KELARUTAN MINERAL KALSIUM (Ca) DAN FOSFOR (P)
BEBERAPA JENIS LEGUM POHON
SECARA IN VITRO
SKRIPSI SUHARLINA
PROGRAM STUDI NUTRISI DAN MAKANAN TERNAK FAKULTAS PETERNAKAN
RINGKASAN
Suharlina. D24101026. 2006. Kelarutan Mineral Kalsium (Ca) dan Fosfor Beberapa Jenis Legum Pohon Secara in Vitro. Skripsi. Program Studi Nutrisi dan Makanan Ternak, Fakultas Peternakan, Institut Pertanian Bogor.
Pembimbing Utama : Dr. Ir. Idat Galih Permana, MSc.Agr. Pembimbing Anggota : Dr. Ir. Luki Abdullah, MSc.Agr
Rumput sebagai pakan ternak umumnya mengandung serat yang tinggi dan protein yang rendah. Ketersediaan rumput juga bergantung pada musim. Pada musim hujan hijauan tersedia cukup melimpah dan menurun jika musim kemarau. Sehingga perlu dicari alternatif hijauan yang ketersediaannya cukup sepanjang tahun seperti legum pohon. Legum pohon merupakan tumbuhan yang dapat tumbuh pada lahan kritis dan tahan musim kemarau selama beberapa bulan. Legum pohon merupakan hijauan sumber protein dan memiliki kandungan mineral yang cukup tinggi.
Penelitian ini bertujuan untuk mengetahui kelarutan mineral makro (Ca dan P) beberapa legum pohon dan nilai fermentabilitasnya didalam rumen. Peubah uang diukur meliputi laju degradasi bahan kering, kelarutaan mineral (Ca dan P), produksi VFA dan NH3 dengan menggunakan teknik in vitro. Materi penelitian berupa lima jenis legum pohon yaitu Angsana (Pterocarpus indicus), Turi (Sesbania grandiflora), Gamal (Gliricidia sepium), Lamtoro (Leucaena leucocephala) dan Kaliandra (Caliandra calothyrsus) yang dinkubasi selama 0, 3, 6, 9, 12, 24, 48 dan 72 jam pada cairan rumen.
Penelitian ini menggunakan Rancangan Acak Kelompok (RAK) yang terdiri dari 5 perlakuan dan 3 kelompok yang didasarkan pada penggunaan cairan rumen yang berbeda. Data yang diperoleh dianalisis dengan sidik ragam (ANOVA). Untuk mengetahui hubungan laju degradasi pakan dengan produksi VFA dan NH3, dilakukan uji korelasi.
Hasil penelitian menunjukkan bahwa kelima jenis legum memiliki produksi VFA yang tidak berbeda nyata (P>0,05). Laju degradasi, kelarutan mineral Ca dan P serta Produksi NH3 masing – masing legum menunjukkan perbedaan yang sangat nyata (P<0,01). Produksi NH3 meningkat jika tingkat degradasi pakan meningkat, tetapi produksi VFA menurun untuk daun turi. Kesimpulan dari penelitian ini adalah Nilai degradasi diurutkan dari tertinggi sampai terendah selama inkubasi 6 hingga 48 jam adalah turi > angsana dan gamal > lamtoro > kaliandra, sedangkan nilai potensi degradasi diurutkan dari yang tertinggi hingga terendah adalah turi dan gamal > angsana dan lamtoro > kaliandra. Kelarutan mineral P setelah inkubasi 24 jam diurutkan dari yang paling tinggi sampai terendah berturut-turut adalah dauun gamal > turi > angsana dan lamtoro > kaliandra. Terdapat hubungan yang erat antara fermentabilitas pakan dengan kelarutan mineral.
ABSTRACT
Solubility of Calcium and Phosphor of Several Tree Legumes Using In Vitro Technique
Suharlina., I.G. Permana., L. Abdullah
An in vitro experiment was carried out to examine the solubility of Ca and P of selected tree legumes in rumen liquor. The legumes were Pterocarpus indicus,
Sesbania grandiflora, Gliricidia sepium, Leucaena leucocephala and Caliandra calothyrsus. The legumes leaves were dried, ground and incubated in rumen liquor at 0, 3, 6, 9, 12, 24, 48, and 72 h. The observed variables were degradation rate of legumes leaves, solubility of Ca and P, concentration of ammonia and total VFA concentration. The data were analyzed using analysis of variance. The results showed that the VFA production of tree legumes was no different statistically (P>0.05). However, the degradation rate, solubility of Ca and P and the ammonia concentration of Sesbania grandiflora were significantly higher than those of other legumes (P<0.05). There was significant relationship between the degradation rate, solubility of Ca and P and ammonia concentration.
KELARUTAN MINERAL KALSIUM (Ca) DAN FOSFOR (P)
BEBERAPA JENIS LEGUM POHON
SECARA IN VITRO
SUHARLINA
D24101026
Srikpsi ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan pada
Fakultas Peternakan Institut Pertanian Bogor
PROGRAM STUDI NUTRISI DAN MAKANAN TERNAK FAKULTAS PETERNAKAN
KELARUTAN MINERAL KALSIUM (Ca) DAN FOSFOR (P)
BEBERAPA JENIS LEGUM POHON
SECARA IN VITRO
Oleh
SUHARLINA
D24101026
Skripsi ini telah disetujui dan disidangkan dihadapan Komisi Ujian Lisan pada tanggal 14 Februari 2006
Pembimbing Utama
Dr. Ir. Idat Galih Permana, MSc.Agr. NIP. 131 956 694
Pembimbing Anggota
Dr. Ir. Luki Abdullah, MSc.Agr. NIP. 131 672 157
Mengetahui,
Dekan Fakultas Peternakan Institut Pertanian Bogor
RIWAYAT HIDUP
Penulis dilahirkan pada tanggal 14 Juli 1982 di Pamekasan Jawa Timur. Penulis adalah anak pertama dari empat bersaudara dari pasangan Bapak Achmad Padli dan Ibu Marsuna.
Penulis mulai menempuh pendidikan di Sekolah Dasar Negeri Pademawu Timur V Kecamatan Pademawu, Kabupaten Pamekasan pada tahun 1989 dan lulus pada tahun 1995. Pendidikan lanjutan tingkat pertama diselesaikan pada tahun 1998 di SLTP Negeri 1 Pademawu dan pendidikan menengah umum diselesaikan pada tahun 2001 di SMU Negeri 3 Pamekasan.
Pada tahun 2001, penulis diterima sebagai mahasiswa Tingkat Persiapan Bersama di Institut Pertanian Bogor (TPB–IPB) melalui Undangan Seleksi Masuk IPB (USMI). Penulis terdaftar sebagai mahasiswa Departemen Ilmu Nutrisi dan Makanan Ternak, Program Studi Nutrisi dan Makanan Ternak, Fakultas Peternakan, Institut Pertanian Bogor.
KATA PENGANTAR
Syukur Alhamdulillah, penulis panjatkan kehadirat Allah SWT. yang Maha Kuasa atas segala karuniaNya sehingga penulis bisa menyelesaikan karya ilmiah dengan judul Kelarutan Mineral Kalsium (Ca) dan Fosfor (P) Beberapa Jenis Legum Pohon Secara in Vitro.
Leguminosa pohon merupakan tumbuhan tropis yang dapat hidup pada lahan yang kurang subur. Sebagai hijauan pakan, leguminosa pohon dikenal sebagai hijauan sumber protein dengan kecernaan yang lebih tinggi dari rumput, kandungan mineral (khususnya kalsium dan fosfor) dan vitamin yang tinggi. Selain itu leguminosa pohon mampu mensuplai protein fermentable dan by pass (dengan adanya tannin). Keuntungan lain dari leguminosa pohon adalah dapat meningkatkan kesuburan tanah, melindungi tanah dari erosi dan merupakan penghasil kayu yang bermutu. Penambahan legum dalam ransum diharapkan dapat mensuplai kebutuhan ternak, sehingga penggunaan ransum lebih efisien.
Penulis berharap semoga skripsi ini dapat memberikan kontribusi bagi pengetahuan dalam bidang ilmu nutrisi dan makanan ternak.
Bogor, Februari 2006
Penulis
DAFTAR ISI
Ketersediaan Mineral Makro dalam Hijauan ... 10
Mineral Ca dan P ... 10
Laju Degradasi Bahan Kering Legum Pohon ... 15
Pengujian Kelarutan Mineral Makro dalam Rumen dengan Teknik in Vitro ... 16
Analisis Data ... 20
HASIL DAN PEMBAHASAN ... 21
Kandungan Nutrisi Legum Pohon ... 21
Degradasi Pakan ... 22
Kelarutan Mineral ... 24
Produksi VFA ... 25
Produksi Amonia ... 26
Hubungan Laju Degradasi dengan Produksi VFA dan NH3 ... 27
KESIMPULAN DAN SARAN ... 29
Kesimpulan ... 29
Saran ... 29
UCAPAN TERIMAKASIH ... 30
DAFTAR PUSTAKA ... 31
LAMPIRAN ... 35
DAFTAR TABEL
Nomor Halaman
1. Komposisi Abu, Ca dan P Beberapa Legum Pohon (%BK) ... 3
2. Kandungan Mineral Makro pada Hijauan di Beberapa Lokasi di Pulau Jawa (%BK) ... 9
3. Larutan Standar Mineral Fosfor ... 18
4. Kandungan Nutrisi Beberapa Jenis Legum Pohon (%BK) ... 21
5. Laju Degradasi Beberapa Legum Pohon (%BK) ... 22
6. Kelarutan Mineral Selama Inkubasi 12 dan 24 Jam (mg/kg BK Sampel) ... 24
7. Produksi VFA Beberapa Legum Pohon ... 26
8. Konsentrasi NH3 Beberapa Jenis Legum Pohon ... 27
DAFTAR GAMBAR
Nomor Halaman
1. Daun Angsana dan Bunga (a), Buah Angsana (b) ... 4
2. Daun Turi (a) dan Bunga (b) ... 5
3. Daun Gamal (a) dan Biji (b) ... 6
4. Daun Lamtoro dan Bunga (a), Buah Polong Lamtoro (b) ... 6
DAFTAR LAMPIRAN
Nomor Halaman
1. Analisis Ragam Degradasi Pakan Masa Inkubasi 0 Jam ... 36
2. Analisis Ragam Degradasi Pakan Masa Inkubasi 3 Jam ... 36
3. Analisis Ragam Degradasi Pakan Masa Inkubasi 6 Jam ... 36
4. Analisis Ragam Degradasi Pakan Masa Inkubasi 9 Jam ... 37
5. Analisis Ragam Degradasi Pakan Masa Inkubasi 12 Jam ... 37
6. Analisis Ragam Degradasi Pakan Masa Inkubasi 24 Jam ... 37
7. Analisis Ragam Degradasi Pakan Masa Inkubasi 48 Jam ... 38
8. Analisis Ragam Degradasi Pakan Masa Inkubasi 72 Jam ... 38
9. Analisis Ragam Fraksi Terlarut (nilai a) ... 38
10.Analisis Ragam Fraksi Tidak Larut (nilai b) ... 38
11.Analisis Ragam Potensial Degradasi (nilai a+b) ... 39
12.Analisis Ragam Laju Degradasi (nilai c) ... 39
13.Analisis Ragam Produksi VFA Masa Inkubasi 12 Jam ... 39
14.Analisis Ragam Produksi VFA Masa Inkubasi 24 Jam ... 39
15.Analisis Ragam Produksi NH3 Masa Inkubasi 12 Jam ... 40
16.Analisis Ragam Produksi NH3 Masa Inkubasi 24 Jam ... 40
17.Analisis Ragam Ca Terlarut Masa Inkubasi 12 Jam ... 40
18.Analisis Ragam Ca Tidak Terlarut Masa Inkubasi 12 Jam ... 41
19.Analisis Ragam P Terlarut Masa Inkubasi 12 Jam ... 41
20.Analisis Ragam P Tidak Terlarut Masa Inkubasi 12 Jam ... 41
21.Analisis Ragam Ca Terlarut Masa Inkubasi 24 Jam ... 42
22.Analisis Ragam Ca Tidak Terlarut Masa Inkubasi 24 Jam ... 42
23.Analisis Ragam P Terlarut Masa Inkubasi 24 Jam ... 42
24.Analisis Ragam P Tidak Terlarut Masa Inkubasi 24 Jam ... 43
PENDAHULUAN
Latar Belakang
Hijauan merupakan bahan pakan yang dibutuhkan ternak ruminansia, namun
ketersediaanya sangat tergantung pada musim. Pada musim kemarau sering kali
terjadi defisiensi mineral yang disebabkan menurunnya kualitas padang
penggembalaan alam. Hal ini menyebabkan produktivitas ternak yang rendah. Upaya
perbaikan gizi pada ternak ruminansia telah banyak dilakukan dengan cara
pemberian hijauan leguminosa.
Hijauan legum umumnya hanya digunakan sebagai sumber protein. Namun,
demikian dengan melihat kandungan mineral makro yang cukup tinggi, pemanfaatan
legum pohon dapat juga ditujukan sebagai sumber mineral makro. Seperti yang
dilaporkan oleh Underwood dan Suttle (1999) bahkan kandungan mineral legum
pohon cukup tinggi dibandingkan rumput. Hal ini juga sejalan dengan penelitian
Sutardi et al. (1994) yang melaporkan bahwa legum pohon pada umumnya
kandungan mineralnya cukup tinggi terutama kalsium, sehingga dapat digunakan
mengatasi kekurangan mineral.
Mineral merupakan zat makanan yang berperan penting pada berbagai proses
fisiologis dalam tubuh ternak dan mempengaruhi efisiensi produksi. Kebutuhan
mineral pada ternak ruminansia tidak hanya untuk mencukupi kebutuhan pokok
ternak sendiri tetapi juga dibutuhkan oleh mikroba didalam rumen. Mineral dalam
rumen digunakan untuk aktivitas pembentukan sel, aktivitas selulolitik dan
pertumbuhan mikroba. Mineral juga berguna dalam mengatur tekanan osmotik,
sebagai larutan penyangga, sebagai potensi reduksi dan mengatur laju kelarutan di
dalam rumen.
Kebutuhan mineral untuk ternak diperoleh dari kuantitas dan ketersediaannya
(bioavailability). Bioavailability mineral adalah mineral yang siap diserap dan
dimanfaatkan oleh ternak. Mineral tersedia yang dimaksud adalah mineral yang
terlarut dalam rumen sehingga siap diserap oleh usus.
Kandungan mineral makro dalam hijauan dipengaruhi oleh beberapa faktor,
diantaranya spesies, umur tanaman, pengelolaan, iklim dan tipe tanah (McDowell
dan Valle, 2000). Pada daerah yang masih mengandalkan sistem ekstensif,
kualitas hijauan relatif baik kandungan mineralnya, sedangkan pada musim kering
akan terjadi sebaliknya.
Perumusan Masalah
Kajian penggunaan hijauan legum pohon pada umumnya hanya ditujukan
sebagai sumber protein, namun dengan melihat kandungan mineral makro yang
cukup tinggi pemanfaatan legum pohon dapat juga ditujukan sebagai sumber mineral
makro. Akan tetapi, informasi mengenai ketersediaan (bioavailability) mineral dalam
rumen yang berasal dari legum pohon masih sedikit. Mekanisme hubungan antara
kelarutan mineral makro di dalam rumen dengan proses fermentasi juga belum
banyak diketahui, terutama sejauh mana peranan mineral dalam menunjang aktivitas
mikroba rumen sehingga diperoleh produk-produk fermentasi yang optimal.
Tujuan
Penelitian ini bertujuan untuk mengetahui kelarutan mineral makro (Ca dan
TINJAUAN PUSTAKA
Leguminosa Pohon
Leguminosa pohon memiliki beberapa karakteristik yang khas antara lain:
kandungan proteinnya yang tinggi (12,5–20,7 %) dengan kecernaan yang lebih tinggi
dari rumput, kandungan mineral (khususnya kalsium dan fosfor) dan vitamin yang
tinggi. Selain itu leguminosa pohon mampu mensuplai protein fermentable dan by
pass karena mengandung zat anti nurisi berupa tannin. Secara ekologis dan ekonomis
leguminosa pohon dapat meningkatkan kesuburan tanah, melidungi tanah dari erosi
dan merupakan penghasil kayu yang bermutu (Allen dan Allen, 1981).
Fleming (1973) mengatakan bahwa secara umum kandungan elemen mineral
pada leguminosa lebih banyak dibandingkan pada rumput. Mineral Ca dan Mg pada
legum pakan lebih tinggi dari rumput (Serra et al., 1996). Hal serupa dilaporkan
Sutardi et al. (1994) yang menganalisa kandungan abu, Ca dan P beberapa legum
pohon ( Tabel 1).
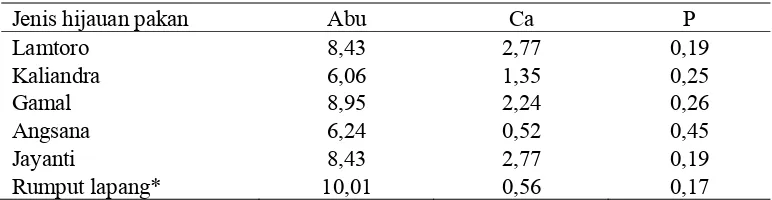
Tabel 1. Komposisi Abu, Ca dan P Beberapa Legum Pohon (%BK)
Jenis hijauan pakan Abu Ca P
Ket: * hasil analisis laboiratorium Ilmu Nutrisi dan Teknologi Pakan
Mineral Ca dalam hijauan pakan umumnya terikat dalam kalsium-oksalat
(Ward dan Herber, 1982) bahkan dalam dinding sel (McManus et al., 1979) sehingga
sulit untuk dicerna. Oleh karena itu ketersediaan mineral tidak selalu dikaitkan
dengan kuantitas mineral. Untuk ternak ruminansia, mineral tidak hanya dibutuhkan
oleh ternak tetapi juga oleh mikroba didalam rumen yang dibutuhkan untuk
menunjang proses fermentasi yang optimal (Durand dan Kawashima, 1980).
Mikroba sangat berperan penting dalam proses fermentasi. Sebagian besar
(70-80%) suplai energi pada ternak ruminansia berasal dari proses fermentasi
didalam rumen, sehingga kebutuhan mikroba harus benar-benar diperhatikan.
fermentasi. Mikroba rumen juga penting dalam utilisasi sulfur pada sapi perah
(Kandylis, 1984). Penambahan sulfur pada rumen yang mengandung urea dapat
meningkatkan retensi nitrogen. Sulfur organik dapat digunakan oleh mikroba untuk
sintesis asam amino.
Angsana (Pterocarpus indicus)
Angsana merupakan legum pohon yang dapat tumbuh dengan cepat mencapai
tinggi tegakan 35-40m (Allen dan Allen, 1981). Penggunaan angsana sebagai pakan
ternak dibatasi oleh adanya tannin yang merupakan senyawa kimia yang bersifat
racun. Dalam jumlah kecil, tannin dipandang menguntungkan bagi ternak ruminansia
karena dapat mencegah degradasi protein yang berlebihan oleh mikroorganisme
rumen, tetapi kandungan tannin yang tinggi dapat mengganggu metabolisme protein
dan karbohidrat sehingga dapat menurunkan penampilan ternak (Leng, 1997).
Bentuk daun, bunga dan buah angsana dapat dilihat pada Gambar 1.
(a) (b)
Gambar 1. Daun Angsana dan Bunga (a), Buah Angsana (b)
Kandungan nutrisi angsana menurut analisa proksimat adalah sebagai berikut:
protein 18,35%, serat kasar 24,12%, kalsium 1,02%, fosfor 0,86% dan kandungan
tannin mencapai 1,15%. Menurut Widiyanto (1992), penggunaan tepung angsana
dalam ransum akan meningkatkan konsumsi bahan kering yang dipengaruhi oleh
Turi (Sesbania grandiflora)
Tanaman turi hanya cocok pada daerah tropika. Tanaman turi termasuk
legum dengan tipe daun majemuk sederhana dan memiliki bunga yang besar
berwarna putih dan merah, berfungsi sebagai makanan ternak dan pupuk hijau.
Tinggi tegak mencapai 12 m, toleran terhadap kekeringan hingga 7 bulan. Daunnya
banyak digunakan sebagai pakan ternak dan mempunyai sifat toleran terhadap tanah
dengan kadar garam tinggi (Rosa, 1998). Bentuk daun dan bunga turi dapat dilihat
pada Gambar 2.
(a) (b)
Gambar 2. Daun Turi (a) dan Bunga (b)
Daun turi muda mengandung karbohidrat 42,30%, protein kasar mencapai
30,13% dan serat kasar 5,1% (Kareem dan Sundarajaj, 1967). Pemberian pada sapi
umumnya 1.8 kg segar/hari. Batasan penggunaanya dikarenakan adanya kandungan
antinutrisi berupa conavanine dan saponin.
Gamal (Gliricidia sepium)
Gamal merupakan legum pohon yang tinggi tegaknya mencapai 10 m, tipe
daunnya majemuk sederhana dan memiliki bunga berbentuk kupu-kupu berwarna
putih dan merah jambu. Tanaman ini toleran terhadap kekeringan hingga 8 bulan dan
toleran terhadap tanah yang memiliki kadar garam yang tinggi (Rosa, 1998). Daun
gamal digunakan sebagai hijauan pakan ternak. Daya cerna dari gamal berkisar
antara 50-75%, dengan kandungan protein kasar 22-27%, serat kasar 14%, NDF 45%
(a) (b)
Gambar 3. Daun Gamal (a) dan Biji (b)
Lamtoro (Leucaena leococephala)
Tanaman lamtoro hanya tumbuh pada daerah tropis dan subtropis, tahan
musim kering sampai 8 bulan bahkan 10 bulan, tidak tahan terhadap embun beku
(Karti, 1998). Merupakan legum pohon yang memiliki tipe daun majemuk ganda.
Banyak digunakan sebagai hijauan makanan ternak, peneduh dan tanaman hias
(Skerman, 1977). Bentuk daun, bunga dan buah lamtoro dapat dilihat pada Gambar
4.
(a) (b)
Gambar 4. Daun Lamtoro dan Bunga (a), Buah Polong Lamtoro (b)
Kecernaan dari lamtoro berkisar antara 65-87% dengan kandungan protein
kasar 25,9%, kalsium 2,36%, fosfor 0,23%. Lamtoro mengandung zat antinutrisi
berupa tannin (10.15%) dan mimosin (3-5%) (Skerman,1977). Pengaruh mimosin
pada ternak yaitu dapat menyebabkan kerontokan bulu dan mempengaruhi fetus pada
Kaliandra (Calliandra calothyrsus)
Kaliandra merupakan tanaman legum yang tinggi tegaknya umumnya
mencapai 4-6 m. Memiliki tipe daun majemuk ganda dan berwarna hijau gelap.
Tanaman ini dapat tumbuh pada periode musim kering 3-6 bulan tanpa kehilangan
daunnya namun tidak tahan genangan. Tamanan ini dapat tumbuh pada tanah yang
kurang subur karena adanya simbiosis yang menguntungkan antara mikroba
rhizobium dan mikoriza. Bentuk daun dan buah polong kaliandra dapat dilihat pada
Gambar 5.
(a) (b)
Gambar 5. Daun Kaliandra (a) dan Buah Polong (b)
Produksi hijauan dan dahan muda, menghasilkan bahan segar 46,2
ton/ha/tahun dengan kandungan protein 22% dan mengandung tannin cukup tinggi
(1-3%) sehingga daya cernanya rendah (35-42%). Pemberian pada ternak tidak
bermasalah, dari penelitian domba yang diberi hijauan kaliandra sebanyak 40-60%
dari pakan menghasilkan respon pertumbuhan yang baik (Karti, 1998).
Kandungan Anti Nutrisi Leguminosa
Anti nutrisi pada leguminosa merupakan faktor pembatas dalam
penggunannya sebagai pakan, karena sebagian besar dari zat tersebut bersifat racun
bagi ternak. Secara alami, anti nutrisi tersebut terdapat dalam tanaman sebagai
proteksi dari serangan hama. Tannin merupakan salah satu anti nutrisi yang sering
terdapat dalam leguminosa (Leng, 1997), selain itu juga ada mimosin, asam sianida
Tannin: merupakan senyawa polifenol yang terdapat alami dengan sifat utamanya dapat berikatan dengan protein. Menurut Prince et al. (1980), tannin memiliki
beberapa sifat yang dapat bersifat sebagai anti nutrisi, yaitu rasa sepat dari tannin
dapat menurunkan konsumsi ternak. Tannin juga dapat membentuk ikatan kompleks
dengan protein dan zat makanan lainnya, dengan protein endogen dan dengan
enzim-enzim pencernaan. Penyerapan tannin dan hasil hidrolisanya dapat meracuni ternak
dan mengganggu fungsi saluran pencernaan karena tanin dapat melukai saluran
pencernaan.
Tanin berpengaruh terhadap ternak terutama pada pakan yang mengandung
tannin tinggi. Pada saat mastikasi tannin terikat dengan saliva dapat menjadikan
pakan tidak palatabel sehingga dapat menurunkan konsumsi pakan (Makkar, 1993;
Cheeke dan Shull, 1985). Lebih lanjut dijelaskan bahwa tanin dapat menonaktifkan
enzim-enzim yang dihasilkan oleh mikroba dan dapat juga menimbulkan keracunan
bagi mikroba (Makkar, 1993).
Menurut McLeod (1974), tannin digolongkan menjadi dua golongan yaitu
tanin terhidrolisis dan tanin terkondensasi. Tanin terhidrolisis merupakan golongan
tanin yang sudah dihidrolisis menjadi gula sederhana dan asam fenolat. Sedangkan
tanin terkondensasi adalah tanin yang terjadi karena kondensasi flavanol, dikenal
sebagai proathocyanidins, tidak mudah dihidrolisis dan terdapat dalam bentuk yang
sangat kompleks (Cheeke dan Shull, 1985).
Mimosin: merupakan asam amino bebas yang sedikit larut dalam air. Suttie (1979) menyatakan bahwa kandungan mimosin dipengaruhi oleh varietas tanaman lamtoro
dan bagian-bagian tanaman. Ternak ruminansia lebih tahan terhadap pengaruh
mimosin dibanding ternak berlambung tunggal. Karena pada rumen terdapat bakteri
yang mampu mengubah mimosin menjadi 3-hidroksi-4-1 (H)-piridon atau DHP yang
kurang beracun.
Beberapa penelitian menyatakan bahwa untuk menurunkan daya racun
mimosin dari lamtoro adalah dengan memanaskan, menghancurkan jaringan daun
lamtoro, menambahkan garam besi sulfat dan pencucian. Matsumoto et al. (1951)
menyatakan bahwa pemanasan daun lamtoro pada suhu 90oC selama 20 jam mampu
menurunkan pengaruh racun mimosin yang dibuktikan dengan memberikan daun
Kandungan Mineral Legum Pohon
Faktor pembatas produksi ruminansia di daerah tropis kekurangan nutrisi
terutama defisien energi dan protein. Dalam keadaan tersebut produksi dan
reproduksi terlihat tidak normal meskipun bahan makanan diberikan dalam jumlah
cukup banyak. Keadaan ini biasanya diduga defisiensi, kelebihan atau
ketidakseimbangan mineral dalam bahan pakan, khususnya hijauan yang merupakan
sumber makanan utama bagi ruminansia. Faktor yang mempengaruhi kebutuhan
mineral adalah (1) jenis dan level produksi (2) level dan bentuk ikatan (3) bangsa
ternak (4) proses adaptasi (5) tingkat konsumsi mineral (6) umur ternak (7) adanya
hubungan dengan zat makanan lain (Parakasi, 1999).
Secara umum mineral dipergunakan dalam memelihara, pertumbuhan, dan
pergantian sel–sel dan jaringan yang rusak dalam tubuh ternak (Chase dan Sniffen,
1998). Berdasarkan jumlah yang dibutuhkan mineral dibagi atas mineral makro,
mikro dan trace element. Mineral makro antara lain kalsium (Ca), magnesium (Mg),
fosfor (P), kalium (K), sulfur (S), natrium (Na) dan klor (Cl). Mineral ini dibutuhkan
dalam jumlah yang cukup, namun seringkali terjadi defisiensi yang nantinya
berakibat fatal pada ternak.
Kandungan mineral pakan sangat bervariasi yang tergantung pada berbagai faktor
diantaranya spesies, tipe tanah, iklim dan umur tanaman (Underwood, 1981). Pada
umumnya kandungan mineral seperti Ca dan Mg pada legum lebih tinggi dari rumput
(Serra et al., 1995). Legum pohon dapat dijadikan sebagai sumber mineral terutama
Ca bagi ternak ruminansia.
Di daerah Nusa Tenggara, kandungan mineral hijauan sangat tergantung pada
ketersediaan air. Pada musim hujan kualitas hijauan relatif baik sehingga kandungan
mineralnya lebih baik, namun pada musim kering terjadi hal yang sebaliknya.
Kondisi ini juga terjadi di Pulau Jawa bahwa kandungan mineral makro pada hijauan
sangat bervariasi seperti yang dilaporkan Kawashima et al., (1989) pada tabel 2.
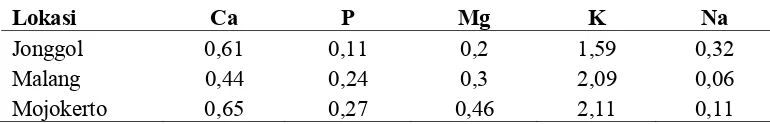
Tabel 2. Kandungan Mineral Makro pada Hijauan di Beberapa Lokasi di Pulau Jawa (%BK)
Lokasi Ca P Mg K Na
Jonggol 0,61 0,11 0,2 1,59 0,32
Malang 0,44 0,24 0,3 2,09 0,06
Ketersediaan Mineral Makro dalam Hijauan
Secara umum spesies legum mengandung mineral makro lebih tinggi dari
pada rumput. Di daerah tropik kandungan mineral umumnya lebih rendah
dibandingkan di daerah temperate. Konsentrasi rata–rata mineral Ca pada legum
untuk daerah tropik dan temperate masing–masing 19,1 dan 14,2 g/kg BK,
sedangkan untuk rumput masing–masing 3,8 dan 3,7 g/kg BK. Demikian juga
konsentrasi Na pada hijauan di daerah tropik 50% lebih rendah dari pada di daerah
temperate.
Kecukupan kalsium pada ternak yang digembalakan umumnya dapat
terpenuhi, khususnya untuk pastura yang mengandung legum. Dengan demikian,
penambahan hujauan pohon pada pastura dianjurkan untuk meningkatakan
ketersediaan mineral makro. Kandungan Ca pada bagian daun hijauan dua kali lebih
tinggi dari pada bagian batang. Penyerapan Ca umumnya terjadi di usus halus,
namun ada sebagian di dalam rumen (Yano et al, 1991). Penyerapan Ca (Calcium
absorbability = ACa) untuk setiap sumber hijauan berbeda-beda, dengan rata–rata
0,68 (AFRC, 1991). Pada beberapa hijauan nilai ACa lebih rendah disebabkan oleh
keberadaan kalsium oksalat yang sulit dicerna didalam rumen (Ward et al., 1979).
Bamualim (2004) melaporkan bahwa di daerah Timor kandungan mineral
fosfor legum adalah 1,5%, relatif lebih tinggi dibandingkan dengan kebutuhan seekor
ternak, namun demikian pada musim kemarau kandungannya relatif rendah dan akan
meningkat kembali pada musim hujan. Defisiensi ini terjadi juga pada mineral Na,
Cu dan Se.
Mineral Ca dan P
Kalsium (Ca) adalah mineral yang paling banyak dibutuhkan di dalam tubuh.
Secara kuantitatif fungsi utama kalsium adalah pada pembentukan tulang. Tulang
tidak saja berfungsi untuk menunjang struktur komponen tubuh tetapi juga
mempunyai fungsi fisiologis penting dalam jaringan dalam menyediakan kalsium
untuk mempertahankan sistem homeostasis tubuh (Piliang, 2001). Fungsi lain dari
kalsium yaitu untuk perkembangan gigi, produksi air susu, transmisi impuls saraf,
pemeliharaan eksitabilitas urat daging yang normal (bersama-sama dengan K dan
Na), regulasi denyut jantung, geraka-gerakan urat daging, pembekuan darah dan
kalsium pada ternak dapat menyebabkan riketsia, osteoporosis, osteomalasia,
pertumbuhan terlambat, hipertropi kelenjar parathiroid, dan milk fever (Underwood,
1981).
Mineral P sangat penting peran biokimia dan fiologisnya. Fosfor dideposit
dalam tulang dalam bentuk kalsium-hidroksi appetite {Ca10(PO4)6(OH)2)}. Fosfor
merupakan komponen dari fosfolipid yang mempengaruhi permiabilitas sel; juga
merupakan komponen dari meilin pembungkus urat saraf; banyak transfer energi
dalam sel yang melibatkan ikatan fosfat yang kaya energi dalam ATP; fosfor
memegang peran dalam sistem buffer dari darah; mengaktifkan beberapa vitamin B
(tiamin, niasin, piridoksin, riboflavin, biotin, dan asam pantotenik) untuk membentuk
koenzim yang dibutuhkan dalam proses fosforilasi awal, fosfor juga merupakan
bagian dari matrik dNA dan RNA (Parakkasi, 1999).
Fungsi fosfor antara lain untuk pembentukan dan pemeliharaan tulang dan
gigi, sekresi normal air susu, aktivator enzim–enzim dan metabolisme asam amino
(Piliang, 2001). Defisien Fosfor pada ternak dapat menyebabkan riketsia, sifat
memakan makanan yang aneh–aneh (pica appetite), menurunkan reproduksi dan
ukuran tubuh kecil (Underwood, 1981).
Durand dan Kawashima (1980) menyatakan bahwa, didalam rumen mineral
dipergunakan untuk berbagai aktifitas antara lain untuk pembentukan sel, aktivitas
selulolitik dan pertumbuhan mikroba. Disamping itu, mineral juga dipergunakan
dalam mengatur tekanan osmotik, buffering capacity, potensial reduksi dan kelarutan
di dalam rumen.
Cairan Rumen
Cairan rumen yang merupakan sumber inokula yang dapat dengan cepat
menghancurkan plasmalema dan banyak struktur sitoplasma dari sel tanaman (Cheng
et al., 1980). Penghuni terbesar dalam cairan rumen adalah bakteri yaitu 1010–
1012/ml cairan rumen dan populasi terbesar kedua diduduki oleh protozoa yang dapat
mencapai 105 – 106 sel/ml, namun demikian karena ukuran tubuhnya lebih besar
dari bakteri maka biomassanya ternyata cukup besar yakni mengandung lebih kurang
Teknik In vitro
Untuk mempelajari pemanfaatan bahan makanan pada ruminansia, ada dua
teknik yang dapat digunakan yaitu in vitro dan in vivo (Tilley dan Terry, 1963).
Teknik in vitro tergantung dari mikroba yang diambil dari hewan. Kecernaan secara
in vitro memiliki keuntungan antara lain pelaksanaannya mudah, mengurangi resiko
kematian ternak, lebih ekonomis, mewakili penampilan ternak. Kelemahannya
adalah medium kecernaan pada in vitro tidak mungkin sama dengan kecernaan in
vivo yang langsung menggunakan ternak. Syarat–syarat yang perlu diperhatikan
dalam membuat teknik in vitro adanya larutan penyangga (buffer) dan media
makanan. Temperatur optimumnya 390C, dengan pH optimum 6,7– ,0.
Metabolisme Rumen
Bahan makanan yang masuk ke dalam alat pencernaan akan mengalami
perubahan fisik dan kimia. Menurut Sutardi (1980) proses pencernaan pada ternak
ruminansia terjadi secara mekanis (di dalam mulut), secara fermentatif (oleh enzim–
enzim yang berasal dari mikroba rumen dan secara hidrolitis (oleh enzim–enzim
pencernaan hewan induk semang). Organ pencernaan ternak ruminansia terdiri atas
mulut, perut, usus halus, dan organ pencernaan bagian belakang. Perut ternak
ruminansia terdiri atas 4 bagian, yaitu rumen (perut beludru), retikulum (perut jala),
omasum (perut buku) dan abomasum (perut sejati) (Arora, 1989).
Rumen adalah suatu ekosistem yang kompleks yang dihuni oleh
beranekaragam mikroba anaerob. Mikroba tersebut terdiri atas bakteri, protozoa dan
fungi yang memegang peranan penting dalam pencernaan pakan. Bakteri merupakan
penghuni terbesar dalam rumen yang kepadatannya mencapai 1010–1012/ml cairan
rumen, diikuti protozoa 105–106/ml cairan rumen (Ogimoto dan Imai, 1981). Adanya
mikroba dalam rumen menyebabkan ruminansia memiliki kemampuan untuk
mencerna Nitrogen Bukan Protein (BNP) dan karbohidrat struktural tanaman
(selulosa dan hemiselulosa) melalui proses fermentasi menjadi asam lemak terbang
(VFA) (Demeyer, 1981), sedangkan protein terfermentasi menjadi amoniak.
Pencernaan mikrobial pada ruminansia memegang peranan yang sangat penting,
karena diperkirakan sekitar 70–85% dari bahan kering pakan yang biasa dikonsumsi
oleh ternak dapat dicerna dalam rumen (Benerjee, 1978). Pencernaan fermentatif
tertentu menjadi senyawa lain yang sama sekali berbeda dari molekul zat asalnya
(Amin,1997). Hasil pencernaan fermentatif berupa VFA, NH3, dan air diserap
sebagian di rumen dan sebagian lagi di omasum. Selanjutnya pakan yang tidak
dicerna mengalir ke abomasum dan dicerna secara hidrolitik oleh enzim–enzim
pencernaan (Sutardi, 1977).
Kontribusi proses fermentatif cukup besar, tetapi pada tahap tersebut
sebagian energi pakan ada yang terbuang sebagai gas metan dan panas fermentasi.
Disamping VFA dan NH3, fermentasi dalam rumen juga menghasilkan gas CO2, H2,
dan metan yang dikeluarkan dari rumen melalui proses eruktasi (Arora, 1989).
Peranan mikroba dalam proses fermentasi sangat besar. Sebagian besar
(70-80%) suplai energi pada ternak ruminansia berasal dari proses fermentasi di dalam
rumen, sehingga kebutuhan mikroba harus benar–benar diperhatikan. Kelarutan
mineral baik langsung maupun tidak langsung akan mempengaruhi produk
fermentasi. Mikroba rumen mempunyai peranan penting dalam utilisasi sulfur pada
sapi perah (Kandylis, 1984). Penambahan sulfur pada rumen yang mengandung urea
dapat meningkatkan retensi nitrogen. Sulfur organik dapat digunakan oleh mikroba
untuk sintesis asam amino.
Volatil Fatty Acids (VFA)
Volatil Fatty Acids (VFA) yang biasa disebut asam lemak terbang juga
merupakan salah satu produk fermentasi di dalam rumen. Asam lemak terbang
merupakan produk fermentasi dari karbohidrat. Konsentrasi VFA cairan rumen dapat
digunakan sebagai salah satu tolak ukur fermentabilitas pakan dan sangat erat
kaitannya dengan aktivitas mikroba rumen (Sewet, 1997). Mikroba rumen
menghidrolisis selulosa menjadi monosakarida yang kemudian membentuk VFA
melalui fermentasi. VFA yang dihasilkan adalah asam asetat, propionat dan butirat.
Asam lemak yang lain terdapat didalam rumen dalam jumlah yang kecil adalah asam
isobutirat, isovalerat, dan lain-lain.
Banyaknya VFA yang dihasilkan di dalam rumen sangat bervariasi
tergantung pada jenis ransum yang dikonsumsi (MC Donald et al. 1988). Konsentrasi
VFA total yang layak bagi kelangsungan hidup ternak adalah 80–160 mM dengan
Amonia (NH3)
Amonia (NH3) merupakan salah satu hasil perombakan protein oleh mikroba
rumen. Konsentrasi NH3 cairan rumen akan meningkat jika populasi protozoa
meningkat karena protozoa ikut berperan dalam proses daur ulang nitrogen (Sewet,
1997). Protozoa akan mencerna protein bakteri yang dimangsanya kemudian
dikeluarkan kembali dalam sistem rumen dalam bentuk peptida, asam-asam amino
dan amonia.
Produksi amonia tergantung pada kelarutan protein ransum, jumlah protein
ransum, lamanya makanan di dalam rumen dan pH rumen (Orskov, 1982). Sebagian
besar mikroba rumen menggunakan NH3 untuk perbanyakan dirinya terutama dalam
proses sintesis selnya (Sutardi, 1979). Konsentrasi optimum NH3 untuk menunjang
pertumbuhan mikroba rumen berkisar 85–300 mg/l atau 6–21 mM (McDonald et al.,
METODE
Lokasi dan Waktu
Penelitian ini dilaksanakan di Laboratorium Ilmu Nutrisi Ternak Perah,
Departemen Ilmu Nutrisi dan Makanan Ternak, Fakultas Peternakan Institut
Pertanian Bogor mulai bulan Januari sampai September 2005. Penelitian ini terdiri
dari tiga tahap pengerjaan yaitu laju degradasi bahan kering legum pohon, pengujian
kelarutan mineral kalsium dan fosfor, dan fermentabilitas legum pohon di dalam
rumen (produksi VFA dan NH3).
Materi
1. Pakan Percobaan
Legum pohon yang mempunyai potensi sebagai sumber mineral makro yang
akan digunakan dalam penelitian ini adalah angsana (Pterocarpus indicus), turi
(Sesbania glandiflora), gamal (Gliricidia sepium), lamtoro (Leucaena leucocephala),
dan kaliandra (Calliandra calothyrsus). Bagian yang digunakan dari legum pohon
tersebut adalah daunnya. Proporsi daun yang tua lebih banyak digunakan dari pada
daun yang muda (pucuk daun). Legum pohon yang digunakan didapatkan di daerah
Bogor.
Daun legum pohon dikeringkan dibawah sinar matahari dan digiling halus.
Daun yang sudah digiling digunakan untuk inkubasi dalam cairan rumen.
2. Cairan Rumen
Cairan rumen yang digunakan dalam penelitian ini adalah cairan rumen
domba yang diambil dari Rumah Potong Hewan (RPH) di Kecamatan Ciampea.
Cairan rumen dimasukkan kedalam termos. Lamanya cairan rumen di dalam termos
selama perjalanan dari RPH ke tempat penelitian kurang lebih 30 menit.
Prosedur
1. Laju Degradasi Bahan Kering Legum Pohon
Parameter yang diukur dalam penelitian ini adalah laju degradasi pakan dari
masing–masing legum. Pengukuran laju degradasi dilakukan dengan menggunakan
teknik in vitro yang dimodifikasi. Sebanyak satu gram sampel dari berbagai jenis
fermentor, kemudian ditambahkan 8 ml cairan rumen dan 12 ml larutan buffer serta
dialiri gas CO2 selama 30 detik. Tabung fermentor ditutup dengan karet berventilasi
dan dimasukkan dalam shaker bath pada suhu 390C dengan waktu inkubasi 0, 3, 6, 9,
12, 24, 48, dan 72 jam, dengan demikian tabung fermentor yang digunakan
sebanyak 72 buah (dua waktu inkubasi x (lima jenis legum + satu blanko) duplo) x
tiga ulangan). Setelah inkubasi disentrifuge pada kecepatan 3.000 rpm selama 15
menit sehingga residu dan supernatannya terpisah. Sampel residu dikeringkan dalam
oven untuk diketahui bahan keringnya, sedangkan supernatan digunakan untuk
analisis VFAtotal dan NH3. Untuk menghitung laju degradasi legum dalam rumen
digunakan model matematik yang dikemukakan oleh Ørskov dan McDonald (1979),
yaitu :
Y = a + b( 1 – e - ct)
Keterangan :
Y = degradasi pakan (%BK)
a = degradasi pakan dari fraksi terlarut
b = degradasi pakan selama fermentasi
c = laju degradasi pakan (%/jam)
t = waktu inkubasi (jam)
2. Pengujian Kelarutan Mineral Makro dalam Rumen dengan Teknik in Vitro Percobaan ini dilakukan untuk mengetahui kelarutan mineral makro legum
pohon dalam rumen dengan menggunakan teknik in vitro.
Sampel yang telah diinkubasi seperti prosedur (1), setelah ketahui bahan
keringnya, diabukan dengan menggunakan metode pengabuan basah (wet ashing).
Mineral yang dianalisis adalah kalsium (Ca) dan Phosphor (P). Analisis mineral
dilakukan dengan menggunakan Atomic Absorption Spectrofotometric (AAS).
Kandungan residu mineral setiap sampel dalam tabung fermentor
memperlihatkan proporsi mineral yang terlarut dalam cairan rumen. Laju kelarutan
mineral dalam rumen tersebut dihitung dengan model :
Y = a + b (1 – e –ct)
Y = kelarutan mineral dalam cairan rumen (mg/gram sample)
b = kelarutan mineral selama dalam cairan rumen(mg/gram sample)
c = laju kelarutan mineral(mg/jam)
t = waktu inkubasi (jam)
Kelarutan mineral dihitung berdasarkan jumlah mineral dalam bahan pakan
dikurangi dengan mineral yang tersisa pada bahan pakan yang telah diinkubasi.
Pengukuran kadar mineral tersebut dilakukan dengan cara pengabuan basah (wet
ashing) berdasarkan prosedur Reitz et al. (1960). Sampel yang telah diinkubasi
secara in vitro dimasukkan kedalam tabung Erlenmeyer dan ditambahkan lima ml
HNO3, lalu dibiarkan selama satu jam sampai menjadi bening atau tidak ada buih.
Labu Erlenmeyer dipanaskan pada hot plate selama kurang lebih empat jam. Setelah
dingin ditambahkan 0.4 ml H2SO4 pekat, labu Erlenmeyer dipanaskan kembali. Pada
saat terjadi perubahan warna volume akan berkurang diteteskan larutan HClO4 dan
HNO3 (perbandingan 2:1). Perubahan warna dimulai dari warna coklat menjadi
kuning dan bening. Setelah bening, dipanaskan kembali selama 15 menit, lalu
ditambahkan dengan dua ml aquades dan 0.6 ml HCl pekat dan dipanaskan kembali
hingga larut. Setelah didinginkan, ditambahkan dengan aquades hingga 100 ml.
Pengukuran kadar Phospor dilakukan dengan menggunakan spektrofotometer
(UV Visible) dengan panjang gelombang 660 nm, sedangkan untuk analisis kalsium
dibaca konsentrasinya pada Spektrofotometer serapan atom (AAS).
a. Prosedur pengenceran untuk analisa mineral Ca
Pengenceran dilakukan untuk menurunkan kadar mineral dalam sampel
supaya tidak melebihi standar yang sudah ditetapkan. Langkah-langkah pengenceran
adalah sebagai berikut: sebanyak 0.5 ml larutan sampel (sesuai dengan tinggi
rendahnya kadar kalsium) yang sudah disiapkan dengan metode pengabuan basah
dilarutkan dengan aquades sampai menjadi lima ml pada tabung reaksi dengan
menggunakan pipet volumetrik, kemudian ditambahkan larutan lantan sebanyak 0.05
ml yang berfungsi sebagai pengikat terhadap ion-ion pengganggu. Larutan sampel
diaduk dengan menggunakan alat pengaduk vortex selama beberapa detik. Sampel
siap untuk dianalisa jumlah kandungan mineral kalsium dengan menggunakan mesin
b. Prosedur pengenceran untuk analisa mineral P
Sebelum pengukuran phospor, terlebih dahulu dibuat larutan standar dengan
konsentrasi 0, 2, 3, 4 dan 5 ppm. Larutan standar dibuat dengan beberapa tahap.
Larutan A (NH4)6Mo7O24) 10% = Ammonium molibdat 10%. Sebanyak sepuluh
gram ammonium molibdat ditambah dengan 60 ml aquades dan 28 ml H2SO4 pekat
secara bertahap, ditambahkan kembali aquades hingga 100 ml. Larutan B dibuat
dengan memasukkan sepuluh ml larutan A ke dalam labu takar 100 ml, kemudian
ditambahkan 60 ml aquades dan 5 gram FeSO4.7H2O. Larutan B dibuat menjadi 100
ml dengan menambahkan aquades. Larutan B digunakan dalam pembuatan larutan
standar seperti terlihat pada Tabel 3.
Tabel 3. Larutan Standar Mineral Fosfor
Sampel yang telah diabukan diambil 0,5 (sesuai dengan tinggi rendahnya
phospor dalam sampel), ditambahkan dua ml larutan B dan aquades hingga menjadi
5.0 ml. Selanjutnya dikocok dengan menggunakan vortex agar dapat homogen.
Sampel dan larutan standar siap untuk dianalisis kadar phospornya dengan
menggunakan spektrofotometer dengan panjang gelombang 660 nm.
3. Pengukuran Produksi VFAtotal dan NH3
Kecukupan mineral makro dalam rumen dapat dilihat dari kemampuan
mikroba dalam memfermentasi pakan. Mineral juga membantu dalam menjaga
kondisi pH optimal rumen. Untuk melihat hubungan kelarutan mineral makro dengan
fermentabilitas hijauan dalam rumen dilakukan pengukuran kadar asam lemak
terbang (Volatile Fatty Acids, VFA) dan amonia secara in vitro.
Analisis VFAtotal dan NH3 menggunakan supernatan yang telah dipisahkan
dari subtrat selama inkubasi 12 dan 24 jam.
a. Teknik Analisis VFA
Kadar VFAtotal diukur dengan menggunakan Steam Destilation Method.
Sebanyak lima ml supernatan sampel (yang diperoleh dari hasil fermentasi secara in
vitro selama 12 dan 24 jam) dimasukkan dalam tabung destilasi Markham dan
ditambahkan satu ml H2SO4 15%. Hasil destilasi ditampung dalam erlenmeyer yang
telah diisi lima ml NaOH 0,5 N hingga destilat mencapai 300 ml. Destilat yang
ditampung diberi 2 – 3 tetes indikator phenopthalein kemudian dititrasi dengan HCl
0,5 N hingga terjadi perubahan warna dari pink menjadi bening. Kadar VFAtotal
dihitung dengan rumus:
Kadar amonia diukur dengan metode Micro Diffusion Conway. Sebanyak
satu ml supernatan ditempatkan disalah satu ruang bersekat cawan conway dan disisi
yang lain ditambahkan satu ml Na2CO3 jenuh, sedangkan dibagian tengah cawan
ditempatkan satu ml asam borat yang beri indikator metil merah dan brom kresol
biru. Pada bagian pinggir cawan diolesi vaselin kemudian ditutup rapat. Cawan
digerakkan perlahan agar supernatan bercampur dengan Na2CO3 jenuh kemudian
diinkubasikan selama 24 jam pada suhu ruangan.
Amonia yang keluar akan diikat oleh asam borat, lalu dititrasi dengan H2SO4
0.005 N sampai terjadi perubahan warna biru menjadi merah jambu (pink). Kadar
amonia dihitung dengan rumus :
NH3 = [(Vs – Vo) x N-H2SO4 x 1000],
Keterangan :
NH3 dalam mmol
Vs : volume titran sampel
Analisis Data
Rancangan percobaan yang digunakan dalam penelitian in vitro adalah
Rancangan Acak kelompok (RAK) 5 x 3, yang terdiri dari 5 perlakuan legum yang
berbeda (angsana, turi, gamal, lamtoro dan kaliandra) dan 3 kelompok berdasarkan
cairan rumen yang berbeda.
Adapun model matematik rancangan tersebut adalah sebagai berikut :
Yij= µ + τi +βj + εij
Yij = Nilai pengamatan dari perlakuan ke-i dalam kelompok ke- j
µ = Nilai tengah populasi
τi = Pengaruh aditif dari perlakuan ke-i (jenis legum)
βj = Pengaruh aditif dari kelompok ke- j (periode cairan rumen)
εij = Pengaruh galat percobaan dari perlakuan ke-i pada kelompok ke-j
Data yang diperoleh dianalisis dengan menggunakan sidik ragam (ANOVA)
dan untuk mengetahui perbedaan antar perlakuan dilakukan uji kontras ortogonal
mengikuti Steel dan Torrie (1991).
Hubungan antara laju degradasi bahan kering legum pohon dalam rumen
dengan produksi VFA dan NH3 dilakukan dengan Uji Korelasi menggunakan SPSS
HASIL DAN PEMBAHASAN
Kandungan Nutrisi Legum Pohon
Legum pohon merupakan hijauan pakan yang dikenal sebagai sumber
protein. Namun, dengan melihat kandungan mineralnya yang tinggi, legum pohon
dapat dimanfaatkan sebagai sumber mineral makro. Kadungan mineral seperti Ca
dan Mg pada legum pohon lebih tinggi dari rumput (Serra et al., 1996). Kualitas
hijauan pakan ditentukan oleh komposisi kimia hijauan. Hasil analisa komposisi
kimia pakan yang digunakan dapat dilihat pada Tabel 4.
Tabel 4. Kandungan Nutrisi Beberapa Jenis Legum Pohon (%BK)
Nutrien Legum
Angsana Turi Gamal Lamtoro Kaliandra
Abu 6,27 7,60 7,62 7,24 4,46
Protein Kasar 20,15 20,99 18,58 22,76 18,70
Serat Kasar 23,25 21,71 19,74 18,47 19,46
Lemak Kasar 1,33 1,33 2,07 3,02 1,45
Beta-N 33,13 28,57 38,53 37,76 42,93
Ca 1,02 1,27 1,45 1,74 0,95
P 0,31 0,37 0,27 0,35 0,25
Ca : P 3 : 1 3 : 1 5 : 1 5 : 1 4 : 1
Keterangan: Hasil analisis Laboratorium Ilmu dan Teknologi Pakan
Legum pohon yang digunakan dalam penelitian ini memiliki kandungan
protein kasar 18,58 sampai 22,76%BK. Lamtoro memiliki kandungan protein kasar
(PK) yang lebih tinggi dan serat kasar (SK) yang lebih rendah dibandingkan dengan
legum yang lain, tetapi rasio Ca : P sangat besar (5:1). Rasio Ca : P dalam ransum
sangat penting dibandingkan dengan jumlahnya, karena kedua mineral tersebut
saling mempengaruhi. Rasio Ca : P yang direkomendasikan adalah (1:1) sampai
(2:1), tetapi pada umumnya ternak ruminansia lebih tahan terhap rasio Ca : P yang
luas dibanding hewan–hewan monogastrik (Parakkasi, 1999). Rasio Ca : P legum
angsana lebih baik diantara legum yang lain (3:1), tetapi kandungan serat kasar
angsana lebih tinggi yaitu 23,25 %BK. Kandungan serat kasar pada pakan dapat
Degradasi Pakan
Tingkat degradasi pakan dapat digunakan untuk mengetahui tinggi rendahnya
kualitas pakan. Jumlah pakan yang didegradasi dapat menunjukkan porsi pakan yang
dapat dicerna. Laju degradasi legum pohon dapat dilihat pada Tabel 5.
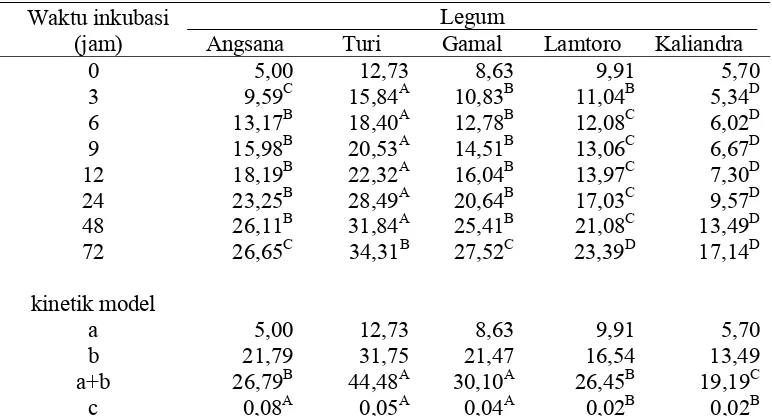
Tabel 5. Laju Degradasi Beberapa Legum Pohon (%BK)
Waktu inkubasi (jam)
Legum
Angsana Turi Gamal Lamtoro Kaliandra
0 5,00 12,73 8,63 9,91 5,70
b 21,79 31,75 21,47 16,54 13,49
a+b 26,79B 44,48A 30,10A 26,45B 19,19C
c 0,08A 0,05A 0,04A 0,02B 0,02B
Keterangan: superkrip yang berbeda pada baris yang sama menunjukkan berbeda sangat nyata (P<0,01)
a : fraksi terlarut (%BK)
b : materi tidak larut tetapi bisa difermentasi(%BK) (a+b) : potensial degradasi(%BK)
c : laju degradasi (jam-1)
Tingkat degradasi mulai menunjukkan perbedaan setelah inkubasi selama tiga
jam. Tingkat degradasi yang paling tinggi selama inkubasi tiga jam adalah daun turi
sedangkan yang paling rendah adalah daun kaliandra. Tingkat degradasi daun
angsana selama inkubasi tiga jam lebih rendah dari daun turi tetapi lebih tinggi dari
tingkat degradasi daun gamal dan lamtoro. Tingkat degradasi selama inkubasi 6
sampai 48 jam dimulai dari yang tertinggi sampai terendah adalah turi > gamal dan
angsana > lamtoro > kaliandra. Tingkat degradasi lamtoro lebih rendah karena
lamtoro dan kaliandra mengandung tannin yang lebih tinggi. Hal ini sesuai dengan
pernyataan Karti (1998), bahwa kaliandra mengandung tannin yang cukup tinggi (1–
3 %) sehingga daya cernanya rendah. Lamtoro mengandung zat anti nutrisi berupa
tannin (10,15%) dan mimosin (3–5%) dari bahan kering. Kecernaan lamtoro berkisar
Daun turi memiliki tingkat degradasi 6,32 % lebih tinggi dari daun lamtoro
dan 12,38% dari daun kaliandra selama inkubasi enam jam. Tingkat degradasi daun
lamtoro lebih rendah 8,35% dari daun turi, 4,22% dari daun angsana dan 2,07% dari
daun gamal, sedangkan tingkat degradasi kaliandra lebih rendah 15,02% dari daun
turi, 10,89% dari daun angsana dan 8,74% dari daun gamal selama inkubasi 12 jam.
Hal ini sesuai dengan pernyataan Khamseekhiew et al., (2001) bahwa degradasi
bahan kering pada gamal lebih tinggi dari lantoro pada periode inkubasi di atas 6
jam.
Selama inkubasi 24 jam tingkat degradasi lamtoro lebih rendah 11,46% dari
daun turi, 6,22% dari daun angsana dan 3,61% dari daun gamal. Demikian pula
kaliandra memiliki tingkat degradasi 13,68%, 18,92% dan 11,07% lebih rendah dari
daun angsana, turi dan gamal. Penambahan waktu inkubasi dari 12 jam hingga 24
jam dapat meningkatkan nilai tercerna pakan dengan rataan mencapai 4,23%.
Daun angsana dan gamal memiliki tingkat degradasi yang lebih tinggi
walaupun kandungan serat kasarnya juga tinggi. Hal ini sangat bertolak belakang
dengan daun lamtoro dan kaliandra yang kandungan serat kasarnya lebih rendah
namun tinggkat degradasinya juga rendah. Rendahnya tingkat degradasi daun
lamtoro dan kaliandra disebabkan adanya zat anti nutrisi berupa mimosin dan tannin
yang dapat melindungi lamtoro dan kaliandra dari mikroba yang akan mencernanya
selama inkubasi.
Fraksi terlarut (a) adalah kandungan bahan pakan yang dapat larut dalam air
dan dapat dicerna tanpa proses fermentasi seperti glukosa. Materi yang tidak larut
tetapi dapat didegradasi dalam waktu tertentu disimbolkan (b). Potensial degradasi
merupakan jumlah dari degradasi fraksi terlarut dan materi yang tidak terlarut (a+b).
Jumlah pakan yang didegradasi setiap satuan waktu disebut laju degradasi (c).
Degradasi fraksi terlarut (a) kelima jenis legum tersebut tidak berbeda. Sama
halnya dengan nilai fraksi terlarut (a) nilai fraksi tidak larut tetapi masih bisa
difermentasi (b) kelima jenis legum tersebut juga tidak menunjukkan perbedaan yang
nyata (P>0.05). Nilai fraksi tidak larut (b) daun kaliandra lebih rendah dibandingkan
dengan nilai b pada daun legum lainnya. Hal ini mengindikasikan bahwa potensi
substrat yang akan terfermentasi pada kaliandra tidak akan berperan signifikan dalam
Potensial degradasi (a+b) kelima jenis legum tersebut sangat berbeda nyata
(P<0.01). Daun turi dan gamal memiliki nilai potensial yang lebih tinggi, sedangkan
daun legum yang memiliki nilai potensial degradasi yang rendah adalah angsana dan
kaliandra. Laju degradasi (c) daun angsana, turi dan gamal sangat berbeda dengan
laju degradasi daun lamtoro dan kaliandra. Laju degradasi (c) daun lamtoro dan
kaliandra lebih rendah karena lamtoro dan kaliandra mengandung mimosin dan
tannin yang menghambat pencernaan oleh mikroba selama inkubasi dalam cairan
rumen.
Kelarutan Mineral
Mineral merupakan elemen–elemen atau unsur kimia selain dari karbon,
hidrogen dan nitrogen (Piliang, 2001). Mineral makro (Ca dan P) yang terlarut dan
yang tidak terlarut dapat dilihat pada Tabel 6.
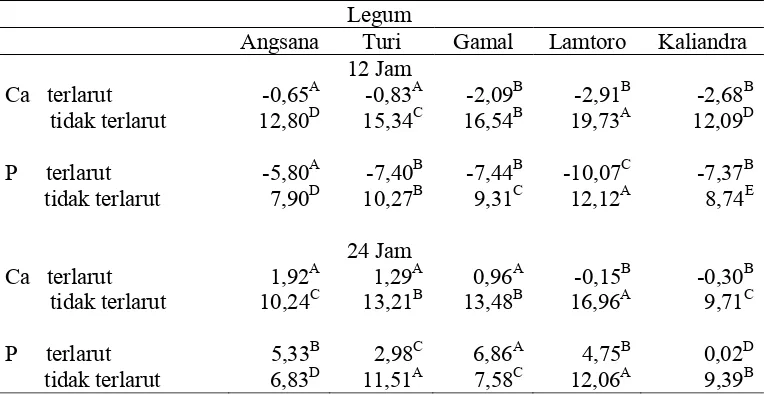
Tabel 6. Kelarutan Mineral Selama Inkubasi 12 dan 24 Jam (mg/kg BK Sampel)
Keterangan: superkrip yang berbeda pada baris yang sama menunjukkan berbeda sangat nyata (P<0,01)
Nilai kelarutan Ca dan P dalam cairan rumen berbeda untuk setiap jenis daun
legum. Selama inkubasi 12 jam kandungan Ca tidak larut yang tertinggi terdapat
pada lamtoro. Jumlah Ca terlarut selama inkubasi 12 jam pada masing–masing legum
bernilai negatif. Hal tersebut menunjukkan bahwa jumlah Ca yang tidak larut lebih
besar dibandingkan jumlah Ca yang terdapat dalam pakan. Hal ini bisa terjadi pada
sehingga Ca yang terdapat dalam daun mengalami proses mobilisasi menjadi
Ca-organik yakni terikat dalam sel mikroba. Dengan demikian jumlah Ca yang tidak
larut lebih tinggi dibandingkan jumlah Ca dalam pakan. Akibatnya jumlah Ca yang
terdapat larutan bernilai negatif.
Tidak jauh berbeda dengan Ca yang tidak terlarut, P yang tidak terlarut
tertinggi juga terdapat pada daun lamtoro, dan sedikit lebih rendah dibawahnya
adalah daun turi. P terlarut selama inkubasi 12 jam juga bernilai negatif. Hal tersebut
karena sifat mineral P yang sangat mobile. Kondisi ini tidak berbeda dengan yang
terjadi pada mineral Ca. Mineral P mengalami mobilisasi dan terikat oleh mikroba
menjadi P-organik atau mineral P diikat oleh mineral Ca yang pada saat itu menjadi
Ca-organik.
Nilai kelarutan Ca dan P dapat terlihat setelah inkubasi 24 jam. Kelarutan Ca
daun angsana, turi dan gamal sangat berbeda dengan lamtoro dan kaliandra.
Kelarutan Ca lamtoro dan kaliandra masih bernilai negatif. Kelarutan mineral P
kelima jenis legum setelah inkubasi 24 jam menunjukkan perbedaan yang sangat
nyata (P<0.01). Kelarutan mineral P diurutkan dari yang paling tinggi sampai
terendah berturut-turut adalah daun gamal > turi > angsana dan lamtoro > kaliandra.
Sama halnya dengan degradasi pakan rendahnya kelarutan Ca dan P pada
lamtoro dan kaliandra disebabkan oleh kandungan anti nutrisi pada kedua legum
tersebut. Lamtoro dan kaliandra mengandung antinutrisi mimosin dan tannin. Keir et
al., (1997) dalam penelitiannya menyebutkan bahwa mimosin dan tannin dapat
mengurangi fermentabilitas pakan oleh mikroba dalam rumen.
Produksi VFA
Volatile Fatty Acids (VFA) yang biasa disebut asam lemak terbang
merupakan hasil pencernaan karbohidrat oleh mikroba dalam cairan rumen.
Komponen asam lemak terbang dalam rumen adalah asam asetat, asam propionat,
asam – asam lemak rantai cabang berasal dari katabolisme protein. Konsentrasi asam
lemak terbang cairan rumen dapat digunakan sebagai salah satu tolak ukur
fermentabilitas pakan dan sangat erat kaitannya dengan aktivitas mikroba rumen
(Sewed, 1997). VFA mempunyai peran ganda yaitu sebagai sumber energi utama
bagi ternak dan sumber kerangka karbon untuk pembentukan protein mikroba
mengindikasikan bahwa energi yang tersedia bagi mikroba rumen juga semakin
tinggi sehingga aktivitas fermentasi mikroba juga dapat meningkat. Konsentrasi VFA
dari berbagai jenis legum pohon dapat dilihat pada Tabel 7.
Tabel 7. Produksi VFA Beberapa Legum Pohon
Produksi VFA (mM) Legum
Angsana Turi Gamal Lamtoro Kaliandra
12 jam 57,73 99,20 79,56 60,24 55,25
24 jam 57,73 75,84 96,08 92,83 93,25
Produksi VFA rataan hasil penelitian ini berkisar 55,25–99,20 mM.
Suryapratama (1999) menyatakan bahwa kisaran konsentrasi VFA total yang layak
bagi kelangsungan hidup ternak adalah 80–160 mM, dengan titik optimum 110 mM.
Secara umum konsentrasi VFA dari berbagai jenis legum pohon tidak
menunjukkan adanya perbedaan (P>0,05). Banyaknya VFA yang dihasilkan didalam
rumen sangat bervariasi tergantung pada jenis ransum yang dikonsumsi (McDonald
et al., 1988). Konsentrasi VFA pada angsana, turi dan gamal selama inkubasi 12 jam
lebih tinggi dari lamtoro dan kaliandra. Akan tetapi, setelah inkubasi 24 jam
konsentrasi VFA dari daun turi mengalami penurunan bertolak belakang dengan
konsentrasi gamal, lamtoro dan kaliandra yang semakin meningkat.
Rendahnya konsentrasi VFA pada lamtoro dan kaliandra selama inkubasi 12
jam pertama disebabkan oleh aktivitas mikroba rumen. Konsentrasi VFA meningkat
setelah inkubasi selama 24 jam mengindikasikan bahwa mikroba rumen
membutuhkan waktu yang lebih lama dalam mencerna lamtoro dan kaliandra.
Lamtoro dan kaliandra mengandung anti nutrisi tannin sehingga daya cernanya
rendah.
Produksi Amonia (NH3)
Amonia (NH3) merupakan salah satu hasil perombakan protein oleh mikroba
rumen. Konsentrasi NH3 cairan rumen akan meningkat jika populasi protozoa
meningkat, karena protozoa ikut berperan dalam proses daur ulang nitrogen (Sewed,
Tabel 8. Konsentrasi NH3 Beberapa Jenis Legum Pohon
Produksi NH3 (mM)
Legum
Angsana Turi Gamal Lamtoro Kaliandra
12 jam 12,28B 28,83A 8,30C 6,30C 2,51C
24 jam 19,93B 39,55A 13,33C 11,30D 3,49E
Keterangan: superkrip yang berbeda pada baris yang sama menunjukkan berbeda sangat nyata (P<0,01)
Konsentrasi NH3 yang ditunjukkan beberapa jenis legum pohon diatas cukup
tinggi. Konsentrasi optimal NH3 untuk menunjang pertumbuhan mikroba rumen
berkisar antara 85–300 mg/l atau 6–21 mM (McDonald, 1995), dengan titik
optimum 8 mM (Suryapratama, 1999). Konsentrasi NH3 pada lamtoro, kaliandra dan
gamal yang diinkubasi selama 12 jam nyata lebih rendah dibandingkan legum yang
lainnya. Legum yang memiliki konsentrasi NH3 yang paling tinggi baik pada
inkubasi selama 12 jam maupun 24 jam adalah turi. Hal ini karena kandungan
protein kasar pada turi lebih tinggi dibanding legum yang lain, sedangkan lamtoro
mempunyai nilai kecernaan yang rendah meskipun protein kasarnya tinggi. Selain itu
kaliandra memiliki zat anti nutrisi tanin yang menghambat kerja mikroba rumen
dalam mencerna pakan. Produksi NH3 tergantung dari kelarutan protein ransum,
jumlah protein ransum, lamanya pakan dalam rumen dan pH rumen (Orskov, 1982).
Hubungan Laju Degradasi Bahan Kering dengan Produksi VFA dan NH3
Hubungan laju degradasi dengan produksi VFA dan NH3 beberapa legum
pohon dapat dilihat pada Tabel 9.
Tabel 9. Hubungan Laju Degradasi Bahan Kering dengan Produksi VFA dan NH3
Hasil penelitian menunjukkan bahwa terdapat hubungan yang sangat erat
antara laju degradasi (c) dengan konsentrasi NH3. Semakin tinggi laju degradasi
terbalik dengan konsentrasi VFA dalam rumen. Hal ini karena legum yang
digunakan dalam percobaan ini mengandung protein yang tinggi. Pakan yang
mengandung protein tinggi dan kandungan serat kasar yang relatif rendah jika
dibandingkan umumnya hijauan pakan, sehingga produksi NH3 tinggi, tetapi
KESIMPULAN DAN SARAN
Kesimpulan
Legum pohon memiliki potensi degradasi dan laju degradasi pakan yang
berbeda. Legum turi memiliki potensi degradasi dan laju degradasi paling tinggi.
Nilai degradasi diurutkan dari tertinggi sampai terendah selama inkubasi 6 hingga 48
jam adalah turi > angsana dan gamal > lamtoro > kaliandra, sedangkan nilai potensi
degradasi diurutkan dari yang tertinggi hingga terendah adalah turi dan gamal >
angsana dan lamtoro > kaliandra.
Perbedaan potensi dan laju degradasi mempengaruhi kelarutan Ca dan P
pada masing–masing legum tersebut. Kelarutan Ca dan P dapat terlihat setelah
inkubasi 24 jam. Kelarutan Ca pada angsana, turi dan gamal lebih tinggi
dibandingkan lamtoro dan kaliandra. Sedangkan kelarutan P tertinggi pada legum
gamal. Kelarutan mineral P setelah inkubasi 24 jam diurutkan dari yang paling tinggi
sampai terendah berturut-turut adalah daun gamal > turi > angsana dan lamtoro >
kaliandra. Terdapat hubungan yang erat antara fermentabilitas pakan dengan
kelarutan mineral.
Saran
Untuk menilai kualitas mineral kalsium (Ca) dan fosfor (P) legum pohon
perlu dilakukan penelitian terhadap kelarutan mineral kalsium (Ca) dan fosfor (P)
UCAPAN TERIMAKASIH
Assalamualaikum Wr. Wb.,
Puji syukur penulis panjatkan kehadirat Allah SWT yang telah melimpahkan
rahmat dan hidayah-Nya, sehingga penulis dapat menyelesaikan skripsi yang diberi
judul Kelarutan Mineral Kalsium (Ca) dan Fosfor (P) Beberapa Jenis Legum
Pohon Secara in Vitro.
Rasa terima kasih yang tulus penulis hanturkan kepada Dr. Ir. Idat G.
Permana MSc.Agr. dan Dr. Ir. Luki Abdullah MSc.Agr. selaku dosen pembimbing,
yang telah memberikan pengarahan dan bimbingan selama penulis melakukan
penelitian hingga penulisan skripsi ini. Juga kepada Ibu dan Bapak yang senantiasa
berdoa, mencurahkan kasih sayang serta memberikan semangat baru bagi penulis.
Ucapan terimakasih juga penulis sampaikan pada Project DUE Like yang
telah memberikan biaya penuh selama penelitian. Kepada teman satu penelitian,
Nunik Puji Haryati, terima kasih atas kerja sama dan bantuannya kepada Penulis.
Juga kepada Ir. Anita S. T. M.RurSc. sebagai dosen penguji seminar, Ir. Ignatius
Kismono MS, dan Ir. Sri Rahayu Msi. sebagai dosen penguji sidang, Dr. Ir. Yuli
Retnani MSc. yang telah memberikan dukungan moral, adikku tersayang (Mamat,
Syaflan dan Jhon), dan keluarga besar di Madura, keluarga besar Bapak Bahfid dan
Bapak Abd. Salam di Madura terima kasih untuk semua perhatian dan kasih
sayangnya.
Kepada Bu Dian, Pak Adi, Pak Iman dan teman-teman yang telah membantu
(Martha, Niken, Edo, Murni, M’Anis, M’Tien, Mamak dan Yudi dan Adik Wulan),
semua INMT’38, Keluarga Mahasiswa Madura, teman-teman Senior Residence,
teman–teman DKM Al Hurriyyah, Adik–adik angkatan 41 dan 42, terimakasih atas
doa, kasih sayang dan dorongan semangatnya. Kepada semua pihak yang tidak bisa
penulis sebutkan satu persatu, terima atas semua bantuan, kritik dan sarannya.
Akhirnya, penulis berharap semoga tulisan ini bisa bermanfaat bagi yang
membutuhkan. Amin.
Wassallamualaikum Wr. Wb.
Bogor, Februari 2006
DAFTAR PUSTAKA
Allen, O.N and E. K. Allen. 1981. The Leguminosae, a source Book of Characteristic, Uses and Nodulation. The University of Wisconsin Press. Wisconsin.
AFRC. 1991. A reapraisal of the calcium and phosphorus requirement of sheep and cattle. Technical Committee on Responses to Nutrient. Report No 6 Nutr. Abstract. Rev. 61 : 571 – 613.
Amin, M. 1997. Pengaruh penggunaan probiotik Saccharomyces cerevisiae dan
Aspergillus niger dalam ransum pada populasi mikroba, aktivitas fermentasi rumen, kecernaan dan pertumbuhan sapi perah dara. Tesis. Pascasarjana. Institut Pertanian Bogor. Bogor.
Arora, S.P. 1989. Pencernaan Mikroba pada Ruminansia. Cetakan Pertama. Gadjah Mada University Press, Yogyakarta.
Bamualim, A. 2004. Strategi pengembangan peternakan pada lahan kering. Makalah. Seminar Nasional Pengembangan Peternakan Berwawasan Lingkungan. Bogor, 15 Januari 2004.
Benerjee, G. C. 1978. Animal Nutrition. Oxford and IBM Pub. Co. New Delhi. 232 – 249.
Chase, E.L and C.J. Sniffen. 1989. Mineral in dairy cattle nutrition. International Seminar on Holstein Frisien. PPSKI–USHA and Padjadjaran University. Bandung.
Cheeke, P. R. and L. R. Shull. 1985. Natural Toxicant in Feeds and Poisonous Plants. Avi Publishing Company. Inc. New York.
Cheng, K. J., J. P. Fay, R. E. Howarth, and J. W. Costerton. 1980. Sequence of events in the digestion of fresh legumes leaves by rumen bacteria. Applied Environment Microbiology 40: 613 – 625.
Demeyer, D. I. 1981. Rumen Microbes and Digestion of Plant Cell Wall Agriculture and Environment. Elsevier Sci. Pub. Co, Amsterdam.
Durand, M. and R. Kawashima. 1980. Influence of mineral in rumen microbial digestion. In: Y. Ruckebusch and P. Thivend (Eds.). Digestive Physiology and Metabolism in Ruminants. MTP Ltd. Lancaster. 275–408.
Fleming,G.A. 1973. Mineral composition of herbage. In:Butler, G.W. and R.W. Bailey, (Eds) Chemistry and Biochemistry of Herbage. Academic Press, London, 529-563.
Kandylis, K. 1984. Toxicology of Sulfur in Ruminants. Review. J. Dairy Sci. 66 : 2263.
Kareem, S. and E.D. Sudararaj. 1967. Why sesbania makes nutritious cattle feed. In: Skerman, P.J. (Eds). 1977. Tropical Forage Legume. Food Agriculture Organisation of The United Nations. Rome.
Kawashima, R., L.A. Sofyan, N. Ishida, J. Jachja, K. Katoh, H. Kumagai and H. Yano. 1989. Study on Mineral in Java island in Indonesia. Kyoto University and IPB Project. Bogor.
Keir, Brenda., Nguyen Van Lai., T. R. Preston and E. R. Orskov. 1997. Nutritive value of leaves from tropical trees and shrubs: 1. In vitro gas production and in sacco rumen degradability. Livestock Research for Rural Development. 9. 4.
Khamseekhiew, B., J. B. Liang, C. C. Wong, and Z. A. Jalan. 2001. Ruminal and intestinal digestibility of some tropical legume forages. AJAS. 14. : 321 – 325.
Leng, R.A. 1997. Tree Foliage In Ruminant Nutrition. Food and Agricultural Organization of The United Nations. Rome.
Makkar, H. P. S.1993. Antinutritional factors in animals feedstuffs mode of actions. Int. J. Anim. Sci. 6:88-94.
Matsumoto, H., E. G. Smith and G. D. Sherman.1951. The effect of elevated temperature on the mimosin content of kao haole (Leucaena glauca). Arch. Bioche. Biophys. 33:201-202.
McDonald, P., R.A. Edwards, J.F.D. Greenhalg, and C.A. Morgan. 1988. Animal Nutrition. 4th Edition. Longman Scientific and Technical. New York
McDonald, P., R.A. Edwards, and J.F.D. Greenhalg, and C. A. Morgan. 1995. Animal Nutrition. 5th Ed. Longman Scientific and Technical, New York.
McDowell, L., and R., G. Valle. 2000. Major mineral in forage. In: D. I. Given, E. owen, R. F. E. Axford and H. M. Omed (Eds) Forage Evaluation in Ruminant Nutrition. CABI Publishing. UK. London.
McLeod. N. 1974. Plant Tannins. Their Role in Forage and Quality Nutrition. Abs and Rev. 44 : 804.
McManus, W.R., R.G. Antony, L.L. Grout, AS. Malin and V.N.E. Robinson. 1979. Biocristallization of mineral material on forage plant cell walls. Aust. J. Agric. Res. 30 : 635–649.
Ogimoto, K. and S. Imai. 1985. Atlas of Rumen Microbiologiy. Japan Science Societies Press, Tokyo.
Ørskov, E.R.1982. Protein Nutrition in Ruminants. Academic Press, London.
Ǿrskov, E. R., and I. McDonald. 1979. The estimation of protein degradability in rumen from incubation measurements weighed according to rate of passage. J. Agri. Sci. 1979.:499-503.
Parakkasi, A. 1999. Ilmu Nutrisi dan Makanan Ternak Ruminan. UI-Press. Jakarta.
Piliang, W. G. 2001. Nutrisi Mineral. Edisi ke-4. Pusat Antar Universitas Ilmu Hayat. Institut Pertanian Bogor. Bogor.
Prince, M. L., A. E. Hagerman and L. G. Butler.1980. Tannin content of cowpeas, chispeas, Pegeonpeas and Mung beans. J. Agric. Food. Chem. 28:459-461.
Rosa, K. R. D. 1998. Nitrogen fixing tress as tool soil builders. FACT. www.winrock.org/forestry/factnet.htm. [5 Oktober 2005]
Sewet, U.1997. Dinamika Populasi dan Aktivitas Fermentasi Mikriba Rumen
Kambing yang Diberi Pakan Kaliandra (Calliandra calothyrsus). Tesis.
Program Pasca Sarjana. Institut Pertanian Bogor. Bogor.
Serra, S.D., A.B. Serra, T. Ichinohe and T. Fujuhara. 1996. Ruminal sulubilization of macrominerals in selected Philippine forages. AJAS. 9 : 75 – 81.
Skerman, P.J. 1977. Tropical Forage Legume. Food Agriculture Organisation of The United Nations. Rome.
Steel, R. G. D. and J. H. Torrie. 1981. Principles and Procedures of Statistic. Mc Grow Hill Book Co. Inc.New York.
Suryapratama, W. 1999. Efek suplementasi asam lemak volatil bercabang dan kapsul lisin serta treonin terhadap nutrisi protein sapi Holstein. Disertasi. Program Pascasarjana. Institut Pertanian Bogor, Bogor.
Sutardi, T. 1977. Ikhtisar Ruminologi. Bahan Penataran Kasus Peternakan Sapi Perah di Kayu Ambon, Lembang. Bandung.
Sutardi, T. 1979. Ketahanan protein bahan makanan terhadap degradasi oleh mikroba dan populasi protozoa rumen dan pemanfaatannya bagi produktivitas ternak. Proseding Seminar Penelitian dan Pengembangan Peternakan. Lembaga Penelitian dan Pengembangan Pertanian. Bogor.
Sutardi, T. 1980. Landasan Ilmu Nutrisi. Departemen Ilmu Nutrisi Makanan dan Ternak. Diktat Kuliah. Fakultas Peternakan. Institut Pertanian Bogor, Bogor.
Sutardi, T. , N. A. Sigit dan T. Toharmat. 1983. Standarisasi mutu protein bahan makanan ruminansia berdasarkan parameter metabolismenya oleh mikroba Rumen. Laporan Penelitian. Fakultas Peternakan. Institut Pertanian Bogor, Bogor.
Sutardi, T., D. Sastradipdradja, T. Toharmat, A. Sardiana dan I.G. Permana. 1994. Peningkatan produksi ternak ruminansia melalui amoniasi pakan serat bermutu rendah, defaunasi dan suplementasi protein tahan degradasi dalam rumen. Laporan Penelitian. Fakultas Peternakan IPB. Bogor.
Suttie, J.M. 1979. Leucaena leucocephala (Lam.) de Wit.
www.Fao.Org./ag/AGP/AGPC/doc./Gbase/DATA/pf000158.htm. [5 Oktober 2005].
Tilley, J. M. A. and R. A. Terry. 1963. A two stage technique for the In vitro
digestion of forage crops. J. British Grassland Society 18, 104 – 111.
Underwood, E.J. 1981. The Mineral Nutrition of Livestock. Common Wealth Agricultural Bereaux. London.
Underwood, E. J. and N. F. Suttle. 1999. The Mineral Nutrition of Livestock. 3rd Edition. CABI Publishing. London.
Ward, G. and L.H. Harbers. 1982. Effect of pH on extractibility of calcium and oxalate from alfafa leafs. J. Dairy Sci. 65 : 154 – 160.
Widiyanto. 1992. Kemungkinan penggunaan tepung daun angsana dalam ransum kelinci ditinjau dari penampilan ternak dan produksi karkas. Karya Ilmiah. Fakultas Peternakan. Institut Pertanian Bogor. Bogor.