TRANSFORMASI GENETIK PADI
INDICA
DENGAN GEN
OsMADS18
MENGGUNAKAN
Agrobacterium tumefaciens
UNTUK MERAKIT
VARIETAS PADI UMUR PENDEK
MUJIBUR RAHMAN
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ABSTRAK
MUJIBUR RAHMAN. Transformasi Genetik Padi
Indica
dengan Gen
OsMADS18
Menggunakan
Agrobacterium tumefaciens
untuk Merakit Varietas
Padi Umur Pendek. Dibimbing oleh I MADE ARTIKA.
Padi adalah makanan pokok lebih dari setengah populasi manusia di dunia.
Kebutuhan padi setiap tahunnya terus meningkat seiring bertambahnya penduduk.
Penelitian ini bertujuan untuk merakit varietas padi berumur pendek dengan
mengintroduksikan gen
OsMADS18
ke dalam genom tanaman menggunakan
Agrobacterium tumefaciens
. Gen
OsMADS18
merupakan gen umur pendek yang
berhasil dikonstruksi ke vektor pCAMBIA 1301 di bawah promotor konstitutif
CaMV35S. Hal ini dibuktikan dengan munculnya pita DNA dengan ukuran 12561
bp, ukuran ini merupakan gabungan antara vektor pCAMBIA 1301 (11819 bp)
dan gen
OsMADS18
(742 bp). Kemudian vektor pCAMBIA 1301-
OsMADS18
ditransformasikan ke
Agrobacterium tumefaciens EHA105
melalui metode
Heat-shock
. Keberhasilan transformasi
Agrobacterium tumefaciens
ditandai dengan
tumbuhnya koloni putih pada media LB padat yang telah diberikan antibiotik,
X-gal, dan IPTG. Kemudian bakteri tersebut digunakan untuk mentransformasikan
50 kalus padi
Indica
dengan cara merendam kalus tersebut ke dalam koloni
bakteri selama 20 menit. Kalus di pindahkan ke media seleksi yang mengandung
higromisin. Setelah 14 hari, kalus yang tidak mengalami nekrotik di pindahkan ke
media regenerasi R3B untuk ditumbuhkan sebagai tanaman transgenik. Dari
penelitian diperoleh 18 planlet transforman
Indica.
Keberhasilan uji
gus-A
di
tunjukkan dengan munculnya warna biru pada jaringan daun. Sedangkan analisis
PCR terhadap 18 planlet transgenik menghasilkan pita berukuran 742 bp.
ABSTRACT
MUJIBUR RAHMAN. Genetic Transformation of
Indica
rice by
OsMADS18
gene using
Agrobacterium tumefaciens
to assemble short life plant.
Supervised by I MADE ARTIKA.
Rice is the staple food of more than half the world's human population. The
need of rice each year continues to increase with increasing population. This study
aimed to assemble a short-lived rice varieties by introducing
OsMADS18
gene
into the genome of plants using
Agrobacterium tumefaciens
.
OsMADS18
gene is a
short age gene which has been successfully constructed to pCAMBIA1301 vector
under CaMV35S constitutive promoter. This is evidenced by the emergence of
12561 bp DNA band size, this measure is a combination among vector pCAMBIA
1301 (11819 bp) and
OsMADS18
genes (742 bp). Then the vector pCAMBIA
1301 that contains
OsMADS18
, was used to transform
Agrobacterium tumefaciens
strain EHA105 through Heat-shock method. The success of transformation
marked by the growth of the white colonies
Agrobacterium tumefaciens
on solid
media LB, this media contained antibiotic, X-gal and IPTG. Then the bacteria are
used to transforms 50 calli Indica rice by soaking the calli into colonies for 20
minutes. The calli moved to selection media that contains hygromycin. After 14
days, calli that resistant to hygromycin moved to regeneration media R3B grown
for transgenic plant. 18 transformants plantlets were obtained.
Gus-A
successful
test on the show with the appearance of blue color in leaf tissues. While PCR
analysis of the 18 transgenic plantlets produced 742 bp sized bands.
TRANSFORMASI GENETIKA PADI INDICA DENGAN GEN
OsMADS18
MENGGUNAKAN
Agrobacterium tumefaciens
UNTUK MERAKIT
VARIETAS PADI UMUR PENDEK
MUJIBUR RAHMAN
Skripsi
Sebagai salah satu syarat memperoleh gelar
Sarjana Sains pada
Departemen Biokimia
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
Judul Skripsi
: Transformasi Genetika Padi
Indica
dengan Gen
OsMADS18
Menggunakan
Agrobacterium Tumefaciens
untuk Merakit
Varietas Padi Umur Pendek
Nama
: Mujibur Rahman
NIM
: G84070020
Disetujui,
Dr. I Made Artika, M.App.Sc
Pembimbing
Diketahui,
Dr. I Made Artika, M.App.Sc
Ketua Departemen Biokimia
\
KATA PENGANTAR
Puji syukur penulis ucapkan kehadirat Tuhan Yang Maha Esa atas segala
berkah dan rahmat-Nya sehingga penulis dapat menyelesaikan karya ilmiah yang
berjudul “Transformasi Genetika Padi
Indica
dengan Gen
OsMADS18
Menggunakan
Agrobacterium Tumefaciens
untuk Merakit Varietas Padi Umur
Pendek”. Karya ilmiah ini
disusun berdasarkan kegiatan penelitian yang dilakukan
dari bulan Februari 2011 hingga bulan Mei 2012, di Laboratorium Kultur
Jaringan, Departemen Agronomi dan Hortikultura, Fakultas Pertanian, IPB.
Penulis mengucapkan terima kasih kepada Dr. I Made Artika M.App.Sc
selaku pembimbing yang telah memberikan bimbingan, masukan, dan arahan
selama penulisan karya ilmiah ini, Direktorat Jenderal Pendidikan Tinggi dan
Kementerian Riset dan Teknologi RI yang telah mendanai penelitian ini. Selain
itu penulis juga mengucapkan terima kasih kepada Indra Kurniawan S, Harryade
Putra, Ridho Pratama, Putri Jumiarti, Cinthya Lestari HD dan Dian Rachmawati
atas bantuan teknis, teoritis maupun materil selama pengerjaan penelitian ini.
Penulis juga mengucapkan terima kasih kepada orang tua dan seluruh keluarga
yang telah mendukung secara moril, dan juga kepada semua pihak yang telah
membantu terselesaikannya skripsi ini.
Penulis menyadari bahwa tulisan ini belum sempurna sehingga saran dan
kritik yang membangun sangat penulis harapkan. Akhirnya penulis berharap
semoga karya ilmiah ini dapat dimanfaatkan bagi kemajuan pengetahuan,
pendidikan, dan penelitian.
Bogor, Februari 2013
RIWAYAT HIDUP
Penulis dilahirkan di Aceh Utara pada tanggal 16 Februari 1989 sebagai
putra kedua dari lima bersaudara pasangan Bapak Zainal Abidin (alm) dan Ibu
Nurul Hidayat. Penulis tinggal dan dibesarkan di Krueng Geukuh dan sekolah di
SD 06 Kec. Dewantara, pada tahun 1998 penulis pindah dan menetap di kota
Banda Aceh dan pada tahun 2001 penulis lulus dari SD 105 Lam Ujong, Ulee
Kareng dan melanjutkan pendidikan ke SMP 10 Lam Teh. Setelah lulus pada
tahun 2004, penulis melanjutkan sekolah ke SMA 4 Negeri Banda Aceh dan
Lulus pada tahun 2007. Pada tahun yang sama penulis diterima di Institut
Pertanian Bogor pada Mayor Biokimia, Departemen Biokimia, Fakultas
Matematika dan Ilmu Pengetahuan Alam IPB, melalui jalur Undangan Seleksi
Masuk IPB (USMI).
Selama duduk di bangku perkuliahan, penulis aktif di berbagai organisasi
kemahasiswaan. Tahun 2007-2008 penulis aktif sebagai Ketua ROHIS A18.
Selain itu penulis juga aktif di Organisasi Mahasiswa Daerah (OMDA) Ikatan
Mahasiswa Tanah Rencong (IMTR). Pada tahun 2009-2010 penulis aktif sebagai
pengurus dan kepanitian Himpunan Keprofesian CREB’s, BEM
-G, dan beberapa
Unit Kegiatan mahasiswa (UKM) di lingkup IPB. Diantaranya pernah menjabat
sebagai Ketua Divisi Keilmuan Metabolisme CREB’s pada tahun 2009/2010,
Ketua Pelaksana PEMIRA dan MUBES CREB’s 2010,
Kepanitian Kajian Ilmiah
CREB’s, LKIP, BCL 2010
(Biochemistry Champion League),
MPD BIOKIMIA
45,
The CEBOCH
,
anggota BP CREB’s 2011
, dan berbagai kegiatan lainnya.
Tahun 2011-2012 penulis pernah menjadi finalis BIOFRONT ITB, finalis TUN
Mipa Untuk Negeri UI 2012, mendapatkan hibah dana penelitian dari DIKTI
melalui Program Kreativitas Mahasiswa (PKM-P dan PKM-GT), mendapatkan
penghargaan setara emas di Penunjang PIMNAS XXIV di Universitas
Hasanuddin, Makassar 2011. Finalis PIMNAS XXV dan mendapatkan mendali
setara emas di Universitas Muhammadiyah Yogyakarta 2012. Selain itu penulis
juga berkesempatan menjadi peserta dalam
4th World Congress of Industrial
Biotechnology
China 2011, dan peserta dalam
annual Meeting of the German
society for Genetic Engineering
Jerman 2011.
DAFTAR ISI
DAFTAR GAMBAR ………
DAFTAR LAMPIRAN ………
PENDAHULUAN
………
.
TINJAUAN PUSTAKA
………
Padi
………
MADS Box
………
Transformasi Genetik
………
Konstruksi DNA Rekombinan
……….….
Vektor
………
Plasmid pCAMBIA 1301
……….
Agrobacterium tumefaciens
………..………
Analisis Tanaman Transgenik
………..
BAHAN DAN METODE
………
..
Alat dan Bahan
……….
Metode
………..
HASIL DAN PEMBAHASAN
……….
Plasmid rekombinan pembawa Gen
OsMADS18
……….………
Agrobacterium tumefaciens
Pembawa vektor pCAMBIA 1301-
OsMADS18...
Tranformasi dan Regenerasi Tanaman
……….
Tanaman Transgenik
………
KESIMPULAN
……….
SARAN
……….
DAFTAR PUSTAKA
………
DAFTAR GAMBAR
1.
Elektroforegram DNA pGEM-T Easy-
OsMADS18
….
………. ……...
2.
Elektroforegram pemotongan pCAMBIA 1301 dengan enzim restriksi
….……
3.
Koloni
Agrobacterium tumefaciesn EHA 105
hasil transformasi ………
4.
Transformasi genetik padi
Indica
……….
5.
Hasil uji
β
-glucuronidase
terhadap daun tanaman padi ………..
6.
Elektroforegram PCR terhadap gen
OsMADS18
………
DAFTAR LAMPIRAN
1.
Diagram alir penelitian ………
2.
Proses umum isolasi plasmid ………
3.
Peta Plasmid pCAMBIA 1301 ……….
4.
Proses umum reaksi ligasi ………
5.
Proses purifikasi DNA dengan metode dari QiAgen (2006) ………
6.
Proses umum isolasi DNA tanaman ……….
7.
Bahan-
bahan pembuatan media ………
...
Halaman
16
17
18
18
19
20
21
Halaman
1
PENDAHULUAN
Padi adalah makanan pokok lebih dari setengah populasi di dunia. Sebagian besar tanaman padi di produksi dan dikonsumsi di Asia (Kibria et al. 2008). Padi memegang peranan paling penting di antara berbagai sumber bahan pangan lainnya di Indonesia, terutama dalam penyediaan pangan yang mendukung ke arah ketahanan pangan nasional dan pemberdayaan ekonomi rumah tangga petani. Mutu suatu varietas sangat mempengaruhi besarnya pendapatan petani (Lestari et al. 2007). Kebutuhan pangan dunia setiap tahunnya semakin meningkat seiring dengan bertambahnya jumlah penduduk dan perkembangan industri pangan. Menurut data dari BPS (2012), produksi gabah pada tahun 2012 sebesar 65,39 juta ton. Namun, jumlah ini belum mampu untuk memenuhi kebutuhan lebih dari 259 juta penduduk Indonesia. Maka diperlukan suatu inovasi untuk memenuhi permintaan konsumen tersebut, salah satunya dengan melakukan pemuliaan tanaman (Herman 2010).
Pemuliaan tanaman merupakan suatu metode yang menggali potensi genetik suatu tanaman untuk memaksimumkan ekspresi dari potensi tersebut pada suatu kondisi lingkungan tertentu (Wangxia 2003, Joseph et al. 2011). Salah satunya dengan merakit varietas padi berumur pendek. Pemulian tanaman bisa dilakukan melalui tranformasi genetik, yaitu dengan menyisipkan gen asing ke dalam genom tanaman ( Heryati 2002). Transformasi genetik dapat dilakukan melalui berbagai metode antara lain elektroporasi (Widodo 2002), polyethylene glycol (Rachmawati 2010), penggunakan serat silicon carbide (Herman 2010), penembakan partikel DNA (Frame et al. 2000, Sutrisno et al. 2000) dan perantara Agrobacterium tumefaciens (Belarmino dan Mii 2000, Joseph et al. 2011). Teknik penyisipan gen dengan menggunakan metode tersebut sudah berhasil dilakukan (Schroeder et al. 2000, Chuanfeng et al. 2010). Penggunaan Agrobacterium untuk proses transformasi genetik pada tanaman monokotil seperti padi, masih belum efisien, akan tetapi beberapa penelitian telah menunjukkan keberhasilan (Cheng et al. 2004, Rachmawati et al. 2004, Rajesh et al. 2008)
Tantangan lain dari teknik transformasi genetik adalah penggunaan promotor yang tidak sesuai, sehingga ekspresi gen yang diinginkan tidak terjadi (Yina et al. 2004). Selain itu teknik penyisipan gen pada tanaman padi dengan Agrobacterium tumefaciens
kurang efektif (Enriquez-Obregon et al. 2000, Opabode 2006). Hal ini disebabkan tidak efektifnya infeksi Agrobacterium tumefaciens pada tanaman monokotil karena tidak adanya respon pelukaan yang menghasilkan senyawa fenolik yang digunakan oleh bakteri untuk menginduksi gen-gen vir (Lucca et al. 2001, Gao et al. 2005), akan tetapi masalah ini bisa diatasi dengan melakukan penyesuaian kondisi, yaitu dengan menambahkan senyawa fenolik, monosakarida, dan penggunaan vektor ekspresi seperti pCAMBIA (Zupan et al. 2000, Cheng et al. 2004). Agrobacterium tumefaciens dapat menginfeksi tanaman secara alami karena memiliki plasmid Ti
(tumor inducer)
, yaitu vektor (pembawa DNA) untuk menyisipkan gen asing (Madigan dan Martinko 2006). Gen asing yang ingin dimasukkan ke dalam tanaman dapat disisipkan di dalam plasmid Ti tersebut. Selanjutnya, Agrobacterium tumefaciens secara langsung akan memindahkan gen pada plasmid tersebut ke dalam genom tanaman (Olhoft et al. 2004).Teknik tranformasi genetik yang bertujuan untuk membuat varietas padi berumur pendek mulai dikembangkan di Indonesia. Salah satu gen yang membawa sifat umur pendek adalah gen OsMADS18 (Masiero et al. 2002). Gen OsMADS18 merupakan gen yang memiliki sifat dalam mempercepat masa transisi tanaman sehingga tanaman dapat berbunga lebih awal (Xiao et al. 2003). Selain itu gen ini juga berfungsi dalam proses perkembangan tanaman, seperti dalam proses pengendalian masa transisi reproduksi vegetatif pada tanaman jagung (Pelucchi et al. 2002), penentu identitas organ pada Antirrhinum (Muller et al. 2001), dan pengaturan pematangan buah (Masiero et al. 2002, Favoro et al. 2003, Xiao et al. 2003).
2
TINJAUAN PUSTAKA
Padi
Padi (Oryza sativa L.) merupakan tanaman pangan terpenting pada wilayah tropis maupun sub tropis. Upaya peningkatan produksi padi merupakan salah satu strategi untuk menghadapi peningkatan kebutuhan pangan akibat pertambahan populasi penduduk (KOMPAS 2011). Selain itu padi juga dapat digunakan sebagai organisme model dalam kajian genetika tumbuhan karena memiliki ukuran kromosom yang relatif kecil, yaitu 1.6~2.3 × 108 pasangan basa (Garris 2004). Dari hasil kajian biologi molekuler padi di bagi ke dalam dua subspesies utama, yaitu indica dan japonica . Selain itu juga terdapat subspesies minor yang beradaptasi berdasarkan ekosistemnya seperti aus, royada, ashina, dan aromatic (Zohary dan Hopf 2000) Regenerasi tanaman dari protoplast padi indica lebih sulit dibandingkan japonica. Untuk memecahkan masalah ini Christou et al. (1991) melakukan bombardment terhadap embrio muda padi indica menggunakan emas pembawa DNA. Tanaman transgenik padi indica terbukti dapat dihasilkan dari kalus hasil transformasi, hal ini membuka peluang pengembangan sistem transformasi padi indica tanpa kultur protoplas. Dalam perkembangan selanjutnya, Datta et al. (1992) berhasil melakukan transformasi pada protoplas padi kelompok indica kultivar IR72.
Regenerasi tanaman dari protoplas beberapa varietas padi indica telah dilaporkan. Datta et al. (1990) mengembangkan teknik regenerasi dari protoplas padi indica Chinsurah Boro dan mentransformasikan gen hygromycin phosphotransferase glycol. Regenerasi tanaman dari protoplas IR54 telah dilaporkan peng et al. (1992) dan diperoleh tanaman dengan gen neomycin phosphotransferase. Hal ini membuktikan transformasi menggunakan protoplas dapat dilakukan pada padi indica, akan tetapi kelemahan dari teknik ini adalah terbatasnya keberhasilan pada kultivar tertentu.
MADS Box
MADS Box merupakan protein faktor trankripsi yang mengendalikan perkembangan tanaman (Masiero et al. 2002). Hasil beberapa penelitian terdahulu menunjukkan bahwa gen homolog MADs Box yang di isolasi dari spesies yang berbeda memiliki fungsi yang sama (Xiao et al. 2003). Misalnya OsMADS18 yang diisolasi dari padi (Oryza sativa) memiliki fungsi dalam perkembangan
tanaman seperti dalam proses pengendalian masa transisi reproduksi vegetatif pada tanaman jagung (Pelucchi et al. 2002), selain itu gen ini juga menjadi penentuan identitas organ, dan pengaturan pematangan buah (Favoro et al. 2003). Gen OsMADS18 banyak terakumulasi pada meristem, ekspresi gen ini lebih awal dapat menentukan faktor histologi yang menunjukkan pembentukan tunas aksiler meristem akan dipercepat (Ferrandiz et al. 2000). OsMADS18 merupakan keluarga dari MADS Box, gen inilah yang menyandi ekspresi umur genjah pada berbagai jenis tanaman (Pelucchi et al. 2002).
Transformasi Genetik
Transformasi genetik merupakan salah satu metode yang dapat dimanfaatkan untuk mempelajari regulasi gen, identifikasi fungsi gen, pengujian metabolisme, mempelajari fisiologi serta perkembangan tanaman (Walkerpeach dan Velten 1994, Utomo 2005). Transformasi genetik tidak hanya dipergunakan untuk studi dasar dalam ilmu tanaman, tetapi juga telah menjadi salah satu cara untuk pengembangan varietas baru. Keberhasilan proses transformasi gen melalui Agrobacterium tumefaciens sangat ditentukan oleh berbagai hal antara lain kesesuaian antara strain Agrobacterium tumefaciens dengan jenis tanaman dan plasmid vektor yang dipergunakan, kerapatan sel Agrobacterium yang digunakan pada saat proses tranformasi genetik, lama waktu ko-kultivasi, tingkat kemasaman media, kondisi kultur in vitro dan sebagainya (Zupan et al. 2000, Cheng et al. 2004). Disamping itu pemanfaatan Agrobacterium tumefaciens pada proses transformasi genetik jenis tanaman monokotil masih memerlukan berbagai penyesuaian dalam upaya meningkatkan efisiensi transformasi (Klee 2000, Utomo 2004). Hal ini disebabkan secara alami bakteri patogen tanah tersebut hanya menginfeksi tanaman dikotil dengan cara mengintroduksi T-DNA dari plasmid Ti bakteri ke dalam inti sel tanaman (Le et al. 2001, Olhoft et al. 2004).
Konstruksi DNA Rekombinan
3
intinya adalah proses kloning gen. Gen merupakan sekuen nukleotida yang menyandi RNA yang dibatasi oleh promotor dan terminator (Watson et al. 2007). Teknologi DNA rekombinan memungkinkan sejumlah gen dari sumber berbeda disatukan untuk membentuk DNA rekombinan. Tahapan dalam kloning gen meliputi: penyisipan fragmen DNA yang mengandung gen target ke dalam molekul DNA vektor, vektor rekombinan dimasukkan kedalam sel inang, vektor dalam sel inang akan diperbanyak seiring dengan pembelahan sel inang (Pray 2008).
Komponen penting dalam rekombinasi DNA adalah enzim-enzim manipulasi DNA serta vektor. Bernard and Pasternak (2002) membagi enzim manipulasi berdasarkan jenis dan reaksi yang dikatalisnya menjadi lima golongan yaitu : a) nuklease yaitu enzim yang mampu memotong molekul asam nukleat, b) ligase adalah enzim yang berfungsi menyatukan molekul asam nukleat, c) polimerase adalah enzim yang dapat mensintesis DNA, d) enzim modifikasi yang mampu menghilangkan atau menambahkan gugus kimia, dan e) topoisomerase adalah enzim yang mengubah DNA tertutup secara kovalen menjadi DNA supercoil. Pada pengklonan gen hanya dua jenis enzim yang berperan yaitu enzim restriksi endonuklease dan enzim ligase.
Vektor
Vektor kloning adalah agen pembawa fragmen DNA masuk ke dalam sel makhluk hidup yang berfungsi untuk memperbanyak fragmen DNA. Beberapa vektor kloning yang umum digunakan adalah plasmid, vektor lamda, virus, kromosom bakteri buatan, kromosom khamir buatan, dan cosmid (Primrose dan Twyman 2006). Suatu vektor kloning harus dapat disambungkan atau menyatu dengan fragmen DNA yang ingin ditransfer kemudian dapat dimasukkan ke dalam sel. Di dalam sel tersebut, fragmen DNA akan diperbanyak jumlahnya. Untuk memilih vektor kloning yang cocok dalam penelitian, beberapa hal yang harus dipertimbangkan adalah ukuran fragmen DNA yang akan ditransfer, jumlah salinan, inkompatibilitas (ketidaksesuaian), marka pemilihan (selectable marker) untuk seleksi, situs kloning, dan fungsi khusus dari suatu vektor (Casali et al. 2003). Dalam penelitian ini vektor yang digunakan adalah pGEM-T Easy, vektor ini mempunyai multiple cloning site, serta tidak perlu dipotong lagi ketika
digunakan. Sedangkan vektor ekspresi yang digunakan adalah pCAMBIA 1301.
Plasmid pCAMBIA 1301
DNA vektor adalah molekul DNA yang dapat bereplikasi secara mandiri dan dapat digunakan sebagai pembawa molekul DNA lain yang tidak memiliki kemampuan bereplikasi sendiri di dalam sel. DNA yang sering digunakan adalah plasmid bakteri. Plasmid adalah bahan genetik ekstrakromosom yang diwariskan secara tetap (Dworkin et al. 2006). Ciri-ciri plasmid antara lain berukuran kecil dan hanya mengandung beberapa gen, membawa informasi genetik, terlepas dari DNA kromosom atau kadang-kadang dapat berintegrasi dengan DNA kromosom, dan dapat diisolasi dengan mudah dari sel bakteri. Ciri lain plasmid adalah mempunyai situs untuk memulai replikasi yang disebut ORI (origin of replication) (Tsen et al. 2002)
4
Plasmid pCAMBIA 1301 (Lampiran 3) mengandung T-DNA, yang merupakan untaian DNA yang akan ditransfer ke sel tanaman. T-DNA pCAMBIA 1301 memiliki gen neomisin fosfotranferase II (nptII) dan gen higromisin fosfotransferase (hpt). Gen npt II menyandi enzim neomisin fosfotranferase yang digunakan untuk menyeleksi bakteri. Gen hpt untuk menyandikan enzim higromisis fosfotransferase sebagai penyeleksi untuk mengetahui telah terintegrasinya T-DNA dari A.tumefaciens ke dalam genom tanaman (Dobhal et al. 2010). Plasmid pCAMBIA 1301 juga mempunyai gen penanda yaitu Gus-A yang disisipkan dibagian yang dapat menyandikan, sehingga ekspresi gen hanya mungkin terjadi bila sudah terintegrasi pada tanaman (Ghasemi et al. 2008).
Agrobacterium tumaficiens
Agrobacterium tumefaciens merupakan bakteri tanah gram positif yang bersifat fitopatogen pada tanaman dikotil. Bakteri ini secara alami mempunyai kemampuan untuk mentransfer potongan DNA atau disebut T-DNA (transfer DNA) ke dalam genom tanaman dan menyebabkan terbentuknya tumor (crown gall). Tumor ini merupakan sumber makanan atau sebagai sumber karbon bagi Agrobacterium (Rajesh et al. 2008). Terdapat tiga komponen genetik penting terlibat dalam proses pembentukan tumor (Tzfira et al. 2004). Pertama, gen virulen kromosom (chromosomal virulence), yang terdapat pada kromosom Agrobacterium dan berfungsi dalam pelekatan bakteri dengan sel tanaman. Kedua, sekelompok gen virulen (vir) yang terdapat dalam plasmid Ti (Tumor inducing), berukuran 200 kb yang berperan dalam menginduksi transfer dan integrasi T-DNA. Ketiga, daerah T-DNA yang juga terletak pada plasmid Ti (Daerah T-DNA, dibatasi oleh LB (left border) dan RB (right border), mengandung gen penting bagi Agrobacterium (Matthews et al. 2001, Zhou et al. 2003, Sha et al. 2004). Daerah T-DNA mengandung gen iaaH, iaaM, dan ipt yang menyandikan enzim-enzim penting dalam biosintesis auksin dan sitokinin, yaitu zat penting untuk pembelahan sel sehingga terjadi pembelahan sel yang tidak terkontrol dan menyebabkan terbentuknya tumor (Azhakanandam et al. 2000).
Analisis Tanaman Transgenik
Tujuan utama dalam proses transformasi genetik adalah mendapatkan ekspresi dari gen-gen asing yang diintroduksikan ke dalam sel
atau jaringan tanaman target. Proses transfer gen dalam tanaman tidakk menjamin bahwa gen asing yang diintroduksikan akan selalu terekspresi. Oleh karena itu diperlukan serangkaian analisis untuk memastikan keberadaan gen yang telah diintroduksikan ke genom tanaman.
Analisis tanaman transgenik dalam proses transformasi genetik tanaman dapat dilakukan pada berbagai tahapan, yaitu; 1) introduksi gen, 2) transkripsi, 3) translasi, 4) pengujian efektifitas protein yang dihasilkan. Saat ini telah dikembangkan berbagai teknik analisis tanapan trasgenik, seperti uji histokimia b-glucuronidase (uji Gus), analisis polymerase chain reaction (PCR), analisis Southern Blot, Nothern, Western, Elysa, dan lainnya.
1.Uji Gus (Glucuronidase)
Uji histokimia β-glucuronidase (uji Gus) dapat dilakukan pada tahap awal setelah ko-kultivasi maupun setelah gen terintegrasi dengan stabil dalam kromosom. Warna biru yang muncul pada sel atau jaringan menunjukkan hasil transformasi positif. Warna biru disebabkan oleh reaksi substrat X-gluc (5-bromo-4-chromo-3-indolyl-B-D-glucuronide) dengan enzim β-glucuronidase menjadi suatu senyawa perantara yang kemudian melalui reaksi dimerisasi oksidatif membentuk senyawa dichloro-dibromoindigo (CIBr-Indigo) yang berwarna biru (Stomp 1992). Warna biru tersebut menunjukkan telah terekspresinya gen gus-A.
5
2.Analisis PCR (Polmerase Chain Reaction)
Teknik PCR merupakan salah satu komponen utama teknologi DNA rekombinan dan pertama kali ditemukan oleh Kary B. Mulis pada tahun 1983. Teknik ini merupakan amplifikasi in vitro sejumlah sekuens DNA dengan menggunakan 2 oligonukleotida sebagai primer yang berhibridisasi secara berlawanan pada sisi daerah target DNA yang diinginkan. PCR mampu mengamplifikasi secara selektif sekuens DNA spesifik, hal inilah yang membuat penggunaan teknik PCR semakin meluas disamping kesederhanaan metodenya (Dieffenbach & Dveksler 2003).
Reaksi PCR adalah tiruan dari proses replikasi DNA, penempatan primer, dan perpanjanagan rantai DNA baru oleh DNA
polimerase dari arah 5’ ke 3’, hanya saja pada
teknik PCR tidak menggunakan enzim ligase dan primerRNA. Secara ringkas, teknik PCR dilakukan dengan cara mencampurkan sampel DNA dengan primer oligonukleotida, deoksiribonukleotida trifosfat, enzim termostabil Taq DNA polimerase dalam larutan yang sesuai, kemudian menaikkan dan menurunkan suhu campuran secara berulang selama beberapa jam sampai diperoleh jumlah sekuen DNA yang diinginkan.
Satu siklus pada teknik PCR terdiri atas tiga tahap, yaitu denaturasi, annealing dan ekstensi. Denaturasi dilakukan pada suhu 90-950C, sehingga terjadi pemisahan utas ganda DNA menjadi dua utas tunggal DNA yang menjadi cetakan (template) tempat penempelan primer dan tempat kerja DNA polimerase. Selanjutnya suhu diturunkan untuk penempelan primer oligonukleotida pada sekuen yang komplementer pada molekul DNA cetakan. Tahap ini disebut annealing. Suhu annealing tiap sekuen DNA sifatnya spesifik dan merupakan penentu utama keberhasilan suatu reaksi PCR. Tahap terakhir adalah tahap ekstensi yang dilakukan pada suhu 720C. Suhu ini merupakan suhu optimum untuk kerja enzim Taq DNA polimerase. Sintesis DNA komplemen dengan DNA cetakan terjadi pada rahap ekstensi. Ketiga tahap tersebut disusun berulang kali dalam mesin PCR, umumnya antara 25-40 siklus, bergantung pada jumlah DNA yang diinginkan. Primer oligonukleotida yang digunakan dalam teknik PCR harus memenuhi beberapa syarat, antara lain memiliki susunan basa yang acak, sehingga tidak terjadi polipurin atau polipirimidin, memiliki jumlah purin dan pirimidin yang seimbang, memiliki titik leleh saling mendekati satu sama lain, dan
sedikit mungkin memiliki basa yang komplementer.Panjang primer yang umum digunakan berkisar 20-30 (Mikkelsen & Corton 2004).
BAHAN DAN METODE
Alat dan Bahan
Alat-alat yang digunakan dalam penelitian ini adalah pipet mikro, tip, tabung Eppendorf, neraca analitik, autoklaf, laminar, vorteks, sentrifus, inkubator bergoyang, cawan petri, alumuniumfoil, stopwatch, penangas air, microwave, mesin PCR, spektrofotometri OD600, elektroforesis, UV illuminator Chemidoc EQ Biorad, cawan petri, api spiritus, erlenmeyer, sheld, parafilm, wadah air, kompor, rak tabung Eppendorf, QIAquick tube, waterbath, cutter.
Bahan-bahan yang digunakan adalah primer OL13F-(GGATTCGGGAGAGGGCC GGTGC) dan OL14R-(GTCGACTCATG TGTGACTTGTCCGGAG) (Masiero et al. 2002), enzim restriksi BamHI, enzim restriksi SalI, Plasmid vektor pCambia 1301 yang disisipi CaMV35S promotor pada MCS, air steril, 2x ligasi buffer, enzim T4 ligase, LB cari, LB padat, glukosa, 1 M Tris-Cl pH 8, 0,5 M EDTA pH 8, 10 N NaOH, 10% SDS, 5 M Kalium asetat, Asam asetat glasial, EtOH 70%, EtOH 95%, TE Buffer, dNTP, MgCl2, 10x PCR buffer, enzim platinum taq polimerase, TAE buffer, leader 1 kb, loading dye, etidium bromida, buffer QG, isopropanol, asetosiringon, buffer PE, buffer EB, Detergen, Baycline, Agrapt, Banlok, Agar, Media MS, media R3B, Media AAM, Media LB padat/cari, Media IK3, Media seleksi I dan II.
Sel bakteri yang digunakan adalah E. coli DH5α yang membawa vektor pGEM-T Easy-OsMADS18, Agrobacterium tumefaciens strain EHA105, dan padi yang digunakan adalah Indica.
Metode
Perbanyakan E.coli DH5α yang mengandung pGEM-T Easy (Sambrook dan Russel 2001)
6
Isolasi DNA Plasmid (QIAGEN 2006)
Isolasi DNA plasmid dilakukan dengan prosedur Mini Preps (QIAGEN 2006). Sebanyak 100 mL kultur bakteri hasil perbanyakan LB cair dimasukkan ke tabung Eppendorf dan disentrifus pada 12000 rpm selama 1 menit (bagian ini diulangi lagi dengan membuang supernatan dan menambahkan lagi kultur ke dalam Ependorf dan disentrifus). Tahap selanjutnya supernatan dibuang dan dilarutkan dengan 100 µL larutan I, di vortek selama 5 menit, kemudian ditambahkan larutan II sebanyak 200 µL (larutan II harus freshmade) dan digoyang bolak-balik 10 kali, kemudian ditambahkan larutan III sebanyak 150 µL, dan di bolak-balik selam 10 kali. Lalu campuran disentrifus dengan kecepatan tinggi (12.000 rpm) selama 10 menit, setelah itu supernatan dipindahkan ke tabung Eppendorf yang baru dan ditambah 450 µL EtOH 95%, disentrifus dengan kecepatan tinggi (12.000 rpm) selama 10 menit, supernatan dibuang dan ditambah 500µL ETOH 70% disentrifus lagi pada kecepatan 12000 rpm selama 10 menit, supernatan dibuang lagi. Tabung Eppendorf diletakkan terbuka pada vakum hingga benar-benar kering, lalu dilarutkan dengan 30 µL buffer TE.
Pemotongan DNA Plasmid dengan Enzim BamHI dan SalI (Invitrogen 2003)
Pemotongan DNA plasmid dengan enzim restriksi sesuai dengan prosedur yang terdapat pada Invitrogen (2003). Plasmid dipotong dengan mencampurkan 20 µL DNA dengan enzim restriksi BamHI sebanyak 1 µL, buffer TE sebanyak 5 µL, dan BSA sebanyak 0,5 µL. Setelah suspensi homogen, kemudian diinkubasi pada suhu optimum selama 2 jam. Setelah inkubasi enzim diinaktifasi dengan dipanaskan pada suhu 700C selama 15 menit. Kemudian dilanjukkan dengan pemotongan menggunakan enzim SalI, yaitu dengan menambahkan sebanyak 1 µL enzim SalI dan diinkubasi lagi selama 2 jam, kemudian enzim tersebut diinaktifasi dengan pemanasan.
Elektroforesis Gel Agarosa (Sambrook dan Russel 2001)
Sampel DNA ditambah dengan bufer loading dye kemudian dimasukkan ke dalam sumur gel agarosa 1%. Elektroforesis dilakukan pada tegangan 75 volt, arus 150 mA. Kemudian gel di cuci dengan Ethidium bromida dan air selama 10 menit. Kemudian
dilihat dibawah UV illuminator Chemidoc EQ Biorad. Untuk pemurnian DNA dari gel agarosa dilakukan dengan mengisolasi gel yang mengandung DNA target yang diketahui dengan membandingkannya dengan marker 1 kb.
Pemurnian DNA dari Gel Agarosa (QIAGEN 2006)
Pemurnian DNA dari gel agarosa dilakukan berdasarkan QIAGEN (2006), gel agarosa dimasukkan ke dalam UV illuminator Chemidoc EQ Biorad, dan diletakkan penanda untuk mengetahui posisi DNA, lalu dilihat dengan sinar ultraviolet. Setelah didapat posisi DNA nya maka dilakukan pemotongan pada agar yang terdapat DNA dan dimasukkan ke dalam tabung Eppendorf yang terlebih dahulu sudah ditimbang bobotnya, setelah dimasukkan agar ke dalam Eppendorf dihitung bobot agar dengan mengurangi bobot Eppendorf kosong dengan Eppendorf yang berisi agar. Kemudian ditambahkan 3x volume buffer QG ke dalam tabung Eppendorf, lalu tabung Eppendorf tersebut dimasukkan ke oven bersuhu 650C selama 10 menit, lalu ditambahkan isopropanol 1 volume dan dimasukkan ke dalam tabung QIAquick dan di sentrifus selama 1 menit, supernatan dibuang dan ditambahkan buffer PE 0,75 mL dan disentrifus kembali selama 1 menit, buang supernatannya. Lalu ke dalam tabung QIAquick ditambahkan buffer EB 50 µL dan disentrifus selama 1 menit, dan disimpan pada suhu -200C.
Penyambungan (Ligasi) DNA (Invitrogen 2003)
Proses ligasi berdasarkan prosedur dari Invitrogen (2003). Ligasi dilakukan dengan nisbah DNA sisipan (OsMADS18) dan vektor 3:1. Proses ligasi dilakukan dengan memasukkan 2 x Ligation Buffer 5 µL ditambah 1 µL vektor pCAMBIA1301 vektor dan 3 µL OsMAD18 serta enzim T4 Ligase sebanyak 1 µL. Semuanya dicampur dalam tabung Eppendorf dan diletakkan pada suhu 11-160C selama 16 jam atau semalam.
Transformasi Agrobacterium tumefaciens (Sambrook dan Russel 2001)
7
detik lalu diinkubasi di dalam es selama 5 menit. Ditambahkan dengan 750 µL media LB kemudian digoyang pada suhu ruang selama 60 menit. Kemudian disentrifus 6000 rpm selama 2 menit. Setelah itu supernatan sebagian dan kemudian disebar pada media LB padat yang sudah ditambahkan dengan rifampicin 50 mg/mL dan kanamycin 50 mg/mL. Penumbuhan Agrobacterium tumefaciens pada media LB padat dilakukan di dalam LAF. Hasil penumbuhan kemudian diinkubasi pada suhu 280C selama 48 jam. Sel Agrobacterium tumefaciens yang terinsersi gen OsMADS18 akan tumbuh membentuk koloni putih pada media LB.
Koloni tunggal pada media LB padat kemudian ditumbuhkan pada 5 mL media LB cair yang sudah ditambahkan dengan rifampicin 50 mg/mL dan kanamycin 50 mg/mL. Penumbuhan koloni tunggal Agrobacterium tumefaciens pada media LB cair dilakukan di dalam LAF. Masing-masing koloni tunggal diambil dengan menggunakan tip setelah itu tip dimasukkan ke dalam media LB cair yang sudah ditambahkan dengan antibiotik. Hasil penumbuhan kemudian digoyang pada suhu ruang selama 48 jam.
Sterilisasi Benih (Coombs dan Franco 2003)
Benih padi yang telah dikupas disterilisasi dengan alkohol 70% selama 1 menit, lalu dibilas 1x dengan aquades steril, setelah itu di rendam dalam larutan bakterisida 10 menit dan larutan fungisida 10 menit, lalu dicuci dengan aquades 3 kali, kemudian di rendam kembali dalam bayclean 20% 15 menit dan bayclean 10% selama 5 menit dan dibilas sebanyak 6x, kemudian ditanam ke media.
Transformasi kalus embriogenik dengan Agrobacterium tumefaciens (Sambrook dan Russel 2001)
Bakteri A.tumefaciens yang telah ditanam pada media AB padat selama 3 hari dipanen dengan menggunakan spatula steril sebayak 2-3 petri (bakteri ditaman merata pada permukaan petri dengan pengaduk kaca segitiga), kemudian dimasukkan ke dalam 100 mL media AAM yang mengandung 100 µM acetoyringone pada erlemeyer steril 250 mL steril, kemudian digoyang perlahan agar homogen, ODnya diukur sampai mencapai OD600 bernilai 1. Kemudian 20 kalus embriogenik yang telah disiapkan direndam dengan suspensi bakteri selama 15 menit, setelah itu kalus hasil infeksi ditiriskan pada kertas steril dengan dirolling dari ujung ke
ujung agar kering dan diganti sampai 2-3 kali. Kemudian bagian atas ditutup juga dengan kertas saring steril dan ditekan sambil digoyang perlahan. Setelah kalus kering ditanam pada media ko-kultivasi (IK3+100 µM asetosiringon) selama 3 hari dengan ditutup plastik hitam.
Seleksi dan Regenerasi (Purnamaningsih 2006)
Kalus dari hasil ko-kultivasi di tanam pada media seleksi I selama 14 hari kemudian kalus yang resisten di subkultur pada media seleksi II selama 14 hari. Kalus yang resisten ditanam pada media regenerasi (R3B: media IK3+0,5mg/L IAA, dan 0,3 mg/L BAP) dan setelah tumbuh planlet, planlet dipindahkan ke media MS sampai perakaran cukup kuat untuk dipindahkan ke media aklimatisasi
Isolasi DNA Tanaman (Jaakola et. al. 2003)
Isolasi DNA dilakukan dengan metode CTAB yang telah dimodifikasi dari Jaakola et al. (2003): Daun muda berukuran sekitar 2 cm atau kalus dimasukkan ke dalam Eppendorf 1,5 mL dan dibekukan dengan nitrogen cair. Daun digerus sampai halus, kemudian ditambahkan dengan 750 ul buffer isolasi yang mengandung buffer lisis [0,2 M tris-HCL pH 7.5, 0.05 M EDTA, 2 M NaCl, 2% (b/v)) CTAB (Hexadecyltrimethyl ammonium bromide)], buffer ekstraksi [0.35 M sorbitol, 0.1 M tris HCl pH 7.5, 5 mM EDTA]. dan 5% (b/v) sarkosil dengan perbandingan 2,5:2,5:1. Selanjutnya campuran diinkubasi 1 jam pada suhu 650C sambil dibolak-balik perlahan. Lalu ditambahkan 750 uL kloroform:isoamil alkohol (24:1). Kemudian sampel disentrifus (12000 rpm, selama 5 menit pada suhu ruang). Lapisan atas diambil dan dipindahkan ke dalam tabung 1,5 mL baru dan ditambah 400 ul isopropanol lalu dibolak-balik hingga gumpalan DNA terlihat. Selanjutnya cairan disentrifus (12000 rpm, selama 6 menit pada suhu ruang) dan cairan dibuang, endapan dicuci dengan 500 ul 70% etanol dan disentrifus lagi (12000 rpm selama 3 menit). Cairan dibuang dan endapan dikeringkan. DNA dilarutkan dalam 50 ul TE (10mM tris-HCL pH 8.0, dan 1 mM EDTA). Sampel DNA disimpan pada -200C
Uji Gus (Rueb dan Hensgen 1989)
8
pH 6.8) yang mengandung 1% Triton X-100 pada suhu 370C selama 1 jam, kemudian dicuci 2 kali dengan buffer fosfat segar, dan divakum selama 5 menit. Setelah itu daun direndam dalam larutan X-gluc dan divakum selama 5 menit, kemudian diinkubasi semalam pada suhu 370C. Buffer fosfat dibuang dan ditambahkan alkohol 70%. Pengamatan dilakukan dengan melihat perubahan warna yang terbentuk pada daun, apabila daun berubah menjadi warna biru maka gen β -glucuronidase terekspresi.
Analisis PCR (Sambrook dan Russel 2001)
Analisis PCR digunakan untuk mendeteksi integrasi gen OsMADS18 pada genom tanaman padi. Komposisi bahan yang digunakan adalah 1x reaksi PCR di dalam volume akhir 15 ul adalah : 1 x buffer PCR yang mengandung Mg2+, 0.05 mM dNTP, 0.05 ul Taq Polymerase, 2.5 ng/ul primer OL13F, 2.5 ng/ul primer OL14R, 1 ul DNA hasil isolasi dan ditambahkan dH2O hingga volume mencapai 15 ul, dan dimasukkan dalam mesin PCR dengan kondisi pra PCR pada susu 950C 5 menit, denaturasi pada 950C 1 menit, annealing pada 550C 1 menit, sintesis DNA pada 720C 1 menit dan ekstensi pada 720C selama 10 menit dengan 40 siklus. Kemudian hasil PCR di elektroforesis.
HASIL DAN PEMBAHASAN
Plasmid rekombinan pembawa Gen OsMADS18
Plasmid rekombinan pembawa gen OsMADS18 berhasil dikonstruksi dengan menyisipkan gen tersebut ke pCAMBIA 1301. Tahap awal kegiatan ini adalah melakukan isolasi plasmid pGEM-T Easy yang membawa gen OsMADS18 dari kultur bakteri Eschericia coli DH5α dan mengkultur bakteri E.coli ke dalam media LB cair yang mengandung antibiotik ampisilin 50 mg/l. Isolasi DNA plasmid dilakukan dengan menggunakan metode lisis alkali merujuk pada Sambrook dan Russel (2001). Adapun prinsip dari metode ini adalah merusak dinding sel dan membran sel dari sel E.coli. Penggunaan Etilen diamin tetra asetat (EDTA) bertujuan untuk merusak dinding sel dengan menghilangkan ion magnesium pada selubung sel, sehingga sel bakteri pecah (Zhiwen et al. 2006). Sodium Deodosil Sulfat (SDS) berfungsi untuk menghilangkan molekul lipid, sehingga akan menyebabkan kerusakan pada membran sel (Davis et al. 2004). Untuk
memisahkan RNA dari DNA ditambahkan enzim RNAse yang akan mendegradasi molekul RNA. Protein dihilangkan dengan penambahan potasium asetat sehingga protein akan mengendap dan dipisahkan melalui sentrifus (Holzmann et al. 2008).
Tahap selanjutnya adalah melakukan pemotongan terhadap vektor dan fragmen gen OsMADS18 menggunakan enzim BamHI dan SalI. Enzim ini mengenali gugus 6 basa spesifik, yaitu G*GATCC (BamHI) dan G*TCGAC (SalI) sehingga hasil pemotongan berujung lenket (Bernard and Pasternak 2002). Pemotongan vektor pCAMBIA 1301 dan gen
OsMADS18 dengan enzim yang sama
bertujuan agar proses ligasi dapat terjadi karena kedua ujung vektor dan DNA sisipan saling berkomplemen. Sedangkan penggunaan dua enzim berbeda bertujuan untuk memastikan arah orientasi gen OsMADS18 ketika terligasi dalam vektor pCAMBIA1301.
Hasil elektroforegram menunjukkan bahwa plasmid pGEM-T Easy yang membawa gen OsMADS18 berhasil diisolasi dari kultur E.coli, dibuktikan dengan adanya pita DNA pada ukuran 3757 bp (Gambar 1; lajur 1). Selanjutnya plasmid dipotong dengan enzim
BamHI dan SalI. Pemotongan ini
menghasilkan 2 pita yaitu pGEM-T Easy (3015 bp) dan OsMADS18 (742 bp) (Gambar 1; lajur 2 dan 3). Sedangkan pemurnian fragmen gen OsMADS18 dari gel agarose merujuk pada metode dari QIAGEN (2006). Tujuan pemurnian adalah untuk mendapatkan fragmen DNA yang diinginkan (OsMADS18) dan menghilangkan pengotor-pengotor lain berupa nukleotida, sisa-sisa garam, dan enzim yang dapat mengganggu proses selanjutnya (Dobhal et al. 2010). Hasil elektroforegram terlihat bahwa pemurnian fragmen gen OsMADS18 berhasil dilakukan, hal ini ditunjukkan dengan adanya pita DNA berukuran 742 bp (Gambar 1; lajur 4).
Gambar 1 Elektroforegram DNA pGEM-T Easy-OsMADS18 (Lajur 1); DNA yang dipotong dengan enzim BamHI dan SalI (Lajur 2 dan 3); Hasil pemurnian fragmen OsMADS18 (Lajur 4)
3015 bp
742 bp 3757 bp
9
Konstruksi plasmid rekombinan dilakukan dengan menyisipkan gen OsMADS18 di bawah kontrol promotor CaMV35S di dalam vektor pCAMBIA 1301 linear. Dari hasil proses ligasi antara vektor pCAMBIA 1301 dengan fragmen gen OsMADS18 diperoleh plasmid rekombinan pCAMBIA
1301-OsMADS18 yang berukuran 12651 bp
(Gambar 2 ; lajur 1). Ukuran ini terdiri atas ukuran vektor pCAMBIA1301 (11819 bp) dan fragmen gen OsMADS18 (742 bp). Sedangkan Gambar 3; lajur 2 adalah hasil restriksi vektor pCAMBIA 1301 – OsMADS18. Pita dengan ukuran 11819 bp adalah vektor pCAMBIA 1301, sedangkan pita berukuran 742 bp adalah gen OsMADS18.
Gambar 2 Elektroforegram pemotongan vector dengan enzim restriksi. lajur 1 ; pCAMBIA 1301 - OsMADS18. lajur 2 ; (11819 bp adalah fragmen vector pCAMBIA 1301, 742 bp adalah fragmen gen OsMADS18) M = λ HindIII
Agrobacterium tumefaciens Pembawa vektor pCAMBIA 1301-OsMADS18
Tahapan selanjutnya adalah transformasi plasmid rekombinan (vertor pCAMBIA 1301
– OsMADS18) ke Agrobaterium tumefacies EHA105 dengan metode kejut panas. Kemudian dilakukan seleksi Agrobacterium tumefaciens EHA105yang mengandung DNA rekombinan berdasarkan pada marka seleksi antibiotik ripamfisin 50 mg/l dan kanamisis 50 mg/l. Selain itu juga dilakukan seleksi koloni biru-putih melalui pemberian IPTG dan X-gal pada media seleksi.
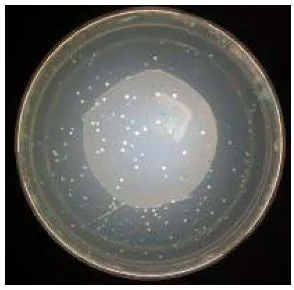
Koloni yang tumbuh pada media seleksi (Gambar 3) adalah koloni yang membawa vector pCAMBIA 1301, hal ini terjadi karena Agrobacterium tumefaciens yang membawa vector ini akan memilik kemampuan resisten terhadap antibiotik ripamfisin dan kanamisin. Sedangkan seleksi koloni biru-putih pada A. tumefaciens EHA105 bertujuan untuk mendeteksi keberadaan gen OsMADS18 dalam vektor berdasarakan marka seleksi Lac-Z. Koloni putih menunjukkan bahwa sel Agrobacterium tumefaciens strain EHA105
membawa gen OsMADS18, sedangkan koloni biru tidak membawa gen OsMADS18.
Gambar 3 terlihat bahwa semua koloni yang terbentuk adalah koloni yang berwarna putih. Terbentuknya koloni putih disebabkan adanya penyisipan fragmen gen OsMADS18 pada lacZ di MCS yang mengakibatkan enzim β-galaktosidase tidak terekspresi (Tsen et al. 2004). Hal ini membuktikan bahwa semua koloni tersebut membawa plasmid pCAMBIA 1301 yang tersisipi gen OsMADS18.
Gambar 3 Koloni Agrobacterium tumefaciesn EHA 105 hasil transformasi dengan pCAMBIA 1301 (koloni putih)
Tranformasi dan Regenerasi Tanaman
Kemampuan Agrobacterium tumefaciens sebagai mediasi transformasi sel tanaman sudah banyak dimanfaatkan, hal ini karena bakteri tersebut mampu membentuk tumor yang akan mengintroduksikan gen asing ke dalam genom tanaman dan gen tersebut akan terintegrasi dan berfungsi dengan baik (Uchimiya et al. 1989). Transformasi kalus dilakukan melalui ko-kultivasi dengan Agrobacterium tumefaciesn EHA105 yang mengandung pCAMBIA 1301 pembawa gen OsMADS18.
Beberapa faktor penting yang menentukan keberhasilan transformasi tanaman padi melalui A.tumefaciens adalah; tipe dan umur jaringan, jenis vektor, genotipe, kondisi kultur jaringan, waktu infeksi. Kalus embriogenik merupakan salah satu faktor penting yang menentukan efisiensi transformasi (Hiei et al. 1997). Secara morfologi, kalus embriogenik mempunyai bentuk yang teratur, compact, bewarna putih atau kekuningan, sedangkan kalus non-embriogenik bersifat lembek, tidak membentuk embrio, dan pertumbuhan kalusnya lambat, tetapi dapat tumbuh besar dalam waktu yang lama (Rueb et al. 1994).
M 1 2
12651 bp 11819 bp
10
Hasil tranformasi diperoleh sejumlah kalus yang membawa gen penyeleksi hpt sehingga bisa hidup dalam media yang mengandung antibiotik higromisin. Jumlah kalus yang resisten terhadap higromisin pada media seleksi adalah 18 kalus. Kalus yang resisten higromisin akan berwarna putih kekuningan. Sedangkan kalus yang tidak resisten akan mengalami gejala nekrotik (warna hitam) (Gambar 4b). Kemudian kalus tersebut dipindahkan ke media regenerasi. Kemampuan regenerasi kalus setelah proses transformasi akan menentukan jumlah planlet yang dihasilkan. Hal ini terkait dengan kombinasi dan konsentrasi hormon yang digunakan pada media regenerasi. Pertumbuhan tunas pertama kali terjadi pada hari ke-4, yaitu dengan menculnya green spot pada kalus (Gambar 4c). Green spot ini akan terus tumbuh hingga menjadi planlet padi trangenik (Gambar 4d).
Gambar 4 Transformasi genetik padi Indica menggunakan A. tumefaciens a)
Kalus yang telah diinfeksi A.tumefaciens pada media ko-kultivasi (kalus hitam mengalami nekrotik = gagal). b) Seleksi kalus resisten higromisin. c)
Pertumbuhan tunas awal pada media regenerasi (wana hijau adalah greenspot yang akan menjadi planlet). d) Planlet padi transgenik.
Tanaman Transgenik
Percobaan ini menghasilkan 18 tanaman transgenik yang mengandung gen OsMADS18. Hal ini dibuktikan dengan uji gus-A dan PCR (Polymerase Chain Reaction). Uji gus-A bisa dilakukan karena plasmid pCAMBIA 1301 mengandung gen gus-A dan gen hpt untuk ketahanan terhadap atibiotik higromisin. Uji histokimia β-glucuronidase dapat dilakukan pada tahap awal segera setelah ko-kultivasi maupun setelah gen diintroduksikan dan terintegrasi dengan stabil dalam genom tanaman. Pada penelitian ini digunakan daun tanaman padi hasil transformasi.
Hasil pengujian terhadap semua tanaman menunjukkan bahwa semua tanaman mengandung gen gus-A. Hasil transformasi positif ditunjukkan dengan adanya warna biru yang muncul pada irisan daun (Gambar 5). Warna biru tersebut disebabkan oleh reaksi substrat X-gluc
(5-bromo-4-chromo-3-indolys-β-D-glucuronide) dengan enzim β -glucuronidase menjadi senyawa intermediet melalui reaksi dimerisasi oksidatif membentuk senyawa dichloro-dibromoindigo (CIBr-Indigo) yang berwarna biru.
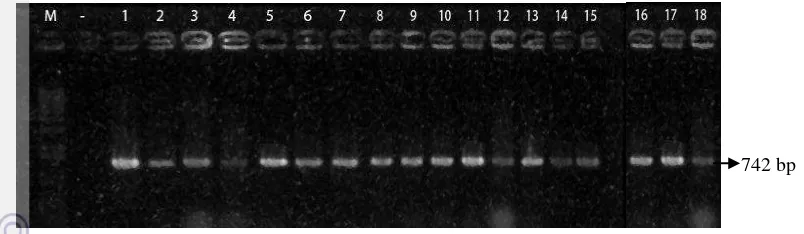
Berdasarkan elektroforegram hasil PCR terhadap 18 tanaman transgenik (Gambar 6), semua tanaman membawa gen OsMADS18. Hal ini ditunjukkan dengan adanya pita yang berukuran 742 bp yang merupakan ukuran dari gen tersebut.
Gambar 5 Hasil uji β-glucuronidase terhadap daun tanaman padi. Hasil positif ditunjukkan dengan terbentuknya warna biru.
KESIMPULAN
Sejumlah 18 planlet Indica berhasil di transformasikan dengan gen OsMADS18. Keberhasilan uji gus-A di tunjukkan dengan munculnya warna biru pada irisan jaringan daun. Sedangkan analisis PCR terhadap 18 planlet transgenik menghasilkan pita berukuran 742 bp, ini menunjukkan bahwa gen OsMADS18 sudah terintegrasi ke dalam genom tanaman padi.
a
d
b
11
SARAN
Tanaman transgenik yang dihasilkan perlu dianalisis lebih lanjut dengan Southern Blot untuk mengetahui jumlah kopi. analisis Western Blot untuk mengetahui ekspresi gen yang diintroduksi. Uji segregasi terhadap keturunan berikutnya untuk mendapatkan tanaman transgenik yang mengandung gen sasaran tetapi tidak mengandung gen penyeleksi antibiotik.
DAFTAR PUSTAKA
Azhakanandam K, MS McCabe, JB POwer, KC LOwe, EC Cocking, MR Davey. 2000. T-DNA transfer, integration, expression and inheritance in rice: Effects of plant genotype and Agrobacterium tumefaciens super-virulence. J Plant Physiol 157:429-439
Belarmino MM. and Mii M. 2000.
Agrobacterium mediated genetic
transformation of a phalaenopsis orchid. Plant Cell Report 19:435-442
Bernard JG, Pasternak JJ. 2002. Molecular
Biotechnology: Principles and
Applications of Recombinant DNA. 3rd Edition. USA: American Society for Microbiology
Casali N, Andrew P, Preston A. 2003. E coli plasmid vectors: methods and applications. Humana Press 12:19-25
Cheng M, Lowe BA ,Spencer M ,Ye X, Armstrong CL. 2004. Factors influencing Agrobacterium-mediated transformation of monocotyledonous species. Dev Biology-Plant 40: 31-45
Christou P. 1990. Inheritance and expression of foreign genes in transgenic soybean plants. Proc. Natl. Acad. Sci. 86:7500-7504. In. Marc de Block. 1993. The cell biology of plant transformation : current state, problem, prospects and the implication for the plant breeding. Kluwer Aad Publisher. Netherlands.
Chuanfeng Z, Jiahe W, Chaozu H. 2010. Induction of chromosomal inversion by integration of T-DNA in the rice genome. J Genet Genomics 37:1-9
Coombs JT, Franco CMM. 2003. Isolation and identification of actinobacterial isolated from surface-sterilized wheat roots. Applied Environ Microbiol 69:5303-5308.
Datta SK, Peterhand A, Datta K and Potrykus I. 1990. Genetically enginered fertile Indica-rice recovered from protoplast. Biotech 8:736-740
Datta SK, Datta K, Soltanifar N, Donn G and Potrykus I. 1992. herbicide resistant Indica-rice plants from IRRI breeding line IR72 after PEG mediated transformation of protoplast. Plant Mol Biol 20:619-629 Davis L et al. 1994. Basic Methods:
Molecular Biology. Jilid 2. Norwola : Appletn & Lange
Dobhal S, P Dinesh, K Anil, A Sanjeev. 2010. Studies on plant regeneration and transformation efficiency of Agrobacterium mediated transformation using neomycin phosphotransferase II (nptII) and glucuronidase (GUS) as a reporter gene. Afr J of Biotech vol 9(41):6853-6859
742 bp
12
Dieffenbach CW, Dveksler GS. 2003. PCR Primer, A Laboratory Manual. 2nd Edition. USA: Spring Harbor Laboratory Pr. Dworkin MM, Falkow W, Rosenberg E,
Schleifer K, Stackebrandt E. 2006. The Prokaryotes. 3rd Edition. USA: Springer.
Enriquez-Obregon GA, Prieto-Samsonov DL, de la Riva GA, Perez MI, Selman-Housein G, Vazquz-Padron RI. 2000. Agrobacterium-mediated Japonica rice transformation a procedure assisted by an antinecrotic treatment. Plant Cell Tiss. Organ Cult. 59: 159-168l
Favaro R, Pinyopich A, Battaglia R, Kooiker M, Borghi L, Ditta G, Yanofsky MF, Kater MM, Colombo L. 2003. MADS box protein complexes control carpel and ovule development in Arabidopsis. Plant Cell 15:2603–2611
Ferrandiz C, Gu Q, Martienssen R, Yanofsky MF. 2000. Redundant regulation of meristem identity and plant architecture by FRUITFUL, APETALA1 and CAULIFLOWER. Development 127:725– 734
Fornara F, Parenicova L, Falasca G, Pelucchi N, Masiero S, Ciannamea S, Lopez-Dee Z, Altamura MM, Colombo L, Kater MM. 2004. Functional characterization of OsMADS18, a member of the AP1/SQUA subfamily of MADS box genes. Plant Physiol 135:2207-2219
Frame B. H Zhang, S Cocciolone, L Sidorenko, C Dictrich, S Pegg, S Zhen, P Schnable, K Wang. 2000. Production of transgenic maize from bombarded type II callus: effect of gold particle size and callus morphology on transformation efficiency. In Vitro Cell Dev Plant 36:21-29
Garris AJ. 2004. Genetic structure and diversity in Oryza sativa L. Genetics 169:1631-1638
Gao Z, Xie X, Ling Y, Muthurrishnan S, Liang HG. 2005. Agrobacterium tumefaciens transformation using an mannose selection system. J Plant Biotech 3:591-597
Ghasemi BK, SV Stepanovich, KG Illich. 2008. Design, construction and cloning of pCAMBIA-MiAMP1 vector for enhancing disease resistance in plants using
Agrobacterium-mediated transformation. J App Bioscien vol 10(2):538-546
Herman M. 2002. Perakitan hasil rekayasa genetik dan pengaturan keamanan hayati Indonesia. Bul Agrobio vol 5(1).
Herman M 2010. Aplikasi teknik rekayasa genetik dalam perbaikan sumber daya genetik tanaman untuk ketahanan cekaman biotik. Bul Plasma Nutfah Vol 16(1):172. Heryati N. 2002. Konstruksi Plasmid
Rekombinan cryIB dan cryIAc di bawah kontrol promotor terinduksi pelukaan [SKRIPSI]. Derpartemen Kimia IPB. Hiei Y, KOmari T, Kubo T. 1997. Efficient
transformation of rice (O.sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. The Plant J 6(0):001-011.
Holzmann J, P Frank, E Löffler, K Bennett, C. Gerner, W Rossmanith .2008. RNase P without RNA: Identification and functional reconstitution of the human mitochondrial tRNA processing enzyme. Cell 135 (3): 462–474
Jaakola L. AM Pirtilla, M Halonen, and A Hohtola. 2001. Isolation of hight-quality DNA from bilberry (Vaccinium myrtillus L.) fruit. Mol Biotech 19:201-203
Jefferson RA. 1987. Assaying chimeric genes in plant : the GUS gene fusion system. Plant Mol Biol Rep 55:387-405
Joseph B, Jini D, Ajisha SU. 2011. Fight global warming wiht genetically altered trees. Asian J Biotech 1-8
Kibria K, Islam MM, and Begum SN. 2008. Screening of aromatic rice lines by phenotypic and molecular markers. J Bot 37(2):141-147
Klee H. 2000. A guide to Agrobacterium binary Ti vectors. Trends in Plant Scie 5:446-451
Lal R and Lal S. 1993. Genetic Enginering of Plants for Crop Improvement. New Delhi: CRS Pr.
13
Lestari AP, Aswidinnorr H, Suwarno. 2007. Uji daya hasil pendahuluan dan mutu beras 21 padi hibrida harapan. Bul Agron 35(1):1-7
Listanto E, Sutrisno, Saptowo JP, Herman M. 2005. Penyisipan gen inhibitor α-amilase pada plasmid biner pCAMBIA 1301. J Agro Biogen 1(2):45-52
Lucca P, Ye X, Potrykus I. 2001. Effective selection and regeneration of transgenic rice plants with mannose as selective agent. Mol Breed 7:43-49
Madigan MT, Martinko JM. 2006. Brock: Biology of Microorganism. 12th edition, San Francisco Perason Education International. p989-990
Masiero S, Imbriano C, Ravasio F, Favoro R, Pelucchi N, Gorla MS, Mantovani R, Colombo L, Kater MM. 2002. Ternary complex formation between MADS-box transcription factor and the histone fold protein NF-YB. J Biol Chem Vol. 277 (29):26429-26435
Matthews PR. Ming-Bo W, Waterhouse PM, Thornton S, Fieg SJ, Gubler F, Jacobsen JV. 2001. Marker gene elimination from transgenic barley, using co-transformation with adjacent "twin T-DNAs" on a standard Agrobacterium transformation vector. Mol Breed 7:195-202
Mikkelsen SR, Corton E. 2004. Bioanalytical Chemistry. New Jersey: John Wiley & Sons.
Olhoft PM, Flaye, LE, Sowers DA. 2004. T-DNA locus structure in a large population of soyabean plant transform using the Agrobacterium-mediated cotyledonany-node methods. Plant Biotech. J. 2: 289-300.
Opabode JT. 2006. Agrobacterium-mediated transformation of plants: emergin factors that influence efficiency. Biotech Mol Biol Rev 1(1):12-20
Pelucchi N, Fornara F, Favalli C, Masiero S, Lago C, Pe ` ME, Colombo L, Kater MM. 2002. Comparative analysis of rice MADS
box genes expressed during flower
development. Sex Plant Reprod 15:113– 122
Peng J, Kononowcz H and Hodges TK. 1992. Transgenic indica rice plants. Theor Appl Genet 83:855-863
Pray L. 2008. Recombinant DNA technology and transgenic animal. Nature Education 1(1):125-133
Primrose SB, Twyman RM. 2006. Principles of Gene Manipulation and Genomics. Ed. Ke-7. Oxford: Blackwell Publishing Purnamaningsih R. 2006. Induksi kalus dan
optimasi regenerasi empat varietas padi melalui kultur In Vitro. J Agro Biogen 2(2):74-80
Rachmawati D, T Hosaka, E Inoue, H Anzai. 2004. Agrobacterium-mediated transformation of javanica rice cv. Rojolele. Biosci Biotech Bioch 68:1193-1200
Rachmawati EE. 2010. Peningkatan Viabilitas (Priming) Benih Juwawut (Setaria italica (L.) P. Beauvois) dengan Polyethylene Glycol (PEG) 6000 [SKRIPSI]. Biologi. Fakultas Sains dan Teknologi. UIN Malang.
Rajesh S, Krishnaveni S, Sudhakar D, Raveendran M, Sivakumar P, Gnanam R, Manickam A. 2008. Agrobacterium-mediated transformation on indica rice (Oryza sativa L.), IR64 with mungbean LEA protein gene for water-stress tolerance. Am J Plant Physiol 3(3):101-110 Rueb S and Hensgens LAM. 1989. Improved
histochemical staining for β
-D-glucuronidase activity in monocotyledonous plant. Genet 6:168-169.
Rueb S, Leneman M, Schilperoort RA, Hensgens LAM. 1994. Efficient plant regenartion through somatic embryogenesis from callus induced on mature rice embryos (Oryza sativa L.). Plant Cell, Tissue And Organ Cult 36:259-264.
Sarkar S, Chaudhuri S, Basu T. 2002. Mechanism of arttificial transformation of E.coli with plasmid DNA-clues from the influence of ethanol. Current Science 83:1376-1380
Sambrook J, Russel DW. 2001. Molecular Cloning : A Laboratory manual.3rd Edition. New York: Cold Spring Harbor Laboratory Pr, Cold Spring Harbor
14
Sha Y, Li S, Pei Z, Luo L, Tian Y, He C. 2004. Generation and flanking sequence analysis of a rice T-DNA tagged population. Theor Appl Genet 108:306-314 Stomp AM. 1992. Histochemical localization of B-glucuronidase. IN : S.R. Gallagher (ed) GUS Protocols : using the GUS gene as reporter of gene expression. London : Academic Press, Inc.
Sutrisno M, Herman S, J Pardal, E Listanto, A Sisharmini, SG Budiarti, D Damayanti, TJ Santoso, R. Wu. 2000. Regenerasi embrio muda jagung Antasena dan Bisma yang ditembak dengan Plasmid pTW-a dan pRQ6. Dalam Moejopawiro S, T Purwadaria, M Herman, A Rukyani, Sutrisno, H Kasim, IN Orbani (eds.). Prosiding Ekspose Hasil Penelitian Bioteknologi Pertanian dalam rangka 25 Tahun Badan Penelitian dan Pengembangan Pertanian. Balitbangtan, Deptan.
Tsen Z et al. 2002. Natural plasmid transformation in Escherichia coli. J Biomel Scine 9:246-252.
Tzfira T, citovsky V. 2002. Partners-in-infection: host proteins involved in the transformation of plant cell by Agrobacterium. Trends in Cell Biol 12:121-130
Tzfira T, Li JX, Lacroix B, Citovsky V. 2004. Agrobacterium T-DNA integration: molecules and models. Trends Genet 20:375-383.
Uchimiya H, Handa T, Brar DS. 1989. Transgenic plant. J Biotech 12:1-20 Utomo SD. 2005. The effect of L-Cysteine on
the efficiency of Agrobacterium-mediated transformation of maize (Zea mays). Bul Agron 33(3):7-16
Walkerpeach C and Velten J. 1994. Agrobacterium-mediated gene transfer to plant cells : cointegrate and binary vector systems. Plant Mol Biol Man B1:1-19 Wangxia W. 2003. Plant responses to drought,
salinity and extreme termperatures: towards genetic engineering for stress tolerance. PLANTA 218(1):1-14
Watson JD, Hopkins NH, Roberts J, Seitz JA, Wainer AM. 2007. Molecular Biology of the Gene. 6 Edition. California: The Benyamin/Cumming Publishing Co Inc.
Widodo 2002. Pengaruh tegangan elektroporasi terhadap efisiensi transformasi plasmid pND968 bakteri asam laktat ke E.coli HB101. J Teknol dan Ind Pang Vol XIII(2):173-177
Xiao H, Wang Y, Liu D, Wang W, Li X, Zhao X, Xu J, Zhai W, Zhu L. 2003. Functional analysis of the rice AP3 homologue OsMADS16 by RNA interference. Plant Mol Biol 52:957–966
Yina K, Biran I, Davie RW. 2004. Simultaneously monitoring gene expression kinetics and genetic noise in single cells by optical well array. Anal Chem 76:6282-6286
Zhiwen Yuan, Jeanne M. VanBriesen. 2006. The formation of intermediates in EDTA and nta biodegradation. Environ Eng Sci 23:533-544.
Zhou HY, et al. 2003. Generating marker-free transgenic tobacco by Agrobacterium-mediated transformation with double T-DNA binary vector. Acta Bot Sin 45:1103-1108
Zohary D, Hopf M. 2000. Domestication of Plants in The Old World. Oxford: Oxford University Pr
16
Lampiran 1 Diagram alir penelitian
Isolasi Plasmid dari
E.coli DH5α
Restriksi dengan enzim
BamHI
dan
SalI
Transformasi ke
Agrobacterium tumefaciens
Transformasi Kalus dengan
A. tumefaciens
Uji gus-A
Kultur Jaringan
Padi Indica
Seleksi dan Regenerasi
Analisis PCR
Seleksi dan Sterilisasi
Benih Padi Indica
Purifikasi DNA dari Gel Agarose
Ligasi DNA ke Vektor pCAMBIA 1301
17
Lampiran 2 Proses umum isolasi plasmid (Sambrook & Russel 2001)
Koloni tunggal bakteri
E.coli
ditumbuhkan ke media LB
diinkubasi selama 24 jam
pada suhu ruang
kultur bakteri dipanen dengan sentrifugasi
pada 10.000 rpm selama 1 menit
pemurnian DNA dengan larutan I, larutan II
dan larutan III
18
Lampiran 3 Peta Plasmid pCAMBIA 1301 (cambia.org)
Lampiran 4 Proses umum reaksi ligasi (Sambrook & Russel 2001)
19
Lampiran 5 Proses purifikasi DNA dengan metode dari QiAgen (2006)
Hasil PCR
+ 3x volume buffer QG dan 1x volume isopropanol
+ Buffer PE 0,75 mL
+ Buffer EB 50 µL
Diperoleh DNA murni
Sentrifuse
Sentrifuse
20
Lampiran 6 Proses umum isolasi DNA tanaman (Sambrook & Russel 2001)
0,5 g bunga padi
Indica
, ditambah N2
lisis sel secara mekanik dengan penggerusan
sel dipanen dengan sentrifugasi
pada 12.000 rpm 10 menit
pemurnian DNA dengan kloroform/isoamil-
alkohol (1:1) dan disentrifugasi pada 12.000 rpm
selama 10 menit
pemekatan DNA dengan etanol absolut
pelet diresuspensi dengan etanol 70%
disentrifugasi pada 12.000 rpm selama 2 menit
dan dikeringkan selama 5 menit
21
Lampiran 7 Bahan-bahan pembuatan media
Media
Komposisi
LB
5 g/l bacto yeast extract, 10 g/l bacto tripton, 5 g/l NaCl, dibuat
pH 7.5; untuk media padat + 15 g/l bacto agar
Makro
elemen
0.65 mg/l NH
4NO
3, 1.9 mg/l KNO
3, 0.37 mg/l MgSO
4.7H
2O,
0.17 mg/l KH
2PO
4, 0.44 mg/l CaCL
2.2H
2O)
Mikro
elemen
0.8 mg/l KI, 6.2 mg/l H3BO3, 22.3 mg/l MnSO2.4H2O, 8.6 mg/l
ZnSO
4.7H
2O, 0.25 mg/l Na2MoO
4.2H
2O, 0.025 mg/l
CuSO4.5H2O, 0.025 mg/l CoCl2.6H2O
IK3
50 ml/l Makro elemen, 2 ml/l Mikro elemen, EDTA 36.7 mg/l,
Thiamin HCL 0.5 mg/l, 30 g/l sukrosa, 2.5 mg/l 2.4-D,
Myoinositol 0.1 g/l, Phytagel 3.75 g/l
Ko-kultivasi media IK3 + 100 uM acetosyringone, pH 5,2
Seleksi I
IK3 + 250 mg/l cefotaxime, 50 mg/l higromisin, 60 mg/l
timentin, pH5,8
Seleksi II
IK3 + 50 mg/l cefotaxime, 50 mg/l higromisin, 50 mg/l timentin,
pH 5,8
Regenerasi
R3B
IK3 + hormon 0.5 mg/l, IAA 0.3 mg/l BAP
MS
50 ml/l Makro elemen, 2 ml/l Mikro elemen, EDTA 36.7 mg/l,
Vitamin, AA (pyridoxine-HCl 0.5 mg/l, thiamin HCl 0.1 mg/l,
nicotinic acid 0.5 mg/l, glycine 3 mg/l), 30 g/l sukrosa, 0.1 g/l
myoinositol, 3.75 g/l phytagel
AAM pH
5,2