VOCAL COMMUNICATION OF WILD CRESTED
MACAQUES (
Macaca nigra
)
MARIA ROSDALIMA PANGGUR
GRADUATE SCHOOL INSTITUT PERTANIAN BOGOR
STATEMENT LETTER
I hereby declare that this thesis entitled Vocal Communication of Macaca nigra is the original result of my own research under supervision of an advisory committee and has never been submitted in any form to any other institution before. All information from other authors cited here are mentioned in the text and listed in the reference list at the end of the thesis.
Bogor, 4 February 2014
Maria Rosdalima Panggur
ABSTRACT
MARIA ROSDALIMA PANGGUR. Vocal Communication of Wild Crested Macaque (Macaca nigra). Supervised by RR DYAH PERWITASARI and ANTJE ENGELHARDT.
Macaca nigra is one of seven Sulawesi macaques’ species, endemic to northern part of Sulawesi. The species is one representative of highly tolerant species of macaques that has received less attention than low tolerance species (e.g. rhesus macaque, M. mulatta). This study presents an overview over the vocal repertoire of wild crested macaques (Macaca nigra) in order to get a comprehensive picture of their communication. The calls were categorized according to the social context in which they were uttered. Each call types within context are presented in the form of spectograms along with descriptive statistics of the different acoustic parameters measured. M. nigra issued 11 call types which were distributed in five distinct social contexts and one loud call. The five social contexts in which the calls were uttered were affiliation, cohesion (including group movement), agonism, predation, and mating contexts. Loud calls were emitted only by males and in various contexts including non-social context. Calls were highly graded, meaning that measures of acoustic parameters overlapped between different contexts. The behavioural contexts and the use of other communication mode can be considered as a source of variation.
Key words: Crested macaque, Macaca nigra, vocal repertoire, social context
ABSTRAK
MARIA ROSDALIMA PANGGUR. Komunikasi Suara pada Macaca nigra. Dibimbing oleh RR DYAH PERWITASARI and ANTJE ENGELHARDT.
terkait perilaku kawin, dan satu tipe suara loud call. Tipe suara loud call
merupakan suara yang dikeluarkan dalam beberapa konteks yang berbeda. Suara
M. nigra tergolong graded, yang berarti bahwa nilai parameter akustik suara-suara saling tumpang tindih antara konteks. Perbedaan konteks sosial dan penggunaan komunikasi multimodal dapat menjadi penyebab variasi suara-suara tersebut. Kata kunci: Crested macaque, konteks sosial, spektogram, vokal repertoire,
SUMMARY
MARIA ROSDALIMA PANGGUR. Vocal Communication of Wild Crested Macaque (Macaca nigra). Supervised by RR DYAH PERWITASARI and ANTJE ENGELHARDT.
Communication is an important aspect in the life of primates. In general, communication in primates involves the visual, auditory, and olfactory system. However, the vocal communication is an effective mode of communication since it may reach further than any other mode, especially for primates living in the forest habitat. Communication has important roles for the social life of primates such as individual or group identity, providing information of the presence of food and predators, and facilitating social interactions in primate social groups.
This research was carried out on wild Macaca nigra, which is one of the seven macaque species endemic to Sulawesi. The species is characterised by a high level of social tolerance. Vocal communication studies on tolerant species are very limited compared to other, more despotic macaque species. The purpose of this study was to determine the variation of vocalizations of M. nigra and the social contexts in which the calls are issued. This study aims at providing a comprehensive overview of the complex social communication of these macaques and indirectly, contributes to the conservation and management of it, both in the wild and in captivity.
A total of 1369 call units were generated from 328 sequences of calls. The calls were categorized based on their acoustic structure and the contexts in which they were uttered. Descriptive statistics of the acoustic parameter measurements are presented.
This study found 11 call types emitted by M. nigra. Based on the physical structure of acoustics, the calls are categorized as tonal calls, non-tonal calls and complex calls (combination of tonal and non-tonal parts). M. nigra emitted the calls in all five main social contexts: affiliation, group cohesion (incl. group movement), agonism, predation and mating behaviour, whereas the loud call was uttered in several different contexts including non-social one. The calls were emitted by all individual, except the calls during mating behaviour and loud calls. Females and males had distinct call structure and calling time during mating behaviour. The loud call was issued only by adult males.
One of the vocalizations can be considered as discrete signals while the others calls show high degrees of grading into each other. The discrete signals include calls in the group cohesion. The graded call signals were issued in the context of affiliation, agonism, the presence of predators and calls related to sexual behaviour. The ability of receiver to assess various clues and the usage of multimodal by the sender would influence the individual’s response to the graded signals.
might give information regarding the emotional state of animals, which is important in ecotourism activities and captive animal management.
RINGKASAN
MARIA ROSDALIMA PANGGUR. Komunikasi Suara Pada Monyet Hitam Sulawesi (Macaca nigra). Dibimbing oleh RR DYAH PERWITASARI dan ANTJE ENGELHARDT.
Komunikasi merupakan salah satu aspek penting dalam kehidupan primata. Secara umum komunikasi pada primata melibatkan sistem visual, auditori, dan olfaktori. Namun, komunikasi dengan suara merupakan moda komunikasi yang efektif terutama bagi primata yang hidup di habitat hutan. Komunikasi memiliki fungsi penting bagi kehidupan sosial primata seperti penanda identitas individu dan kelompok, informasi keberadaan makanan dan predator, dan memfasilitasi interaksi sosial dalam kelompok sosial primata.
Penelitian ini dilakukan pada Macaca nigra liar yang merupakan salah satu primata endemik Pulau Sulawesi. Spesies ini termasuk dalam kelompok dengan tingkat toleransi sosial yang tinggi. Belum banyak penelitian tentang ukuran variasi vokalisasi pada spesies toleran jika dibandingkan dengan spesies Macaca dengan toleransi sosial yang rendah.
Tujuan penelitian ini adalah untuk mengetahui variasi vokalisasi M. nigra
dan konteks sosial saat suara tersebut dikeluarkan. Penelitian ini diharapkan dapat memberi gambaran yang komprehensif tentang komunikasi sosial M. nigra yang kompleks dan secara tidak langsung berkontribusi pada konservasi dan managemen spesies baik di alam maupun di penangkaran.
Jumlah unit suara yang diperoleh dalam penelitian ini adalah 1369 unit yang dihasilkan dari 328 sekuens suara. Suara dibedakan satu sama lain berdasarkan struktur akustik yang dapat terlihat pada spektogram. Hasil pengukuran parameter akustik disajikan dalam analisis statistik deskriptif.
Hasil penelitian ini menemukan 11 tipe suara yang dikeluarkan oleh M. nigra. Berdasarkan struktur fisik akustik, suara M. nigra dibedakan atas suara tonal, non-tonal dan suara kompleks (gabungan antara tonal dan non-tonal). M. nigra bersuara pada lima konteks utama yaitu konteks afiliasi, kohesi (termasuk pergerakan kelompok), agonistik, predasi dan perilaku kawin. Sedangkan loud call merupakan suara yang dikeluarkan dalam beberapa konteks yang berbeda. Semua suara yang diteliti dikeluarkan oleh semua individu pada semua kelompok jenis kelamin kecuali suara pada perilaku kawin dan loud call. Jantan dan betina mengeluarkan suara terkait perilaku kawin yang berbeda secara struktur akustik dan waktu bersuara. Loud call hanya dikeluarkan oleh jantan dewasa.
Suara yang dikeluarkan terbagi atas sinyal diskret dan graded. Kelompok suara diskret sinyal meliputi suara dalam konteks kohesi kelompok (termasuk pergerakan kelompok). Sinyal suara yang termasuk kelompok graded meliputi suara-suara yang dikeluarkan dalam konteks afiliasi, agonistik, kehadiran predator dan suara terkait perilaku kawin. Spesies ini mampu membedakan suara graded
Sebagai anggota kelompok monyet toleran, M. nigra memiliki variasi suara yang lebih beragam dibandingkan dengan spesies monyet lain. Perbedaan variasi suara dengan spesies dari genus Macaca yang lain kemungkinan dipengaruhi berbagai faktor antara lain karakter sosial spesies, perilaku dan habitat.
Data vokalisasi M. nigra dapat melengkapi data penunjang taksonomi dan kegiatan konservasi. Suara yang juga merupakan gambaran kondisi emosi satwa penting dalam kegiatan ekowisata dan manajemen di penangkaran yang berbasis pada kenyamanan satwa.
Copyright © 2013 Bogor Agriculture University Copyright are protected by law,
It is prohibited to cite all or part of this thesis or dissertation without referring to and mentioning the sources. Citation only permitted for the sake of education, research, scientific writing, report writing, critical writing or reviewing scientific problem. Citation does not inflict the name and honour of Bogor Agriculture University.
Thesis
as a partial fulfilment of the requirements for a Master Degree in Animal Bioscience Master Program of Graduate
School of Bogor Agriculture University
VOCAL COMMUNICATION OF WILD CRESTED
MACAQUES (
Macaca nigra
)
MARIA ROSDALIMA PANGGUR
GRADUATE SCHOOL INSTITUT PERTANIAN BOGOR
Title : Vocal Communication of Wild Crested Macaques (Macaca nigra) Name : Maria Rosdalima Panggur
NIM : G35100191
Certified by, Advisory Committee
Dr Ir RR Dyah Perwitasari, MSc Chairman
Acknowledged by,
Dr Antje Engelhardt Member
Coordinator Major of Animal Bioscience
Dr Ir RR Dyah Perwitasari, MSc
Dean of Graduate School
Dr Ir Dahrul Syah, MSc. Agr
PREFACE
I praise and grateful to Almighty God over the completion of this scientific work. This study is part of research collaboration between Macaca Nigra Project, German Primate Center and Bogor Agricultural University. The study was conducted from April 2010 to April 2011. This research topic is part of the study on Social Communication of Macaca nigra by Dr. Jerome Micheletta.
I thank profusely to Dr. Ir. Dyah Perwitasari, MSc and Dr. Antje Engelhardt as the supervising committee, which have supervised and put so much trust to me during my master study and the completion of the thesis. Study funding, research and acoustic training in Germany (April-June 2012) was provided by Dr. Antje Engelhardt from Macaca Nigra Project.
I sincerely thank to Dr. Ir. Entang Iskandar MSi as the thesis defence examiner and to Dr. Rika Raffiudin from Biology Department for insightful comments and correction of the thesis.
I specially thank to Dr. Jerome Micheletta for the invaluable assistance during field data collection and writing of the thesis. Thank you for the patience, knowledge and experience that you shared, as well as the ideas and inspiration that had raised my interest on animal communication. I thank Dr. Kurt Hammerschmidt for the help and guidance in the analysis of acoustic and statistical analysis. I knew nothing when I first met you. And thank to other scientists Dr. Christof Neuman, Dr. Julie Dubosq, Daphne Kerhoas for numerous help and inspiration. I also would like to thank to all the lectures in Animal Bioscience Master Program, who at the end shaping my knowledge up through their high dedication into the research and teaching.
I also would like to thank BKSDA Manado for allowing me conducted the study and to Dr. Muhammad Agil, Edith Sabara, Siti Aminah (IPB) for all the help. This study would not have succeeded without the technical assistance and moral support by the staff of Macaca Nigra Project (Stephen M. Lentey, the late Benediktus Giyarto, Mathilde Chanvin, Meldy Tamengge, Irawan Halir) and all Macaca Nigra research team. The endless gratitude is addressed to my parents, brothers and sister who have supported me by their own way during the study. The least, but very important, I would like to thank to all my friends; class of BSH 2011, BSH students, Andi Darmawan, Rahayu Oktaviani, Andy N Cahyana, Yuni Fanggi Tasik, Yane Moy, Ican Hakim and Sasi Kirono.
Hopefully this scientific work will be useful.
Bogor, 4 February 2014
CONTENT LIST
CONTENT LIST v
LIST OF TABLES vi
LIST OF FIGURES vi
1 INTRODUCTION 1
2 LITERATURE REVIEW 2
Mechanism of Vocal Production of Primates 2
Sound Transmission 3
The Constraint of Sound Propagation in the Field 3
Sound Analysis 4
3 METHODS 5
Study Site and Time 5
Behavioural Observation and Recording Methods 5
Acoustic Analysis 6
Statistical Analysis 7
4 RESULTS AND DISCUSSION 8
Results 8
Acoustic Measurement of Call Type 8
Call Variation per Context 9
1. Affiliation 9
2. Group Cohesion 12
3. Agonism 14
4. Mating 15
5. Predation 16
6. Others Category: Loud call 16
Discussion 17
Discrete and Graded Signal 17
Comparison to Other Macaque Species 17
Implication to Taxonomy and Conservation Issues 18
5 CONCLUSIONS 20
REFERENCES 20
APPENDICES 25
LIST OF TABLES
Table 1 Detail description of acoustic parameters measured in the analysis 6 Table 2 Description of social contexts in which the calls were emitted 9 Table 4 The comparison of Macaque’s vocal communication. 19
LIST OF FIGURES
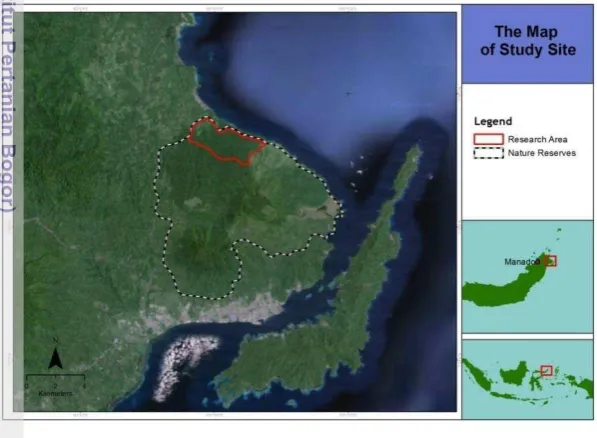
Figure 1 The map of study site of Macaca Nigra Project in Tangkoko-Batuangus NR and Dua Saudara NR, North Sulawesi, Indonesia 5
Figure 2 Spectograms and parameters of call 7
Figure 3 Spectograms of calls used in the affiliation context 13 Figure 4 Spectograms of calls used in the group cohesion and movement
context 13
Figure 5 Spectograms of calls used in the agonistic context 15 Figure 6 Spectograms of calls used in sexual context 16 Figure 7 Spectograms of calls used in the predation context 16
Figure 8 Spectograms of loud calls 16
LIST OF APPENDICES
Appendix 1 The social tolerance among macaque’s society as adapted from
Thierry (2007) 25
Appendix 2 Classification result of discriminant function analysis; main context
1
1
INTRODUCTION
Most primates live within complex social systems. Primates mainly use the visual (gestural and facial) and vocal communication channel in their communication system. Cercophithecinae uses vocalizations more widely since most of them live in dense forest habitats that often limit the transmission of visual signals (Altmann 1967). The calls are given by monkeys in social contexts such advertising individual identity, alerting con-specific about predators, maintaining cohesion during group movement, signalling the presence of food and facilitating social interaction (Cheney and Seyfarth 2010). It can thus be noted that vocal communication played a key role in the evolution of primate social behaviour (McComb and Semple 2005).
The crested macaque (Macaca nigra) is one of seven endemic macaque species that inhabit the island of Sulawesi (Fooden 1969). Similarly to the other Sulawesi macaques, crested macaques are a socially tolerant species where interactions among group members are weakly constrained by kinship and dominance hierarchy (Thierry et al. 2004, Duboscq et al. 2013). This social tolerance is thought to have led to the evolution of a larger communicative repertoire than more despotic species. In a meta-analysis carried out on several macaque species, Dobson (2012) showed that increased social tolerance correlates with a large repertoire of facial expressions. A similar study carried out by Maestripieri (2005) additionally suggests that species with increased social tolerance also have a larger repertoire of communicative gestures. Freeberg et al. (2012) proposed a hypothesis that social communication complexity increases alongside social complexity. The social complexity (group size and time spent grooming) is likely to increase vocal complexity (McComb and Semple 2005). Since the social tolerance might play role in social complexity, the study therefore suggests that in a species as highly tolerant as the crested macaques individuals will also display a high degree of vocal repertoire.
Research on vocalization has been carried out in many species of macaques such as M. fuscata (Green 1975), M. silenus (Hohmann and Herzog 1985), M. radiata (Hohmann 1989), M. fascicularis (Palombit 1992), M. sylvanus (Fischer and Hammerschmidt 2002) and M. cyclopis (Hsu et al. 2005). However, most available data are from less tolerant species, and those carried out on Sulawesi macaques (M. nigra: Lewis 1985; M. tonkeana: Masataka and Thierry 1993) were conducted in captivity where the full extent of a species repertoire might not be expressed. The only reports on the vocalisation repertoire of wild Sulawesi macaques are only descriptive and anecdotal (Thierry et al. 2000). No quantitative descriptions on the vocal patterns of any of the highly tolerant Sulawesi macaques are so far available. Such a study is therefore urgently needed in order to get a more comprehensive picture of the relationship between social tolerance and the complexity of communication in macaques.
2
human disturbance in general raises animal anxiety in the presence of humans, which in turn leads to difficulties in monitoring wild individuals. Since at least certain vocalisations are species specific (Hartwig 2005, Rupell 2009) and individual specific (Bouchet et al. 2012, Neumann et al. 2010, Geissman and Nijman 2006), knowing a species repertoire and an individual’s call pattern will help population monitoring activities for conservation purposes. It has been demonstrated by Adi et al. (2009) that a population census in Emberiza hortulana
using vocal recognition can predict the population density and structure simpler and more accurate as compared to other population monitoring methods. Furthermore, different types of vocalisations might represent the internal emotion of animals either positive state (Boissy et al. 2007) or negative state (Dupjan et al. 2008, Fichtel et al. 2001), so the understanding of macaque's vocalization will be useful for the species’ management and to ensure their welfare.
The aim of this study therefore is to provide quantitative data of crested macaque vocalizations to get a better understanding about the communication system of this species. The study also attempt to categorize calls based on the context in which they were emitted and examine whether acoustic features overlap within and between calls in different contexts to provide an estimation of the graded nature of the crested macaque vocal communication system. In addition to support species management and conservation, this research will provide a key of future studies on the vocal communication of Sulawesi macaques.
2
LITERATURE REVIEW
Mechanism of Vocal Production of Primates
3 and Linker 1995). These differences have presumably limited the ability of vocal variation in non-human primate.
The air puff from the larynx produces sound, which can be either tonal or noisy sound. Tonal sound is characterized by one excitation frequency, which is sometimes followed by harmonics or overtones. The harmonics are integer multiple of the fundamental frequency. While noisy sound is a periodic signal, which may also be produced at the vocal folds as a result of air turbulence. Noisy call can be produce either at the larynx or above the larynx (due to constriction elsewhere in the vocal tract, Fitch and Hauser 1995). The combination of tonal and non-tonal call can be called complex calls (Micheletta 2012).
Sound Transmission
The sound waves are only able to propagate through a medium. The sound waves that travel through gasses and liquid are called longitudinal waves while sound waves that transmit through solid medium are called transversal waves (Bradbury and Vehrencamp 1998). Hence the vocal exchange in primate is a longitudinal sound wave that is transmitted through air. The sound wave transmission is simply described as the following explanation; when there is no vibration from any sound source, the air pressure is called ambient pressure. The vibration of the sound source increases the collision of molecules in the air. When the molecules move and stay away due to the collision, the air pressure is below ambient pressure and increase again when the next tremor occurs. The state when the molecule assembled is called compression while the state when the molecule expanded is called rarefaction. The temporal pattern of rise and fall in signal amplitude is called the waveform of the signal. The largest distance of air pressure toward ambient pressure is called amplitude. The distance between two successive peaks of high pressure called wavelength. The number of cycles (one rise and fall of the signal) per second is called frequency. The unit of frequency, cycles/second is called Hertz. The time is needed by a cycle is called period (Bradbury and Vehrencamp 1998).
The Constraint of Sound Propagation in the Field
4
The constraints of sound wave transmission in the field can affect the quality of sound recordings (Maciej et al. 2011). Signal quality can also be masked by background noise such as the sound of other animals or observer. Large distances between sound emitter and observer will potentially enhance sound degradation. Some simple efforts such as pointing the microphone towards to the sound source within close proximity, can overcome the problem. Depending on the call, e.g. low amplitude calls like grunts, good sound quality may be obtained from 3-5 m distance while high amplitude signals might be recorded at 15 m (Fischer et al. 2013). Nevertheless, for further analysis using computers, preliminary assessment of the reliability of recordings needs to be done for any signals recorded in large distance (Maciej et al. 2011).
Sound Analysis
Fischer et al. (2013) provided a detailed explanation of the study of bioacoustic in primates along with a guideline for playback experiments. Data collected in the field is transferred into a computer for analysis. Computer program analyses that are widely used to extract acoustic features of sound signal are Avisoft SAS Lab (R. Specht, Berlin), RAVEN (Cornell Lab of Ornithology), PRAAT (Institute of Phonetic Science, http://www.praat.org) or Signal (Engineering Design, Belmont, MA). The computer-aided analysis still faces problems related to the complexity of signal, high variation of vocalization, environmental effect and distance and spatial orientation of the animal towards the microphone. Using a multi-parameter approach, as provided by the software LMA (Schrader and Hammerschmidt 1997), can reduce the importance of these problems.
Sound is a complex signal, mostly made up of non-pure sinusoidal waveforms. To deal with this acoustic complexity, the Fourier transformation technique is required. The main principle of this technique is to transform any continuous waveform into a pure sine wave. Sound signals in pure sinusoidal form are easy to compare or to analyse their frequency, amplitude and relative phase. In a computer program, the Fast Fourier Transform (FFT) breaks down the signal into smaller unit samples and records the amplitude at each point. The advantage of this technique is that sound signals can be analysed rapidly. The disadvantage is that when the number of signal points is too small, the computer will have difficulties to determine the frequency of the signal. This problem can be solved by applying the Nyquist-Shanon equation in which the sampling rate of recording equipment is set at twice the frequency of the signal frequency (Bradburry and Vehrencamp 1998). The sampling rate for most mammal vocal recordings can be set at 40 kHz, which is twice the maximum capability of human hearing, 20 kHz. Yet, the sampling rate that is commonly used is 44.1 kHz, parallel to the sampling rate of a signal in compact disc storage.
5 of the transformation window, which is chosen with respect to the quality of the signal (Schrader and Hammerschmidt 1997).
3
METHODS
Study Site and Time
The research was conducted on two wild groups of crested macaques living in the Tangkoko Nature Reserve (NR), North Sulawesi (Figure 1). The research was carried out under the Macaca Nigra Project (research collaboration between Bogor Agricultural University, the University Sam Ratulangi and the German Primate Center), which has been running since 2006 (www.macaca-nigra.org). Monkeys are well habituated to the presence of researchers. The two groups of crested macaques, which were the focus of research, are group Rambo 1 (R1) and group Pantai Batu (PB), each consisting of 65-75 individuals. The field data collection was conducted from September 2010 until March 2011.
Figure 1 The map of study site of Macaca Nigra Project in Tangkoko-Batuangus NR and Dua Saudara NR, North Sulawesi, Indonesia
Behavioural Observation and Recording Methods
6
2006-2012). Ad-libitum and 30 min long focal animal sampling were used to note the behaviour associated with call production (Altmann 1974). The focal individual was chosen according to a pseudo-randomised order so that all adults were followed every 2 days. The behavioural data of each focal individual were obtained equally between three different time periods: morning (0600-1000), midday (1000-1400) and afternoon (1400-1800). The observational data was recorded using a Psion handheld computer with Windows mobile and Excel (Microsoft). All social interactions were videotaped with a Panasonic HDC-SD700 HD camcorder that was connected to a Sennheiser K6-ME66 directional microphone. The sampling rate was 44.1 kHz. In this way, the video and sound were simultaneously obtained.
Acoustic Analysis
For the acoustic analysis, each sequence of calls was inspected using Avisoft SAS Lab Pro 5.2 (Avisoft Bioacoustics, Berlin, Germany). Call sequences were cut off per unit call for subsequent analysis. Each selected unit calls were free from unwanted signals like bird song, insect or other sound. Call units were down sampled from 44025 to 11025 Hz for better frequency resolution. Fast Fourier transform (1024-pt FFT; frame size 100%; window: Hamming; overlap 93.73%) were conducted for all the calls. The resulting time resolution was 5.8 ms at 11 Hz of frequency resolution. All the call units were saved in the ASCII format files for further analysis using LMA, a software tool to extract different measures of acoustic parameters from acoustic signals (Schrader and Hammerschmidt 1997).
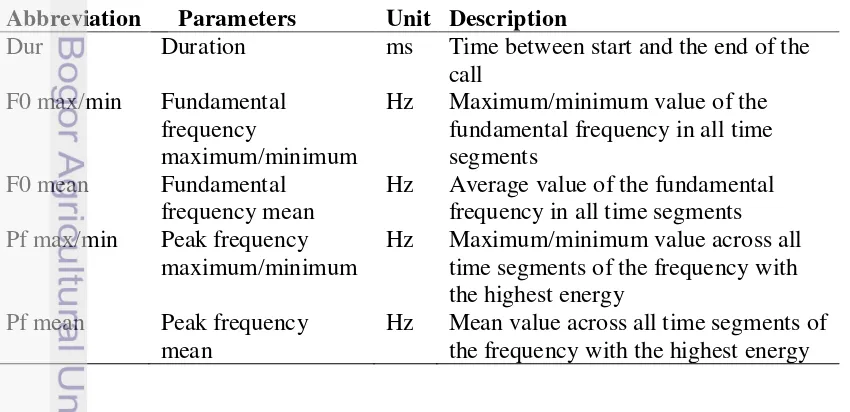
An interactive macro was applied to calculate acoustic parameter of each call unit. The cut-off frequency was set at 200 Hz to reduce the noise signal. In this way, all frequencies ranging below 200 Hz were excluded from the analysis. The start and end threshold was set at 20 %, meaning that all time segments in the beginning and the end of calls that were considered for analysis were more than 20 % of maximal amplitude. Detailed information of acoustic parameter measured in the study was presented in Table 1 and Figure 2.
Table 1 Detailed description of acoustic parameters measured in the analysis Abbreviation Parameters Unit Description
Dur Duration ms Time between start and the end of the call
F0 max/min Fundamental frequency
maximum/minimum
Hz Maximum/minimum value of the fundamental frequency in all time segments
F0 mean Fundamental frequency mean
Hz Average value of the fundamental frequency in all time segments Pf max/min Peak frequency
maximum/minimum
Hz Maximum/minimum value across all time segments of the frequency with the highest energy
Pf mean Peak frequency mean
7
Figure 2 Spectograms and parameters of call (a: call start, b: call end, c: duration (ms), Fmax: Maximum frequency, Pf: Peak frequency, F0: Fundamental frequency).
Statistical Analysis
A step-wise discriminant function analysis (DFA) was used to assign the calls to each context by determining which acoustic variables would maximize the differences between contexts. First, the assignment test was applied to determine whether the calls would be reliably classified to the main contexts; affiliation, cohesion, agonism, predation, mating and loud calls. Since there were at least two different calls within the main contexts (named as sub-context calls), the second test was applied to check the assignment of sub-context calls within each main contexts. The sub-context calls refer as call types except for predation calls where sub-context calls differentiated by the predator type. Those contexts of interaction and call types were used as grouping variables and all acoustic parameters as predictors. The leave-one-out procedure of cross validation was applied to check the validation of the discrimination result by which each case is classified by the function derived from all cases other than that case. The assignment of calls into graded and discrete was determined by 70 % threshold of correctly assignment value.
To test whether the acoustic structure of call parameters (duration, F0 max/min/mean, Pf max/min/mean) varied related to the context and sex, a linear mixed model was applied. The sex and sub contexts were used as fixed factors while identity of caller was set as random factors. The test was done to each acoustic parameter as dependent factors. The Least Significant Differences (LSD) was used as adjustment for multiple comparisons. For mating context, the differences between copulation calls of males and females were compared with
F
re
que
nc
y (
kH
z
8
Anova. Sex was set as factor and the acoustic parameters were used as dependent variables. All data analysis was carried out with SPSS 20 (IBM).
4
RESULTS AND DISCUSSION
Results
A total 1369 call units were generated from 328 sequences of calls. The analyses resulted in 11 call types distributed in five social contexts. Another category of call, the loud calls was used in several different contexts (Table 2). The general physical structures of calls were tonal, atonal and complex. All individuals issued all types of calls except sexual related call and loud call.
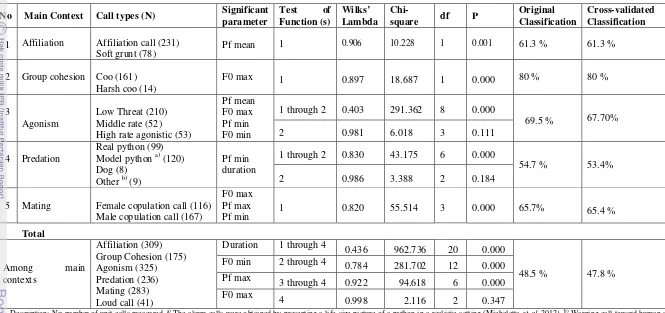
Acoustic Measurement of Call Type
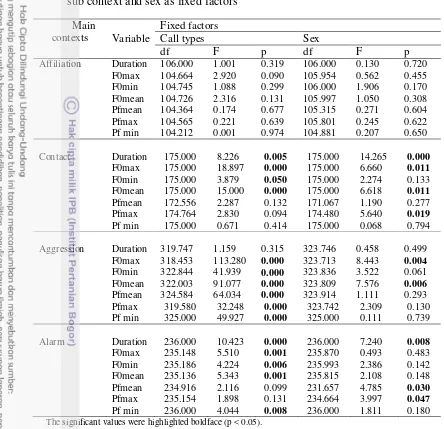
The assignment into main context categories resulted in a percentage of correctly classified calls of 47.8 %. The calls were discriminated based on the sub context within each main context. For example, the affiliation context consists of two sub-contexts, which was discriminated by 61.3 % after cross validation. The variables that significantly discriminated the two groups of sub contexts were Pf mean, duration, F0 max, F0 min (Table 3). Descriptive statistics of the acoustic parameter for each call type per each context is presented in the Table 4.
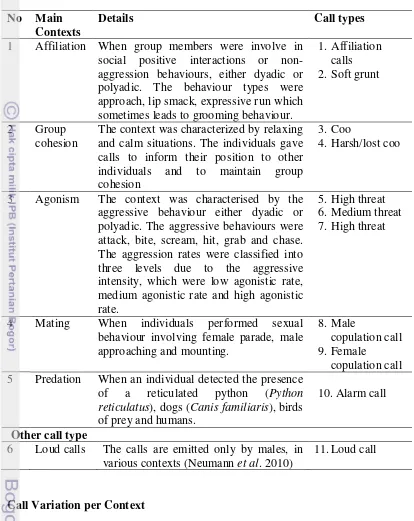
9 Table 2 Description of social contexts in which the calls were emitted No Main
Contexts
Details Call types
1 Affiliation When group members were involve in social positive interactions or non-aggression behaviours, either dyadic or polyadic. The behaviour types were approach, lip smack, expressive run which sometimes leads to grooming behaviour.
1. Affiliation calls 2. Soft grunt
2 Group cohesion
The context was characterized by relaxing and calm situations. The individuals gave calls to inform their position to other individuals and to maintain group cohesion
3. Coo
4. Harsh/lost coo
3 Agonism The context was characterised by the aggressive behaviour either dyadic or polyadic. The aggressive behaviours were attack, bite, scream, hit, grab and chase. The aggression rates were classified into three levels due to the aggressive intensity, which were low agonistic rate, medium agonistic rate and high agonistic rate.
5. High threat 6. Medium threat 7. High threat
4 Mating When individuals performed sexual behaviour involving female parade, male approaching and mounting.
8. Male
copulation call 9. Female
copulation call 5 Predation When an individual detected the presence
of a reticulated python (Python reticulatus), dogs (Canis familiaris), birds of prey and humans.
10.Alarm call
Other call type
6 Loud calls The calls are emitted only by males, in various contexts (Neumann et al. 2010)
11.Loud call
Call Variation per Context
1. Affiliation
10
Table 3 The classification of calls based on the discriminant function analysis (DFA). Discriminant analysis to the loud call provided by other study (Neumann et al. 2010)
No Main Context Call types (N) Significant parameter
Test of Function (s)
Wilks’ Lambda
Chi-square df P
Original Classification
Cross-validated Classification
1 Affiliation Affiliation call (231)
Soft grunt (78) Pf mean
1 0.906 10.228 1 0.001 61.3 % 61.3 %
2 Group cohesion Coo (161) Harsh coo (14)
F0 max 1 0.897 18.687 1 0.000 80 % 80 %
3
Agonism
Low Threat (210) Middle rate (52) High rate agonistic (53)
Pf mean F0 max Pf min F0 min
1 through 2 0.403 291.362 8 0.000
69.5 % 67.70% 2 0.981 6.018 3 0.111
4 Predation
Real python (99) Model python a) (120) Dog (8)
Other b) (9)
Pf min duration
1 through 2 0.830 43.175 6 0.000
54.7 % 53.4% 2 0.986 3.388 2 0.184
5 Mating Female copulation call (116) Male copulation call (167)
F0 max Pf max
Pf min 1 0.820 55.514 3 0.000 65.7% 65.4 %
Total
Among main contexts
Affiliation (309) Group Cohesion (175) Agonism (325) Predation (236) Mating (283) Loud call (41)
Duration 1 through 4 0.436 962.736 20 0.000
48.5 % 47.8 % F0 min 2 through 4 0.784 281.702 12 0.000
Pf max 3 through 4 0.922 94.618 6 0.000
F0 max 4
0.998 2.116 2 0.347
11 Table 4 Mean and standard deviation for each call type per context, which is obtained with LMA
No
Main Context Sub Context N Dur (ms) F0max (Hz) F0min (Hz) F0mean (Hz) Pfmean (Hz) Pfmax (Hz) Pfmin (Hz)
1 Affiliation
Affiliation call 231
192.15 ± 74.84 438.36 ± 114.82 261.82 ± 76.65 355.58 ± 97.56 877.75 ± 645.96 1592.19 ± 1257.82
398.89 ± 365.96
Soft grunt 78
228.73 ± 130.09 393.13 ± 94.58 266.13 ± 73.65 332.45 ± 85.18 1154.56 ± 583.85 1759.26 ± 993.07 617.17 ± 527.51
2 Contact call
Coo 161
281.23 ±115.41 546.21± 138.86 423.68± 113.93 494.11± 109.33 958.32± 345.28 1416.14± 675.23 582.26± 364.18
Harsh coo 14
338.89 ±115.97 737.36± 281.22 495.14± 315.96 635.64± 297.77 1117.71± 358.86 1665.29± 986.33 705.86± 322.81
3 Aggression
Low Threat 210
236.0± 118.20 472.33± 182.3 319.57± 157.43 403.24± 165.20 1194.89± 577.68 1975.47± 1061.24 604.27± 521.02
Middle rate 52
215.72± 126.78 558.06± 325.34 393.5± 267.37 472.71± 277.72 1515.17± 519.84 2085.04± 919.87 854.25± 445.73 High rate
agonistic 63
201.53± 127.97 1040.08± 545.42 534.95± 381.57 829.92± 484.86 2631.87± 1023.2 4151.35± 2400.74 1444.57± 600.14
4 Alarm
Real python 99
210.38± 59.63 681.18± 264.76 515.46± 287.74 601.98± 261.05 1338.38± 716.06 1911.16± 1052.43 870±31 748.91
Model python a) 120
197.09± 81.92 642.68± 172.57 450.65± 188.79 557.51± 172.38 1483.9± 542.59 1977.98± 901.78 1086.8± 461.38
Dog 8
332.05± 85.06 538.13± 285.48 336.38± 271.62 444.13± 296.45 1114.25± 254.96 1845.75± 673.16 399.63± 351.76
Other b) 9
313.20± 227.01648 535.67± 211.29 404.00± 210.61 472.56± 207.92 1015.67± 629.99 1305.89± 821.22 666.00± 659.50
5 Sexual
Female
copulation call 116
102.6± 33.6 362.92± 187.98 308.51± 190.14 332.59± 187.75 843.80± 485.23 976.09± 692.4 726.5± 457.88 Male copulation
call 167
123.81± 47.52 568.32± 333.03 421.44± 285.32 499.89± 307.15 1398.49± 1210.54 1842.25± 1647.77 853.91± 952.79
6 Loud call 41
494.1± 103.43 1077.29± 119.85 977.12± 139.44 1022.97± 119.42 1061.51± 211.05 1251.05± 852.93 983.95± 159.73
12
Table 5 LMM results per main context. Subject ID was set as random factor and sub context and sex as fixed factors
Main contexts
Fixed factors
Variable Call types Sex
df F p df F p
Affiliation Duration 106.000 1.001 0.319 106.000 0.130 0.720 F0max 104.664 2.920 0.090 105.954 0.562 0.455 F0min 104.745 1.088 0.299 106.000 1.906 0.170 F0mean 104.726 2.316 0.131 105.997 1.050 0.308 Pfmean 104.364 0.174 0.677 105.315 0.271 0.604 Pfmax 104.565 0.221 0.639 105.801 0.245 0.622 Pf min 104.212 0.001 0.974 104.881 0.207 0.650
Contact Duration 175.000 8.226 0.005 175.000 14.265 0.000 F0max 175.000 18.897 0.000 175.000 6.660 0.011 F0min 175.000 3.879 0.050 175.000 2.274 0.133 F0mean 175.000 15.000 0.000 175.000 6.618 0.011 Pfmean 172.556 2.287 0.132 171.067 1.190 0.277 Pfmax 174.764 2.830 0.094 174.480 5.640 0.019 Pf min 175.000 0.671 0.414 175.000 0.068 0.794
Aggression Duration 319.747 1.159 0.315 323.746 0.458 0.499 F0max 318.453 113.280 0.000 323.713 8.443 0.004 F0min 322.844 41.939 0.000 323.836 3.522 0.061 F0mean 322.003 91.077 0.000 323.809 7.576 0.006 Pfmean 324.584 64.034 0.000 323.914 1.111 0.293 Pfmax 319.580 32.248 0.000 323.742 2.309 0.130 Pf min 325.000 49.927 0.000 325.000 0.111 0.739
Alarm Duration 236.000 10.423 0.000 236.000 7.240 0.008 F0max 235.148 5.510 0.001 235.870 0.493 0.483 F0min 235.186 4.224 0.006 235.993 2.386 0.142 F0mean 235.136 5.343 0.001 235.815 2.108 0.148 Pfmean 234.916 2.116 0.099 231.657 4.785 0.030 Pfmax 235.154 1.898 0.131 234.664 3.997 0.047 Pf min 236.000 4.044 0.008 236.000 1.811 0.180
The significant values were highlighted boldface (p < 0.05).
Table 6 Results of anova test to the male and female copulation calls
Variables df F p
Duration 1 18.163 0.000
F0max 1 35.463 0.000
F0min 1 13.672 0.000
F0mean 1 26.709 0.000
Pfmean 1 21.892 0.000
Pfmax 1 28.527 0.000
Pfmin 1 1.786 0.182
13
1 2 3
2 4 6 8 1 0
1 s 2
[image:30.595.83.514.41.772.2]4 6 8 1 0
Figure 3 Spectograms of calls used in the affiliation context: a) An affiliative call by a female toward other female with her baby, b) A male gives a soft grunt during an affiliative interaction toward other male which involve genital grasp.
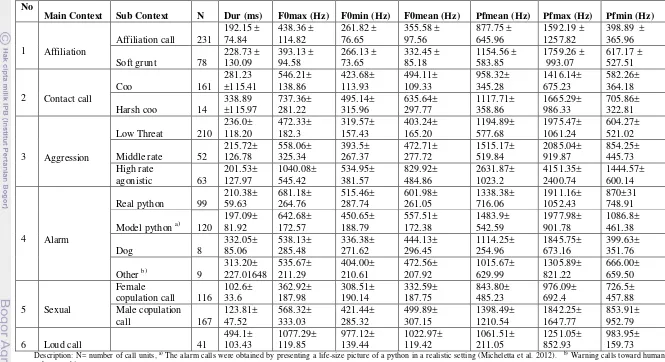
2. Group Cohesion
The call types in this context were called contact calls or coo calls. Most of these calls had a rich tonal harmonic pattern. There were three types of contact calls in this study, which were also presented by Thierry et al. (2000); clear coo (Figure 4a), low coo (Figure 4b), and harsh coo (Figure 4c). Low coos were mostly exchanged by individuals in close proximity. Clear coos were uttered when an individual separated >10m from the main group. By ear, the low coo and clear coo were not easily distinguished each other. While harsh coo has higher frequency and amplitude than the other two, which is easily recognise by observer.
1 2 3 4
2 4 6 8 0
1 2 3
2 4 6 8 0
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 2
[image:30.595.111.504.442.679.2]4 6 8 10
Figure 4 Spectograms of calls used in the group cohesion and movement context; a) low coo exchanges between two females at a distance, b) clear coo of a female who was far from the group, c) Harsh call emitted by adult female who was separated from the group, d) female contact call for her baby.
F re q ue nc y ( kH z) Time (s) F re q ue nc y ( kH z) F re q ue nc y ( kH z) F re q ue nc y ( kH z) Time (s)
Time (s) Time (s)
1 2 3 4 5
2 4 6 8 1 0 F re q ue nc y ( kH z)
a) b)
c) d)
a) F re q ue nc y ( kH z) F re q ue nc y ( kH z) b)
14
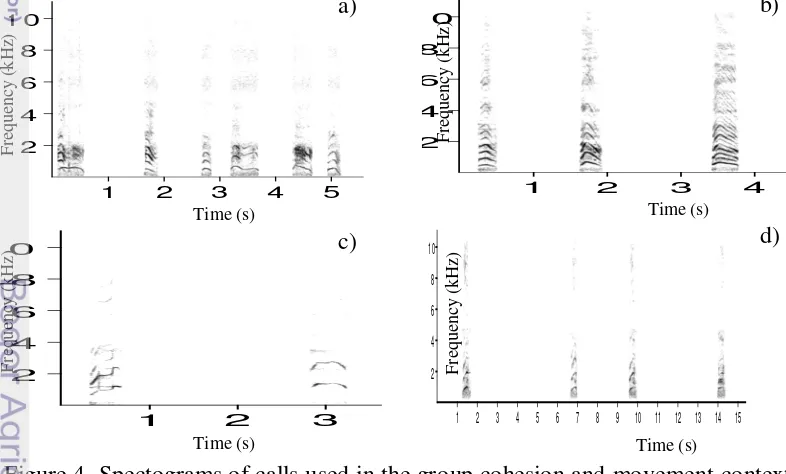
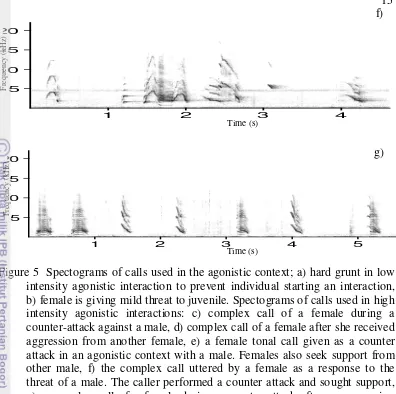
3. Agonism
There were several different types of calls in this context, which seemed to correspond to the intensity of the aggressive interactions: low threat (Figure 5a), medium threat (Figure 5b), and high threat (Figure 5 c-g). Due to the high variability of acoustic features of agonistic calls, the calls fell into three main call groups, namely pulsed calls, tonal screams and complex scream (Micheletta 2012). One aggressive interaction could comprise a sequence of many basic patterns of calls.
Low threat calls were generally issued when initiating aggression. Mild threat were frequently accompanied by the half-open mouth display and seemed to prevent intervention or as a protest to avoid interaction initiated by other individuals. The open mouth bared-teeth face accompanied high threat calls. High threat calls were issued to initiate aggression, to counter attack and during aggressive interventions. 1 2 5 10 15 20 1 5 0 5 0
1 2 3 4
5 0 5 2 0
1 2 3 4 5
5 0 5 20
1 2 3 4 5 6 7
5 10 15 20 c) F re que nc y (k H z ) F re que nc y (k H z ) d) F re que nc y (k H z ) e)
Time (s) Time (s)
F re q ue nc y ( kH z) F re q ue nc y ( kH z) Time (s) Time (s) Time (s)
15
1 2 3 4
5 10 15 20
1 2 3 4 5
[image:32.595.115.511.65.459.2]5 0 5 20
Figure 5 Spectograms of calls used in the agonistic context; a) hard grunt in low intensity agonistic interaction to prevent individual starting an interaction, b) female is giving mild threat to juvenile. Spectograms of calls used in high intensity agonistic interactions: c) complex call of a female during a counter-attack against a male, d) complex call of a female after she received aggression from another female, e) a female tonal call given as a counter attack in an agonistic context with a male. Females also seek support from other male, f) the complex call uttered by a female as a response to the threat of a male. The caller performed a counter attack and sought support, g) a complex call of a female during a counter attack after an aggression initiated by a juvenile
4. Mating
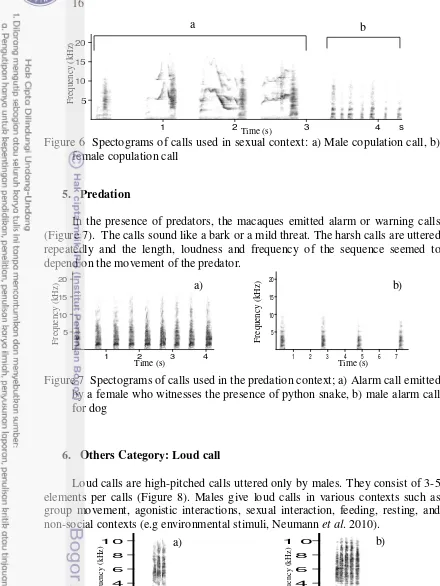
Sexually-related calls consist of male copulation calls and female copulation calls. Male copulation calls were characterized by a series of similar repetitive squeaks (Figure 6a). Female copulation call consisted of repetitive short-pulsed calls (Figure 6b). Female copulation calls varied on an individual level with the number of call units ranging from 3 to 8 units call. Males emitted the call during copulation. Females called after males had demounted, but not all females were observed emitting the calls.
f)
g)
F
re
que
nc
y
(k
H
z
)
F
re
que
nc
y
(k
H
z
)
Time (s)
16
1 2 3 4 s
[image:33.595.41.481.53.639.2]5 10 15 20
Figure 6 Spectograms of calls used in sexual context: a) Male copulation call, b) female copulation call
5. Predation
In the presence of predators, the macaques emitted alarm or warning calls (Figure 7). The calls sound like a bark or a mild threat. The harsh calls are uttered repeatedly and the length, loudness and frequency of the sequence seemed to depend on the movement of the predator.
1 2 3 4
5 10 15 20
1 2 3 4 5 6 7
5 10 15 20
Figure 7 Spectograms of calls used in the predation context; a) Alarm call emitted by a female who witnesses the presence of python snake, b) male alarm call for dog
6. Others Category: Loud call
Loud calls are high-pitched calls uttered only by males. They consist of 3-5 elements per calls (Figure 8). Males give loud calls in various contexts such as group movement, agonistic interactions, sexual interaction, feeding, resting, and non-social contexts (e.g environmental stimuli, Neumann et al. 2010).
T im e (se 1 2 4 6 8 10
T i m e (
1 2 4 6 8 1 0
Figure 8 Spectograms of loud calls: Male loud call consists of three elements (a), and up to five elements (b). The ones depicted were emitted during aggression F re q ue nc y ( kH z) F re q ue nc y ( kH z) F re q ue nc y ( kH
z) a) b)
F re que nc y (k H z ) F re que nc y (k H z )
a) b)
b a
Time (s)
17 Discussion
Discrete and Graded Signal
The vocal repertoire of M.nigra was found to be highly variable, and consisting of a mix of discrete and graded signals. A discrete signal is a specific class of signal that can be easily distinguished from other types of signals. Conversely, a signal is considered as graded when its acoustic features and the context in which it is uttered overlap with other signals (Marler 1977, Fischer and Hammerschmidt 2002). All the calls emitted by crested macaques can be considered as graded signals except those calls used in the context of group cohesion. The loud call is a specific call that is only issued by the males graded in various contexts (Neumann et al. 2010). The calls in the affiliation, agonistic context, predation context and mating context graded with each other between sub-contexts. This graded vocal repertoire within the genus Macaca has been reported in other species such as M. radiata (Hohman1989), M. sylvanus (Fischer and Hammerschmidt 2002) and M. cyclopis (Hsu et al. 2005).
The ability of primates to extract information from graded signal and respond appropriately has been confirmed by several playback studies (Rendall et al. 1999, Fischer et al. 2001). Even in the agonistic context, highly variable screams can be discriminated by the receivers. Rhesus macaque and chimpanzees, for example are able to infer the intensity of aggression when hearing a given scream (Scolombe et al. 2009, Gouzoules et al. 1984). The context in which the calls are uttered might facilitate the individual's decision to call or respond. In an experiment, Michelatta et al. 2012 presented crested macaques with life-sized models of reticulated pythons in the realistic setting in order to get the predator alarm calls. The experiment corroborates the assumption that graded calls in crested macaques encode severity of a situation, since in this study an individual who saw the python picture gave stronger vocal responses than an individual who did not see the picture. Individuals who did not see the snake picture approached the scene but did not give any call. The study suggests that some primates assess context based on various clues before deciding to issue a signal. This ability was recently supported by a study on green monkeys (Chlorocebus sabaeus) who gave grade responses with acoustic difference cues with appropriate predator escape behaviour (Price and Fischer 2013).
The usage of multimodal communication by the sender might be another reason why receivers can understand graded signal. With multimodal signals, the caller intent can be reinforced by a gesture or a facial expression and vice versa (Partan and Marler 1999, Micheletta et al. 2013). Crested macaques are mostly terrestrial, which allows each individual to communicate with a variety of sensory modalities. For example, in close communication, the combination of lipsmack and affiliative calls are more likely elicit the affiliative contact (Micheletta et al. 2012).
Comparison to Other Macaque Species
18
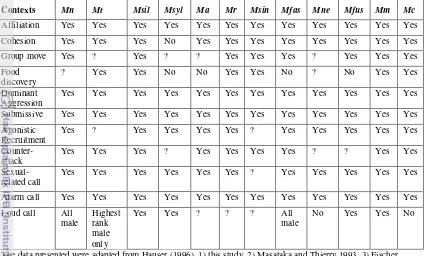
shared in all species in the genus Macaca except M. sylvanus (Fischer and Hammerschidt 2002). Research on M. fuscata (Green 1975) noted the difference in contact coo in the very various contexts includes during foraging. While Sulawesi Macaque such as M. nigra and M. tonkeana (Masataka and Thierry 1993) has two basic acoustic patterns used in three general contexts. The contact calls related to foraging might need further investigation in this species. The second difference is the alarm calls. The alarm call is sex-specific call in M. radiata, male particularly has different alarm call type toward leopard and dog (Hohman 1989). This study suggested that M. nigra has one call type for all of predators but further investigation including detailed acoustic analyses of alarm calls emitted in response to different predators and playback experiments are required before reaching firm conclusions. It thus could not be compared with studies on M. tonkeana in captivity (Masataka and Thierry 1993). The third difference is the loud call. Loud calls among Sulawesi macaques differ in terms of the acoustic pattern. Loud calls of M. tonkeana consist of a single phrase with 2 elements, while M. nigra has one phrase with 3-5 elements. In M. nigra, all males in the group issue loud calls, whereas in M. tonkeana only the highest-ranking male usually produces loud calls. The loud calls in this species are known to convey individual identity and dominance rank (Neumann et al. 2010). The fourth difference is sexually-related calls. Inspection of spectograms shows the difference between female sexually-related call of female M. nigra and female M. tonkeana. The timing of utterance is also different; female M. nigra call after the male has demounted refer as female copulation call, while female M. tonkeana
emitted call during sexual swelling, refers as oestrous call. The presence/absence of call types related to socio-biological context is shown in Table 4 for different macaque species.
Communication is very complex among macaque. Some calls are widely used by some species, but some are absent (Table 4). Each species are suspected to display a different combination of multimodal communication for a particular context. The usage of multimodal communication can either enhance or diminish the intensity of the signal and therefore modulate its meaning (Partan & Marler 1999). For examples, a mix of complex gestures and facial expressions, which may alter the meaning of the calls, accompanies vocalizations in the context of aggression. Thus the acoustic variation alone might not able to explain the social complexity between high and less tolerance species. The future research will be needed in order to get a comprehensive assessment of social communication. In general it might show its differences on social complexity, habitat, or phylogenetic relationships of Sulawesi Macaques.
Implication to Taxonomy and Conservation Issues
19 Table 3 The comparison of Macaque’s vocal communication
Contexts Mn Mt Msil Msyl Ma Mr Msin Mfas Mne Mfus Mm Mc
Affiliation Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes
Cohesion Yes Yes Yes No Yes Yes Yes Yes Yes Yes Yes Yes
Group move Yes ? Yes ? ? Yes Yes Yes ? Yes Yes Yes
Food discovery
? Yes Yes No No Yes Yes No ? No Yes Yes
Dominant Aggression
Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes
Submissive Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes
Agonistic Recruitment
Yes ? Yes Yes Yes Yes ? Yes Yes Yes Yes Yes
Counter-attack
Yes Yes Yes ? Yes Yes Yes Yes ? ? Yes Yes
Sexual-related call
Yes Yes Yes Yes Yes Yes ? Yes Yes Yes Yes Yes
Alarm call Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes Yes
Loud call All
male
Highest rank male onl y
Yes Yes ? ? ? All
male
No Yes Yes No
The data presented were adapted from Hauser (1996), 1) this study, 2) Masataka and Thierry 1993, 3) Fischer and Hammerschmidt 2002). Mn= Macaca nigra; Mt= M. tonkeana; Msil: M. silenus; Msyl= M. sylvanus; Mne= M. nemestrina; Ma= M. arctoides; Mr= M. radiata; Msin= M. sinica; Mfas= M. fascicularis; Mfus= M. fuscata; Mm= M. mulata; Mc; M. cyclopis. The social tolerance decreases from right to the left.
The current taxonomy determination has widely used vocalization data to support the endorsement of the new taxa from the morphological and molecular differences. It had been applied in the Sulawesi tarsier in which the seven species members of this genus can be differentiated successfully based on the morphology, genetics and vocalization (Shekelle 2008). New discovery of
Cercopithecus iomamiensis in Africa was determined by the complete assessment and comparison to other closely related species including the vocal type difference (Hart et al. 2012). Thus, the vocal recognition of the species becomes a crucial part in taxonomic determination.
M nigra is one of the endemic species of Sulawesi macaques. The differences between those species are morphological, genetic and related to their distribution area. The vocal data might also be needed to differentiate between this species and M. nigrescens, which has the similar borderland with M. nigra. The vocal communication has been known to play a role in occurrence of hybridization in Tarsius lariang and and T. dianae in their border habitat in Sulawesi (Merker et al. 2009). The urgent investigations on the vocal communication of all Sulawesi macaque species need to be done for better understanding of each communication system in wild species especially for exploring the probability of hybridization.
20
population estimation could be using vocalization data. The use of this method has more advantages where the observer does not have to habituate animals, no causing unnecessary anxiety of animals as shown by the study of estimation population of the Gibbon (Geissmann and Nijman 2006) and Norwegian ortolan bunting (Adi et al. 2009). Here the differences of call types between two Sulawesi macaques can be used as species recognition. Each call type is needed to explore whether they are differ among individual. Detailed studies on individual or sex differences will be useful for the future population monitoring (species presence, group size, sex composition) in wild habitat.
Furthermore, vocalizations are known to describe the internal condition of primate psychology. The conditions assessed by the context in which the sound issued and wildlife response to certain environmental conditions (Seyfarth and Cheney 2003). Understanding the vocalizations will help people to implement better ecotourism activities while respecting the comfort of the animal itself. Calls in aggressive contexts which represent emotional states of the animal, can be used as a sign for visitors to keep distance from the monkeys. This action could help to avoid redirection behaviour from the monkeys to the visitors, which can lead to severe injury. The low threat calls also often emitted by the female with infant in order to prevent the close proximity of human. When monkeys encounter snake, the alarm call can be used as an alert to the presence of snake to other group member and visitor as well. The information is also useful in the management of wildlife both in the wild and in captivity.
5
CONCLUSIONS
The vocal production of M. nigra is highly variable. The calls were used in the contexts of affiliation, group cohesion, agonism, mating behaviour, presence of predator context and non-social categories for some of the loud calls. The repertoire consists of discrete and graded signals. Acoustic features varied between tonal calls, non-tonal calls and complex screams. The calls that are different between M. nigra and other species are contact calls, alarm calls, loud calls and sexually-related calls. The differences of ecology, geographic, social and behaviour might be the source of variation in macaques’ vocal communication.
REFERENCES
Adi K, Johnson MT, Osiejuk TS. 2009. Acoustic censusing using automatic vocalization classification and identity recognition. J. Acoust. Soc. Am. 127(2): 874–883.
Altmann J. 1974. Observational study of behavior: sampling methods. Behaviour. 49(3):227-267.
21 Bouchet H, Blois-Heulin C, Pellier AS, Zuberbühler K , Lemasson A. 2012.
Acoustic variability and individual distinctiveness in the vocal repertoire of red-capped mangabeys (Cercocebus torquatus). J Comp Psychol 26(1): 45-56. Boissy A, Manteuffel G, BJ Margit, Moe RO, Spruijt B, Keeling LJ, Winckler C,
Forkman B, Dimitrov I, Langbein J, et al. 2007. Assessment of positive emotions in animals to improve their welfare. Physiol Behav 92:375–397. Bradbury JW, Vehrencamp SL. 1998. Principles of Animal Communication.
Sunderland, Massachusetts (US): Sinauer Associates.
Cheney DL, Seyfarth RM. 2010. Production, usage, and comprehension in animal vocalizations. Brain Lang. 115(1):92-100.
Dobson SD. 2012. Coevolution of facial expression and social tolerance in macaques. Am J Primatol. 4(3):229-235. doi: 10.1002/ajp.21991.
Dupjan S, Schön P-C, Puppe B, Tuchscherer A, Manteuffel G. 2008. Differential vocal responses to physical and mental stressors in domestic pigs (Susscrofa). J Applanim. 114:105–115.
Duboscq J, Micheleta J, Agil M, Hodges K, Thierry B, Engelhardt A. 2013. Social tolerance in wild female crested macaques (Macaca nigra) in Tangkoko-Batuangus Nature Reserve, Sulawesi, Indonesia. Am J Primatol. 00:1-15. doi: 10.1002/ajp.22114.
Fichtel C, Hammerschmidt K, Jürgens U. 2001. On the vocal expression of emotion. A multi-parametric analysis of different states of aversion in the Squirrel monkey. Behaviour 138:97-116.
Fischer J, Hammerschmidt K. 2002. An overview of the Barbary macaque,
Macaca sylvanus, vocal repertoire. Folia Primatol. 73(1):32-45.
Fischer J, Metz M, Cheney DL, Seyfarth RM. 2001. Baboon responses to graded bark variants. Anim Behav. 16(5): 925-931.
Fischer J, Noser R, Hammerschmidt K. 2013. Bioacoustic Field Research: A primer to acoustic analyses and playback experiments with primates. Am. J. Primatol. 9999:1-21.
Fitch WT, Hauser MD. 1995. Vocal production in nonhuman primates: acoustics, physiology, and functional constraints on "honest" advertisement. Am J Primatol. 37(3): 191-219.
Fooden J. 1969. Taxonomy and evolution of the monkeys of Celebes (Primates: Cercopithecidae). Basel (CH): Karger.
Freeberg TM, Dunbar RIM, Ord TJ. 2012. Social complexity as a proximate and ultimate factor in communicative complexity. Proc R Soc . 367:1785-1801. Geissmann T, Nijman V. 2006. Calling in Wild Silvery Gibbons (Hylobates
moloch) in Java (Indonesia): behavior, phylogeny, and conservation. Am J Primatol. 68:1-19.
Gouzoules S, Gouzoules H, Marler P. 1984. Rhesus monkey (Macaca mulatta) screams: representational signaling in the recruitment of agonistic aid. Anim Behav. 32(1):182-193.
Green S.1975. Variation in vocal pattern with social situation in the Japanese monkey (Macaca fuscata): a field study. Rosenblum A, editor. Primate behavior: developments in field and laboratory research. New York (US): Academic Pr, p 1-102.
22
Monkey Endemic to the Democratic Republic of Congo and Implications for Conservation of Congo’s Central Basin. Plos one 7(9): e44271. doi:10.1371 /journal.pone.0044271.
Hartwig S. 2005. Individual acoustic identification as a non-invasive conservation tool: An approach to the conservation of the African Wild Dog Lycaonpictus
(Temminck 1820). Bioacoustics. 15: 35-50.
Hauser MD. 1996. Vocal communication in macaques: causes of variation. Fa JE, Lindburg DG. Editor. Evolution and Ecology of Macaque Societies. Cambridge (UK): Cambridge University Press. p: 551-578.
Hohmann G, Herzog MO. 1985.Vocal communication in lion-tailed macaques
(Macaca silenus). Fol Primatol 45:148–178.
Hohmann GM. 1989. Vocal communication of wild bonnet macaques (Macaca radiata). Primates. 30: 325-345.
Hsu MJ, Chen LM, Agoramoorthy G. 2005. The vocal repertoire of Formosan macaques, Macaca cyclopis: Acoustic structure and behavioural context. Zool Stud. 44(2):275-294.
Lewis SA. 1985. The vocal repertoire of the Celebes black ape (Macaca nigra) [disertation]. Georgia (US): University of Georgia.
Maciej P. Ndao I. Hammerschmidt K. Fischer J. 2011. Vocal communication in a complex multi-level society: constrained acoustic structure and flexible call usage in Guinea baboons. Front. Zool.10:58.
Maestripieri D. 2005. Gestural communication in three species of macaques (Macaca mulatta, M. nemestrina, M. arctoides): use of signals in relation to dominance and social context. Gesture. 5(1-2):57-73.
Marler P. 1977. The structure of animal communication sounds. In TH Bullock, ed. Recognition of complex acoustic signals. Berlin (DE): Springer-Verlag, p 17-35.
Masataka N, Thierry B. 1993. Vocal communication of Tonkean macaques in confined environments. Primates. 34(2):169-180.
McComb K, Semple S. 2005. Coevolution of vocal communication and sociality in primates. Biology Lett. 1(4):381-385. doi:10.1098/rsbl.2005.0366.
Merker S, Driller C, Perwitasari-Farajallah D, Pamungkas J, Zischler H. 2009. Elucidating geological and biological processes underlying the diversification of Sulawesi tarsiers. PNAS 106 (21): 8459–8464.
Micheletta J, Waller BM, Panggur MR, Neumann C, Duboscq J, Agil M, Engelhardt A. 2012. Social bonds affect anti-predator behaviour in a tolerant species of macaque, Macaca nigra. Proc R Soc B. 279(1744): 4042-4050.
Micheletta J, Engelhardt A, Matthews L, Agil M, Waller BM. 2013. Multicomponent and multimodal lipsmacking in Crested macaques (Macaca nigra). Am J Primatol. 75(7):763-773.
Micheletta J. 2012. Social communication in crested macaques (Macaca nigra). [dissertation]. Portsmouth (UK): University of Portsmouth.
Neumann C, Assahad G, Hammerschmidt K. Perwitasari-Farajallah D, Engelhardt A. 2010. Loud calls in male crested macaques, Macaca nigra: a signal of dominance in a tolerant species. Anim Behav. 79(1):187-193. doi:10.1016/j.anbehav.2009.10.026.
23 Monje JC, Samyn Y, VandenSpiegel D, editor. Manual on eld recording
techniques and protocols for all taxa biodiversity inventories and monitoring. 8th Volume. Belgium (BE): Belgian Development Cooperation. p 69-99.
Owren MJ, Linker CD. 1995. Some analysis methods that may be useful to acoustic primatologists. Zimmermann E, Newman JD, Jürgens U. Editor.
Current topics in primate vocal communication. New York (US): Plenum. p 1-28.
Palacios JFG, Engelhardt A, Agil M, Hodges K, Bogia R, Waltert M. 2011. Status of, and conservation recommendations for, the critically endangered crested black macaque Macaca nigra in Tangkoko, Indonesia. Oryx. 46(2):290-297. Palombit RA. 1992. A preliminary study of vocal communication in wild
long-tailed macaques (Macaca fascicularis). I. Vocal repertoire and call emission.
Int J Primatol 13(2): 143-182.
Partan SR, Marler P. 1999.Communication goes multimodal. Science. 283(5406): 1272-1273.
Price T, Fischer J. 2013. Meaning attribution in the West African green monkey: influence of call type and context. Anim Cogn. doi:10.1007/s10071-013-0660-9.
Rendall D, Seyfarth RM, Cheney DL, Owren MJ. 1999. The meaning and function of grunt variants in baboons. Anim Behav. 57:583–592.
Rosenbaum B, O'Brien TG, Kinnaird MF, Supriatna J. 1998. Population densities of Sulawesi crested black macaques (Macaca nigra) on Bacan and Sulawesi, Indonesia: effects of habitat disturbance and hunting. Am J Primatol. 44(2):89-106.
Rupell JC. 2009. Vocal diversity and taxonomy of Nomascus in Central Vietnam and Southern Laos. Int J Primatol 31:73–94.
Seyfarth RM , Cheney DL. 2003. Meaning and emotion in animal vocalizations. Ann NY Acad Sci 1000: 32–55.
Schrader L, Hammerschmidt K. 1997. Computer-aided analysis of acoustic parameters in animal vocalizations: a multi-parametric approach. Bioacoustics.
7(4):247-265.
Seyfarth RM, Cheney DL. 2003. Meaning and emotion in animal vocalizations.
Ann NY Acad Sci.1000:32-55.
Shekelle M. 2008. Distribution of Tarsier acoustic form, north and central Sulawesi: with notes on the primary taxonomy of Sulawesi’s tarsiers. Fitch-Snyder H, Groves C. Maryanto I, Schulze, Shekelle M, editors. Primates of the Oriental Night. Bogor (ID): LIPI Press. p 35-50.
Slocombe KE, Townsend SW, Zuberbühler K. 2009. Wild chimpanzees (Pan troglodytes schweinfurthii) distinguish between different scream types: evidence from a playback study. Anim Cogn. 12:441–449.
Sugardjito J, Southwick CH, Supriatna J, Kohlhaas A, Baker SC, Erwin J, Froehlich J, Lerche N. 1989. Population survey of macaques in northern Sulawesi. Am J Primatol. 18(4):285-301.
Supriatna J, Andayani N. 2008. Macaca nigra. In: IUCN 2012. The IUCN Red List of Threatened Species.Version 2012.2.[Internet]. [downloaded on 17th
October 2012]. Available at:
24
Thierry B, Bynum EL, Baker S, Kinnaird MF, Matsumura S, Muroyaman Y, O'Brien TG, Petit O, Watanabe K. 2000. The social repertoire of Sulawesi macaques. Primate Res. 16:203-226.
Thierry B, Singh M, Kaumanns W. 2004. Social Epigenesis. Thierry B, Singh M, Kaumanns W, editor. Macaque societies: a model for the study of social organization. Cambridge (GB): Cambridge Univ Pr. p 267-294.
Wiley RH, Richards DG. 1978. Physical constraints on acoustic communication: implications for the evolution of animal vocalizations. Behav Ecol Sociobiol
25
APPENDICES
Appendix 1 The social tolerance among macaque’s society as adapted from Thierry (2007). The level of tolerance increase from left (grade 1) to right (grade 4). The social tolerance of the species in the bracket is less known.
Grade 1 Grade 2 Grade 3 Grade 4
M. mulatta M. fascicularis M. sylvanus M. tonkeana
M. fuscata M. nemestrina M. silenus M. maura
(M. cyclopis) (M. assamensis) M. arctoides M. nigra
(M. thibetana) M. radiata M. brunnescens
M. sinica M. ochreata
26
27
AUTHOR’S BIOGRAPHY
The author was born in the District of East Manggarai, East Nusa Tenggara Province on August 23, 1985 as the second child of the couple Arnold Panggur and Mathilda Muwa. Authors pursued the Bachelor degree from the Department of Forest Resources Conservation and Ecotourism, Faculty of Forestry, Bogor Agricultural University, graduating in 2009. In 2011, the authors attended graduate school at the Animal Bioscience Master Program, Department of Biology, Bogor Agricultural University. The master program funded by Macaca Nigra Project (a collaborative research program between the German Primate Center and the Bogor Agricultural University).
ABSTRACT
MARIA ROSDALIMA PANGGUR. Vocal Communication of Wild Crested Macaque (Macaca nigra). Supervised by RR DYAH PERWITASARI and ANTJE ENGELHARDT.
Macaca nigra is one of seven Sulawesi macaques’ species, endemic to northern part of Sulawesi. The species is one representative of highly tolerant species of macaques that has received less attention than low tolerance species (e.g. rhesus macaque, M. mulatta). This study presents an overview over the vocal repertoire of wild crested macaques (Macaca nigra) in order to get a comprehensive picture of their communication. The calls were categorized according to the social context in which they were uttered. Each call types within context are presented in the form of spectograms along with descriptive statistics of the different acoustic parameters measured. M. nigra issued 11 call types which were distributed in five distinct social contexts and one loud call. The five social contexts in which the calls were uttered were affiliation, cohesion (including group movement), agonism, predation, and mating contexts. Loud calls were emitted only by males and in various contexts including non-social context. Calls were highly graded, meaning that measures of acoustic parameters overlapped between different contexts. The behavioural contexts and the use of other communication mode can be considered as a source of variation.
Key words: Crested macaque, Macaca nigra, vocal repertoire, social context
ABSTRAK
MARIA ROSDALIMA PANGGUR. Komunikasi Suara pada Macaca nigra. Dibimbing oleh RR DYAH PERWITASARI and ANTJE ENGELHARDT.
terkait perilaku kawin, dan satu tipe suara loud call. Tipe suara loud call
merupakan suara yang dikeluarkan dalam beberapa konteks yang berbeda. Suara
M. nigra tergolong graded, yang berarti bahwa nilai parameter akustik suara-suara saling tumpang tindih antara konteks. Perbedaan konteks sosial dan penggunaan komunikasi multimodal dapat