PERILAKU, POLA PELETAKAN TELUR DAN EFISIENSI
PEMARASITAN PARASITOID Trichogramma chilotraeae
NAGARAJA & NAGARKATTI (HYMENOPTERA:
TRICHOGRAMMATIDAE) PADA BERBAGAI
JUMLAH INANG DAN KEPADATAN PARASITOID
HASRIYANTY
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
1
ABSTRAK
HASRIYANTY. Perilaku, Pola Peletakan Telur dan Efisiensi Pemarasitan Parasitoid Trichogramma chilotraeae Nagaraja & Nagarkatti (Hymenoptera: Trichogrammatidae) Pada Berbagai Jumlah Inang dan Kepadatan Parasitoid. Dibimbing oleh DAMAYANTI BUCHORI dan PUDJIANTO.
Sebagian besar parasitoid Hymenoptera, termasuk parasitoid
Trichogramma chilotraeae merupakan organisme yang mempunyai pola
reproduksi haplodiploidi. Penentuan kelamin keturunan sepanjang peletakan telur ditentukan oleh induk betina berdasarkan keadaan lingkungan, dalam hal ini jumlah inang dan kepadatan parasitoid. Jumlah inang dan kepadatan parasitoid tersebut dalam perbanyakan massal juga dapat mempengaruhi berbagai ciri kebugaran sekaligus menentukan efisiensi pemarasitan. Perilaku dan pola urutan peletakan telur penting dikaji untuk lebih memahami perilaku parasitoid untuk tujuan memaksimalkan keberhasilan reproduksi dan keberlangsungan keturunannya nanti, yang secara langsung memberikan arti penting untuk memperbaiki keefektifan pengendalian hama sasaran di lapangan dalam suatu program pengendalian hayati. Selain itu kajian berbagai ciri kebugaran serta efisiensi pemarasitan penting dilakuan dalam kaitannya dengan kegiatan perbanyakan massal di laboratorium. Dalam penelitian ini dipelajari perilaku reproduksi melalui pengamatan urutan peletakan kelamin keturunan pada perlakuan jumlah inang 3, 6, 12, 24 dan 48 dan dipaparkan pada 1 dan 2 parasitoid; berbagai ciri kebugaran parasitoid T. chilotraeae pada jumlah inang 3, 6, 12, 24 dan 48; serta pengaruh kepadatan parasitoid, yakni kepadatan 1, 2, 3 dan 4 parasitoid betina terhadap nisbah kelamin keturunan.
Jumlah inang berpengaruh terhadap perilaku dan pola peletakan telur. Pada umumnya parasitoid T. chilotraeae memulai rangkaian peletakan telur dengan menghasilkan keturunan betina. Faktor jumlah inang dan kepadatan parasitoid betina berpengaruh pada perilaku selfsuperparasitism. Jumlah inang juga berpengaruh terhadap berbagai ciri kebugaran dan efisie nsi pemarasitan. Rasio 24 inang dengan 1 parasitoid betina lebih efisien dibandingkan rasio 48 inang dengan 1 parasitoid, dengan pemarasitan paling efektif terjadi pada tiga jam pertama hari pertama. Meningkatnya kepadatan parasitoid betina berpengaruh pada nisbah kelamin (proporsi betina). Nisbah kelamin semakin menurun seiring bertambahnya kepadatan parasitoid.
2
PERILAKU, POLA PELETAKAN TELUR DAN EFISIENSI
PEMARASITAN PARASITOID Trichogramma chilotraeae
NAGARAJA & NAGARKATTI (HYMENOPTERA:
TRICHOGRAMMATIDAE) PADA BERBAGAI
JUMLAH INANG DAN KEPADATAN PARASITOID
HASRIYANTY
T e s i s
Sebagai salah satu syarat untuk memperoleh gelar
Magister Sains pada Program Studi Entomologi dan Fitopatologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
3
PRAKATA
Puji syukur dipanjatkan kehadirat Allah SWT, atas limpahan rahmat dan karuniaNya sehingga penulis dapat menyelesaikan penelitian dan penulisan tesis yang berjudul “Perilaku, Pola Peletakan Telur dan Efisiensi Pemarasit an Parasitoid Trichogramma chilotraeae Nagaraja & Nagarkatti (Hymenoptera: Trichogrammatidae) pada Berbagai Jumlah Inang dan Kepadatan Parasitoid”, dilaksanakan di Laboratorium Bioekologi Parasitoid dan Predator, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanaian Bogor pada bulan Juni 2005 hingga Maret 2006.
Terima kasih penulis ucapkan kepada ibu Dr. Ir. Damayanti Buchori, M.Sc. selaku ketua komisi pembimbing dan bapak Dr. Ir. Pudjianto, M.Si. sebagai anggota atas segala bimbingan dan petunjuknya selama penelitian dan penulisan tesis. Terima kasih juga disampaikan kepada Ketua Program Studi serta seluruh staf pengajar Pascasarjana Program Studi Entomologi Fitopatologi yang telah memberikan pengetahuan yang tak ternilai harganya selama masa perkuliahan. Disamping itu, ungkapan terima kasih juga disampaikan kepada Rektor dan Dekan Sekolah Pascasarjana IPB, Rektor serta Dekan Fakultas Pertanian Universitas Tadulako yang telah memberikan kesempatan kepada penulis untuk melanjutkan pendidikan program Magister Sains pada Sekolah Pascasarjana IPB. Demikian pula pada Pengelola Biaya Pendidikan Program Pascasarjana (BPPS) Dikti atas dukungan biaya selama masa studi.
Penghargaan dan ungkapan terima kasih kembali penulis sampaikan kepada ibu Dr. Ir. Damayanti Buchori, M.Sc. selaku penanggungjawab Laboratorium Bioekologi Parasitoid dan Predator atas perizinan untuk menggunakan semua bahan dan peralatan penelitian di laboratorium sehingga penelitian ini dapat terlaksana dengan baik. Terima kasih juga kepada Adha Sari, Bandung Sahari dan Akhmad Rizali atas diskusi dan masukan-masukannya serta kepada seluruh anggota Laboratorium Bioekologi Parasitoid dan Predator untuk segala pertemanan dan menciptakan suasana kekeluargaan di laboratorium, juga kepada mahasiswa pascasarjana Entomologi Fitopatologi angkatan 2004 terima kasih untuk persahabatan, bantuan serta dukungan dalam menyelesaikan studi.
Pada akhirnya, karya ini penulis persembahkan buat ayahanda dan ibunda tercinta karena dengan doa dan keridhaan semua ini dapat berjalan lancar, serta kepada suami terkasih Andi Amran Mappasittung, kedua anakku Andi Aura Amaliyah dan Andi Azra Amaliyah atas keikhlasan, pengertian, doa dan dorongan serta semangat untuk pencapaian ini.
Semoga karya ilmiah ini bermanfaat bagi kita semua.
Bogor, 25 Agustus 2006
4
Judul tesis : Perilaku, Pola Peletakan Telur dan Efisiensi Pemarasitan Parasitoid Trichogramma chilotraeae Nagaraja & Nagarkatti (Hymenoptera: Trichogrammatidae) pada Berbagai Jumlah Inang dan Kepadatan Parasitoid
Nama : Hasriyanty
NIM : A451040011
Disetujui
Komisi Pembimbing
Dr. Ir. Damayanti Buchori, M.Sc. Dr. Ir. Pudjianto, M.Si. Ketua Anggota
Diketahui
Ketua Program Studi Entomologi dan Fitopatologi
Dr. Ir. Sri Hendrastuti Hidayat, M.Sc.
Dekan Sekolah Pascasarjana
Dr. Ir. Khairil Anwar Notodiputro, MS.
5
RIWAYAT HIDUP
Penulis dilahirkan di Soni, Kecamatan Dampal Selatan, Kabupaten Tolitoli Sulawesi Tengah pada tanggal 27 Oktober 1972 dari Ayah Hi. Latjella Mannessa dan Ibu Hj. Djuhrana. Penulis merupakan anak kedua dari tiga bersaudara.
16
SURAT PERNYATAAN
Dengan ini saya menyatakan bahwa tesis yang berjudul :
Perilaku, Pola Peletakan Telur dan Efisie nsi Pemarasitan Parasitoid
Trichogramma chilotraeae Nagaraja & Nagarkatti (Hymenoptera: Trichogrammatidae) pada Berbagai Jumlah Inang dan Kepadatan Parasitoid
adalah benar karya saya sendiri dan belum pernah diajukan dalam bentuk apapun kepada Perguruan Tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun yang tidak diterbitkan dari penulis lain telah disebutkan dalam tesis dan dicantumkan dalam Daftar Pustaka di bagian akhir
tesis ini.
Bogor, 25 Agustus 2006
17
© Hak cipta milik Institut Pertanian Bogor, tahun 2006 Hak cipta dilindungi:
Dilarang mengutip dan memperbanyak tanpa izin tertulis dari Institut Pertanian Bogor, sebagian atau seluruhnya dalam bentuk apapun,
6
DAFTAR ISI
Halaman
DAFTAR TABEL ... xi
DAFTAR GAMBAR ... xii
PENDAHULUAN ... ... 1
Latar Belakang ... 1
Tujuan Penelitian ... ... 4
Manfaat Penelitian ... ... 4
TINJAUAN PUSTAKA ... ... 5
Parasitoid Telur Trichogrammatidae dalam Pengendalian Hayati ... 5
Pengendalian Hayati Pada Hama Plutella xylostella ... 7
Trichogramma chilotraeae Nagaraja & Nagarkatti dan Sebarannya ... 9
Alokasi Jenis Kelamin Keturunan pada Trichogrammatidae ... 10
BAHAN DAN METODE ... 15
Waktu dan Tempat ... 15
Persiapan Penelitian ... 15
Koleksi dan Perbanyakan Parasitoid ... 15
Perbanyakan serangga inang Corcyra cephalonica ... 15
Pelaksanaan Penelitian ... 16
Identifikasi Parasitoid ... 16
Pola Urutan Peletakan Telur ... 17
Studi Pengaruh Jumlah Inang terhadap Efisiensi Pemarasitan .. 18
Studi Waktu Pemarasitan Paling Efektif ... 19
Studi Pengaruh Jumlah Betina terhadap Nisbah Kelamin ... 20
HASIL DAN PEMBAHASAN ... 21
Identifikasi Parasitoid ... 21
Perilaku Penemuan dan Penerimaan Inang ... 23
Pola Urutan Peletakan Telur Parasitoid T. chilotraeae ... 25
7
Parasitisasi ... 34
Persentase Selfsuperparasitism ... 37
Efisiensi Pemarasitan Berbagai Jumlah Inang ... 41
Waktu Pemarasitan Paling Efektif ... 45
Pengaruh Kepadatan Betina Terhadap Nisbah Kelamin Keturunan 46 KESIMPULAN DAN SARAN ... 50
Kesimpulan ... 50
Saran ... 50
8
DAFTAR TABEL
Halaman
1 Alokasi jenis kelamin keturunan yang dihasilkan induk parasitoid
T. chilotraeae pada peletakan telur ke-1 sampai peletakan telur
ke-n... 28 2 Frekuensi berbagai variasi pola urutan peletakan tiga telur
pertama, parasitoid T. chilotraeae ... 32 3 Rata-rata nisbah kelamin (proporsi betina) pada berbagai
kepadatan inang dan kepadatan parasitoid ... 34 4 Total inang terparasit, persentase inang terparasit, persentase
keturunan betina dan persentase keturunan jantan pada berbagai
jumlah inang... 35 5 Persentase inang terparasit, persentase keturunan betina dan
persentase keturunan jantan pada berbagai kepadatan parasitoid
inang ... 36 6 Persentase selfsuperparasitism pada berbagai jumlah inang... 37 7 Persentase selfsuperparasitism pada berbagai kepadatan
parasitoid ...
38 8 Frekuensi berbagai variasi selfsuperparasitism pada peletakan
tiga telur pertama parasitoid T. chilotraeae pada berbagai jumlah
inang ... 40 9 Total inang terparasit, rata-rata inang terparasit per hari,
persentase kemunculan imago dan lama hidup parasitoid T.
9
DAFTAR GAMBAR
Halaman
1 Karakter morfologi T. chilotraeae hasil identifikasi; antena,
sayap dan genitalia imago jantan... 22 2 Karakter morfologi T. chilotraeae berdasarkan kunci identifikasi;
antena, sayap dan genitalia imago jantan..... 22 3 Persentase keturunan betina yang dihasilkan pada setiap urutan
peletakan telur pada berbagai jumlah inang, pemaparan satu
parasitoid betina ... 26 4 Persentase keturunan betina yang diletakkan pada setiap urutan
peletakan telur pada berbagai jumlah inang, pemaparan dua
parasitoid betina ... 30 5 Nisbah kelamin keturunan dari satu induk betina yang
meletakkan telur pada berbagai kepadatan inang ... 33 6 Nisbah kelamin keturunan dari dua induk betina yang
meletakkan telur pada berbagai kepadatan inang ... 33 7 Persentase selfsuperparasitism sepanjang peletakan telur,
pemaparan satu parasitoid betina ... 39 8 Persentase selfsuperparasitism sepanjang peletakan telur,
pemaparan dua parasitoid betina ... 39 9 Grafik jumlah inang terparasit perhari ... 42 10 Nisbah kelamin total satu parasitoid betina pada berbagai jumlah
inang ... 43 11 Sebaran data nisbah kelamin parasitoid betina pada berbagai
jumlah inang ... 44 12 Proporsi keturunan betina pada setiap hari pengamatan selama
masa hidup induk betina ... 45 13 Grafik jumlah inang terparasit pada setiap 3 jam pemaparan ... 45 14 Nisbah kelamin parasitoid T. chilotraeae pada berbagai
18
PENDAHULUAN
Latar Belakang
Hama ulat daun kubis Plutella xylostella Lin. (Lepidoptera: Yponomeutidae) merupakan salah satu hama penting pada pertanaman kubis yang dapat menimbulkan kerusakan dan bisa mengakibatkan kegagalan panen. Selama ini, ditingkat petani pengendalian hama ini biasanya menggunakan insektisida sintetik, namun dari beberapa penelitian membuktikan bahwa hama ini mampu mengembangkan sistem kekebalan sehingga menjadi resisten terhadap insektisida. Bila hal ini terjadi maka biasanya petani akan meningkatkan dosis atau mencampur berbagai jenis insektisida (Kfir 2002). Sebagai contoh, hasil penelitian Rauf et al. (2005) melaporkan bahwa petani di tiga lokasi sentra produksi kubis di pulau Jawa menggunakan 35 produk insektisida untuk mengendalikan hama P. xylostella dan Crocidolomia vaponana (Fabricus) (Lepidoptera: Noctuidae). Penggunaan insektisida yang berlebihan dan dengan sifat insektisida yang berspektrum luas akan ikut mematikan berbagai jenis musuh alami hama ini di lapangan. Sebagai akibatnya, bila tidak ada faktor alami yang menekan maka populasi hama akan semakin tinggi dan lebih sulit untuk dikendalikan (Kfir 2002).
P. xylostella dilaporkan merupakan serangga hama yang mempunyai
kemampuan untuk mengembangkan sifat resisten terhadap berbagai jenis insektisida. Penggunaan DDT secara luas pada tahun 1940-an, telah mengakibatkan hama ini dengan cepat menjadi resisten terhadap bahan aktif insektisida tersebut. Selanjutnya dilaporkan bahwa P. xylostella resisten terhadap
diclorvos, cartap, methomiyl, chlorpyrifosmethyl dan malathion serta berbagai bahan aktif insektisida lainnya (Kfir 2002). Hama ini bahkan merupakan serangga hama pertama yang dilaporkan resisten terhadap bahan aktif insektisida mikroba
Bacillus thuringiensis Berl. (Tabone et al. 2003; Tabashnik et al. 1990 dalam
19
Pengendalian hayati menggunakan parasitoid merupakan teknik pengendalian yang bisa mengatasi permasalahan serangga hama karena parasitoid pada umumnya mempunyai kemampuan untuk memarasit ratusan inang sehingga dapat dengan cepat menekan populasi hama. Parasitoid juga mempunyai sifat selektif sehingga tidak menimbulkan dampak pada musuh alami lainnya dan tidak menimbulkan resistensi pada hama sasaran (Nofemela 2004).
Di Indonesia, parasitoid P. xylostella yang selama ini dilaporkan hanya ada satu spesies yakni Diadegma semiclausum Hellen (Hymenoptera: Ichneumonidae) yang merupakan parasitoid larva (Kalshoven 1981). Tetapi dari hasil penelitian Meilin (1999) melaporkan, dari hasil eksplorasi di beberapa daerah di wilayah Jawa, ternyata didapatkan tiga spesies parasitoid telur dari famili Trichogrammatidae yang menyerang telur P. xylostella yakni
Trichogrammatoidea cojuangcoi Nagaraja, Trichogrammatoidea armigera
Nagaraja dan Trichogramma flandersi Nagaraja & Nagarkatti. Dari eksplorasi yang dilakukan di daerah Pangalengan Bandung pada penelitian ini, ternyata diperoleh satu spesies parasitoid Trichogrammatidae lainnya yang menyerang telur P. xylostella dan diidentifikasi sebagai Trichogramma chilotraeae Nagaraja & Nagarkatti. Pemanfaatan parasitoid telur Trichogrammatidae ini dapat menjadi pilihan karena dapat mengendalikan hama pada fase paling awal sehingga kerusakan tanaman dapat dicegah sedini mungkin. Parasitoid ini juga dapat dikembangbiakkan secara massal di laboratorium dengan menggunakan inang pengganti Corcyra cephalonica Stointon (Lepidoptera: Pyralidae).
Berkenaan dengan tujuan untuk memanfaatkan parasitoid T. chilotraeae ini, banyak aspek mendasar yang perlu dikaji baik dalam kaitannya dengan kegiatan pembiakan massal, maupun untuk mengetahui berbagai karakter yang berhubungan dengan kemampuan parasitoid ini untuk dapat berperan di lapangan nantinya. Aspek tentang hubungan inang-parasitoid, kajian biologi reproduksi untuk pengembangan metode pembiakan massal perlu diperhatikan untuk mendapatkan parasitoid yang berkualitas tinggi, demikian pula aspek ekologi dan perilaku reproduksinya.
20
berasal dari populai yang berbeda memiliki ciri-ciri biologi yang berbeda. Faktor lingkungan, dalam hal ini kepadatan inang dan kepadatan parasitoid (foundres), juga berpengaruh pada biologi dan perilaku reproduksi yang nantinya akan menentukan kualitas parasitoid yang dihasilkan. Hasil penelitian Khan et al. (2004) melaporkan bahwa parasitoid Trichogramma chilonis Ishii dapat merespon kondisi kepadatan inang dan kepadatan induk parasitoid dan hal ini dapat berpengaruh pada berbagai indikator kualitas yang diukur.
Jumlah inang dan kepadatan parasitoid akan berpengaruh pada perilaku reproduksi, dalam hal ini pengaturan alokasi kelamin keturunan yang dihasilkan oleh induk betina. Studi tentang pengaturan alokasi kelamin keturunan oleh induk betina sebagai respon terhadap lingkungan dapat dilakukan dengan pengamatan urutan peletakan telur. Pengamatan ini akan memberikan gambaran mengenai perilaku reproduksi parasitoid T. chilotraeae ini karena diketahui bahwa setiap spesies parasitoid Hymenoptera mempunyai strategi peletakan telur yang berbeda. Wanjberg (1993) melaporkan bawa pada parasitoid Trichogramma brassicae
Bezdenco, akan memulai rangkaian peletakan telur dengan meletakkan keturunan jantan pada awal-awal peletakan telur. Demikian pula Colazza dan Wajnberg (1998) melaporkan bahwa urutan peletakan telur parasitoid Trissolcus basalis
(Wollaston) (Hymenoptera: Scelionidae) pada berbagai jumlah inang dalam suatu paket tidak diletakkan secara acak. Peletakan telur pertama pada semua perlakuan berbagai jumlah inang selalu jantan (strategi “jantan pertama”), tetapi bentuk alokasi kelamin keturunan akan berubah sebagai respon terhadap variasi jumlah inang.
Tujuan Penelitian
21
Manfaat Penelitian
22
Pengendalian hayati menggunakan parasitoid telur family Trichogrammatidae untuk mengendalikan berbagai jenis hama, terutama hama dari ordo Lepidoptera telah banyak dilakukan secara rutin di beberapa negara, penggunaan parasitoid Trichogrammatidae ini menjadi pilihan karena dapat mengendalikan hama pada fase paling awal sehingga kerusakan tanaman dapat dicegah sedini mungkin, tidak meninggalkan residu pada produk pertanian dan terutama aman terhadap kesehatan manusia. Dilaporkan bahwa sampai saat ini lebih dari 32 juta hektar lahan pertanian di seluruh dunia menggunakan parasitoid Trichogramma untuk mengendalikan hama tanaman (Li 1994). Di Indonesia, penggunaan parasitoid telur ini telah banyak digunakan dan berhasil mengendalikan hama penggerek tebu, penggerek padi dan penggerek kedelai (Kalshoven 1981), dan saat ini telah banyak dibiakkan secara massal sebagai teknologi alternatif non pestisida.
Nisbah kelamin keturunan parasitoid Hymenoptera diketahui bervariasi antar satu spesies dengan spesies lainnya, bahkan dalam satu spesies akan mempunyai nisbah kelamin keturunan yang berbeda tergantung dari kondisi lingkungan (Godfray 1994). Hal ini dapat terjadi karena diketahui parasitoid Hymeoptera mempunyai pola reproduksi haplodiploidi, dimana jantan haploid berkembang dari telur yang tidak dibuahi dan betina diploid akan berkembang dari telur yang dibuahi, dan penentuan kelamin keturunan dapat diatur oleh induk betina berdasarkan keadaan lingkungan, jadi berdasarkan konsep ini maka nisbah kelamin keturunan dapat menjadi ekstrim bias betina atau bias jantan (Colazza & Wajnberg 1998).
Penelitian ini akan melihat bagaimana respon betina parasitoid T. chilotreae
23
TINJAUAN PUSTAKA
Parasitoid Telur Trichogrammatidae dalam Pengendalian Hayati
Pengendalian hayati merupakan suatu strategi pengendalian hama yang saat ini banyak dikembangkan untuk menggantikan penggunaan pestisida sintetik yang cenderung menimbulkan dampak negatif seperti resistensi dan resurgensi pada hama sasaran, munculnya hama sekunder, pencemaran lingkungan dan pengaruhnya pada kesehatan manusia serta residu pada produk pertanian dan hewan. Istilah pengendalian hayati pertama kali diartikan sebagai penggunaan musuh alami untuk mengendalikan hama. Akan tetapi kemudian berkembang dan diartikan sebagai pengendalian hama tanpa menggunakan senyawa kimia termasuk teknik pengembangan tanaman yang resisten terhadap hama dan penyakit, modifikasi kultur teknis, pengaturan waktu tanam, eradikasi, pelepasan jantan steril atau dengan menggunakan feromon (Wilson & Huffaker 1976). Sedangkan menurut DeBach (1973), pengendalian hayati diartikan sebagai kegiatan parasitoid, predator dan patogen yang memelihara dan menjaga keseimbangan kepadatan polulasi suatu organisme lain pada suatu tingkat populasi rata-rata tanpa pengendalian lain.
Salah satu kelompok musuh alami serangga hama yang banyak dikembangkan adalah parasitoid. Parasitoid umumnya merupakan serangga dari ordo Hymenoptera, tetapi beberapa spesies parasitoid juga dapat ditemukan pada Ordo Diptera, Strepsiptera, Coleoptera dan Lepidoptera (Gord et al. 1999). Salah satu famili dari ordo Hymenoptera yang anggotanya banyak berperan sebagai parasitoid adalah famili Trichogrammatidae.
digabungkan dan kompatibel dengan metode pengendalian lain (Alba 1988)
sehingga dapat mendukung pelaksanaan PHT di lapangan. Parasitoid ini terutama banyak digunakan untuk menekan hama penting dari ordo Lepidoptera. Beberapa spesies Trichogramma juga dapat memarasit telur Coleoptera, Diptera, Heteroptera, Hymenoptera dan Neuroptera (Smith 1996; Knutson 2002).
Dalam Famili Trichogrammatidae terdapat 80 genera dan semuanya diketahui merupakan parasitoid telur (Pinto & Stouthamer 1994). Dua genera yang terkenal adalah Trichogramma dan Trichogrammatoidea (Clausen 1940; Nagarkatti & Nagaraja 1977). Di Indonesia, beberapa parasitoid Trichogrammatidae yang pernah dilaporkan antara lain: Trichogramma japonicum
Ashmead, Trichogrammatoidea nana Zehnt (Kalshoven 1981), Trichogramma
australicum Girault (Nagarkatti & Nagaraja 1977; Kalshoven 1981),
T. chilotraeae, Trichogrammatoidea armigera Nagaraja (Nagarkatti & Nagaraja 1977; Nurindah & Bindra 1989), Trichogrammatoidea guamensis Nagaraja (Nurindah & Bindra 1989), Trichogramma flandersi dan Trichogrammatoidea cojuangcoi Nagaraja (Meilin, 1999).
Sejarah penggunaan Trichogramma untuk mengendalikan berbagai jenis serangga hama telah berlangsung sejak lama. Namun demikian, baru pada tahun 1926 Trichogramma mulai dibiakkan secara massal ketika Flander untuk pertama kalinya mengembangkan sistem pengembangbiakan massal dengan menggunakan
telur inang pengganti Sitotroga cerealella Oliver (Lepidoptera: Gelechiidae) (Li 1994). Selama 20 tahun terakhir parasitoid Trichogramma telah digunakan
secara luas terutama untuk mengendalikan hama-hama pada tanaman pangan, misalnya padi, jagung, gandum dan sorgum; pada tanaman industri seperti tebu, kapas dan kedelai; serta sayuran dan buah-buahan (Li 1994). Di Indonesia, Trichogramma telah digunakan untuk mengendalikan hama pada tanaman kapas, bit gula, anggur, kubis, apel, tomat dan padi (Smith 1996). Beberapa hasil penelitian juga menunjukkan keefektifan penggunaan parasitoid Trichogramma
dan Trichogrammatoidea untuk mengendalikan berbagai jenis hama di lapangan. Penelitian Ramlan (2001) menunjukkan bahwa tingkat parasitisasi T. armigera
rumpun tanaman. Hasil penelitian Herlinda et al. (1997) menunjukkan bahwa
Trichogrammatoidea bactrae bactrae Nagaraja berhasil menurunkan kerusakan
polong akibat serangan hama Etiella zinkenella (Treitschke) (Lepidoptera: Pyralidae) pada tanaman kedelai. Di luar negeri, penggunaan parasitoid Trichogramma ini telah dilakukan secara komersial pada lebih dari 32 juta hektar lahan per tahun di Cina, Eropa, Asia, Amerika Selatan, Australia dan Amerika Serikat (Li 1994). Pada tahun 1972, dilaporkan bahwa di India telah dikembangkan pelepasan T. armigera bersama-sama T. australicum,
Trichogramma achaeae Nagaraja & Nagarkatti dan Telenomus sp. untuk
mengatasi serangan Helicoverpa armigera Hubn. (Lepidoptera: Noctuidae) pada berbagai tanaman yang bermanfaat secara ekonomi.
Pengendalian Hayati pada Hama Plutella xylostella
Beberapa agens hayati dilaporkan dapat digunakan untuk mengendalikan populasi P. xylostella. B. thuringiensis merupakan salah satu mikroba dari
golongan bakteri yang diketahui efektif untuk mengendalikan hama ini.
B. thuringiensis sudah sejak lama diproduksi secara komersial dan dikemas dalam bentuk formulasi yang mudah diaplikasikan di lapangan (Kfir 2002). Di Indonesia, beberapa petani di Cianjur dan Bandung dilaporkan telah menggunakan formulasi bioinsektisida B. thuringiensis ini dalam berbagai merek dagang, antara lain Agrisal WP, Bactospeine WP, Dipel WP dan Thuricide HP (Rauf et al, 2005). Polyhedral, dan granulosis virus juga dilaporkan pernah digunakan untuk mengendalikan hama ini. Cendawan Beauveria brongniartii
(Saccardo) telah digunakan di Prancis dan Beauveria bassaiana (Balsamo) dalam bentuk formulasi komersil Boverin telah digunakan di Amerika (Kfir 2002).
Dari kelompok parasitoid, beberapa spesies parasitoid larva yakni Cotessia
plutella Kurdjumov (Hymenoptera: Braconidae) dan Diadegma mollipla
(Holmgren) (Hymenoptera: Ichneumonidae) telah digunakan secara luas di pertanaman kubis di Afrika Selatan (Nofemela 2004). Di Kanada bagian barat dilaporkan parasitoid Diadegma insulare Cresson (Hymenoptera: Ichneumonidae), Microplitis plutellae Muesbeck (Hymenoptera: Braconidae) dan
2000). Di Indonesia, dilaporkan ada satu spesies parasitoid larva yakni D.
semiclausum (Kalshoven 1981).
Akhir-akhir ini, banyak dikaji penggunaan musuh alami parasitoid telur dari famili Trichogrammatidae yang berpotensi sebagai agens pengendali hayati yang efektif. Parasitoid telur mempunyai keuntungan dibanding dengan parasitoid larva, karena menyerang telur hama, sehingga dapat mengendalikan hama pada fase paling awal sebelum hama merusak tanaman. Di Indonesia, khususnya di pulau Jawa telah dilaporkan ada tiga spesies parasitoid Trichogrammatidae yang diidentifikasi sebagai parasitoid telur P. xylostella yakni T. flandersi, T. cojuangcoi, dan T. armigera (Meilin 1999). Sedangkan di luar negeri, beberapa yang dapat disebutkan antara lain: di Cina Selatan dilaporkan ada 5 spesies Trichogramma dan Trichogrammatoidea yakni Trichogramma confusum
Nagaraja, Trichogrammatoidea bactrae Nagaraja, Trichogramma ostriniae Pang & Chen, Trichogramma raoi Nagaraja dan Trichogramma bilingensis He & Pang (Yurong et al. 2003; Huang et al. 2005), di Australia T. bactrae dan
Trichogramma pretiosum Riley (Liu et al. 2004), di Jepang, parasitoid
Trichogramma chilonis Ishii serta di Filipina Trichogramma evanescens
Westwood dan T. armigera (Alba 1988). Miura (2003) melaporkan bahwa penggunaan T. chilonis mampu mengendalikan P. xylostella dalamrumah kaca dengan tingkat parasitisasi mencapai 80%. Klem et al. (1992) melaporkan terdapat 27 spesies Trichogramma dan Trichogrammatoidea yang memarasit
P. xylostella yang berasal dari USSR, Prancis, Cina, Amerika, Taiwan dan
Trichogramma chilotraeae Nagaraja & Nagarkatti dan Sebarannya
Dalam klasifikasi, Trichogramma chilotraeae termasuk dalam famili Trichogrammatidae, superfamili Chalcidoidea, Ordo Hymenoptera (Borror et al.
1992). Identifikasi Trichogramma dan Trichogrammatoidea sangat sulit dilakukan karena keduanya mempunyai perbedaan morfologis yang sangat kecil, spesies-spesies dari kedua genus tersebut mempunyai sebaran luas dan strain yang bervariasi. Perbedaan kedua genus tersebut dapat dilihat pada karakter sayap depan, antena jantan dan genitalia imago jantan (Nagarkatti & Nagaraja 1977; Alba 1988). Identifikasi spesies di dasarkan pada beberapa karakter dari genitalia jantan yang sangat khas untuk masing- masing spesies. Untuk melakukan identifikasi maka sebelumnya harus dilakukan pembuatan preparat mikroskop.
T. chilotraeae memiliki dorsal expansion gonobase (DEG) kurang lebih
berbentuk segitiga dengan chelat structures (CS) yang besar dan terletak jauh dari ujung gonoforceps (GF) (Alba 1988).
Informasi penyebaran parasitoid T. chilotraeae di Indonesia sampai saat ini belum diketahui secara pasti karena terbatasnya eksplorasi dan penelitian-penelitian yang dilakukan. Untuk pulau Jawa, beberapa penelitian-penelitian menunjukkan bahwa parasitoid T. chilotraeae dijumpai menyerang telur H. armigera pada tanama n jagung (Nurindah & Bindra 1989), Chillo suppressalis (Wlk.) (Lepidoptera: Pyralidae) pada tanaman padi, dan Ostrinia furnacalis (Guenee) (Lepidoptera: Pyralidae) pada tanaman jagung (Kalshoven 1981), tetapi belum pernah di laporkan menyerang telur P. xylostella. Di Malaysia, India, Kambodja, Filipina dan Thailand, parasitoid ini juga dilaporkan menyerang telur hama
Alokasi Jenis Kelamin Keturunan pada Trichogrammatidae
Pada umumnya parasitoid Hymenoptera adalah haplodiploid. Jantan haploid berkembang dari telur yang tidak dibuahi dan betina diploid berkembang dari telur yang dibuahi. Betina yang tidak berkopulasi akan menghasilkan keturunan yang semuanya jantan, sementara betina yang telah berkopulasi akan menyimpan sperma dalam spermateka dan pada saat peletakan telur, betina dapat mengatur alokasi kelamin keturunan sepanjang peletakan telur berdasarkan kondisi lingkungan (Godfray 1994; Ode & Hunter 2002).
Strategi dalam mengatur alokasi kelamin keturunan oleh induk betina dilaporkan mempunyai pola tertentu. Strategi tersebut dilakukan sebagai tanggap terhadap kondisi lingkungan, dalam hal ini jumlah betina (Wanjberg 1994) atau kondisi jumlah inang (Colazza & Wajnberg1998). Pada umumnya, induk betina meletakkan keturunan jantan pada awal-awal peletakan telur, atau biasa disebut “strategi jantan pertama”. Penelitian Wanjberg (1993) menunjukkan bahwa pada jumlah inang tetap, induk betina parasitoid T. brassicae, akan memulai rangkaian peletakan telur dengan peletakan telur jantan dilakukan pada suatu jarak yang teratur. Sebaliknya hasil penelitian Tagawa (2000) menjelaskan bahwa pada parasitoid Cotesia glomerata L. (Hymenoptera: Braconidae) betina yang telah berkopulasi tidak mempunyai pola kecenderungan untuk meletakkan keturunan jantan atau betina pada awal ataupun pada akhir rangkaian peletakan telur.
Alokasi jenis kelamin keturunan yang diletakkan oleh induk betina, tergambar pada suatu pola peletakan telur yang pada akhirnya dapat dilihat pada nisbah kelamin keturunan. Alokasi keturunan ini meletakkan keturunan jantan pada awal peletakan telur. Strategi peletakan keturunan jantan pada awal-awal peletakan telur juga dilaporkan terjadi pada parasitoid T. basalis (Colazza & Wajnberg1998) dan pada parasitoid Telenomus busseolae Gahan (Hymenoptera: Scelionidae) (Bayram et al. 2004). Pada kedua penelitian ini dilaporkan bahwa strategi ”jantan pertama” terjadi pada semua perlakuan jumlah inang. Bila jumlah inang bertambah maka pada dasarnya dipengaruhi oleh faktor parasitoid dan faktor inang. Faktor parasitoid yakni kepadatan betina yang mengeksploitasi suatu
temperatur yang ekstrim juga akan mempengaruhi perilaku betina (Gordh et al.
1999).
Penelitian tentang alokasi kelamin keturunan karena pengaruh faktor kepadatan betina yang mengeksploitasi suatu patch, dilakukan dengan melihat pengaruhnya terhadap nisbah kelamin. Secara teoritikal, faktor kepadatan betina yang mempengaruhi nisbah kelamin keturunan dapat diprediksi dan dijelaskan berdasarkan teori alokasi kelamin keturunan. Teori Fisher memprediksi nisbah kelamin keturunan secara umum yaitu 50 : 50 (Godfray 1994). Kondisi ini dapat terjadi dengan asumsi setiap betina pada suatu populasi menghasilkan keturunan betina dan keturunan jantan dengan proporsi yang sama, atau dapat terjadi karena separuh betina menghasilkan keturunan jantan saja dan separuhnya menghasilkan keturunan betina saja. Model prediksi nisbah kelamin keturunan berdasarkan kepadatan parasitoid betina yang akhir-akhir ini banyak dikaji adalah menggunakan pendekatan teori local mate competition (LMC) yang dikemukakan oleh Hamilton (1967) yang memprediksi bahwa betina dapat me ngubah alokasi kelamin keturunan sebagai respon terhadap sibmating dan kepadatannya dalam suatu patch. Betina dapat memprediksi dan meletakkan lebih banyak telur yang tidak dibuahi pada kondisi dimana kepadatan betina tinggi. Berdasarkan teori ini didapatkan model bahwa nisbah kelamin (proporsi jantan) untuk setiap n
foundres adalah (n – 1)/2n (Hamilton 1967). Persamaan ini selanjutnya
(bias jantan). Parasitoid betina mampu mengubah penentuan kelamin keturunan
berdasarkan respon sibmating dan kepadatan betina dalam suatu kelompok. Wajnberg (1994) melaporkan bahwa pada parasitoid T. brassicae, nisbah kelamin keturunan akan berbeda bila hanya 1 parasitoid betina yang mengkolonisasi dengan nisbah kelamin 0,1791, dibandingkan dengan bila ada 2 atau 4 betina, maka nisbah kelamin meningkat berturut-turut menjadi 0,2525 dan 0,2735. Prediksi nisbah kelamin menggunakan pendekatan model LMC ini juga dilaporkan oleh Gauthier et al. (1997) pada parasitoid soliter Dinarmus basalis
Rond. (Hymenoptera: Pteromalidae) dan Debout et al. (2002) pada parasitoid
Leptopilina heterotoma (Hymenopetara: Eucoliidae) menjelaskan bahwa, proporsi keturunan jantan akan meningkat seiring dengan peningkatan kepadatan induk betina. Hal yang sama juga ditunjukkan dari hasil penelitian Flanagan et al.
(1998) yang menunjukkan bahwa bila terdapat dua betina parasitoid Nasonia vitripennis Walker (Hymenopetara: Pteromalidae) pada suatu patch, maka nisbah kelamin akan berkorelasi negatif dengan jumlah telur yang diletakkan. Jumlah telur yang diletakkan semakin sedikit seiring bertambahnya kepadatan betina. Hal lain yang juga dijelaskan adalah bahwa dalam suatu kondisi kepadatan betina yang sedikit (2 betina) maka nisbah kelamin yang dihasilkan berkorelasi positif dengan ukuran tubuh betina lainnya pada suatu patch. Betina yang mempunyai ukuran tubuh lebih besar akan meletakkan telur lebih banyak dan akibatnya akan menyebabkan nisbah keturunan menjadi bias betina.
Terjadinya superparasitisme merupakan pendekatan lain yang dapat menjelaskan kenapa nisbah kelamin akan meningkat bila kepadatan betina pada suatu patch bertambah. Bila kepadatan betina meningkat sedangkan jumlah inang tetap, maka ketersediaan inang menjadi terbatas, pada keadaan ini dapat terjadi superparasitisme. Bila terjadi superparasitisme pada satu inang, biasanya imago yang berhasil hidup adalah imago jantan (Werren 1984; Godfray 1994; Gauthier
et al. 1997).
relatif lebih besar dan keturunan jantan pada inang yang relatif lebih kecil
(Godfray 1994). Teori ini telah dibuktikan pada beberapa parasitoid Hymenoptera. Bernal et al. (1999) melaporkan bahwa ukuran inang berpengaruh nyata terhadap nisbah kelamin keturunan yang dihasilkan parasitoid betina Metaphycus stanleyi
Compere dan M. flavus Howard (Hymenoptera: Encyrtidae). Ode dan Heinz (2002) juga melaporkan bahwa keturunan betina pada parasitoid Dyglyphus isaeae
Walker (Hymenoptera: Eulophidae) akan muncul dari inang yang relatif lebih besar dan keturunan jantan akan muncul dari inang yang relatif lebih kecil. Hal yang sama juga dilaporkan oleh Mayhew dan Godfray (1997) pada prasitoid
Laelus pedatus Say. (Hymenoptera: Bethylidae), yang merupakan parasitoid yang dapat meletakkan satu sampai lima telur pada satu inang. Bila hanya meletakkan satu keturunan, keturunan jantan akan diletakkan pada inang berukuran kecil, sementara keturunan betina diletakkan pada inang berukuran besar. Bila meletakkan lebih dari satu telur pada satu inang maka pada inang yang besar akan diletakkan satu telur jantan dan satu atau lebih telur betina.
perlakuan umur inang pada inang Trichoplusia ni Hubner (Lepidoptera:
Noctuidae) dan Pieris rapae L (Lepidoptera: Pieridae), sementara pada inang P. xylostella umur inang berpengaruh pada nisbah kelamin.
Pengaruh jumlah inang yang tersedia pada satu kelompok juga diketahui dapat mempengaruhi nisbah kelamin keturunan. Colazza dan Wajnberg (1998) melaporkan bahwa nisbah kelamin keturunan yang dihasilkan parasitoid betina
T. basalis secara signifikan berbeda nyata pada jumlah inang yang berbeda.
BAHAN DAN METODE
Waktu dan Tempat
Penelitian dilaksanakan pada bulan Juni 2005 sampai dengan Maret 2006 bertempat di Laboratorium Bioekologi Parasitoid dan Predator, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor.
Persiapan Penelitian
Koleksi dan Perbanyakan Parasitoid
Parasitoid T. chilotraeae yang digunakan pada penelitian ini berasal dari areal pertanaman kubis di daerah Pangalengan Bandung. Koleksi parasitoid dilakukan dengan cara mengumpulkan kelompok-kelompok telur P. xylostella
yang ada pada daun kubis dengan menggunakan tangan (hand collection method). Telur yang terkumpul dibawa ke laboratorium dan diinkubasikan sampai parasitoid muncul dari telur yang terparasit. Parasitoid tersebut kemudian dikembangbiakkan pada telur inang pengganti C. cephalonica. Parasitoid dari perbanyakan ini kemudian diidentifikasi dan dibiakkan lebih lanjut sebagai bahan penelitian.
Perbanyakan parasitoid dilakukan dengan cara menempelkan telur inang pengganti C. cephalonica menggunakan gom arabik pada suatu pias (potongan karton yang berukuran 1 x 4 cm). Telur inang pada pias kemudian didinginkan di dalam freezer selama 2 jam dengan tujuan untuk membunuh embrio dalam telur. Telur inang selanjutnya dimasukkan ke dalam tabung reaksi untuk dipaparkan pada parasitoid. Parasitoid yang digunakan pada penelitian ini adalah keturunan F6 – F33.
Perbanyakan Serangga Inang Corcyra cephalonica
52 karton dengan diameter 8 cm dan tinggi 20 cm dengan bagian atas dan bawah yang ditutup dengan kawat kasa 25 mesh sebagai tempat bertelur C. cephalonica. Selang satu hari, telur-telur yang menempel pada kawat kasa dipanen dengan menggunakan kuas dan ditempatkan pada cawan petri. Sebagian telur digunakan untuk perbanyakan C. cephalonica itu sendiri dengan cara menaburkan telur pada kotak perbanyakan yang berisi media campuran pakan ayam dan dedak dengan ketebalan sekitar 3 cm dan sebagian lainnya digunakan untuk inang perbanyakan parasitoid dan inang pada pelaksanaan percobaan.
Pelaksanaan Penelitian
Identifikasi Parasitoid
Parasitoid Trichogrammatidae yang diperoleh dari lapangan dikembangbiakkan di laboratorium, selanjutnya diidentifikasi secara morfologi untuk menentukan genus dan spesies. Identifikasi didasarkan pada ciri-ciri sayap depan, sayap belakang, antena dan genitalia jantan berdasarkan kunci identifikasi Nagarkatti dan Nagaraja (1977), Alba (1988) dan Nurindah (2002).
Indentifikasi dilakukan dengan cara menyiapkan spesimen imago jantan parastoid. Untuk melihat morfologi luar parasitoid, dibuat dalam bentuk preparat sementara yaitu spesimen langsung ditata pada gelas obyek yang sebelumnya ditetesi larutan larutan Hoyer dan ditutup dengan gelas penutup. Sedangkan untuk melihat alat genitalia, spesimen parasitoid disiapkan dalam bentuk preparat permanen dengan cara dilakukan penjernihan terlebih dahulu. Dalam proses penjernihan, imago jantan ditempatkan dalam tabung reaksi yang diberi larutan KOH 10% kemudian direbus dalam air selama ± 1 jam. Setelah jernih, dilakukan penataan parasitoid di atas gelas obyek yang sebelumnya ditetesi larutan Hoyer dan ditutup dengan gelas penutup. Preparat kemudian dipanaskan pada hot plate
53
Pola Urutan Peletakan Telur
Parasitoid yang digunakan pada percobaan ini adalah parasitoid yang telah berumur 1 hari dan diperkirakan sudah berkopulasi serta telah diberi makan cairan madu 10%. Parasitoid ditempatkan pada tabung reaksi sesuai dengan perlakuan kepadatan parasitoid, yakni satu dan dua parasitoid. Sementara itu, pias ditempeli telur inang dengan cara merekatkan telur satu per satu dan disusun dalam barisan-barisan dimana jumlahnya disesuaikan dengan perlakuan, yakni 3, 6, 12, 24 dan 48 telur tiap pias. Pias kemudian dimasukkan ke dalam tabung reaksi yang sudah berisi parasitoid dan selanjutnya diamati di bawah mikroskop. Untuk mencatat setiap urutan telur inang yang diparasit maka terlebih dahulu dibuat skema yang sama dengan barisan-barisan telur yang ada pada pias pada selembar kertas. Pada skema inilah dicatat atau diberi kode dari setiap urutan pemarasitan yang teramati. Selama pengamatan berlangsung, betina yang akan melakukan superparasitisme (mendatangi inang yang sebelumnya sudah diparasit) dicegah dengan cara mengganggu betina dengan menggunakan kuas halus.
Setelah semua telur inang terparasit atau pemarasitan telah berlangsung selama 3 jam pada kelompok inang yang banyak, pias dikeluarkan dari dalam tabung. Telur inang yang sudah diparasit kemudian dipisahkan satu persatu menggunakan cutter dan selanjutnya dimasukkan ke dalam microtube dan diberi kode. Imago yang keluar kemudian diamati di bawah mikroskop untuk mengetahui kelamin keturunan pada setiap urutan peletakan telur.
Pada masing- masing kepadatan satu dan dua parasitoid betina, percobaan dilaksanakan dalam Rancangan Acak Lengkap (RAL) dengan lima perlakuan jumlah inang yakni 3, 6, 12, 24 dan 48 inang. Setiap perlakuan jumlah inang dilaksanakan dalam 20 ulangan sehingga terdapat 200 unit perlakuan.
Data kecenderungan kelamin keturunan yang dihasilkan pada setiap urutan peletakan telur pada masing- masing kepadatan parasitoid dianalisis secara deskriptif, diambil dari data persentase betina selama urutan peletakan telur. Analisis deskriptif juga dilakukan untuk menggambarkan kecenderungan
selfsuperparasitism sepanjang rangkaian peletakan telur.
54 menggunakan metode nonparametrik dengan uji Kruskal Wallis dan dilanjutkan dengan uji Sign test dan uji Wilcoxon signed rank test.
Data jumlah inang terparasit, persentase inang terparasit, persentase keturunan betina, persentase keturunan jantan dan persentase selfsuperparasitism
serta nisbah kelamin yang dihasilkan pada masing- masing perlakuan kepadatan satu dan dua parasitoid betina dianalisis dengan ANOVA dan dilajutkan dengan uji Tukey pada a 0,05. Pengaruh kepadatan parasitoid pada masing- masing perlakuan jumlah inang dianalisis menggunakan uji t. Data diolah menggunakan program Minitab.
Studi Pengaruh Jumlah Inang terhadap Efisiensi Pemarasitan
Percobaan ini dilakukan dengan membuat pias dengan jumlah inang berbeda yakni; 3, 6, 12, 24, dan 48 telur. Masing- masing pias ditempatkan dalam tabung reaksi. Pada setiap tabung reaksi tersebut dimasukkan satu betina parasitoid yang telah berumur 1 hari dan telah berkopulasi. Pemaparan dilakukan selama 24 jam. Setiap hari pias diganti dengan telur inang yang baru. Masing-masing perlakuan diulang sebanyak 20 kali. Telur yang sudah terparasit selanjutnya dipelihara pada suhu ruang 27oC. Pada hari ketiga dan keempat, telur yang tidak terparasit dihilangkan dan pias dipelihara kembali sampai imago muncul. Sexing dilakukan dengan bantuan mikroskop untuk menghitung proporsi keturunan yang dihasilkan.
55 Parameter yang diamati menggunakan rumus;
1. Total inang terparasit dihitung dengan menjumlahkan inang terparasit setiap hari selama parasitoid hidup.
2. Rata-rata inang terparasit per hari ( x ) dihitung dengan rumus; total inang terparasit
x =
lama masa produksi
3. Persentase pemunculan imago (PPI) diperoleh dengan cara menghitung persentase imago yang keluar dari telur inang yang terparasit;
m
PPI = x 100% N
Dimana:
m = jumlah imago yang muncul N = Total telur inang yang terparasit
3. Lama hidup dihitung mulai hari pertama imago keluar dari inang sampai imago mati.
4. Nisbah kelamin (NK) = proporsi betina r
NK = R
Dimana :
r = jumlah imago betina yang muncul R = Total imago yang muncul
Studi Waktu Pemarasitan Paling Efektif
56 rata-rata jumlah inang terparasit dianalisis dengan analisis deskriptif dan ditampilkan dalam bentuk grafik Histogram.
Studi Pengaruh Jumlah Betina terhadap Nisbah Kelamin
Percobaan diawali dengan membuat pias pada kepadatan 48 telur inang. Pias selanjutnya ditempatkan dalam tabung reaksi dan dipaparkan pada 1, 2, 3 dan 4 parasitoid betina. Masing- masing perlakuan diulang sebanyak 20 kali. Telur yang sudah terparasit selanjutnya dipelihara pada suhu ruang 27oC . Pada hari ketiga dan keempat, jumlah telur terparasit dihitung dan telur yang tidak terparasit dihilangkan. Pias dipelihara kembali sampai imago muncul. Sexing dilakukan dengan bantuan mikroskop untuk menghitung proporsi keturunan yang dihasilkan.
Data nisbah kelamin (proporsi betina) yang diamati dianalisis dengan ANOVA yang dilajutkan dengan Tuckey a 0,05. Data diolah menggunakan program Minitab.
Studi pengaruh jumlah betina parasitoid pada suatu kelompok inang yang tetap terhadap nisbah kelamin keturunan: Percobaan ini bertujuan untuk melihat apakah pertambahan jumlah parasitoid yang mengkolonisasi suatu kelompok inang yang tetap berpengaruh terhadap variasi nisbah kelamin keturunan. Percobaan dilakukan juga diawali dengan membuat pias dengan kepadatan inang berbeda yakni, 3, 6, 12, 24, dan 48 telur, selanjutnya setiap kepadatan inang dipaparkan pada 1, 2, 3 4, 5 dan 10 parasitoid betina yang telah berumur 1 hari dan telah berkopulasi, pemaparan dilakukan selama 24 jam. Telur yang sudah terparasit selanjutnya dipelihara pada suhu ruang 27oC sampai imago muncul, dan dengan bantuan mikroskop dihitung proporsi keturunan yang dihasilkan. Selanjunya hasil pengamatan dibandingkan dengan prediksi Local Mate Competititon oleh Hamilton : r = [(n – 1)/2n] [4n -2] / [4n – 1], dimana (r = Prediksi nisbah kelamin; n = jumlah betina parasitoid).
Peubah yang diamati menggunakan rumus :
1. Persen parasitisme (PP), dihitung dari persentase telur inang yang terparasit setiap harinya :
n
PP = x 100% N
57 Dimana :
n = telur yang terparasit
N = Total telur inang dalam setiap perlakuan
2. Persentase pemunculan imago (PPI), diperoleh dengan cara menghitung persentase imago yang keluar dari telur yang terparasit :
m
PPI = x 100% M
Dimana :
m = jumlah imago yang muncul N = Total telur inang yang terparasit
3. Lama masa reproduksi, dihitung mulai hari pertama imago betina meletakkan telur sampai imago mati.
4. Nisbah kelamin (NK) = (proporsi betina) r
NK = x 100% R
Dimana :
58
HASIL DAN PEMBAHASAN
Identifikasi Parasitoid
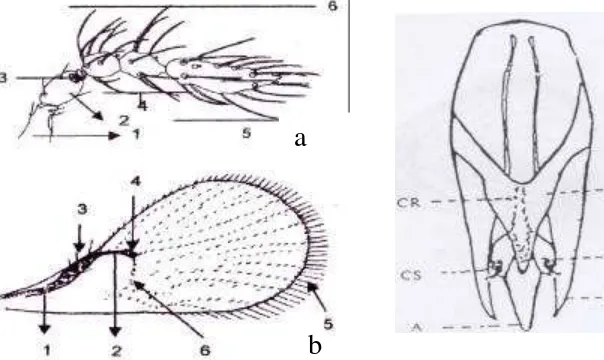
Berdasarkan hasil identifikasi terhadap karakter antena, sayap depan dan alat genitalia imago jantan menunjukkan bahwa parasitoid Trichogrammatidae yang digunakan pada penelitian ini adalah spesies T. chilotraeae (Gambar 1). Pada karakter antena ditemukan bagian club tidak bersegmen dengan rambut yang panjang (Gambar 1A). Sayap depan mempunyai setae yang letaknya tersusun dalam suatu susunan barisan, mempunyai tiga sampai empat trichia pada Rs1 dengan stigma yang mengarah kebawah dan fringe setae yang berukuran lebih pendek jika dibandingkan dengan genus Trichogrammatoidea (Gambar 1B). Alat genitalia jantan mempunyai dorsal expansion gonobase (DEG) kurang lebih berbentuk segitiga dengan ujung yang runcing, chelat structure (CS) yang besar dan terletak jauh dari ujung gonoforceps (GF) serta mempunyai aedagus yang panjang dan runcing. Hal ini sesuai dengan deskripsi berdasarkan sumber Nagarkatti dan Nagaraja (1977), Alba (1988) dan Nurindah (2002) (Gambar 2a, 2b dan 2c).
59
Gambar 2 Karakter morfologi T. chilotraeae berdasarkan kunci identifikasi; antena (a): 1-scapus, 2-pedicel, 3-anneli, 4-funicle, 5-club, 6-flagellum; Sayap (b): 1-submarginal vein, 2-pre marginal vein, 3-marginal vein, 4-stigma vein, 5-fringe setae,
6-Rs1; Alat genitalia (c): DEG (dorsal expansion gonobase); CR (chitized ridge); CS (chelate structure); MVP (median ventral projection) GF (gonoforceps); A (Aedagus). (a dan b sumber Nurindah 2002; c sumber Alba 1988).
a
b
c
Gambar 1 Karakter morfologi T. chilotraeae hasil identifikasi; Antena (A); sayap depan (B) dan genitalia imago jantan (C). Hasil pemotretan (kamera Olympus BX 50 pembesaran 400 x ), Laboratorium Bioekologi Parasitoid IPB (20 April 2006).
A
60
Spesies T. chilotraeae yang memarasit telur P. xylostella yang ditemukan pada penelitian ini relatif baru diketahui dan berbeda dengan spesies Trichogrammatidae yang dilaporkan sebelumnya. Meilin (1999) melaporkan, dari hasil eksplorasi di beberapa tempat di pulau Jawa, ditemukan tiga spesies Trichogrammatidae yang menyerang telur P. xylostella yakni T. flandersi, T. cojuangcoi dan T. armigera, di Filipina di laporkan dari spesies T. evanescens
yang diintroduksi dari Jerman (Alba 1988), sedangkan di India yakni spesies
T. armigera (Manjunath 1972 dalam Alba 1988). Selama ini, T. chilotraeae
dilaporkan memarasit telur H. armigera pada tanaman kapas dan jagung, O. furnacalis pada tanaman jagung,dan C. infuscatella pada tanaman tebu (Alba
1989 ; Nurindah & Bindra 1989), tetapi belum pernah dilaporkan menyerang telur
P. xylostella. Di Malaysia, India, Kamboja, Filipina dan Thailand, parasitoid ini dilaporkan menyerang telur hama penggerek batang padi bergaris C. suppressalis
pada tanaman padi dan O. furnacalis (Suasa 2002).
Perilaku Penemuan dan Penerimaan Inang
61 Pada saat pengamatan parasitoid betina terlihat melakukan gerakan memasukkan ovipositornya dalam inang, ternyata parasitoid tidak selalu meletakkan telur. Menurut Salt (1935) dalam Arthur (1981), untuk dapat diterima atau tidak suatu inang, maka selain melakukan drumming dan tapping, parasitoid akan kembali memeriksa dan mengukur inang pada saat memasukkan ovipositor. Bila inang dianggap cocok maka parasitoid akan segera meletakkan telur dan bila inang dianggap tidak cocok, maka ovipositor akan kembali dicabut tanpa melakukan kegiatan peletakan telur.
Parasitoid T. chilotraeae mampu mengenali inang yang sudah terparasit. Hal ini ditunjukkan oleh parasitoid yang tidak mau melakukan oviposisi pada telur inang yang sudah terparasit sebelumnya. Sejalan dengan penjelasan Doutt et al.
(1976) bahwa imago betina Trichogramma mempunyai kemampuan untuk membedakan antara inang yang sehat dan inang yang sudah terparasit melalui reseptor penginderaan pada antena, tarsi dan ovipositornya. Lebih jauh hasil penelitian Salt (1937) dalam Arthur (1981) menunjukkan bahwa parasitoid Trichogramma mempunyai kemampuan untuk membedakan antara telur yang bersih dan telur yang hanya dilewati oleh betina parasitoid lain dalam satu spesies. Jika inang hanya dilewati, maka parasitoid dapat dengan cepat membedakan dari
sehatnya inang dan kemudian menyerangnya. Sebagai contoh, parasitoid betina
T. evanescens menggunakan feromon untuk menandai telur yang telah dicoba atau telah diparasit dan ini memungkinkan betina lain untuk dapat membedakan antara telur inang yang siap diparasit dengan telur yang tidak bisa diparasit. Kemampuan untuk membedakan telur yang sudah terparasit ini menurut Jackson (1968) dalam
Arthur (1981) karena adanya feromon yang juga dimasukkan ke dalam inang bersamaan dengan peletakan telur. Sekresi ini kemungkinan dihasilkan dalam acid
62
Pola Urutan Peletakan Telur Parasitoid T. chilotraeae
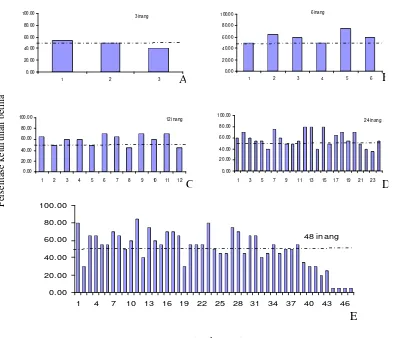
Parasitoid betina T. chilotraeae memberikan respon yang berbeda terhadap jumlah inang dan kehadiran betina lainnya berupa pengaturan kelamin keturunan ketika proses peletakan telur. Pada pemaparan satu parasitoid betina, pada perlakuan jumlah inang yang terbatas (3 inang), parasitoid meresponnya dengan selalu menghasilkan satu keturunan jantan pada setiap rangkaian tiga peletakan telur pertama. Keturunan jantan cenderung dihasilkan pada peletakan telur kedua atau ketiga, sehingga pola peletakan telur pada perlakuan 3 inang umumnya adalah ”betina-betina-jantan”, dengan nilai 80% pada peletakan telur pertama, 55% pada peletakan telur kedua dan 35% pada peletakan telur ketiga (Gambar 3A). Ketika jumlah inang bertambah, parasitoid betina tidak selalu menghasilkan keturunan jantan pertama pada tiga peletakan telur pertama.
Meskipun ada kecenderungan bahwa persentase keturunan betina berkurang pada peletakan telur ketiga, tetapi persentasenya masih lebih tinggi bila dibandingkan pada perlakuan 3 inang. Pada perlakuan 6 dan 12 inang, persentase keturunan betina paling rendah terjadi pada peletakan telur keempat, sehingga pola kelamin keturunan pada tiga peletakan telur pertama masih cenderung betina-betina-betina. Pada perlakuan 24, meskipun persentase keturunan betina menurun pada peletakan telur ketiga, tetapi pada peletakan telur kedua dihasilkan keturunan betina dengan persentase yang tinggi. Pada perlakuan 48 inang, persentase keturunan betina paling rendah baru dihasilkan pada peletakan telur ketujuh.
63
sampai beberapa hari kemudian, meskipun dengan jumlah yang semakin berkurang dibanding dengan hari pertama. Keturunan betina proporsinya juga terlihat semakin berkurang setiap hari pengamatan (Gambar 12). Alasan lain yang kemungkinan lebih tepat untuk menjelaskan hal ini adalah penjelasan Bayram et al. (2004), yang menyatakan bahwa perilaku tersebut merupakan strategi induk betina untuk mengantisipasi adanya kematian keturunan jantan yang diletakkan pada awal rangkaian peletakan telur.
Berdasarkan hasil tersebut, dapat dikatakan bahwa parasitoid T. chilotraeae
ini mempunyai kemampuan untuk mengukur jumlah inang yang ada dan meresponnya dengan mengubah alokasi keturunan betina atau jantan sepanjang peletakan telur. Kemampuan parasitoid untuk menaksir jumlah inang sesaat
64 sebelum melakukan peletakan telur dan mengatur alokasi kelamin keturunan pada suatu pola tertentu, kemungkinan disebabkan oleh beberapa petunjuk fisik yang telah diterima oleh parasitoid. Parasitoid yang mengeksploitasi suatu kelompok inang mempunyai kemampuan untuk mendapatkan informasi dengan cara berjalan dan mengetukkan antenanya pada atau dekat pinggiran suatu kelompok inang sebelum melakukan peletakan telur pertama (Colazza & Wajnberg1998). Selain itu, adanya petunjuk kimia yang dihasilkan oleh inang juga dapat dijadikan petunjuk oleh parasitoid untuk mengukur jumlah inang (Bayram et al. 2004). Mekanisme pertama kemungkinan yang digunakan oleh parasitoid T. chilotraeae
ini. Hal ini didasarkan pada pengamatan bahwa parasitoid mempunyai perilaku berjalan cepat secara acak melewati beberapa inang sambil mengetuk-ketukkan antenanya sebelum menetapkan pilihan pada salah satu inang. Perilaku ini kemungkinan merupakan cara parasitoid untuk mendapatkan informasi tersebut.
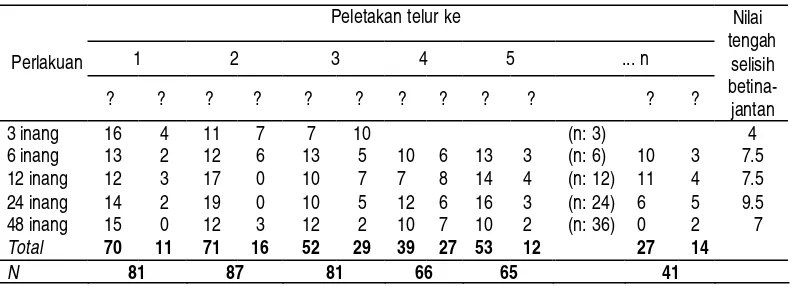
65 Tabel 1 Alokasi jenis kelamin keturunan yang dihasilkan induk parasitoid T.
chilotraeae pada peletakan telur ke-1 sampai peletakan telur ke- n Peletakan telur ke
Ket: n = peletakan telur terakhir
Berdasarkan hasil pengujian Kruskal-Wallis Test ternyata bahwa perlakuan jumlah inang 3, 6, 12, 24 dan 48 tidak berpengaruh nyata terhadap rasio kelamin keturunan betina-jantan pada setiap urutan peletakan telur (P = 0,403). Uji lanjut dengan Sign test dan Wilcoxon signed rank test, diperoleh nilai P= 0,000, yang berarti perbandingan kelamin keturunan (betina : jantan) bukan 50 : 50, dengan nilai tengah dari selisih keturunan betina dikurangi keturunan jantan adalah 8 dengan metode Sign test, dan nilai 7,5 dengan metode Wilcoxon signed rank test.
Hal ini menunjukkan bahwa pada dasarnya alokasi kelamin keturunan pada parasitoid T. chilotraeae ini tidak sejalan dengan prediksi menurut teori Fisher yang menyatakan bahwa setiap betina pada suatu populasi akan menghasilkan keturunan betina dan jantan dengan proporsi yang sama.
66 implikasi penting pada kegiatan pelepasan parasitoid di lapangan. Jika kondisi inang di lapangan rendah dan bila peletakan telur terakhir bias jantan, maka hal ini akan memberikan implikasi tertentu pada perkembangan populasi parasitoid.
Menurunnya proporsi betina pada akhir rangkaian peletakan telur pada perlakuan 24 dan 48 inang (juga digambarkan pada Gambar 1 D dan 1 E), ternyata disebabkan karena rendahnya jumlah inang terparasit pada akhir rangkaian peletakan telur, yakni hanya terdapat 11 inang terparasit pada perlakuan 24 inang dan 2 inang terparasit pada perlakuan 48 inang. Rendahnya inang terparasit pada akhir rangkaian peletakan telur disebabkan karena beberapa parasitoid sudah tidak memarasit lagi. Hal ini menunjukkan bahwa terdapat suatu kisaran optimum jumlah inang yang dapat diparasit oleh parasitoid T. chilotraeae
ini dalam jangka waktu 3 jam.
67
Meskipun dengan persentase yang berbeda, tetapi dapat dikatakan bahwa, induk betina parasitoid T. chilotraeae cenderung akan meletakkan keturunan betina pada peletakan telur awal. Hal ini terjadi baik pada pemaparan satu parasitoid maupun pada pemaparan dua parasitoid betina. Hasil ini berbeda bila dibandingkan dengan beberapa parasitoid Hymenoptera lainnya yang mempunyai pola reproduksi haplodiploid, dimana dilaporkan memulai rangkaian peletakan telur dengan menghasilkan keturunan jantan pada peletakan telur awal. Hasil penelitian Wanjberg (1993) menunjukkan, parasitoid T. brassicae, akan memulai rangkaian peletakan telur dengan menghasilkan keturunan jantan pada peletakan telur awal. Demikian pula pada hasil penelitian Colazza dan Wajnberg (1998)
3 inang
Gambar 4 Persentase keturunan betina yang dihasilkan pada setiap urutan peletakan telur pada berbagai kepadatan inang, pemaparan dua parasitoid betina.
( --- = 50% keturunan betina) Urutan peletakan telur
A B
C D
68 pada parasitoid T. basalis dan penelitian Bayram et al. (2004) pada parasitoid T. busseolae, yang melaporkan bahwa urutan peletakan telur pada berbagai jumlah inang dalam suatu paket tidak diletakkan secara acak, peletakan telur pertama pada semua perlakuan berbagai jumlah inang selalu jantan (strategi “jantan pertama”).
Dijelaskan lebih lanjut bahwa, parasitoid betina sebenarnya tidak mempunyai kemampuan untuk dapat menaksir jumlah inang sesaat sebelum melakukan rangkaian peletakan telur pada suatu kelompok inang. Pola peletakan telur pada berbagai jumlah inang hanya merupakan pengaruh tidak langsung dari strategi jantan pertama. Bila inang bertambah, maka setelah meletakkan keturunan jantan pertama pada awal peletakan telur, parasitoid selanjutnya akan meletakkan keturunan jantan kedua setelah meletakkan keturunan betina dalam jumlah yang banyak. Wajnberg (1993), menjelaskan bahwa perilaku ini sebenarnya ditentukan oleh kontrol genetik.
Bila dibandingkan dengan parasitoid T. chilotraeae pada penelitian ini, yang didasarkan pada pola alokasi kelamin keturunan pada tiga peletakan telur pertama (Tabel 2), dapat dikatakan bahwa parasitoid T. chilotraea juga mempunyai kecenderungan mengalokasikan keturunan jantan pertama pada peletakan telur ketiga dengan pola betina-betina-jantan, atau pada peletakan kedua dengan pola betina-jantan-betina. Akan tetapi, pola betina-betina-jantan atau betina-jantan-betina ini umumnya terjadi pada perlakuan 3 inang. Sedangkan pada pada perlakuan 6, 12, 24 inang umumnya pola betina-betina-betina. Jadi berdasarkan hal ini, dapat dikatakan bahwa, pengaturan alokasi kelamin keturunan sepanjang peletakan telur, dan dengan strategi peletakan keturunan betina pada awal rangkaian peletakan telur pada parasitoid T. chilotraea ini, kemungkinan bukan karena mekanisme kontrol genetik, akan tetapi memang disebabkan karena mekanisme kemampuan parasitoid untuk mengukur jumlah inang dan kepadatan parasitoid sebelum memulai rangkaian peletakan telur.
69 keturunan kemungkinan juga dapat dipengaruhi oleh spesies inang yang digunakan. Nurafiatin (2000) melaporkan bahwa proporsi keturunan betina pada parasitoid T. cojuangcoi akan berbeda bila dibiakkan pada inang berbeda. Proporsi keturunan betina parasitoid T. cojuangcoi lebih tinggi pada telur inang P. xylostella, dibandingkan pada telur inang C. cephalonica atau H. armigera.
Tabel 2 Frekuensi berbagai variasi pola urutan peletakan tiga telur pertama, parasitoid T. chilotraeae
Frekuensi (kali) Pola
peletakan
telur 3 inang 6 inang 12 inang 24 inang 48 inang
Total
? ? ? 8 7 5 5 25
? ? ? 11 3 6 7 2 29
? ? ? 7 5 4 1 9 26
? ? ? 3 2 2 1 8
? ? ? 1 1 1 3
? ? ? 1 1
? ? ? 1 1
Nisbah Kelamin
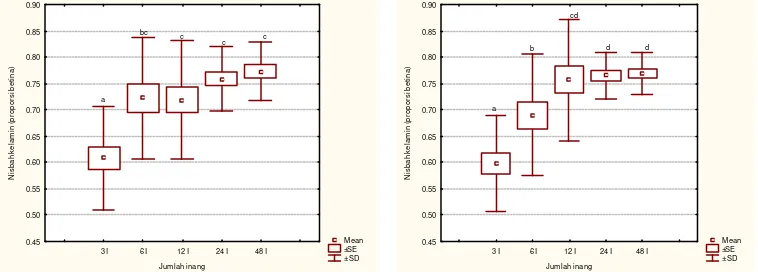
70
inang lainnya. Pada kepadatan inang 12, 24 dan 48 tidak berbeda nyata. Nisbah kelamin yang rendah pada perlakuan jumlah inang rendah, disebabkan karena selalu diletakkannya satu keturunan jantan pada tiga peletakan telur pertama. Sejalan dengan yang dikemukakan Wajnberg (1994), bahwa umumnya pada parasitoid Trichogramma, nisbah kelamin akan bias betina bila jumlah inang meningkat.
Bila melihat pengaruh kepadatan parasitoid terhadap nisbah kelamin, berdasarkan uji t, ternyata tidak terdapat pengaruh yang berbeda nyata antar perlakuan kepadatan betina pada masing- masing perlakuan (Tabel 3). Hasil ini sebenarnya bertolak belakang dengan hasil pengamatan pada bagian lain penelitian ini (perlakuan pengaruh kepadatan betina terhadap nisbah kelamin, (Gambar 14), dan tidak sejalan dengan prediksi local mate competition, yang menyatakan, nisbah kelamin akan menurun seiring bertambahnya kepadatan parasitoid. Akan tetapi hasil ini sejalan dengan hasil penelitian Flanagan et al.(1998) yang menjelaskan bahwa, pada kelompok/kepadatan parasitoid yang kecil (dua parasitoid) maka ternyata nisbah kelamin parasitoid Nasonia vitripennis
Walker (Hymenoptera: Pteromalidae) akan ikut meningkat. Tetapi dijelaskan lebih lanjut, hasil ini disebabkan karena faktor ukuran tubuh betina. Betina yang berukuran relatif lebih besar pada suatu kelompok inang, akan meletakkan telur dalam jumlah ya ng banyak, dan biasanya dengan proporsi betina yang lebih tinggi. Karena faktor tersebut akan meningkatkan nisbah kelamin keturunan.
Gambar 6 Nisbah kelamin keturunan dari dua induk betina yang meletakkan telur pada berbagai kepadatan inang Gambar 5 Nisbah kelamin keturunan dari
71 Pada penelitian ini, kemungkinan disebabkan karena tidak terjadi superparasitisme oleh betina lain, karena selama percobaan berlangsung, selalu dicegah terjadinya superparasitisme dengan cara menyentuh betina yang akan melakukan superparasitisme dengan menggunakan kuas halus. Superparasitisme diketahui sebagai penyebab menurunnya nisbah kelamin, karena jika terjadi superparasitisme pada suatu inang, umumnya keturunan yang berhasil menjadi imago adalah jantan.
Tabel 3 Rata-rata nisbah kelamin (proporsi betina) pada berbagai kepadatan inang dan kepadatan parasitoid
Nisbah kelamin pada berbagai kepadatan parasitoid betina (x ± SD) Kepadatan inang
1 parasitoid betina 2 parasitoid betina
P
3 inang 0,61 ± 0,10a 0,60 ± 0,09a 0,719 6 inang 0,72 ± 0,12a 0,69 ± 0,11a 0,354 12 inang 0,72 ± 0,11a 0,76 ± 0,11a 0,300 24 inang 0,76 ± 0,06a 0,76 ± 0,04a 0,699 48 inang 0,76 ± 0,06a 0,77 ± 0,40a 0,871 Ket : Nilai rataan dan simpangan baku selajur yang diikuti huruf sama tidak berbeda nyata
berdasarkan uji t pada a = 0.05
Pengaruh Jumlah Inang dan Kepadatan Parasitoid Terhadap Parasitisasi
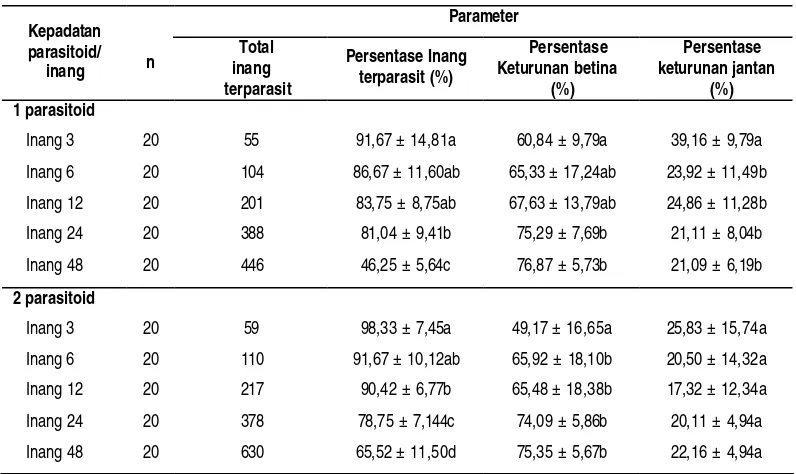
Analisis sidik ragam menunjukan bahwa pada pemaparan satu parasitoid betina, jumlah inang berpengaruh nyata terhadap persentase inang terparasit (P = 0,000), persentase keturunan betina (P = 0,000) dan persentase keturunan jantan (P = 0,000). Pada pemaparan dua parasitoid betina, jumlah inang berpengaruh nyata terhadap persentase inang terparasit (P = 0,000) dan persentase keturunan betina (P = 0,000), tetapi tidak berbeda nyata pada persentase keturunan jantan (P
= 0,210) (Tabel 4).
72 pemaparan dua parasitoid betina. Hasil ini menunjukkan bahwa parasitoid betina mempunyai kemampuan yang terbatas untuk memarasit inang dalam jumlah yang banyak pada jangka waktu 3 jam.
Tabel 4. Total inang terparasit, persentase inang terparasit, persentase keturunan betina dan persentase keturunan jantan pada berbagai jumlah inang.
Parameter
Ket : Pada masing-masing perlakuan kepadatan parasitoid, nilai rataan dan simpangan baku sekolom yang diikuti huruf yang sama tidak berbeda nyata berdasarkan uji Tukey pada a = 0,05
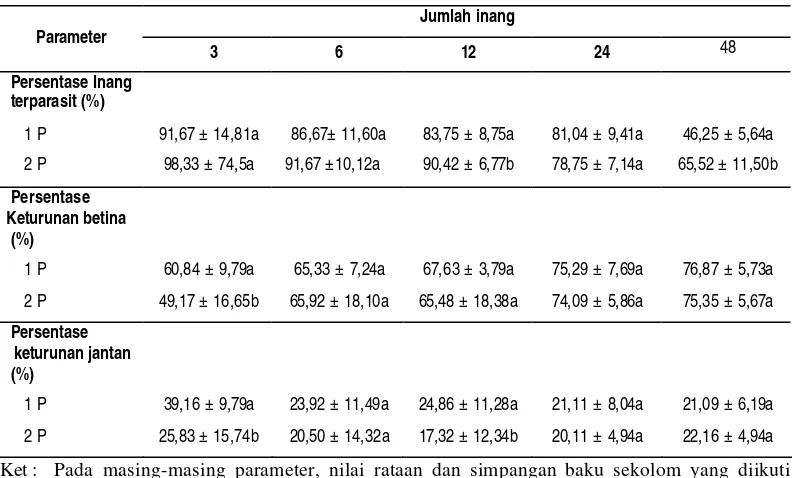
Pada pemaparan dua parasitoid betina, menunjukkan bahwa persentase inang terparasit pada setiap perlakuan memang lebih tinggi dibanding pada pemaparan satu parasitoid betina. Akan tetapi, berdasarkan uji t, menunjukkan bahwa kepadatan parasitoid berpengaruh nyata terhadap persentase inang terparasit pada perlakuan 12 inang (P = 0,011) dan 48 inang (P = 0,000) (Tabel 5). Meskipun berbeda nyata, tetapi parasitoid betina tidak mampu mengoptimalkan kemampuan memarasit secara optimum (dalam waktu 3 jam, total inang terparasit tidak menunjukkan angka dua kali lipat angka pada pemarasitan satu parasitoid betina). Hal ini menunjukkan bahwa kehadiran betina lain akan mempengaruhi kegiatan parasitisme.
73 keturunan jantan menjadi semakin berkurang. Berdasarkan hasil ini dapat dikatakan, secara umum parasitoid betina mempunyai perilaku untuk mengoptimalkan pemanfaatan sumberdaya yang ada untuk memaksimalkan kemampuan reproduksinya.
Bila dibandingkan antar perlakuan kepadatan betina. Berdasarkan uji t,
menunjukkan bahwa kepadatan parasitoid berpengaruh nyata terhadap persentase keturunan betina pada perlakuan 3 inang (P = 0,011), dan berpengaruh nyata pada persentase keturunan jantan pada perlakuan 3 inang (P = 0,003) dan 12 inang (P = 0,050).
Tabel 5 Persentase inang terparasit, persentase keturunan betina dan persentase keturunan jantan pada berbagai kepadatan parasitoid.
Jumlah inang Parameter
3 6 12 24 48
Persentase Inang terparasit (%)
1 P 91,67 ± 14,81a 86,67± 11,60a 83,75 ± 8,75a 81,04 ± 9,41a 46,25 ± 5,64a
2 P 98,33 ± 74,5a 91,67 ±10,12a 90,42 ± 6,77b 78,75 ± 7,14a 65,52 ± 11,50b
Persentase Keturunan betina
(%)
1 P 60,84 ± 9,79a 65,33 ± 7,24a 67,63 ± 3,79a 75,29 ± 7,69a 76,87 ± 5,73a
2 P 49,17 ± 16,65b 65,92 ± 18,10a 65,48 ± 18,38a 74,09 ± 5,86a 75,35 ± 5,67a
Persentase keturunan jantan (%)
1 P 39,16 ± 9,79a 23,92 ± 11,49a 24,86 ± 11,28a 21,11 ± 8,04a 21,09 ± 6,19a
2 P 25,83 ± 15,74b 20,50 ± 14,32a 17,32 ± 12,34b 20,11 ± 4,94a 22,16 ± 4,94a
Ket : Pada masing-masing parameter, nilai rataan dan simpangan baku sekolom yang diikuti huruf sama tidak berbeda nyata berdasarkan uji t pada a = 0,05
74
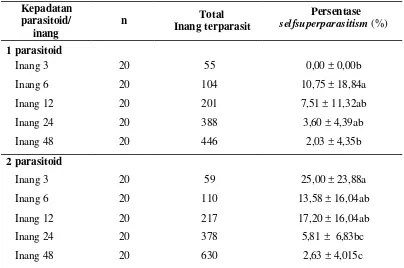
Persentase selfsuperparasitism
Analisis sidik ragam menunjukan bahwa jumlah inang berpengaruh nyata terhadap persentase selfsuperparasitim, baik pada pemaparan satu parasitoid betina (P = 0,009), maupun pada pemaparan dua parasitoid betina (P = 0,000) (Tabel 6). Pada pemaparan satu parasitoid betina, meskipun ada pengecualian pada inang 3, tetapi terjadi kecenderungan persentase selfsuperparasitism semakin berkurang seiring bertambahnya jumlah inang (10,75% pada perlakuan 6 inang dan menurun menjadi 2,03% pada perlakuan 48 inang). Pada pemaparan dua parasitoid betina juga terjadi kecenderungan yang sama bahwa persentase
selfsuperparasitism semakin berkurang seiring bertambahnya jumlah inang
(25,00% pada perlakuan 3 inang, menjadi 2,63% pada perlakuan 48 inang).
Tabel 6 Persentase selfsupeparasitsm pada berbagai jumlah inang
Kepadatan
Ket: Pada masing-masing perlakuan kepadatan parasitoid, nilai rataan dan simpangan baku sekolom yang diikuti huruf yang sama tidak berbeda nyata berdasarkan uji Tukey pada a = 0,05