Embriogenesis Somatik Tidak Langsung Jeruk Keprok Batu 55 (Citrus reticulata L) Hasil Perlakuan Kolkisin
Bebas
47
0
0
Teks penuh
Gambar


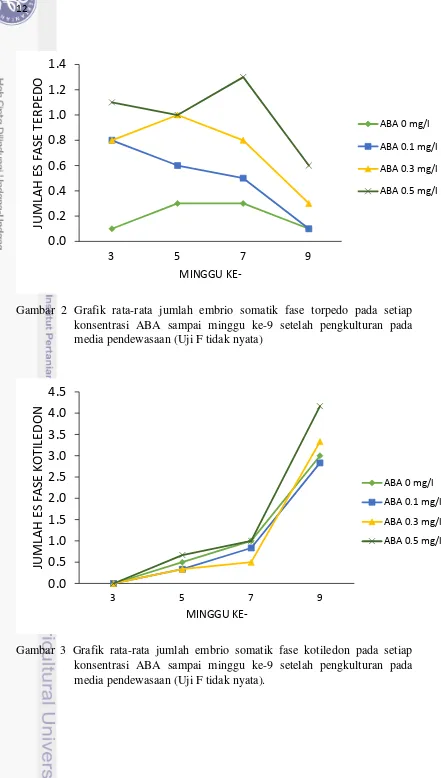
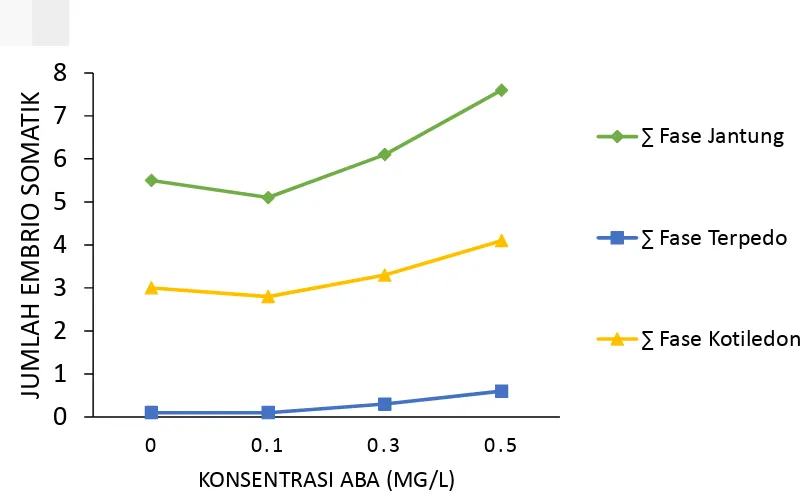
+7

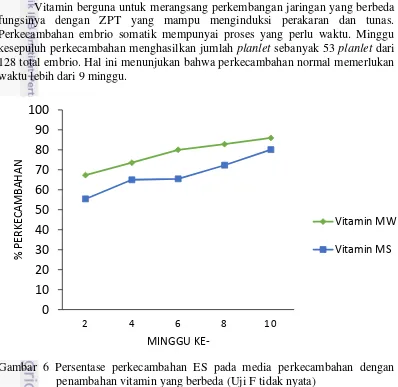
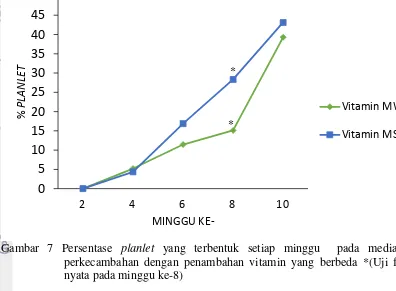


Dokumen terkait
