DINAMIKA KOLONISASI TIGA FUNGI EKTOMIKORIZA
Scleroderma spp. DAN HUBUNGANNYA DENGAN
PERTUMBUHAN TANAMAN INANG
MELYA RINIARTI
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI
DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi yang berjudul “Dinamika Kolonisasi Tiga Fungi Ektomikoriza Scleroderma spp. dan Hubungannya dengan Pertumbuhan Tanaman Inang” adalah karya saya sendiri dengan arahan komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Juli 2010
MELYA RINIARTI. Colonization Dynamics of Three Ectomycorrhizal Fungi Scleroderma spp. and Their Role on Host Plant Growth. Supervised by IRDIKA MANSUR, ARUM SEKAR WULANDARI DAN CECEP KUSMANA.
Ectomycorrhizal symbiosis represented one of the most important mutualistic association in forest ecosystem. Scleroderma spp. have already been known as ectomycorrhizal fungi which could form symbioses with Dipterocarpaceae, Pinaceae, and Gnetaceae. This study was conducted in three steps, (1) Compatibility test of three ectomycorrhizal fungi Scleroderma spp. under in vitro condition. The aim of this study was to determine the mycelial antagonism and interaction patterns among the fungus under in vitro condition. Three species Scleroderma spp. were cultured on solid Modified Melin Norkrans (MMN) medium. Each fungus was cultured as single, pairs and triple combinations. A completely randomized design were used with 7 treatments and 8 replicates Petri dishes. We found that the radial growth of S. sinnamariense mycelia was two times faster than S. columnare and S. dictyosporum. There was no antagonism pattern between fungal mycelia when they grown together. (2) Colonization dynamics of three ectomycorrhizal fungi Scleroderma spp. and their relation to host plant growth. The aim of this study was to determine the compatibility and specificity of each ectomycorrhizal fungi with the host plants, and the effect on host plants growth. S. columnare, S. dictyosporum and S. sinnamariense were inoculated in single-, two-, and three- species treatments to Shorea pinanga, Pinus merkusii and Gnetum gnemon seedlings using spores suspension. After 6, 8, and 10 months of inoculation, seedlings were harvested to determine colonization percentage, height and diameter growth, and plant biomass. The results revealed that S. sinnamariense did not form association with S. pinanga and P. merkusii but form association with G. gnemon. On the other hand, S. columnare and S. dictyosporum both in single- and multi- species treatments could form association with all the host plants. Generally, Scleroderma spp. increased the growth of host plants. The last was (3) Root characteristics of ectomycorrhizal fungi on S. pinanga, P. merkusii, and G. gnemon. The aim of this study was to determine the morphology (hyphae colour, branching pattern, clamp-connection) and anatomy (mantel and Hartig net) of ectomycorrhizal roots in each host plants which associated with the ectomycorrhizal fungus. This result revealed that morphology were affected by fungi, while branching pattern and Hartig net were affected by plants. S. pinanga dan G. gnemon were have monopodial branching pattern which P. merkusii was dichotomously branching pattern. The depth of mantels and Hartig net were increase by time. The Hartig net was only at
epidermis S. pinanga and G. gnemon, while its penetrated in cortex on P. merkusii.
MELYA RINIARTI. Dinamika Kolonisasi Tiga Fungi Ektomikoriza Scleroderma spp. dan Hubungannya dengan Pertumbuhan Tanaman Inang. Di bawah bimbingan IRDIKA MANSUR, ARUM SEKAR WULANDARI DAN CECEP KUSMANA.
Fungi ektomikoriza memainkan peranan penting dalam berbagai ekosistem, melalui keterlibatannya dalam siklus karbon dan nutrisi, jejaring makanan di bawah tanah, serta dinamika komunitas tanaman. Fungi ektomikoriza merupakan salah satu komponen penting dalam ekosistem hutan, dan diketahui membentuk simbiosis dengan sebagian besar tanaman kehutanan. Scleroderma merupakan genus fungi ektomikoriza yang diketahui dapat membentuk simbiosis dengan Dipterocarpaceae, Pinaceae, dan Gnetaceae. Simbiosis dengan fungi ektomikoriza akan membantu pertumbuhan dan ketahanan hidup tanaman inang karena fungi ektomikoriza akan membantu dalam penyerapan unsur hara dan air, meningkatkan ketahanan terhadap patogen dan meningkatkan ketahanan terhadap kekeringan. Beberapa fungi ektomikoriza juga menghasilkan tubuh buah yang dapat dikonsumsi (edible mushroom).
Penelitian ini dilaksanakan menggunakan tiga tanaman inang yang mewakili tiga famili, yaitu Shorea pinanga (Dipterocarpaceae-Angiospermae); Pinus merkusii (Pinaceae-Gymnospermae) dan Gnetum gnemon (Gnetaceae-peralihan gymnospermae ke angiospermae). Fungi ektomikoriza Scleroderma spp. yang digunakan adalah jenis yang secara alami bersimbiosis dengan ketiga tanaman inang yang digunakan yaitu: S. columnare bersimbiosis dengan S. pinanga; S. dictyosporum bersimbiosis dengan P. merkusii dan S. sinnamariense bersimbiosis dengan G. gnemon. Tujuan umum penelitian ini ialah mempelajari interaksi antar fungi ektomikoriza pada tanaman inang yang sama, yang berkaitan dengan fungsi dan peranan masing-masing fungi ektomikoriza sehubungan dengan upaya peningkatan pertumbuhan dan ketahanan hidup tanaman inang.
Penelitian dilaksanakan dalam tiga tahapan. Penelitian 1 berjudul “Uji Kompatibilitas Tiga Jenis Fungi Ektomikoriza Scleroderma spp. Secara In Vitro”. Penelitian ini bertujuan untuk memperoleh informasi tentang pertumbuhan dan perkembangan serta hubungan yang terbentuk antar fungi ektomikoriza Scleroderma spp. secara in vitro. Pada penelitian ini ketiga jenis Scleroderma spp. dibiakkan pada media kultur MMN padat (Modified Melin Norkrans). Perlakuan yang diberikan ialah pembiakan miselium Scleroderma spp. secara terpisah, dua isolat bersamaan dan tiga isolat bersamaan. Penelitian ini disusun dalam Rancangan Acak Lengap (RAL) dengan 7 perlakuan dan delapan ulangan. Peubah yang diamati ialah kecepatan awal waktu pertumbuhan miselium dan diameter pertumbuhan koloni masing-masing fungi ektomikoriza.
dengan menggunakan suspensi spora (jumlah spora 25 x 10 ), dengan perlakuan inokulasi tunggal, dual dan triple fungi ektomikoriza, sehingga untuk tiap jenis tanaman inang didapat 8 perlakuan. Pengamatan terhadap persen kolonisasi dan pertumbuhan tanaman (pertambahan tinggi, pertambahan diameter, berat kering akar, pucuk dan total, serta penyerapan N dan P) dilakukan pada bulan ke-6, 8, dan 10 setelah inokulasi. Rancangan yang digunakan ialah Rancangan Acak Lengkap untuk tiap jenis tanaman dengan 8 perlakuan dan 3 ulangan, masing-masing ulangan terdiri atas 5 tanaman.
Penelitian 3 (bab V) berjudul “Karakterisitik Akar Berektomikoriza Pada Shorea pinanga, Pinus merkusii, dan Gnetum gnemon”. Tujuan penelitian ini ialah untuk memperoleh informasi tentang karakteristik morfologi dan anatomi akar pada S. pinanga, P. merkusii dan G. gnemon yang telah diinokulasi dengan tiga jenis Scleroderma spp. selama 6, 8, dan 10 bulan. Bahan yang digunakan ialah akar berektomikoriza segar dan akar hasil proses histologi. Parameter yang diamati ialah warna miselium, bentuk percabangan, clamp connection, dan ketebalan mantel, serta kedalaman Hartig net.
Hasil Penelitian 1 menunjukkan bahwa S. sinnamariense adalah fungi ektomikoriza yang kecepatan perkembangan dan diameter pertumbuhan koloninya mencapai dua kali lipat dibandingkan dengan S. columnare dan S. dictyosporum pada media MMN padat. Saat dibiakkan secara bersama-sama, ketiga jenis fungi ektomikoriza dapat tumbuh dengan baik tanpa adanya hambatan pertumbuhan satu dengan lainnya. Hal ini ditunjukkan dengan tidak adanya barrier pada miselium ketiga fungi tersebut, yang menunjukkan tidak ada sifat antagonis di antara ketiganya. Hasil pengamatan pada hifa yang tumbuh diantara fungi menunjukkan pertumbuhan yang normal, ditandai dengan tidak adanya lisis pada dinding hifa.
Hasil penelitian 2 menunjukkan adanya spesifisitas yang tinggi pada S. sinnamariense. Fungi ektomikoriza ini hanya dapat bersimbiosis dengan G. gnemon yang merupakan tanaman inang alaminya. Sementara S. columnare dan S. dictyosporum dapat membentuk simbiosis dengan ketiga tanaman inang yang diujicobakan. Pada ketiga tanaman inang, S. columnare dan S. dictyosporum baik pada perlakuan tunggal maupun secara bersamaan, mampu meningkatkan kolonisasi dan pertumbuhan tanaman inang, sedangkan S. sinnamariense hanya dapat berkontribusi pada G. gnemon. Perlakuan multi fungi ektomikoriza tampaknya memberikan hasil yang lebih baik dalam penyerapan N dan P dibandingkan dengan perlakuan tunggal dan kontrol.
Hasil penelitian 3 menunjukkan bahwa karakteristik morfologi (warna dan clamp connection) dipengaruhi oleh fungi, sementara bentuk percabangan akar berektomikoriza dan kedalaman Hartig net merupakan karakteristik yang lebih dipengaruhi oleh jenis tanaman, pada P. merkusii percabangan berbentuk dikotomus dan Hartig net mencapai kortek. Pada S. pinanga dan G. gnemon bentuk percabangan akar berektomikoriza adalah monopodial dengan Hartig net hanya mencapai jaringan epidermis. Ketebalan mantel dan kedalaman Hartig net meningkat seiring dengan waktu.
© Hak cipta milik IPB, tahun 2010
Hak cipta dilindungi undang-undang
1. Dilarang mengutip sebagian atau seluruh karya disertasi tanpa mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar Institut Pertanian Bogor
DINAMIKA KOLONISASI TIGA FUNGI EKTOMIKORIZA
Scleroderma spp. DAN HUBUNGANNYA DENGAN
PERTUMBUHAN TANAMAN INANG
MELYA RINIARTI
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Ilmu Pengetahuan Kehutanan
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
Penguji Luar Komisi :
Sidang Tertutup :
1. Dr. Ir. Elis Nina Herliyana, M.Si. 2. Dr. Ir. Sri Wilarso Budi R.
Sidang Terbuka
1. Dr. Ir. Nampiah Sukarno
spp. dan Hubungannya dengan Pertumbuhan Tanaman Inang
Nama : Melya Riniarti NRP : E061050011
Program Studi : Ilmu Pengetahuan Kehutanan
Disetujui Komisi Pembimbing
Dr. Ir. Irdika Mansur, M.For.Sc Ketua
Dr. Ir. Arum Sekar Wulandari, MS
Anggota Prof. Dr. Ir. Cecep Kusmana, MS Anggota
Diketahui
Ketua Program Studi Ilmu Pengetahuan Kehutanan
Prof. Dr. Ir. Imam Wahyudi, MS.
Dekan Sekolah Pasca Sarjana
Prof. Dr. Ir. Khairil Anwar Notodiputro, MS.
Puji syukur penulis panjatkan ke hadirat Allah SWT, karena hanya dengan rahmat dan ridho-Nya penelitian dengan judul “Dinamika Kolonisasi Tiga Fungi Ektomikoriza Scleroderma spp. dan Hubungannya dengan Pertumbuhan Tanaman Inang” dapat diselesaikan dengan baik.
Pada kesempatan ini, penulis menyampaikan penghargaan dan mengucapkan terima kasih kepada Dr. Ir. Irdika Mansur, M.For.Sc., Prof. Dr. Ir. Cecep Kusmana, M.S., dan Dr. Ir. Arum Sekar Wulandari M.S. selaku komisi pembimbing. Dr. Istomo, Dr. Elis Nina Herliyana , Dr. Sri Wilarso Budi R, Dr. Nampiah Sukarno dan Dr. Made Hesti Lestari Tata selaku penguji prelim, penguji luar komisi dalam ujian tertutup, dan Penguji luar komisi dalam ujian terbuka. Dirjen Pendidikan Tinggi, Departemen Pendidikan Nasional RI, atas bantuan Beasiswa Pendidikan Pascasarjana (BPPS) dan DP3M Dikti atas Hibah Doktor 2009. Rektor, Dekan Sekolah Pascasarjana IPB dan segenap civitas akademika IPB atas kesempatan studi yang diberikan. Segenap staf pengajar dan manajemen Program Studi Ilmu Pengetahuan Kehutanan IPB. Rektor Universitas Lampung, Dekan Fakultas Pertanian Unila, dan Jurusan Kehutanan Unila yang telah memberikan kesempatan bagi penulis untuk melanjutkan tugas belajar. Dr. Supriyanto (Departemen Silvikultur IPB), Dr. Dorly (Jurusan Biologi IPB), Ibu Endang (LIPI Cibinong), Mbak Faiq, Mbak Susan, Ibu Esti, Pak Yadi, Pak Atang dan Bi Ita, dan berbagai pihak dari berbagai laboratorium yang telah membantu dalam pelaksanaan penelitian ini. Pak Julius dan keluarga, Pak Budi Sulistyawan dan keluarga, Mbak Dina, Ibu Diana, Arida, Jusuf Dibisono, Suhasman, Syahidah, Pak Abimanyu, , Pak Iskandar, Ibu Widi, Teh Nur, Mbak Yuniar, Rara, Nani, Yano, Warji dan keluarga, dan seluruh rekan-rekan SPS IPB dan Unila Keluarga besar Ir. S. Hasan Syahrin dan Achadi (Alm.) atas dukungan dan kasih sayangnya. Suami, Achmad Baehaqi S.P., M.Si., dan ketiga anakku, Achmadyan Raya, Adinda Tami Rahmani, dan Aqilya Puti Gemilang, untuk segala dukungan, pengorbanan dan kasih sayang.
Akhirnya penulis menyadari bahwa penelitian ini masih banyak keterbatasan dan kekurangannya. Namun demikian, penulis mengharapkan tulisan ini dapat bermanfaat khususnya bagi penulis sendiri dan orang-orang yang memerlukannya terlebih lagi bagi perkembangan dunia pendidikan dan ilmu pengetahuan.
Bogor, Juli 2010
Pada kalian
Yang selalu berkata aku pasti bisa Ketika langkahku menyurut
Jalan ini tampaknya akan segera kusudahi Dengan kemenangan di tangan
Walau tiada gemilang dengan kilau Namun menorehkan sejarah dalam hidupku Karena perjalanan ini menganugerahiku Banyak cinta untuk diterima dan dibagi Terima kasih untuk kalian
Yang selalu berkata kami bangga padamu Ketika aku meragukan diriku
Karena tanpa segala dukungan dan doa Tak kan mampu aku singkirkan
Segala kerikil dan terjalnya pendakianku Terima kasih untuk kalian…
Segenap cinta dan kasih Tak kan mampu membalas Segala yang telah kalian berikan Pada ku dan keluarga ku
Selama menempuh perjalanan ini….. Kepada Allah lah segalanya ku pinta…. Untuk membalas…..
RIWAYAT HIDUP
Penulis dilahirkan di Palembang, pada tanggal 3 Mei 1977 sebagai anak pertama dari lima bersaudara pasangan Ir. S. Hasan Syahrin dan Maryati.
Penulis menyelesaikan pendidikan SLTA di Sekolah Menengah Atas Negeri 2 Bandar Lampung pada tahun 1994. Pada tahun yang sama penulis diterima di Program Studi Ilmu Tanah Fakultas Pertanian Universitas Lampung (Unila) melalui jalur PMKA dan memperoleh gelar sarjana pada tahun 1998. Pada tahun 1999 penulis melanjutkan jenjang pendidikan S2 di Program Studi Ilmu Pengetahuan Kehutanan Program Pascasarjana IPB dan mendapatkan gelar Magister Sains pada tahun 2002. Tahun 2005 penulis mendapat kesempatan meneruskan pendidikan pada Program Studi Ilmu Pengetahuan Kehutanan pada Sekolah Pascasarjana IPB. Beasiswa pendidikan pascasarjana diperoleh melalui program BPPS dari Ditjen Dikti Departemen Pendidikan Nasional
Sejak tahun 2002 sampai dengan sekarang penulis bekerja sebagai staf pengajar di Jurusan Kehutanan Fakultas Pertanian Universitas Lampung. Selama mengikuti program S3 penulis menjadi anggota AMI (Asosiasi Mikoriza Indonesia) dan mengikuti berbagai kegiatan seminar baik nasional maupun internasional yang berhubungan dengan penelitian dan keilmuan. Beberapa hasil penelitian S3 telah dipublikasikan pada berkala ilmiah yaitu (1) Uji kompetisi tiga fungi ektomikoriza Sclerodema spp. secara in vitro. Vegetasi (7):63—67. Dan (2) Karakteristik Akar Berektomikoriza pada Shorea pinanga, Pinus merkusii dan Gnetum gnemon. Perennial (6):11—19.
Halaman
DAFTAR TABEL ... ii
DAFTAR GAMBAR ... iii
DAFTAR LAMPIRAN ... iv
1 PENDAHULUAN ... 1
2 TINJAUAN PUSTAKA ... 7
3 UJI KOMPATIBILITAS TIGA JENIS FUNGI EKTOMIKORIZA Scleroderma spp. SECARA IN VITRO ... 29
Abstrak ... 29
Abstract ... 29
Pendahuluan ... 29
Metode Penelitian ... 31
Hasil dan Pembahasan ... 33
Kesimpulan ... 37
Daftar Pustaka ... 37
4 DINAMIKA KOLONISASI TIGA FUNGI EKTOMIKORIZA Scleroderma spp. DAN HUBUNGANNYA DENGAN PERTUMBUHAN TANAMAN INANG ... 39
Abstrak ... 39
Abstract ... 39
Pendahuluan ... 40
Metode Penelitian ... 42
Hasil dan Pembahasan ... 47
Kesimpulan ... 59
Daftar Pustaka ... 59
5 KARAKTERISTIK AKAR BEREKTOMIKORIZA PADA Shorea pinanga, Pinus merkusii, dan Gnetum Gnemon ... 63
Abstrak ... 63
Abstract ... 63
Pendahuluan ... 63
Metode Penelitian ... 65
Hasil dan Pembahasan ... 66
Kesimpulan ... 76
Daftar Pustaka ... 77
6 PEMBAHASAN UMUM ... 79
7 KESIMPULAN DAN SARAN ... 85
Halaman 1. Nama spesies Scleroderma, yang dikelompokkan berdasarkan bentuk
spora dan keberadaan clamp-connections ... 11 2. Daftar penelitian fungi ektomikoriza Scleroderma spp. pada berbagai
tanaman inang dari Dipterocarpaceae, Pinaceae dan Gnetaceae ... 18 3. Rincian perlakuan yang dilakukan secara in vitro ... 33 4. Rincian perlakuan inokulasi fungi ektomikoriza pada tiga jenis tanaman
inang ... 46 5. Nilai rata-rata pengamatan histologi akar P. merkusii yang diinokulasi
oleh S. columnare dan S. dictyosporum pada bulan ke 6, 8, dan 10 ... 67 6. Nilai rata-rata pengamatan histologi akar G. gnemon yang diinokulasi
oleh S. columnare, S. dictyosporum dan S. sinnamariense pada bul ke 6, 8, dan 10 ... 70 7. Nilai rata-rata pengamatan histologi akar S. pinanga yang diinokulasi
Halaman
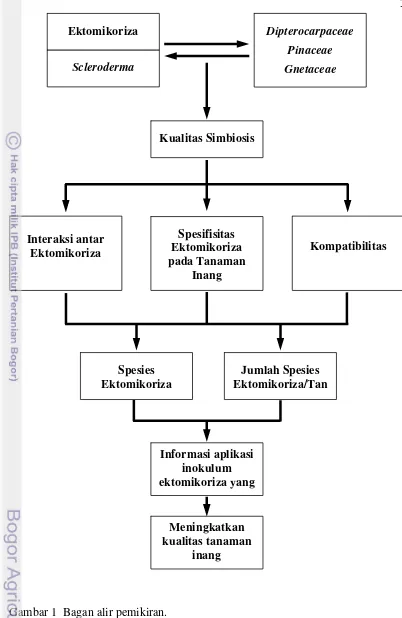
1. Bagan alir pemikiran ... 3
2. Simbiosis ektomikoriza pada tanaman ... 9
3. Empat tipe morfotipe spora Scleroderma dilengkapi dengan ornamen-ornamennya ... 10
4. Jumlah makalah jurnal dari tahun 2000 – 2008 yang berhubungan dengan mikoriza di hutan tropis (kata kunci pencarian: mycorrhiza & tropical & forest), dibandingkan dengan total literatur tentang mikoriza (kata kunci pencarian: mycorrhiza) ... 17
5. Bahan yang digunakan dalam penyiapan miselium fungi ... 32
6. Penempatan inokulum miselium fungi ektomikoriza pada cawan Petri . 32 7. Perhitungan diameter pertumbuhan koloni dalam cawan Petri dengan menggunakan delapan arah mata angin ... 33
8. Perkembangan diameter koloni tiga jenis fungi Scleroderma dengan beberapa perlakuan pada setiap minggu pengamatan ... 33
9. Diameter pertumbuhan koloni pada setiap minggu pengamatan ... 35
10. Tiga jenis tanaman inang yang digunakan dalam penelitian ... 41
11. Tiga fungi ektomikoriza yang digunakan dalam penelitian ... 44
12. Kolonisasi fungi ektomikoriza Scleroderma pada tanaman inang ... 48
13. Persentase kolonisasi ektomikoriza pada S. pinanga, P. merkusii, dan G. gnemon pada 6, 8, dan 10 bulan setelah inokulasi fungi ektomikoriza ... 49
14. Pertambahan tinggi dan diameter pada S. pinanga, P. merkusii, dan G. gnemon pada 6, 8, dan 10 bulan setelah inokulasi fungi ektomikoriza.... 51
15. Berat kering total, berat kering akar dan berat kering pucuk S. pinanga, P. merkusii, dan G. gnemon pada 6,8, dan 10 bulan setelah inokulasi fungi ektomikriza ... 52
16. Penyerapan N dan P pada S. pinanga, P. merkusii, dan G. gnemon pada 6, 8, dan 10 bulan setelah inokulasi dengan fungi ektomikoriza ... 54
17. Ujung akar berektomikoriza ... 67
merkusii berektomikoriza dengan beberapa perlakuan fungi ektomikoriza
dan umur inokulasi ... 69
20. Komposisi ketebalan mantel, Hartig net dan jaringan akar pada akar G. gnemon berektomikoriza dengan beberapa perlakuan fungi ektomikoriza dan umur inokulasi ... 70
21. Komposisi ketebalan mantel, Hartig net dan jaringan akar pada akar S. pinanga berektomikoriza dengan beberapa perlakuan fungi ektomikoriza dan umur inokulasi ... 71
22. Citra SEM akar berektomikoriza ... 72
23. Citra SEM ujung akar berektomikoriza ... 73
Halaman 1. Komposisi media MMN ... 101
2. Analisis histologi akar ektomikoriza ... 102 3. Teknik pewarnaan akar ektomikoriza ... 103 4. Hasil analisis korelasi antara berat kering total tanaman dengan
Latar Belakang
Sebagian besar tanaman kehutanan sangat bergantung pada mikoriza untuk kelangsungan pertumbuhan dan perkembangannya (Nehls et al. 2007). Mikoriza merupakan bentuk hubungan simbiosis mutualisme antara fungi dengan akar tumbuhan tingkat tinggi, tanaman inang memperoleh hara nutrisi sedangkan fungi memperoleh senyawa karbon hasil fotosintesis (Smith dan Read 2008). Mikoriza akan membantu penyerapan unsur hara (Allen et al. 2003; Dehlin et al. 2004; Lilleskov et al. 2002; Baghel et al. 2009), meningkatkan ketahanan terhadap kekeringan (Dell 2002; Dunabeitia et al. 2004), meningkatkan ketahanan terhadap penyakit (Onguene dan Kuyper 2002; Whipps 2004), dan pada beberapa spesies fungi ektomikoriza menghasilkan tubuh buah yang dapat dikonsumsi (edible mushroom) (Hall et al. 2003; Wulandari 2002; Yamada et al. 2001; 2007).
Fungi ektomikoriza memiliki kecenderungan untuk lebih spesifik dalam membentuk simbiosis, dan umumnya hanya pada tanaman berkayu (Brundrett 2002) walaupun terdapat laporan adanya fungi ektomikoriza pada perdu dan semak (Bidartondo et al. 2003; Dickie et al. 2004; Nara dan Hogetsu 2004). Fungi ektomikoriza merupakan komponen penting dalam ekosistem hutan dan umum ditemukan pada hutan-hutan tropis di daerah Asia (Simard dan Durral 2004; Dell 2002; Brearly et al. 2007; Amaronpitak et al. 2006).
2007; Irianto 2004; Obase et al. 2009), dan Gnetaceae (Watling et al. 2002; Wulandari 2002; Suhardi 1997) telah banyak dilaporkan, meskipun demikian masih belum banyak yang diketahui tentang interaksi antar fungi ektomikoriza dan spesifisitas tanaman inang dengan kualitas simbiosis yang terbentuk yang berhubungan dengan kompatibilitas antara tanaman inang dan fungi, dan kemampuannya dalam meningkatkan kualitas tanaman inang, sehingga perlu dikaji lebih dalam (Gambar 1).
Munculnya interaksi antar fungi ektomikoriza dan spesifisitas pada tanaman inang berhubungan dengan tingginya jumlah fungi ektomikoriza yang ada, menurut perkiraan Rinaldi et al. (2008) ada sekitar 20.000—25.000 spesies fungi ektomikoriza yang berasal dari 343 genus, sementara hanya ada 8000 tanaman yang dapat bersimbiosis dengan fungi ini. Berbagai hasil penelitian menemukan adanya berbagai spesies fungi ektomikoriza yang dapat berasosiasi dengan akar tanaman yang sama (Walbert et al. 2009; Valentine et al. 2004; Tedersoo et al. 2006; Obase et al. 2009; Richard et al. 2004). Hal ini akan menimbulkan interaksi antar fungi ektomikoriza (Koide et al. 2005), interaksi ini dapat bersifat saling melengkapi (Newsham et al. 1995; Mollina et al. 1992) atau kompetisi (Kennedy et al. 2007a; Kennedy dan Bruns 2005; Kennedy et al. 2007b; Hortal et al. 2008).
Dipterocarpaceae
Pinaceae
Gnetaceae
Ektomikoriza
Gambar 1 Bagan alir pemikiran. Scleroderma
Kualitas Simbiosis
Spesifisitas Ektomikoriza pada Tanaman
Inang Interaksi antar
Ektomikoriza Kompatibilitas
Spesies Ektomikoriza
Jumlah Spesies Ektomikoriza/Tan
Informasi aplikasi inokulum ektomikoriza yang
Meningkatkan kualitas tanaman
Hedh et al. (2009) menemukan dari 35 strain Paxillus involutus yang diujicobakan pada tanaman birch dan spruce terdapat beberapa strain yang hanya kompatibel dengan salah satu tanaman, dan strain yang kompatibel dengan kedua tanaman inang. Spesifisitas fungi ektomikoriza akan berhubungan dengan kompatibilitas antara fungi ektomikoriza dan tanaman inang, kesesuaiannya akan mempengaruhi optimalisasi fungsi dan peran fungi ektomikoriza dalam membantu pertumbuhan tanaman inang.
Pemilihan ketiga spesies tanaman yang digunakan dalam penelitian ini didasarkan pada pertimbangan bahwa masing-masing spesies mewakili tiga kelas yang berbeda yaitu angiospermae (Shorea pinanga), gymnospermae (Pinus merkusii), dan peralihan antara angiospermae dan gymnospermae (Gnetum gnemon) sehingga diharapkan dapat menjabarkan secara luas hubungan antara fungi ektomikoriza dengan berbagai spesies tanaman.
Fungi ektomikoriza Scleroderma spp. yang digunakan adalah spesies yang secara alami ditemukan pada ketiga tanaman inang yang digunakan yaitu: Scleroderma columnare bersimbiosis dengan S. pinanga; S. dictyosporum bersimbiosis dengan P. merkusii dan S. sinnamariense bersimbiosis dengan G. gnemon. S. columnare, S. dictyosporum dan S. sinnamariense merupakan spesies fungi ektomikoriza yang dikenal sebagai “early-stage ectomycorrhiza” (Supriyanto dan Irawan 1997) yaitu spesies fungi ektomikoriza yang membentuk simbiosis pada awal pertumbuhan tanaman inang.
ketergantungan yang tinggi terhadap fungi ektomikoriza, sehingga dapat mengoptimalkan peranan ektomikoriza dalam meningkatkan pertumbuhan tanaman.
Tujuan Penelitian Tujuan umum penelitian ini ialah:
Mempelajari interaksi antar fungi ektomikoriza pada tanaman inang yang sama, yang berkaitan dengan fungsi dan peranan masing-masing fungi ektomikoriza sehubungan dengan upaya peningkatan pertumbuhan dan ketahan hidup tanaman inang.
Tujuam khusus penelitian ini adalah:
(1) Mempelajari pertumbuhan dan perkembangan serta hubungan yang terbentuk antara S. columnare, S. dictyosporum dan S. sinnamariense secara in vitro.
(2) Mempelajari kompatibilitas dan spesifisitas fungi ektomikoriza S. columnare, S. dictyosporum dan S. sinnamariense terhadap S. pinanga. P. merkusii dan G. gnemon.
(3) Mempelajari hubungan antara fungi ektomikoriza S. columnare, S. dictyosporum dan S. sinnamariense terhadap S. pinanga. P. merkusii dan G. gnemon.
(4) Mempelajari karakteristik morfologi dan anatomi akar S. pinanga, P. merkusii, dan G. gnemon yang telah diinokulasi dengan S. columnare, S. dictyosporum, dan S. sinnamariense selama 6, 8 dan 10 bulan.
Manfaat Penelitian
Hasil penelitian diharapkan akan memberikan manfaat sebagai berikut:
(2) Bagi Pemerintah, khususnya bidang kehutanan, hasil penelitian ini diharapkan dapat memberikan informasi tentang keunggulan penggunaan fungi ektomikoriza pada tanaman kehutanan sehingga dapat dijadikan pedoman dalam kegiatan-kegiatan penanaman di lahan-lahan hutan.
(3) Bagi masyarakat diharapkan dapat memberikan informasi kegunaan fungi ektomikoriza pada berbagai tanaman sehingga memperbaiki kualitas tanaman.
Hipotesis Hipotesis yang diajukan pada penelitian ini adalah:
(1) Terjadi interaksi dalam pertumbuhan dan perkembangan S. columnare, S. dictyosporum, dan S. sinnamariense secara in vitro.
(2) Fungi ektomikoriza S. columnare, S. dictyosporum, dan S. sinnamariense dapat membentuk simbiosis dengan S. pinanga, P. merkusii dan G. gnemon.
(3) Aplikasi fungi ektomikoriza akan meningkatkan pertumbuhan S. pinanga, P. merkusii dan G. gnemon.
Ektomikoriza
Mikoriza merupakan bentuk hubungan simbiosis mutualistik antara fungi dengan akar tumbuhan tingkat tinggi, tanaman inang memperoleh hara nutrisi sedangkan fungi memperoleh senyawa karbon hasil fotosintesis (Smith dan Read 2008). Istilah tersebut pertama kali diperkenalkan oleh Frank pada tahun 1877 di Jerman (Brundrett 2004). Saat ini diketahui terdapat 7 tipe mikoriza yaitu (1) arbuskular mikoriza, (2) ektomikoriza, (3) ektendomikoriza, (4) arbutoid mikoriza, (5) monotropoid mikoriza, (6) ericoid mikoriza, dan (7) orchid mikoriza. Pembagian ini didasarkan pada karakter-karakter (1) ada/tidaknya septa; (2) intraseluler kolonisasi (3) keberadaan mantel dan Hartig net serta (4) acrophyl (Smith dan Read 2008).
Fungi ektomikoriza umumnya dari golongan Basidiomisetes dan
Askomisetes. Beberapa genera fungi Basidiomisetes pembentuk ektomikoriza di antaranya adalah Amanita, Boletellus, Boletinus, Boletus, Pisolithus, Scleroderma, Suillus, Russula, dan Laccaria (Brundrett et al. 1996; Rinaldi et al. 2008).
patogen akar, perlindungan ini terjadi karena adanya lapisan hifa sebagai pelindung fisik dan antibiotika yang dikeluarkan oleh mikoriza (Whipps 2004; Martin-Pinto et al. 2006; Zadworny et al. 2007); dan (4) menghasilkan beberapa zat pengatur tumbuh. Fungi mikoriza dapat menghasilkan hormon auksin, sitokinin, gibberelin, dan vitamin yang bermanfaat untuk inangnya (Allen et al. 2003; Dell 2002). Auksin dapat berfungsi untuk mencegah atau menghambat proses penuaan dan suberinasi akar sehingga umur dan fungsi akar dapat diperpanjang. Manfaat lainnya ialah (5) beberapa fungi ektomikoriza menghasilkan tubuh buah yang dapat dimakan/dikonsumsi oleh manusia, sehingga memberikan hasil hutan non kayu yang bernilai ekonomi dan gizi yang tinggi (Hall et al. 2003; Yamada et al. 2001; 2007).
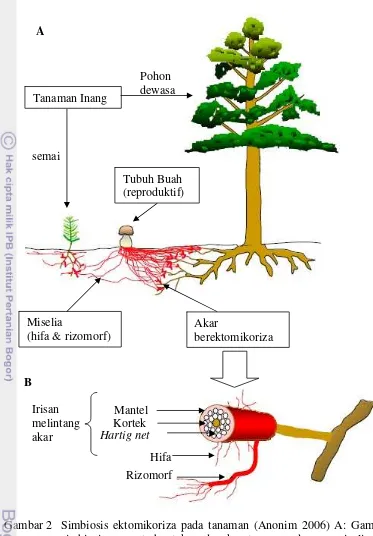
Interaksi antara fungi dan inang dalam medium pertumbuhan sangatlah kompleks dan dipengaruhi oleh sejumlah interaksi biokimia, fisiologi dan proses lingkungan. Simbiosis ektomikoriza pada tanaman disajikan pada Gambar 2.
Scleroderma
Gambar 2 Simbiosis ektomikoriza pada tanaman (Anonim 2006) A: Gambaran simbiosis yang terbentuk pada akar tanaman dengan miselia fungi ektomikoriza, miselia fungi ektomikoriza dapat menghubungkan dua tanaman yang berbeda. B: Komponen fungi ektomikoriza yang terbentuk pada akar tanaman, yang terdiri dari mantel pada lapisan luar akar tanaman, Hartig net yang terbentuk di sela-sela epidermis dan/atau hingga kortek, serta hifa eksternal.
Tanaman Inang
semai
Pohon dewasa
Tubuh Buah (reproduktif)
Miselia
(hifa & rizomorf) Akar berektomikoriza
Hifa Rizomorf Mantel Kortek Hartig net Irisan
melintang akar
A
.
Gambar 3 Empat tipe morfotipe spora Scleroderma dilengkapi dengan ornament-ornamennya (a) Echinulate, (b) Verrucose, (c) Subreticulate, dan (d) Reticulate (Chen 2006)
Tabel 1 Nama spesies Scleroderma, yang dikelompokkan berdasarkan bentuk spora dan keberadaan clamp-connection (Rinaldi et al. 2008).
No. Nama Spesies
Kelompok Aculcatispora – echinulate atau verrucose, tidak reticulate; tanpa clamp-connection
1 S. albidum Pat. Et Trab. 2 S. areolatum Ehrenb. 3 S. cepa Pers.
4 S. columnare Berk. & Broome 5 S. cyaneoperidiatum Watling & Sims 6 S. laeve Lloyd
7 S. leptopodium (Durieu & Mont.) De Toni 8 S. mayama Grgurinovic
9 S. texense Berk.
10 S. uruguayense (Guzmán) Guzmán 11 S. verrucosum Pers.
Kelompok Caloderma – sedikit reticulate dengan duri sangat ramping dan pendek; dengan clamp-connection
12 S. stellatum Berk.
Kelompok Sclerangium – Subreticulate ; dengan clamp-connection 13 S. bermudense Coker.
Kelompok Scleroderma – reticulate sempurna; dengan clamp-connection 20 S. bougheri Trappe, Castellano & Giachini
21 S. bovista Fr.
22 S. congolense Demoulin & Dring 23 S. dictyosporum Pat.
24 S. fuscum (Corda) E. Fisch. 25 S. hypogeum Zeller
26 S. meridionale Dem. & Mal. 27 S. michiganense (Guzmán) Guzmán 28 S. schmitzii Demoulin & Dring 29 S. septentrionale Jeppson 30 S. sinnamariense Mont.
Scleroderma columnare , S. dictyosporum, dan S. sinnamariense
Species S. columnare, S. dictyosporum dan S. sinnamariense dapat diklasifikasikan sebagai berikut (Brundrett et al. 1996; The National Science Museum 1998):
Divisi : Basidiomycota Kelas : Holobasidiomycetes Anak kelas : Gasteromycetes Bangsa : Sclerodermatales Famili : Sclerodermataceae Genus : Scleroderma
Species : Scleroderma columnare Scleroderma dictyosporum Scleroderma sinnamariense
Menurut Zeller (1948) S. columnare telah ditemukan sejak tahun 1888 oleh Berkeley dan Broome, mempunyai tubuh buah yang tertutup, peridium relatif tebal dan keras, himenium tidak jelas. Tubuh buah S. columnare berbentuk hampir bundar atau tidak beraturan dengan diameter 2—12 cm, berwarna coklat tua dan tangkai. Sporanya berwarna coklat muda hingga coklat tua dan mempunyai diameter 1—12 µm, bentuk spora echinulate atau verrucose, tidak reticulate (Chen 2006). Hifanya berwarna kuning pucat, berdinding tebal, bercabang dan bersepta sederhana (Smits 1994). Chen (2006) menemukan bahwa fungi ini tidak memiliki clamp-connection, namun Wulandari (2002), Tata (2001) dan Santoso (1997) menemukan clamp-connection pada hifa S. columnare yang diteliti.
Pertumbuhan S. dictyosporum relatif lambat. Anyaman hifa yang terbentuk seperti woll dan sulit dilepaskan dari tanah yang menempel, serta berwarna putih kekuningan (Sannon et al. 2009), bersepta dan memiliki clamp-connection (Chen 2006, Wulandari 2002).
Tubuh buah S. sinnamariense dapat ditandai dengan warna kuning yang khas, baik pada gleba hingga hifa yang terbentuk. Bentuk dan ukurannya sangatlah bervariasi, mulai berbentuk sangat bulat hingga berbentuk tidak beraturan. Spora berwarna coklat tua hingga kehitaman dan memiliki diameter 7—12 µm, bentuk spora reticulate sempurna dan hifa memiliki septa dan clamp connection (Chen 2006). Ciri khas lain fungi ini ialah ketika masak akan pecah dengan pola menyerupai bunga yang mekar. Pada umumnya ditemukan bersimbiosis dengan melinjo, dan di berbagai daerah di Indonesia tubuh buahnya yang masih muda (warna glebanya masih putih) dikonsumsi sebagai bahan sayuran (Watling et al. 2002: Wulandari 2002). Menurut Watling (2006) S. sinnmariense adalah species Scleroderma yang paling banyak sinonimnya, yaitu: S. aureum, Mass, 1889, dari New Guinea, S. chyrsasrum Mart, 1954 dari Panama, S. endosatrum Petch, 1919, dari Srilangka, S. flavocrocattum Sacc & de toni, 1887, dari Malaysia, S. lutuem Llyod, 1914, Sewarang, S. pantherinum Matt, 1931 dari Indonesia, S. pisiforme P. Henn, 1859, Camerun.
Tanaman Inang
Shorea pinanga
ekonomi yang rendah. Buahnya digunakan sebagai tengkawang (Mindawati dan Rustaman 2004).
Pohon S. pinanga berukuran sedang hingga besar, tinggi 50 m, diameter 1,25 m, banir kecil dengan tinggi 1,5 m; daun jorong hingga bulat telur menyempit, berkulit tipis, 11-24 cm x 4-9 cm dengan 10-20 pasang tulang daun sekunder, daun penumpu 6 cm; benang sari 15, kepala sari seperti bola memanjang; stolopodium panjang dan pipih (Sumarhani 2007).
Pada umumnya ditemukan pada tanah lempung dan khususnya pada dataran yang luas hingga ketinggian 200 m dpl. Kerapatannya kayu 305-630 kg/m3 pada kadar air 15%. Species tengkawang menghasilkan lemak yang dikenal dengan green butter, yang digunakan sebagai bahan dasar pembuatan coklat, margarine, malam, sabun dan kosmetik. Biji tengkawang dikenal dengan nama borneo tallow, illipe nuts, dan tengkawang kernels. Produksi biji tengkawang dapat mencapai ± 1,25 ton biji kering/ha/panen (Mindawati dan Rustaman 2004).
Pinus merkusii (Pinus)
P. merkusii termasuk famili Pinaceae (Earle 2008a). Species ini merupakan salah satu species pohon industri yang penting di Indonesia. Di Sumatera dikenal dengan beberapa nama daerah, seperti di Aceh disebut damar atau sala, di Gayo disebut uyam, di Tapanuli disebut tusam, di Kerinci sugi, di daerah Alas dikenal dengan nama Sulu, sedangkan di Sumatera Barat disebut susugi (Hendi 2000).
Species kayu pinus dapat dimanfaatkan sebagai bahan baku pembuatan pulp dan kertas, batang dan kotak korek api, dan kayu pertukangan. Sifat kayu tanaman ini ialah (1) kayunya berwarna kuning kecoklatan; (2) gubalnya berwarna putih; (3) berat species antara 0,42—0,59; dan (4) panjang serat rata-rata 4 mm. Vinir dan getah pinus banyak digunakan untuk bahan terpentin (sebagai bahan pelarut atau minyak pengering, semir sepatu dan logam) dan gondorukem (untuk campuran bahan batik; campuran sabun, cat, vernis kertas dan pestisida) (Kalima 2005).
P. merkusii merupakan salah satu species pinus di daerah tropik yang unik, karena penyebarannya secara alami melintasi khatulistiwa ke arah belahan bumi bagian selatan pada ketinggian 150—900 mdpl, curah hujan lebih dari 1900 mm per tahun dan pada tanah sarang serta baik drainasenya. Tanaman ini secara alami tersebar di Aceh, Tapanuli dan Jambi (Gunung Kerinci) (Yafid dan Jafarsidik 2005). Di luar negeri terdapat di Philipina, Laos, Khmer, Thailand dan Birma. P. merkusii dapat tumbuh pada ketinggian mulai dari beberapa meter dari permukaan laut hingga 1800 m dpl dan pada bermacam-macam jenis tanah dengan bahan induk yang berlainan.
Gnetum gnemon (Melinjo)
Tanaman ini berasal dari Malaysia, kemudian menyebar sampai ke India, Indonesia dan kepulauan Fiji. Genus melinjo terdiri dari 33 species yang tersebar di beberapa wilayah, di Amerika Utara ada tujuh species, Afrika Tropik ada dua species, dan sisanya di Asia Tropik termasuk di Indonesia. Di Indonesia terdapat 14 species. Di Indonesia tanaman ini dapat ditemukan di seluruh propinsi (Sentra Informasi IPTEK 2005).
sedikit asam hingga netral (4—6). Walaupun demikian tanaman ini tampaknya tidak tahan pada lahan-lahan yang tergenang. Di Indonesia melinjo terdapat pada daerah pantai yang berhawa panas sampai pegunungan dengan ketinggian 1200 m dpl.
Pohon melinjo memiliki batang yang ramping dan ditutupi oleh cabang yang banyak. Ketinggiannya dapat mencapai 15 m dengan diameter batang 25 cm. Pertumbuhan cabang tidak pernah melampaui batang pokok sehingga selalu tampak lebih jelas. Sistem percabangan yang demikian ini membuat perawakan pohon melinjo tampak seperti kerucut (Anonim 2007).
Daun melinjo berbentuk elips meruncing pada ujungnya dan bertepi rata dengan panjang antara 7,2 —20 cm dan lebarnya antara 2—10 cm. Daun melinjo berjenis tunggal dengan duduk daun berhadapan. Setiap buah melinjo mengandung satu biji melinjo berbentuk elips, ujung meruncing pendek dengan panjang 1—3,5 cm (Cheng dan Cheng 2007). Biji melinjo membutuhkan waktu beberapa bulan hingga satu tahun untuk berkecambah, hal ini disebabkan embrio yang terbentuk ketika biji gugur belum sempurna (dormansi embrio), perkembangan embrio disempurnakan ketika biji disemaikan.
Perkembangan Penelitian Scleroderma Pada Dipterocarpaceae, Pinaceae dan Gnetaceae
Menurut Alexander dan Selosse (2009) penelitian mengenai mikoriza pada hutan tropis masih sangat sedikit, hal ini didasarkan pada hasil penelusuran pada jurnal ilmiah sejak tahun 2000 hingga 2008, dari sekitar 5600 artikel ilmiah yang diterbitkan hanya sekitar 170 artikel yang membahas tentang mikoriza pada hutan tropis (Gambar 4).
Tahun
Gambar 4 Jumlah makalah jurnal dari tahun 2000 – 2008 yang berhubungan dengan mikoriza di hutan tropis (kata kunci pencarian: mycorrhiza & tropical & forest), dibandingkan dengan total literatur tentang mikoriza (kata kunci pencarian: mycorrhiza) (Alexander dan Selosse 2009).
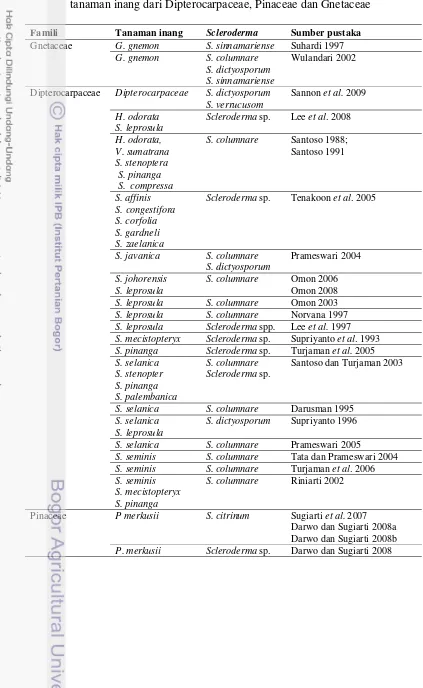
Tabel 2 Daftar penelitian fungi ektomikoriza Scleroderma spp. pada berbagai tanaman inang dari Dipterocarpaceae, Pinaceae dan Gnetaceae
Famili Tanaman inang Scleroderma Sumber pustaka Gnetaceae G. gnemon S. sinnamariense Suhardi 1997
G. gnemon S. columnare
S. dictyosporum S. sinnamariense
Wulandari 2002
Dipterocarpaceae Dipterocarpaceae S. dictyosporum S. verrucusom
S. columnare Santoso 1988;
Santoso 1991
S. javanica S. columnare
S. dictyosporum
Prameswari 2004
S. johorensis S. leprosula
S. columnare Omon 2006
Omon 2008
S. leprosula S. columnare Omon 2003
S. leprosula S. columnare Norvana 1997
S. leprosula Scleroderma spp. Lee et al. 1997 S. mecistopteryx Scleroderma sp. Supriyanto et al. 1993
S. pinanga Scleroderma sp. Turjaman et al. 2005
Santoso dan Turjaman 2003
S. selanica S. columnare Darusman 1995
S. selanica S. leprosula
S. dictyosporum Supriyanto 1996
S. selanica S. columnare Prameswari 2005
S. seminis S. columnare Tata dan Prameswari 2004
S. seminis S. columnare Turjaman et al. 2006
S. seminis S. mecistopteryx S. pinanga
S. columnare Riniarti 2002
Pinaceae P merkusii S. citrinum Sugiarti et al. 2007
Darwo dan Sugiarti 2008a Darwo dan Sugiarti 2008b
Tabel 2 (Lanjutan)
Famili Tanaman inang Scleroderma Sumber pustaka Pinaceae P. patula S. citrinum Mohan et al. 1993
P. pinaster Scleroderma sp. Nieto & Carbone 2009
Pera &Alvarez 1995
P. pinaster S. citrinum Parlade et al. 1996
P. pinea Scleroderma sp. Rincon et al. 1999
P. pinea S. verrucosum Rincon et al. 2001
P. radiate S. citrinum Dunabeitia et al. 2004
P. radiata P. ellioti
Scleroderma spp. Chen et al. 2006
P. taedae Scleroderma sp. Giachini et al. 2004
Penelitian yang terbanyak bertujuan untuk melihat kompatibilitas dan pengaruh inokulasi fungi ektomikoriza pada tanaman inang dengan berbagai macam inokulum yang digunakan, seperti inokulum suspensi spora (Santoso 1988; Santoso 1991 ; Chen et al. 2006; Tata dan Prameswari 2004; Turjaman et al. 2005; Turjaman et al. 2006; Prameswari 2005; Prameswari 2004;Norvana 1997; Sugiarti et al. 2007), Tablet spora (Omon 2006; Omon 2008; Riniarti 2002), miselium (Supriyanto et al. 1993; Darusman 1995; Lee et al. 2008; Wulandari 2002; Rincon et al. 1999, 2001). Hasil penelitian secara umum menunjukkan kesesuaian antara tanaman inang yang diujicobakan dengan fungi ektomikoriza Scleroderma spp. Hasil lainnya adalah kemampuan fungi ektomikoriza untuk meningkatkan pertumbuhan tanaman inang dan juga membantu penyerapan unsur hara bagi tanaman inang.
Beberapa penelitian tentang upaya pengembangan Scleroderma spp. secara in vitro juga telah dilakukan. Media yang digunakan adalah MMN (Modified Melin Norkrans) padat dan cair (Supriyanto dan Irawan 1997; Tata 2003; Ba et al. 1999; Sannon et al. 2009; Diedhiou et al. 2004; Ray et al. 2005; Zeng et al. 2003; Zadworny et al. 2007)
Selain dengan Dipterocarpaceae, Pinaceae dan Gnetaceae, Scleroderma juga ditemukan banyak bersimbiosis dengan tanaman dari famili Fagaceae, seperti pada Eucalypyus globulus dan E. urophylla (Brundrett et al. 2005; Chen et al. 2007), E. tereticornis (Reddy dan Satyanarayana 1998), Quercus garryana (Valentine et al. 2004), Caesalpiniaceae, yaitu pada Afzelia africana (Bâ et al. 1999; Diedhiou et al. 2004), Uapaca bojeri L. (Ramanankierana et al. 2007), Betulaceae yaitu pada Alnus (Moreau et al. 2006), yang berada di hutan subtropis.
pada hutan tanaman ataupun pada kegiatan-kegiatan rehabilitasi hutan yang saat ini semakin gencar dilakukan oleh pemerintah.
Pengetahuan tentang hubungan species-species fungi ektomikoriza dengan tanaman inang akan memberikan informasi penting untuk penelitian-penelitian di bidang silvikultur dan ekologi, serta dalam pengambilan keputusan manajemen ekosistem hutan, informasi-informasi ini dibutuhkan untuk memahami peranan dan fungsi fungi ektomikoriza bagi tanaman inang.
DAFTAR PUSTAKA
Alexander I, Selosse M-A. 2009. Mycorrhizas in tropical forests: a neglected research imperative. New Phytol 182:14–16.
Allen MF, Swenson W, Querejeta JJ, Warburton LME, Treseder KK. 2003. Ecology of mycorrhizae: A conceptual framework for complex interactions among plants and fungi. Annu Rev Phytopathol 41:271–303. Anonim. 2006. Mushroom in Asia. www.edinburgh.ceh.ac.uk/tropica/
vietmushManintro.pdf. [4 April 2006].
Anonim. 2007. Gnetum gnemon. www.wordagroforestrycentre.org/sea/ PRODUCT/FDbases/AF/asp/Spesciesinfo.asp?SPID=1751 [4 November 2007].
Bâ AM, Sanon KB, Duponnois R, Dexheimer J. 1999. Growth response of Afzeli africana Sm. seedlings to ectomycorrhizal inoculation in a nutrient-deficient soil. Mycorrhiza 9:91–95.
Brundrett M, Bougher N, Dell B, Grove T, Malajczuk N. 1996. Working with Mycorrhiza in Forestry and Agriculture. Canberra: Australian Centre for International Agricultural Research.
Brundrett M, Malajczuk N, Mingqin G, Daping Xu, Snelling S, Dell B. 2005. Nursery inoculation of Eucalyptus seedlings in Western Australia and Southern China using spore and mycelia inoculum of diverse ectomycorrhizal fungi from different climatic regions. For Ecol Man 209:163–205.
Chen YL, Kang LH, Malajczuk N, Dell B. 2006. Selecting ectomycorrhizal fungi for inoculating plantations in south China: effect of Scleroderma on colonization and growth of exotic Eucalyptus globulus, E. urophylla, Pinus elliottii, and P. radiata. Mycorrhiza 16:251–259.
Chen YL, Liu S, Dell B. 2007. Mycorrhizal status of Eucalyptus plantations in south China and implication for management. Mycorrhiza 17:527–535. Cheng CY, Cheng WC. 2007. Gnetum, Flora of China. Acta Phytotax Sin
13:88–90.
Darusman LK. 1995. Telaah Biokimia Proses Asosiasi Shorea selanica dan Scleroderma columnare [disertasi]. Bogor: Program Pascasarjana, Institut Pertanian Bogor.
Darwo, Sugiarti. 2008a. Beberapa jenis cendawan ektomikoriza di kawasan hutan Sipirok, Tongkoh, dan Aek Nauli, Sumatera Utara. J Pen Hut & KA 5:157–173.
Darwo, Sugiarti. 2008b. Pengaruh dosis serbuk spora cendawan Scleroderma citrinum Persoon dan komposisi media terhadap pertumbuhan tusam di persemaian. J Pen Hut & KA 5:461-472.
Dickinson TA, Hutchison LJ. 1997. Numerical taxonomic methods, cultural characters, and the systematics of ectomycorrhizal agarics, boletes and gasteromycetes. Mycol Res 101:477–492.
Diedhiou AG, Verpillot F, Gueye O, Dreyfus B, Duponnois R, Bâ AM. 2004. Do concentrations of glucose and fungal inoculum influence the competitiveness of two early-stage ectomycorrhizal fungi in Afzelia africana seedlings? For Ecol And Man 203:187–194.
Diouf D, Diop TA, Ndoye I. 2003. Actinorhizal, mycorrhizal, and rhizobial symbioses: how much do we know? Af J Biotech 2:1–7.
Duñabeitia MK, Hormilla S, Garcia-Plazaola JI, Txarterina K, Arteche U, Becerril JM. 2004. Differential responses of three fungal species to environmental factors and their role in the mycorrhization of Pinus radiata D. Don. Mycorrhiza 14:11–18.
Earle CJ. 2008a. Pinus, The Gymnosperm Databases. www.conifers.Org/pi/pin/ index.html. [12 November 2008]
Earle CJ. 2008b. Pinus merkusii Junghuhn & de Vriese 1845. www. Conifers. Org/pi/pin/merkusii.htm [12 November 2008].
Giomaro G, Sisti D, Zambonelli A, Amicucci A, Cecchini M, Comandini O, Stocchi V. 2002. Comparative study and molecular characterization of ectomycorrhizas in Tilia americana and Quercus pubescens with Tuber brumale. FEMS Microbiol Letters 216:9–14.
Hall IA, Yun W, Amicucci A. 2003. Cultivation of edible ectomycorrhizal mushrooms. Trends in Biotechnol 21:433–438.
Hendi. 2000. Kajian Tehnik Konservasi Pinus merkusii Strain Kerinci. www.dephut.go.id/files/Hendi.pdf [12 November 2008]
Kalima T. 2005. Mengenal panorama alam Pinus merkusii Jungh de Vries di Sumatera Utara. Warta Pusat Litbang dan Konservasi Alam 11:6–7. Kasuya T. Guzmán G, Ramirez-Guillèn F, Kato T. 2002. Scleroderma laeve
(Gasteromycetes, Sclerodermatales), new to Japan. Mycoscience 43:475– 476.
Koide RT, Shumway DL, Xu B, Sharda JN. 2007. On temporal partitioning of a community of ectomycorrhizal fungi. New Phytol 174:420–429.
Lee LS, Alexander IJ, Watling R. 1997. Ectomycorrhizas and putative ectomycorrhizal fungi of Shorea leprosula Miq. (Dipterocarpaceae). Mycorrhiza 7:63–81.
Lee SS, Patahayah M, Chong WS, Lapeyrie F. 2008. Successful ectomycorrhizal inoculation of two dipterocarp species with a locally isolated fungus in Peninsular Malaysia. J of Trop For Sci 20:237–247.
Luo Z-B, Janz D, Jiang X, Göbel C, Wildhagen H, Tan Y, Rennenberg H, Feussner I, Polle A. 2009. Upgrading root physiology for stress tolerance by ectomycorrhizas: Insights from metabolite and transcriptional profiling into reprogramming for stress anticipation. Plant Physiol 151:1902–1917. Martín-Pinto P, Pajares J, Díez J. 2006. In vitro effects of four ectomycorrhizal
fungi, Boletus edulis, Rhizopogon roseolus, Laccaria laccata and Lactarius deliciosus on Fusarium damping off in Pinus nigra seedlings. New Forests 32:323–334.
Mindawati N, Rustaman B. 2004. Mengenal Tengkawang. Warta Pusat Litbang Hutan dan Konservasi Alam 1:9–10.
Mohan V, Natrajan K, Ingleby K. 1993. Anatomical studies on ectomycorrhizas. III The ectomycorrhizas produced by Rhizopogon luteolus and Scleroderma citrinum on Pinus patula. Mycorrhiza 3:51–56.
Muchovej RM. 2002. Importance of Mycorrhizae for Agricultural Crops. http:edis.ifas.ufl.edu/BODY_AGI16 [10 Maret 2006].
Niemi K, Häggman H, Sarjala T. 2002a. Effects of exogenous diamines on the interaction between ectomycorrhizal fungi and adventitious root formation in Scots pine in vitro. Tree Physiol 22:373–381.
Niemi K, Vuorinen T, Ernstsen A, Häggman H. 2002b. Ectomycorrhizal fungi and exogenous auxins influence root and mycorrhiza formation of Scots pine hypocotyl cuttings in vitro. Tree Physiol 22:1231–1239.
Nieto MP, Carbone SS. 2009. Characterization of juvenile maritime pine (Pinus pinaster Ait.) ectomycorrhizal fungal community using morphotyping, direct sequencing and fruitbodies sampling. Mycorrhiza 19:91–98.
Norvana UI. 1997. Peranan Sleroderma columnare dan media tumbuh bahan organik terhadap pertumbuhan stek Shorea leprosula Miq [tesis]. Bogor: Program Pascasarjana, Institut Pertanian Bogor.
Omon RM. 2003. Pengaruh tablet mikoriza terhadap persen akar bermikoriza stek Shorea leprosula Miq. di rumah kaca Wanariset Semboja, Kalimantan Timur. Bul Pen Hutan 16: 23—30.
Omon RM. 2006. Pengaruh suhu dan lama penyimpanan tablet mikoriza terhadap pertumbuhan stek Shorea johorensis Foxw. Di rumah kaca. J Pen Hut & KA 3:83–93.
Omon RM. 2008. Pengaruh dosis tablet mikoriza terhadap pertumbuhan dua jenis meranti merah asal benih dan stek di HPH PT. ITCIKU, Balikpapan. Kalimantan Timur. Info Hutan 5:329–35.
Onguene NA, Kuyper TW. 2002. Importance of ectomycorrhiza network for seedling survival and ectomycorrhiza formation in rain forests of South Cameroon. Mycorrhiza 12:13–17.
Parladé J, Per J, Alvarez IF. 1996. Inoculation of containerized Pseudotsuga menziesii and Pinus pinaster seedlings with spores of five species of ectomycorrhizal fungi. Mycorhiza 6:237–245.
Peay KG, Kennedy PG, Davies SJ, Tan S, Bruns TD. 2009. Potential link between plant and fungal distributions in a dipterocarp rainforest: Community and phylogenetic structure of tropical ectomycorrhizal fungi across a plant and soil ecotone. New Phytol doi: 10.1111/j.1469-8137.2009.03075.x.
Pera J, Alvarez IF. 1995. Ectomycorrhizal fungi of Pinus pinaster. Mycorrhiza 5:193–200.
Peterson RL, Massicotte HB, Melville LH. 2004. Mycorrhiza: Anatomy and Cell Biology. Ottawa: NRC Research Press.
Prameswari D, Tata MHL. 2004. Effect of planting media on the growth of Shorea pinanga Scheff seedlings. J For Res 1:25–30.
Prameswari D. 2004. Pengaruh inokulasi cendawan ektomikoriza dan media tumbuh terhadap pertumbuhan Shorea javanica K & V. [tesis]. Bogor: Program Pascasarjana, Institut Pertanian Bogor.
Prameswari D. 2005. Aplikasi beberapa cendawan ektomikoriza untuk meningkatkan pertumbuhan semai Shorea selanica. J Hut Trop 1:13–18. Querejeta JI, Egerton-Warburton LM, Allen MF. 2003. Direct nocturnal water
transfer from oaks to their mycorrhizal symbionts during severe soil drying. Oecologia 134:55–64.
Ramanankierana N, Ducousso M, Rakotoarimanga N, Prin Y, Thioulouse J, Randrianjohany E, Ramaroson L, Kisa M, Galiana A, Duponnois R. 2007. Arbuscular mycorrhizas and ectomycorrhizas of Uapaca bojeri L. (Euphorbiaceae): sporophore diversity, patterns of root colonization, and effects on seedling growth and soil microbial catabolic diversity. Mycorrhiza 17:195–208.
Ray P, Tiwari R, Reddy UG, Adholeya A. 2005. Detecting the heavy metal tolerance level in ectomycorrhizal fungi in vitro. World J Microbiol Biotech 21:309–315.
Reddy MS, Satyanarayana T. 1998. Inoculation of micropropagated plantlets of Eucalyptus tereticornis with ectomycorrhizal fungi. New Forests 16:273– 279.
Rinaldi AC, Comandini O, Kuyper TW. 2008. Ectomycorhizal fungal diversity: Separating wheat from the chaff. Fungal Diversity 33:1–45.
Rincón A, Àlvarez IF, Pera J. 1999. Ectomycorrhizal fungi of Pinus pinea L. in northeastern Spain. Mycorrhiza 8:271–276.
Rincón A, Àlvarez IF, Pera J. 2001. Inoculation of containerized Pinus pinea L. seedlings with seven ectomycorrhizal fungi. Mycorrhiza 11:265–271. Riniarti M, Setiadi Y, Sopandie D. 2005. Aplikasi asam organik dan inokulasi
ektomikoriza untuk meningkatkan pertumbuhan semai Shorea pinanga. J Hut Trop 1:25–29.
Sanon KB, AM Bâ, Delaruelle C, Duponnois R, Martin F. 2009. Morphological and molecular analysis in Scleroderma species associated with some Caesalpinioid legumes, Dipterocarpaceae, and Phylanthaceae trees in southern Burkina Faso. Mycorrhiza 19:571–584.
Santoso E, Turjaman M, Irianto RSB. 2007. Aplikasi mikoriza untuk meningkatkan kegiatan rehabilitasi hutan dan lahan terdegradasi. Di dalam: Siran AS, Bismark M, Samsoedin I, Suhaendi H, Pratiwi, Haryono, Mardiah, editor. Konservasi dan Rehabilitasi Sumberdaya Alam. Prosiding Ekspose Hasil-Hasil Penelitian. Padang, 20 Sep 2006. Bogor: Pusat Penelitian dan Pengembangan Kehutanan. Hlm 71–80.
Santoso E. 1988. Pengaruh mikoriza terhadap diameter batang dan bobot kering anakan Dipterocarpaceae. Bul Pen Hutan 504:11–21.
Santoso E. 1991. Pengaruh beberapa fungi mikoriza terhadap penyerapan unsur hara pada 5 jenis Dipterocarpaceae. Bul Pen Hutan 532:11–18.
Santoso E. 1997. Hubungan perkembangan ektomikoriza dengan populasi jasad renik dalam rhizosfer dan pengaruhnya terhadap pertumbuhan Eucalytus pellita dan Eucalyptus urophylla [disertasi]. Bogor: Program Pascasarjana, Institut Pertanian Bogor.
Santoso E. Turjaman M. 2003. Tipe-tipe Struktur Ektomikoriza Pada Shorea selanica, S. stenoptera, S. pinanga, dan S. palembanica (Dipetocarpaceae) di Hutan Penelitian Haurbentes Jawa Barat. Bul Pen Hutan 636:33–37. Satomura T, Hasimoto Y, Kinoshita A, Horikoshi T. 2006. Methods to study the
role of ectomycorrhizal fungi in forest carbon cycling: Introduction to the direct methods to qualify the fungal content in ectomycorrhizal fin roots. Root Research 15:119–124.
Sentra Informasi IPTEK.2005. Melinjo. www.iptek.net.id/ind/teknologi_pangan/ index.php?id=270 - 20k [20 Maret 2006].
Smith SE, Read DJ. 2008. Mycorrhizal Symbiosis. Third Edition. London: Academic Press.
Smits WTM. 1994. Dipterocarpaceae: Mycorrhizae and Regeneration. Wageningen: The Tropenbos Foundation.
Sugiarti, Darwo, Panjaitan DJ. 2007. Efektivitas bentuk inokulum cendawan Scleroderma citrinum Persoon dalam meningkatkan pertumbuhan semai Pinus merkusii Jungh. Et de Vriese. J Pen Hut & KA 4:63–74.
Sumarhani. 2007. Pemanfaatan dan konservasi jenis meranti merah penghasil biji tengkawang (Shorea stenoptera Burk dan Shorea pinanga Scheff). Info Hutan 4:177–185.
Supriyanto, Irawan US. 1997. Inoculation techniques of ectomycorrhizal. Prosiding: Seminar on Mycorrhizae. Balikpapan 28 Februari 1992. Hlm 36–40.
Supriyanto, Setiawan I, Omon M. 1993. Effect of Scleroderma sp. on the growth of Shorea mecistopteryx Ridl. seedlings. Prosiding: Bio-Refor Proceeding of Yogyakarta Workshop. Yogyakarta 20–23 September 1993. Hlm 186– 188.
Tata HL, van Noordwijk M, Summerbell R, Werger MJA. 2010. Limited response to nursery-stage mycorrhiza inoculation of Shorea seedlings planted in rubber agroforest in Jambi, Indonesia. New Forests 39:51–74. Tata MHL, Prameswari D. 2004. Pengaruh inokulasi tablet spora ektomikoriza
Scleroderma columnare terhadap pertumbuhan Shorea seminis dan efektivitasnya pada berbagai dosis arang. Di dalam: Simarmata T, Arief DH, Sumarni Y, Hindersah R, Azirin A, Kalay AM, editor. Teknologi Produksi dan Pemanfaatan Inokulan Endo-Ektomikoriza untuk Pertanian, Perkebunan, dan Kehutanan. Prosiding Seminar Mikoriza; Bandung, 16 Sep 2003. Bandung: Asosiasi Mikoriza Indonesia – Jawa Barat bekerja sama dengan Universitas Padjadjaran Bandung. Hlm 163-170.
Tata MHL. 2001. Pengaruh kebakaran hutan terhadap daya tahan hidup fungi ektomikoriza dipterocarpaceae [Thesis]. Bogor: Program Pascasarjana, Institut Pertanian Bogor.
Tata MHL. 2003. Nutrient acquisition of ectomycorrhizae fungus Scleroderma columnare. Presented at the Open Science Meeting Indonesia and Netherlands: Back to the Future. Jakarta. Indonesia, 1–2 Sep 2003.
Tennakoon MMD, Gunatilleke IAUN, Hafeel KM, Seneviratne G, Gunatilleke CVS, Ashton PMS. 2005. Ectomycorrhizal colonization and seedling growth of Shorea (Dipterocarpaceae) species in simulated shade environments of a Sri Lankan rain forest. For Ecol and Man 208:399–405. Turjaman M, Irianto RSB, Santoso E. 2002. Teknik inokulasi dan produksi
massal cendawan ektomikoriza. Info Hutan 152:1–16.
Turjaman M, Tamai Y, Segah H, Limin SH, Osaki M, Tawaraya K. 2006. Increase in early growth and nutrient uptake of Shorea seminis seedlings inoculated with two ectomycorrhizal fungi. J of Trop For Sci 18:243–249. Valentine LL, Fiedler TL, Hart AN, Petersen CA, Berninghausen HK, Southworth
D. 2004. Diversity of ectomycorrhizas associated with Quercus garryana in southern Oregon. Can J Bot 82:123–135.
Watling R, Lee SS, Turnbull E. 2002. The Occurrence and Distribution of Putative Ectomycorrhizal Basidiomycetes in a Regenerating South East Asian Rain Forest. Di Dalam: Watling R, Frankland JC, Ainsworth AM, Isaac S, Robinson CH, editor. Tropical Mycology Volume 1, Macromycetes. New York: CABI Publishing. Hlm 116—203.
Watling R. 2006. The sclerodermatoid fungi. Mycoscience 47:18–24.
Whipps JM. 2004. Prospects and limitations for mycorrhizas in biocontrol of root pathogens. Can J Bot 82:1198–1227.
Wulandari AS. 2002. Beberapa gatra biologi ektomikoriza Scleroderma pada melinjo [disertasi]. Bogor: Program Pascasarjana, Institut Pertanian Bogor. Yafid B, Jafarsidik YS. 2005. Permudaan Pinus merkusii Jungh et de Vriese
galur kerinci, potensi dan komposisi tegakan di kawasan hutan Bukit Tapan, Taman Nasional Kerinci Seblat, Jambi. Info Hutan 2:145–152. Yamada A, Kobayashi H, Ogura T, Fukada M. 2007. Sustainable fruit body
formation of edible mycorrhizal Tricholoma spesies for 3 years in open pot culture with pine seedling hosts. Mycoscience 48:104–108.
Yamada A, Ogura T, Ohmasa M. 2001. Cultivation of mushrooms of ectomycorrhizal fungi associated with Pinus densyflora by in vitro mycorrhizal synthesis. Mycorrhiza 11:67–81.
Zadworny M, Smolinski DJ, Idzikowska K, Werner A. 2007. Ultrastructural and cytochemical aspects of the interaction between the ectomycorrhizal fungus Laccaria laccata and the saprotrophic fungi, Trichoderma harzianum and T. virens. Biocontrols Sci and Technol 17:921–932.
Zeller SM. 1948. Note on certain Gasteromycetes including two new orders. Mycologia 40:639–668.
Scleroderma spp SECARA IN VITRO
(Compatibility Test of Three ectomycorrhizal Fungi Scleroderma spp. Under In Vitro Condition)
ABSTRAK
Interaksi antar fungi ektomikoriza dalam mengkolonisasi akar merupakan bagian penting yang harus dipelajari untuk dapat memahami pola pembentukan mikoriza pada akar. Sebuah percobaan secara in vitro di laboratorium telah dilakukan untuk mengidentifikasi pola pertumbuhan miselium fungi ektomikoriza. Tiga spesies Scleroderma spp. dikulturkan pada media MMN (Modified Melin Norkrans). Masing-masing fungi dikulturkan secara tunggal, berpasangan, dan tripel. Percobaan menggunakan rancangan acak lengkap dengan 7 perlakuan dan 8 ulangan, pengamatan dilakukan selama lima minggu. Hasilnya menunjukkan bahwa diameter pertumbuhan koloni S. sinnamariense dua kali lebih cepat daripada S. columnare dan S. dictyosporum. Ketiga fungi ektomikoriza dapat berkembang secara bersama-sama, tidak terdapat antagonisme di antara ketiganya.
Kata kunci: fungi ektomikoriza, kompatibilitas, miselium, Scleroderma ABSTRACT
Understanding on interaction among ectomycorrhizal fungus would led the knowledge of roots colonization. We conducted an in vitro experiment in laboratory condition to identify the compatibility of mycelia among the fungus. Three spesies Scleroderma spp. were cultured on solid Modified Melin Norkrans (MMN) medium. Each fungus was cultured as single, pairs and triple. A completed randomized design were used with 7 treatments and 8 replicates Petri dishes. We found that the radial growth of S. sinnamariense mycelia was two times faster than S. columnare and S. dictyosporum. There was no antagonism pattern among mycelium.
Key words: compatibility, ectomycorrhizal fungi, mycelia, Scleroderma
PENDAHULUAN
Kompetisi didefinisikan sebagai pengaruh negatif suatu spesies terhadap spesies lainnya yang berhubungan dengan alokasi sumberdaya, atau pembatasan akses terhadap sumberdaya yang ada (Keddy 2007). Adanya kompetisi akan menyebabkan terjadinya pembagian niche (Kennedy et al. 2007a), yang akan dipengaruhi oleh berbagai faktor lingkungan seperti kelembaban tanah (Kennedy et al. 2007b), kedalaman tanah (Dickie et al. 2004), status nutrisi (Püttsepp et al. 2004), dan sebagainya.
Hasil penelitian Kennedy et al. (2009) menunjukkan bahwa waktu pembentukan kolonisasi merupakan salah satu faktor yang menyebabkan kompetisi antar fungi ektomikoriza. Hal ini berhubungan dengan kecepatan perkecambahan spora fungi ektomikoriza (Smith dan Read 2008), semakin cepat spora berkecambah maka kemungkinan untuk mengkolonisasi akar akan semakin besar. Fungi ektomikoriza yang lebih dulu menginfeksi akar umumnya akan menjadi spesies fungi ektomikoriza yang dominan (Kennedy dan Bruns 2005).
Pada kondisi alam, kompetisi yang terjadi berada pada tingkat hifa dan miselium (Daza et al. 2006). Miselium merupakan bagian yang paling dinamis dan berfungsi luas dalam membentuk simbiosis. Kecepatan pertumbuhan dan perkembangan miselium pada akar tanaman akan menentukan besarnya persen kolonisasi yang dapat dibentuk oleh fungi (Leake et al. 2004), karena 80% dari biomassa fungi ektomikoriza adalah ekstraradikal miselium (Wallander et al. 2001). Upaya mempelajari perilaku miselium dapat dilakukan secara in vitro (Zeng et al. 2003), kompetisi dapat dijabarkan dengan melihat adanya sifat antagonis pada miselium tiap spesies fungi ektomikoriza yang dibiakkan, untuk memahami perilaku miselium di alam.
METODE PENELITIAN
Waktu dan Tempat
Penelitian ini dilaksanakan sejak Juli 2008 hingga November 2009, di Laboratorium Silvikultur Departemen Silvikultur Fakultas Kehutanan IPB.
Bahan dan Metode Penyiapan media dan miselium fungi
Media yang digunakan untuk menumbuhkan fungi ektomikoriza secara in vitro ialah Modified Melin Norkrans (MMN) (Lampiran 1). Miselia yang dikembangkan berasal dari tubuh buah Scleroderma columnare, S. dictyosporum dan S. sinnamariense. Tubuh buah yang digunakan ialah tubuh buah yang masih muda (ditandai dengan tubuh buah yang masih keras dan gleba yang masih kompak). Tubuh buah dikumpulkan dari beberapa lokasi di lingkungan Fakultas Kehutanan IPB, pada bulan Mei 2008. Bagian tubuh buah yang digunakan ialah bagian gleba, dicungkil dengan menggunakan jarum oase dan diletakkan di tengah-tengah media MMN. Miselium yang dikembangkan pada media MMN selama empat minggu digunakan sebagai bahan penelitian (Gambar 5). Semua proses pengembangan miselia dilakukan dalam keadaan aseptik.
Uji in vitro pada media agar
Gambar 5 Bahan yang digunakan dalam penyiapan miselium fungi: bagian atas (a) tubuh buah dan bagian bawah (b) isolat hasil pembiakan tubuh buah; (1) S. sinnamariense, (2) S. columnare, dan (3) S. dictyosporum.
Gambar 6 Penempatan inokulum miselium fungi ektomikoriza pada cawan Petri: (a) perlakuan tripel (CDS); (b) perlakuan ganda (CD, CS, dan DS); (c) perlakuan tunggal (C, D, dan S).
Peubah yang diamati
Peubah yang diamati ialah (1) awal pertumbuhan miselium (2) diameter pertumbuhan koloni (Gambar 7), dan (3) ruang tumbuh. Rancangan yang
c
a b
1 mm
1 cm
1 cm 1 cm
1 a
1 b
2 a 3 a
2 b 3 b
1 mm 1 mm
digunakan adalah Rancangan Acak Lengkap, dengan tujuh perlakuan dan delapan ulangan sehingga jumlah satuan percobaannya adalah 56.
Tabel 3 Rincian perlakuan yang dilakukan secara in vitro
No Perlakuan Ulangan
Fungi Ektomikoriza
1 S. columnare + S. dictyosporum +S. sinnamariense 8 2 S. columnare + S. dictyosporum 8 3 S. columnare + S. sinnamariense 8 4 S. dictyosporum + S. sinnamariense 8
5 S. columnare 8
6 S. dictyosporum 8
7 S. sinnamariense 8
Gambar 7 Perhitungan diameter pertumbuhan koloni dalam cawan Petri dengan menggunakan delapan arah mata angin.
HASIL DAN PEMBAHASAN Hasil
Awal pertumbuhan miselium Scleroderma spp. yang dibiakkan dengan media MMN menggambarkan kecepatan pertumbuhan masing-masing spesies. S. sinnamariense merupakan spesies yang paling cepat tumbuh. Pada hari kelima miselium S. sinnamariense mulai tumbuh. Sementara S. columnare dan S. dictyosporum rata-rata baru mulai tumbuh di hari kedelapan setelah penanaman dalam cawan Petri.
pesat diameter pertumbuhan koloninya baik ketika dibiakkan sendiri maupun saat berpasangan. Kecepatan pertumbuhan S. sinnamariense dapat mencapai dua kali lipat dari diameter pertumbuhan koloni S. columnare dan S. dictyosporum. Sedangkan diameter pertumbuhan koloni S. dictyosporum sangat lambat, pertumbuhan radial baru dapat diukur pada minggu ketiga setelah penanaman.
Perbedaan kecepatan diameter pertumbuhan koloni pada ketiga fungi Scleroderma spp. ini tidak menyebabkan timbulnya kompetisi antar fungi tersebut. Hal ini ditandai dengan tidak adanya barrier saat miselium ketiganya tumbuh bersama dalam satu cawan Petri (Gambar 9), yang menunjukkan tidak adanya sifat antagonisme pada ketiga spesies Scleroderma. Hasil pengamatan secara mikroskopis menunjukkan hifa yang tumbuh pada bagian yang bertumpukan antara dua dan tiga fungi tidak ada lisis dan kerusakan pada hifa-hifa fungi yang ada.
D : Perlakuan tunggal S. dictyosporum d : MiseliumS. dictyosporum
S : Perlakuan tunggal S. sinnamariense s : MiseliumS. sinnamariense
CD : Perlakuan ganda S. columnare + S. dictyosporum
CS : Perlakuan ganda S. columnare + S. sinnamariense
DS : Perlakuan ganda S. dictyosporum + S. sinnamariense
CDS : Perlakuan triple S. columnare + S. dictyosporum + S. sinnamariense
d : S. dictyosporum se
Gambar 9 Diameter p r i pada minggu kelima pengamatan.
Miselium dari fungi ektomikor organ penting yang berperan dalam
menunjukkan bahwa S. sinnamariense merupakan fungi yang
atnya perkembangan miselium S. dictyosporum pada media MMN juga
Keterangan: c : S. columnare
1 cm 1 cm
c s
d c
d
s
s : S. sinnamarien
e tumbuhan kolon
Pembahasan iza merupakan
penyerapan unsur hara pada tanaman inang (Nara 2006). Miselia fungi ektomikoriza juga merupakan sumber jembatan hifa bagi tanaman di sekitarnya sehingga membentuk suatu jaringan yang menghubungkan berbagai tanaman (Obase et al. 2009).
Hasil penelitian
memiliki kecepatan awal perkembangan dan diameter pertumbuhan koloni yang paling pesat dibandingkan dua spesies Scleroderma lainnya. Kecepatan S. columnare berada di level menengah sementara S. dictyosporum tumbuh dan berkembang dengan sangat lambat. Dengan demikian, dapat diduga di alam S. sinnamariense akan memiliki kemampuan untuk lebih dulu berkembang pada akar tanaman.
Lamb
, pertu
mene
inokulum. Zeng et al. (2003) menyatakan sebagian besar fungi ektomikoriza sangat lambat pertumbuhannya pada media kultur dibandingkan spesies fungi non ektomikoriza, namun menurut Sanon et al. (2009) dan Chen (2006), miselium Scleroderma merupakan spesies yang mudah tumbuh dan berkembang baik di media kultur maupun di alam, sehingga mudah digunakan untuk aplikasi pada tanaman-tanaman yang memiliki ketergantungan tinggi terhadap ektomikoriza.