago jantan dan betina C. connexa yang ditemukan di lapangan adalah mendekati 1 : 1. Hasil penelitian ini sama dengan hasil penelitian sebelumnya yang dilakukan di laboratorium (Widayanti et al. 1999). Nisbah kelamin 1:1 adalah normal dan baik untuk kelangsungan hidup lalat selanjutnya.
3-5 minggu (21-35 hari). Dengan demikian, siklus hidup lalat puru C. connexa
adalah 8-11 minggu (56-77 hari). Siklus hidup lalat di lapangan ini tidak jauh berbeda dengan siklus di laboratorium, yaitu 47-73 hari (Chenon et al. 2002).
Siklus hidup lalat C. connexa yang sama antara di laboratorium dan lapangan sangat menguntungkan. Hal ini berarti kondisi di lapangan mendukung kehidupan lalat. Kondisi ini memungkinkan lalat untuk hidup dan berkembang biak sehingga pertambahan generasi berjalan terus.
Berdasarkan jumlah individu total pada Gambar 4, dapat dihitung mortalitas lalat C. connexa pada masing-masing stadia yang tercantum pada Tabel 3. Mortalitas pada telur sangat rendah. Mortalitas banyak terjadi pada stadia larva, terutama sesudah minggu ke-2, sedangkan pada stadium pupa dan imago relatif rendah.
Mortalitas pada telur sangat rendah karena telur berada di dalam pucuk yang daunnya belum membuka sehingga telur terlindung dari kekeringan yang disebabkan oleh cuaca buruk atau sinar matahari. Penetasan telur ini lebih tinggi dari hasil penelitian Widayanti et al. (1999) yang menemukan bahwa penetasan telur yang diletakkan dalam cawan petri yang dialasi kapas basah yang diamati di laboratorium adalah 76,3%. Hal ini menunjukkan bahwa kondisi alami di lapangan merupakan kondisi yang optimum untuk penetasan telur. Selain itu, selama 10 hari sesudah pelepasan imago, kurungan percobaan dikurung dengan kain sehingga relatif aman dari musuh alami. Mortalitas pada telur lebih disebabkan oleh telur yang tidak menetas.
Tabel 3 Mortalitas lalat puru C. connexa pada masing-masing fase perkembangan di lapangan
Fase perkembangan Mortalitas (%)
Telur 9,061 *
Larva 58,808 **
Pupa 27,862 **
Larva instar 1 yang baru keluar dari telur mudah diinfeksi oleh parasitoid dan dimangsa oleh predator, karena masih berada pada permukaan pucuk dan daun-daun di pucuk mulai membuka. Parasitoid yang menyerang larva adalah dari famili Braconidae dan Ormyridae. Braconidae diduga menyerang larva C.
connexa instar muda, sedangkan Ormyridae menyerang larva instar akhir dan
pupa muda. McFadyen et al. 2003 menemukan bahwa Ormyridae adalah endoparasitoid soliter pada larva-pupa C. connexa. Predator yang menyerang larva adalah semut (Formicidae), kumbang kubah predator (Coccinellidae), belalang sembah (Mantidae), dan laba-laba (Lycosidae) (Tabel 4).
Pada minggu ke-2 puru sudah terbentuk dan larva terlindung di dalamnya. Namun demikian, diduga larva masih dapat diserang oleh parasitoid dari famili Braconidae. Parasitoid ini mempunyai ovipositor panjang dan mampu menembus puru muda yang belum berkayu. Gambar 5a menunjukkan larva berumur 3 minggu yang terserang parasitoid famili Braconidae. Seperti pada puru tumbuhan Solidago altissima yang disebabkan oleh lalat puru Eurosta
solidaginis (Diptera: Tephritidae), larva dalam puru diserang oleh parasitoid
Calliephialtes notandus (Hymenoptera: Ichneumonidae) yang mempunyai
ovipositor panjang dan mampu menembus puru muda (Anonim 2006).
Selain karena serangan musuh alami, mortalitas larva diduga disebabkan oleh adanya kompetisi. Hal ini terlihat dari rata-rata jumlah larva pada puru
Tabel 4 Parasitoid dan predator yang menyerang lalat C. connexa pada masing-masing fase perkembangan
Fase perkembangan Parasitoid Predator
Telur - -
Larva Braconidae, Ormyridae
Formicidae, Coccinellidae Mantidae, Reduviidae
Pupa Ormyridae Formicidae, Mantidae
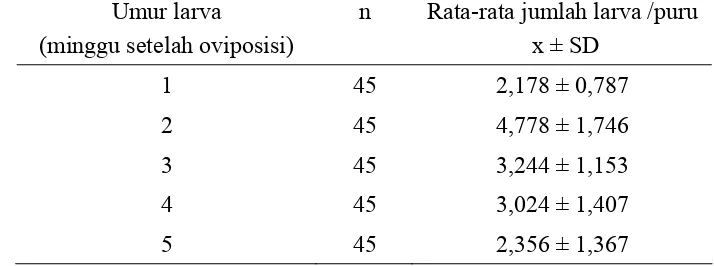
Tabel 5 Rata-rata jumlah larva/puru berumur 1-5 minggu sesudah oviposisi.
Umur larva
(minggu setelah oviposisi)
n Rata-rata jumlah larva /puru x ± SD
umur muda yang lebih banyak daripada minggu-minggu berikutnya (Tabel 5). Selain itu, ukuran larva dalam satu puru biasanya juga berbeda.
Keadaan ini menunjukan bahwa larva yang kuat akan tumbuh lebih besar dari yang lemah. Jika larva terlalu banyak dalam satu puru, beberapa diantaranya akan mati. Hal ini sesuai dengan penelitian Widayanti et al. (1999) yang menemukan bahwa kepadatan telur yang tinggi dalam satu pucuk menyebabkan tingginya tingkat kompetisi diantara larva-larva yang baru keluar dari telur. Oleh karena itu, pada minggu ke-3 sesudah oviposisi jumlah larva menurun tajam.
Pada larva instar 4 akhir dan pupa, puru sudah berjendela sehingga mudah ditembus oleh parasitoid dan predator. Parasitoid yang menyerang larva dan pupa yaitu dari famili Ormyridae. Predator yang menyerang larva yaitu semut, kepik (Reduviidae), serta belalang sembah. Kepik menyerang larva dengan menusukkan stilet ke dalam puru dan menghisap larva melalui jendela puru. Belalang sembah merusak puru dengan cara menggigit puru sehingga puru koyak dan terbuka (Gambar 5b). McFadyen et al. (2003) juga menemukan bahwa di beberapa daerah puru terbuka dan larva dimangsa oleh burung.
(b) (a)
Gambar 5 Larva lalat puru C. connexa yang terserang musuh alami: (a) larva yang terserang parasitoid famili Braconidae dan (b) puru koyak dan larva di dalamnya dimangsa oleh predator
mencari pucuk dan bertelur yang memerlukan konsentrasi tinggi dan pada saat bertelur lalat diam tidak bergerak sehingga mudah dimangsa oleh predator. Menurut McFadyen et al. (2003), imago lalat ini juga dimangsa oleh kadal.
Secara umum hasil penelitian menunjukkan bahwa mortalitas banyak terjadi pada stadium larva. Faktor-faktor eksternal yang kurang menguntungkan bagi pertumbuhan populasi lalat puru C. connexa di lapangan antara lain adalah musuh alami terutama predator. Hal ini sesuai dengan hasil penelitian McFadyen et al. (2003) yang menyebutkan bahwa serangan predator cukup signifikan yaitu dapat mencapai 50%, sedangkan serangan parasitoid hanya berkisar antara 1-15%.
Instar Larva Lalat Puru C. connexa
(2002), hasil pengukuran diasumsikan terdistribusi normal dan membentuk puncak-puncak, setiap pucak mewakili satu instar. Kurva distribusi frekuensi hasil pengukuran panjang dan lebar mandibel dalam selang kelas ukuran tertentu menghasilkan 4 puncak terpisah yang tidak tumpang tindih. Hal ini menunjukkan bahwa terdapat 4 instar pada perkembangan larva C. connexa.
Pendugaan instar juga dilakukan dengan mengukur panjang dan lebar sklerit hipofaring, dan distribusi frekuensi pengukuran kedua struktur ini ditampilkan pada Gambar 6c dan 6d. Berdasarkan pada pengukuran panjang dan lebar sklerit hipofaring ini, dihasilkan pola data yang sama dengan pengukuran panjang dan lebar mandibel yaitu 4 puncak yang menandakan terdapat 4 instar. Hal ini menunjukkan bahwa panjang dan lebar mandibel, serta panjang dan lebar hipofaring dapat digunakan sebagai indikator instar pada perkembangan larva C. connexa. Hasil penelitian sebelumnya menduga bahwa perkembangan larva C. connexa melewati 3 instar (Chenon et al. 2002), tetapi tidak ada informasi indikator yang digunakan untuk pendugaan instar. Perbedaan ini diduga karena perbedaan indikator instar yang digunakan.
Ukuran mandibel dan hipofaring larva C. connexa pada setiap instar disajikan pada Tabel 6. Ukuran mandibel meningkat pesat yaitu 4 kali lebih besar pada pergantian instar 1 ke instar 2. Pada pergantian instar 2 ke 3, dan
instar 3 ke 4 ukurannya meningkat 2 kali lebih besar. Ukuran hipofaring pada semua pergantian instar meningkat 2 kali lebih besar.
Selain perubahan ukuran mandibel yang ekstrem, perubahan instar 1, 2, dan 3 juga ditandai oleh adanya perubahan bentuk mandibel (Gambar 7). Pada larva instar 1, bentuk mandibelnya sederhana dengan kait tumpul dan warna lebih terang. Pada larva instar 2, kaitnya yang berbentuk runcing mulai terlihat melengkung ke bawah dan warnanya lebih gelap, tetapi belum ada modifikasi gigi asesori. Pada larva instar 3, kaitnya berbentuk runcing dan melengkung ke bawah, berwarna gelap, dan terjadi modifikasi mandibel menjadi satu gigi asesori. Gigi asesori ini berfungsi untuk melubangi puru yang biasa disebut dengan jendela puru (windowed gall) sebagai tempat keluarnya imago. Pada larva instar 4, bentuk dan warna mandibelnya tidak berbeda dengan mandibel instar 3, tetapi ukurannya lebih besar.
Dari pengukuran keempat struktur ini dapat diketahui bahwa ukuran mandibel paling baik untuk menduga instar larva C. connexa. Adanya ntuk
Instar 1 Instar 2
Instar 3 Instar 4
Gigi asesori
Gigi asesori
Gambar 7 Perkembangan mandibel dan sklerit hipofaring larva C. connexa
perbedaan bentuk mandibel pada instar 1, 2, dan 3 dapat digunakan upendugaan instar secara cepat tanpa mengukur mandibel. Pendugaan instar 4 tidak dapat dilakukan hanya dari pengamatan bentuk mandibel saja, tetapi perlu dilakukan juga pengukuran mandibel karena bentuk mandibel sama dengan instar 3. Demikian pula untuk pendugaan instar dengan ukuran sklerit hipofaring harus dilakukan dengan pengukuran sklerit hipofaring karena bentuk struktur ini sama pada setiap instar.
Hasil pengukuran volume larva C. connexa ditampilkan dalam bentuk distribusi frekuensi volume larva (Gambar 8). Distribusi frekuensi volume larva tidak menunjukkan pengelompokkan yang nyata dan saling tumpang tindih, sehingga sulit diamati perubahan instar yang terjadi. Dengan demikian volume larva tidak dapat digunakan untuk menduga jumlah instar larva. Hal ini sesuai dengan hasil penelitian Rocha et al. (2004) yang menemukan bahwa pengukuran jaringan lunak tidak dapat dipercaya untuk identifikasi instar larva. Hal ini disebabkan oleh banyaknya perbedaan yang diperoleh dari hasil pengukuran jaringan lunak pada metode fiksasi dan mounting yang berbeda.
Hasil pengamatan terhadap perkembangan larva C. connexa
menunjukkan bahwa lama perkembangan instar larva C. connexa bervariasi untuk masing- masing instar (Gambar 9). Pada minggu pertama sesudah oviposisi, telur baru menetas menjadi larva instar 1, tetapi sebagian kecil larva telah masuk instar 2. Larva instar 1 berbentuk silindris dan berwarna putih transparan. Lekuk-lekuk segmen belum terlihat. Pada minggu ke-2 setelah oviposisi, ditemukan larva instar 1, instar 2, dan sebagian kecil telah masuk instar 3. Dengan demikan periode instar 1 berlangsung paling singkat yaitu 1-2 minggu. Pada minggu ke-3 setelah oviposisi, larva yang ditemukan sebagian besar adalah larva instar 2 dan sebagian kecil instar 3. Pada minggu ke-4 sampai minggu ke-7 setelah oviposisi, larva yang ditemukan adalah larva instar 3 dan 4. Dengan demikian periode instar 2 berlangsung 1-3 minggu. Larva instar 2 berbentuk oval silindris dengan abdomen membesar dan berwarna putih. Lekuk-lekuk segmen sudah terlihat. Pada minggu ke-8 setelah oviposisi, larva yang ditemukan hanya larva instar 4 dan pada minggu ke-9 seluruh larva telah menjadi pupa. Oleh karena itu periode perkembangan instar 3 adalah cukup
0
Instar 1 Instar 2 Instar 3 Instar 4
panjang yaitu 2-7 minggu, sedangkan periode instar 4 adalah 4-8 minggu. Larva instar 3 mempunyai abdomen sangat membesar dan berwarna putih krem. Lekuk-lekuk segmen terlihat sangat jelas.
Penggunaan metode morfometri dengan program tpsdig untuk pengukuran morfologi sangat akurat bahkan untuk serangga-serangga dengan ukuran yang sangat kecil. Pemilihan struktur karakteristik yang akan diukur sangat menentukan dapat tidaknya struktur tersebut digunakan sebagai indikator instar. Penemuan metode pendugaan instar ini sangat bermanfaat bagi penelitian-penelitian perkembangan instar larva terutama untuk serangga-serangga yang hidupnya tersembunyi dan sulit diamati dengan pengamatan visual.
Keterkaitan antara Perkembangan Puru dengan Perkembangan Larva Perkembangan diameter dan panjang puru batang kirinyuh per minggu pengamatan menunjukkan pola yang sama (Gambar 10). Pada minggu pertama setelah oviposisi, sebagian telur sudah menetas tetapi puru belum terlihat secara visual. Puru mulai terbentuk dan dapat terlihat secara visual dua minggu setelah oviposisi. Perkembangan puru terus berlanjut sampai mencapai maksimal pada minggu ke-5 setelah oviposisi. Pada minggu ke-6 setelah oviposisi dan seterusnya panjang dan lebar puru batang relatif tetap.
0
0
Gambar 11 Korelasi antara diameter dan panjang puru dengan jumlah larva
Walaupun ukuran puru tidak dapat digunakan untuk memprediksi jumlah larva yang ada di dalamnya, namun puru sering digunakan sebagai tolok ukuruntuk menduga populasi dan intensitas serangan lalat C. connexa di lapangan. Kepadatan populasi lalat diukur dari jumlah puru per tanaman atau per satuan luas tertentu. Chenon et al. (2002) mengamati adanya peningkatan jumlah puru di kebun kelapa sawit di Sumatra Utara, dari 11 pada 110 hari setelah pelepasan menjadi 79 pada 290 hari sesudah pelepasan. Keuntungan lain dari puru selain larva di dalamnya relatif terlindung dari musuh alami, puru juga mudah dikirim ke berbagai tempat bahkan diekspor ke luar negeri.
Lama Hidup dan Keperidian Lalat Puru C. connexa di Lapangan
Tabel 7 Parameter kehidupan imago lalat puru C. connexa di lapangan
Parameter kehidupan imago n hari Lama hidup imago betina 27 8 - 17
Lama hidup imago jantan 27 7 - 11
Masa praoviposisi 27 < 1
Masa oviposisi 27 8 - 16
Masa pascaoviposisi 27 1 - 2
Masa praoviposisi kurang dari 1 hari. Masa oviposisi 1- 16 hari dengan rata-rata 11,06 ± 2,868 hari. Puncak masa oviposisi terjadi pada hari ke-5 (Gambar 12). Pada hari ke-6 dan seterusnya, jumlah telur yang diletakkan semakin menurun dan berhenti pada hari ke-17. Puncak masa oviposisi ini berbeda dengan hasil penelitian Widayanti et al. (1999) yang menyebutkan bahwa puncak oviposisi terjadi pada hari ke-3. Hal ini terjadi karena pada penelitian ini lama hidup imago dan masa oviposisi lebih lama.
Berdasarkan pengamatan jumlah telur harian (Gambar 12) dapat diketahui pula persentase telur harian. Sampai dengan hari ke-5, telur yang diletakkan
0
ternyata telah mencapai 51,356% dari total telur yang diletakkan. Oleh karena itu, jika pelepasan lalat dilakukan pada stadia imago sebaiknya dilakukan segera (1-2 hari) sesudah imago muncul sebelum produksi telur menurun.
Rata-rata jumlah telur harian adalah 25,087 ± 13,368 butir telur/hari/betina. Keperidian lalat puru C. connexa adalah 140-329 butir telur/betina dengan rata-rata 219,167 ± 47,620 butir telur/betina. Keperidian hasil penelitian ini lebih tinggi daripada penelitian sebelumnya yang menyebutkan bahwa keperidian rata-rata lalat puru adalah 57,98 butir telur/betina (Widayanti et al. 1999).
Secara umum, lalat puru C. connexa sebagai agens hayati gulma kirinyuh yang diintroduksi dari Colombia telah dapat beradaptasi dengan kondisi lingkungan khususnya di Bogor. Setelah ± 10 tahun dilepaskan di lapangan, diketahui bahwa siklus hidup berjalan normal, lama hidup lebih lama dan keperidian lebih banyak di lapangan daripada di laboratorium. Kondisi ini sangat menguntungkan karena berarti kondisi di lapangan sangat mendukung keberhasilan reproduksi dan kehidupan internal lalat puru C. connexa. Hal ini berarti akan lebih banyak tanaman inang yang diserang dan berdampak baik untuk pengendalian gulma kirinyuh. Namun, yang perlu diwaspadai adalah serangan musuh alami terutama predator yang dapat menggangu pertumbuhan populasi lalat.
kompetisi yang kuat (Tjitrosoedirdjo 1989) sehingga dampak penyerangan lalat puru relatif kecil dibanding pertumbuhan kirinyuh.
Karakteristik tanaman kirinyuh yang lain adalah tanaman mati pada musim kemarau tetapi akarnya tetap hidup dan pada musim hujan tumbuh pesat. Menurut Tjitrosemito (1999), pengendalian akan efektif apabila dilakukan pada saat kirinyuh masih muda yang baru tumbuh pada awal musim hujan. Sementara pada saat itu, populasi lalat puru sangat rendah karena banyak yang mati pada saat kirinyuh mati pada musim kemarau. Kondisi ini dapat diatasi dengan cara inundasi. Pada musim kemarau disaat kirinyuh di lapangan mati, dapat dilakukan perbanyakan lalat puru di laboratorium. Pada awal musim hujan, disaat kirinyuh baru tumbuh segera dilakukan pelepasan lalat puru hasil perbanyakan di laboratorium.
Dengan melihat kemampuan lalat puru C. connexa yang belum efektif mengendalikan gulma kirinyuh, mungkin diperlukan kombinasi dengan agens hayati lain agar diperoleh hasil yang lebih maksimal. Menurut Chenon et al. (2002), komplek musuh alami (complex of natural enemy) akan memberikan hasil pengendalian yang lebih baik. Pemilihan agens hayati harus dilakukan dengan cermat dengan mempertimbangkan beberapa hal antara lain efektif mengendalikan gulma, mudah dikembangbiakan, ada tidaknya musuh alami di habitat baru, dan relung (nice) agens hayati. Agens hayati yang dipilih sebaiknya mempunyai relung yang berbeda dengan agens hayati sebelumnya agar tidak terjadi kompetisi. Seperti pengendalian hayati kirinyuh di Sumatra. Menurut Chenon et al. (2002), asosiasi antara P. pseudoinsulata dan C. connexa
Kisaran Inang Lalat Puru C. connexa
Hasil uji kisaran inang menunjukkan bahwa lalat puru C. connexa mampu melakukan infestasi dan oviposisi pada semua tanaman uji dengan tingkat yang berbeda. Pada tanaman kirinyuh (kontrol), lalat puru C. connexa tumbuh dan berkembang dengan baik sehingga mampu menyelesaikan tahap-tahap siklus hidupnya yaitu telur, larva, pupa dan imago (Tabel 8 dan Gambar 13a). Pada larva, dan bahkan dapat terbentuk puru walaupun lalat tidak dapat menyelesaikan siklus hidupnya (Tabel 8, Gambar 13b). Pada tanaman daun tanah dan babanjaran, lalat mampu meletakkan telur, telur menetas menjadi larva dan hidup dalam jaringan batang tetapi tidak terbentuk puru pada batang dan lalat tidak dapat menyelesaikan siklus hidupnya (Tabel 8, Gambar 13c dan 13d).
Pada Tabel 9 ditunjukkan tingkat infestasi dan jumlah telur C. connexa
pada tiga tanaman uji dan kontrol. Tingkat infestasi tertinggi terjadi pada tanaman kirinyuh (kontrol) yaitu 99,33%, kemudian daun tanah yaitu mencapai 83,33% dan tidak berbeda nyata dengan kontrol. Pada tanaman babadotan, tingkat infestasi mencapai 71,33% dan berbeda nyata dengan kontrol tetapi tidak berbeda nyata dengan daun tanah. Pada tanaman babanjaran, tingkat infestasi paling rendah yaitu mencapai 22,67% dan berbeda nyata dengan kontrol maupun babadotan dan daun tanah. Jumlah telur pada tanaman daun tanah dan babadotan, tidak berbeda nyata dengan kontrol, sedangkan pada babanjaran
Tabel 8 Perkembangan lalat puru C. connexa pada beberapa tanaman uji
No. Tanaman uji Telur Larva Pupa Imago Puru
(d) (b)
(c) (a)
Gambar 13 Tiga tanaman uji dan kontrol yang digunakan dalam uji kisaran inang: (a) kirinyuh (kontrol), (b) babadotan, (c) daun tanah, (d) babanjaran. Selain pada kontrol, puru juga terbentuk pada tanaman babadotan
sangat sedikit dan berbeda nyata dengan kontrol, babadotan, maupun daun tanah.
Tabel 9 Tingkat infestasi dan jumlah telur C. connexa pada tiga tanaman uji dan
Huruf yang sama pada satu kolom menunjukkan tidak berbeda nyata (BNT, α = 5%).
dan babadotan (Chenon et al. 2002). Tanaman babanjaran tidak menarik C.
connexa untuk meletakkan telur. Hal ini diduga karena batangnya keras dan
permukaannya kasar sehingga menyulitkan lalat betina untuk meletakkan telur. Menurut Hedin et al. (1977) morfologi atau struktur tanaman dapat mempengaruhi serangga dalam memilih tanaman sebagai inang, tempat berlindung, atau tempat meletakkan telur.
Telur yang diletakkan dalam jaringan pucuk tanaman uji menetas menjadi larva dalam waktu ± satu minggu. Walaupun jumlah telur yang diletakkan pada tanaman daun tanah dan babadotan jumlahnya hampir sama dengan kontrol, jumlah larva yang ditemukan lebih sedikit daripada kontrol (Gambar 14). Hal ini berarti banyaktelur pada kedua tanaman uji yang mati. Larva pada tanaman uji hanya mampu hidup sampai dengan minggu ke-2. Pada minggu ke-3 semua larva telah mati. Hal ini diduga karena nutrisi tanaman uji tidak cocok untuk pertumbuhan lalat puru C. connexa. Selain itu diduga pada tanaman uji terdapat allelokimia tertentu yang bersifat toksik dan mematikan larva. Menurut Tabashnik dan Slansky (1987), allelokimia dapat bersifat toksik pada herbivor yang tidak beradaptasi.
Puru pada kirinyuh (kontrol) maupun babadotan mulai terbentuk pada minggu ke-2 setelah oviposisi. Pada babadotan, infestasi larva lalat puru
C. connexa mampu menstimulasi tanaman untuk membentuk puru, walaupun
0
Kirinyuh (kontrol) Babadotan Daun tanah Babanjaran
Gambar 14 Jumlah larva/pucuk lalat puru C. connexa pada tiga tanaman uji dan kontrol
dan Drees (2006), puru terbentuk ketika tanaman bereaksi terhadap stimuli dari serangga. Stimulus dapat berupa keberadaan dan aktivitas serangga di dalam jaringan tanaman. Pada dua tanaman uji yang lain, yaitu daun tanah dan babanjaran puru tidak terbentuk, walaupun terdapat larva di dalam jaringan batang. Hal ini menunjukkan bahwa sejumlah larva yang terdapat dalam jaringan batang belum dapat menstimulasi pembentukan puru.
Diameter dan panjang puru pada babadotan lebih kecil dari puru pada kontrol (Gambar 15). Pada minggu ke-4 dan seterusnya, diameter dan panjang puru pada babadotan relatif tetap, sedangkan pada kirinyuh masih berkembang terus. Perbedaan ukuran puru disebabkan oleh ukuran batang tanaman babadotan yang lebih kecil dibandingkan dengan batang kirinyuh. Pada minggu ke-4, perkembangan batang babadotan telah mencapai maksimal sedangkan pada kirinyuh masih berkembang terus.
0
Gambar 15 Perkembangan diameter dan panjang puru pada kirinyuh dan babadotan
akan merugikan pertumbuhan populasi lalat karena terjadi pemborosan telur. Telur-telur yang seharusnya berkembang sampai dewasa akan mati. Tetapi hal ini hanya terjadi dalam keadaan terpaksa, pada saat tidak ada kirinyuh. Hasil penelitian Sipayung dan Chenon (1995) juga menemukan bahwa pada uji dengan pilihan, tidak terdapat telur pada tanaman lain selain kirinyuh.
Berdasarkan hasil-hasil penelitian ini diketahui bahwa lalat puru
C. connexa yang diintroduksi dari Colombia sangat spesifik inang dan hanya
Kesimpulan
Perkembangan pradewasa lalat puru C. connexa berada di dalam puru dan imago hidup bebas. Siklus hidup lalat puru C. connexa 56-77 hari. Mortalitas banyak terjadi pada stadia larva yang disebabkan oleh musuh alami terutama predator. Tidak ada korelasi antara ukuran puru dengan jumlah larva.
Terdapat empat instar pada perkembangan larva C. connexa. Mandibel dan sklerit hipofaring dapat digunakan sebagai indikator instar pada perkembangan larva C. connexa. Indikator instar yang paling baik adalah mandibel, karena selain ukurannya berbeda terdapat perubahan bentuk mandibel. Volume larva tidak dapat digunakan sebagai penduga instar pada perkembangan larva C. connexa.
Lama hidup yang lebih lama dan keperidian yang lebih banyak di lapangan sangat menguntungkan karena berarti kondisi di lapangan sangat mendukung keberhasilan reproduksi dan kehidupan lalat puru C. connexa. Hal ini berarti akan lebih banyak tanaman inang yang diserang dan berdampak baik untuk pengendalian gulma kirinyuh.
Lalat puru C. connexa telah berhasil mapan khususnya di Bogor, tetapi belum efisien mengendalikan gulma kirinyuh. Komplek musuh alami diharapkan akan memberikan hasil yang lebih baik bagi pengendalian hayati kirinyuh.
Pada tanaman babadotan dan daun tanah, lalat puru mampu meletakkan telur dan telur menetas menjadi larva tetapi larva hanya bertahan hidup selama 2 minggu. Pada tanaman babadotan, lalat puru C. connexa bahkan mampu menstimulasi tanaman untuk membentuk puru, walaupun tidak dapat menyelesaikan hidupnya.
Saran
DAFTAR PUSTAKA
Alencar YB, Hamada N, Magni-Darwich S. 2001. Morphometric comparison of
Simulium perflavum larvae (Diptera: Simuliidae) in relation to season and
gender in Central Amazonia, Brazil. [terhubung berkala] http://www.ncbi. nlm.nih.gov/entrez/query.fcgi?cmd= Retrieve&db=Pu [16 Juni 2006].
[Anonim]. 2006. Ecological relationship among goldenrod plant and gall insects. [terhubung berkala]. http://www.oswego.edu/wscp/GOLDGALL. html [23 Mei 2006].
Bennet DM, Hoffmann AA. 1998. Effect of size and fluctuating asymmetry on field fitness of parasitoid Trichogramma carverae (Hymenoptera: Trichogrammatidae). Ann Ecol 67:580-591. http://www.life.bio.sunysb. edu/ morph/ morph.html.
Binggeli P. 1997 Chromolaena odorata (L.) King & Robinson (Asteraceae). [terhubung berkala]. http://www.members.lycos.co.uk/ WoodyPlantEcology/ docs/web-sp4.htm. [13 Oktober 2003].
Borror DJ, Triphlehorn CA, Johnson NF. 1992. An Introduction to the Study of
Insects. 6th edition. New York: Saunders College Publishing.
Chenon RD, Sipayung A, Sudharto P. 2002. A decade of biological control against Chromolaena odorata at the Indonesian Oil Palm Research Institute in Marihat. Di dalam: Zachariades C, Muniapan R, Strathie LW, editor. Proceedings of the 5th International Workshop on Biological
Control and Management of Chromolaena odorata; Durban, South Africa,
23-25 October 2000. ARC-PPRI. hlm 46-52.
Cock MJW. 1984. Possibilities for biological control of Chromolaena odorata.
Tropical forest management 30: 7-13.
Cruttwell RE. 1974. The insects and mites attacking Eupatorium odoratum in the Neotropics. Tech Bull Commonwealth Institute of Biological Control 17: 87-125.
Dorchin N, Freidberg A, Aloni R. 2002. Morphogenesis of stem gall tissues induced by larvae of two cecidomyiid species (Diptera: Cecidomyiidae)
on Suaeda monoica (Chenopodiaceae) [abstrak]