OVER-EKSPRESI
GEN OsDREB1A GUNA PERBAIKAN
TOLERANSI CEKAMAN KEKERINGAN PADA PADI
BUDI SANTOSA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Over-ekspresi Gen OsDREB1A guna Perbaikan Toleransi Cekaman Kekeringan pada Padi adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Juli 2012
ABSTRACT
BUDI SANTOSA. Over-expression of OsDREB1A gene for improvement of drought tolerance on rice. Under supervision of Sobir, Sriani Sujiprihati (Alm), Kurniawan Rudi Trijatmiko, and Sudirman Yahya.
RINGKASAN
BUDI SANTOSA. Over-ekspresi Gen OsDREB1A guna Perbaikan Toleransi Cekaman Kekeringan pada Padi. Dibimbing oleh Sobir, Sriani Sujiprihati (Alm), Kurniawan Rudi Trijatmiko, dan Sudirman Yahya.
Beras merupakan bahan pangan pokok yang paling dominan di Indonesia, dan kebutuhan akan beras setiap tahun bertambah terus seiring dengan meningkatnya jumlah penduduk. Peningkatan jumlah penduduk ini jika tanpa disertai dengan peningkatan produksi padi yang seimbang akan menyebabkan kerawanan pangan. Untuk memenuhi kebutuhan dalam negeri pemerintah telah mengimpor beras selain meningkatkan produksi padi dalam upaya menjaga kestabilan harga beras agar tetap terjangkau oleh semua pihak. Untuk mengurangi impor beras perlu diupayakan peningkatan produksi padi. Namun upaya peningkatan produksi padi ini menghadapi berbagai permasalahan, antara lain penyempitan areal lahan sawah karena beralihnya lahan potensial untuk tanaman padi menjadi pemukiman dan industri, iklim yang tidak menentu yang menyebabkan musim kemarau menjadi lebih lama dan persediaan air untuk tanaman menjadi berkurang.
Kehilangan hasil pada tanaman padi akibat cekaman kekeringan antara lain tergantung dari lamanya cekaman dan fase pertumbuhan tanaman waktu mengalami cekaman. Karakter toleran terhadap cekaman pada tanaman padi sangat komplek dan dikendalikan oleh banyak gen, sehingga menyulitkan para pemulia untuk merakit tanaman padi yang toleran terhadap kekeringan.
Perakitan varietas padi toleran kekeringan dapat dilakukan melalui persilangan klasik maupun transformasi genetik. Pada penelitian ini digunakan gen OsDREB1A yang termasuk gen faktor transkripsi. Over-ekspresi gen OsDREB1A dapat menginduksi ekspresi yang kuat dari sejumlah gen-gen target yang produknya dapat memberikan proteksi tanaman terhadap efek yang merusak dari cekaman kekeringan. Gen OsDREB1A dengan promotor 35S yang ada dalam plasmid pCambia 1301 ditransformasikan ke dalam genom padi Nipponbare dengan bantun A. tumefaciens. Hasil transformasi diperoleh 81 tanaman padi putatif transgenik (T0) dan positif mengandung pita DNA berukuran 500 bp berdasarkan analsis PCR menggunakan primer HPTII
Evaluasi tanaman transgenik hasil transformasi untuk mendapatkan genotipe padi transgenik yang toleran terhadap kekeringan dilaksanakan di rumah kaca. Benih padi dari sembilan belas genotipe padi transgenik ditanam pada media 100 mg/l higromisin. Bibit padi transgenik yang lolos seleksi higromisin kemudian ditanam pada pot yang berisi media tanah, pasir, dan kompos untuk uji kekeringan. Hasil evaluasi diperoleh lima tanaman padi transgenik yang lebih toleran dari Nipponbare non-transgenik dan kontrol toleran (varietas Kalimutu) dan mempunyai 2 – 4 salinan transgen berdasarkan analisis Southern hibridisasi. Kelima genotipe tersebut adalah B14, B11, C14, C10, dan C3.
diperoleh tujuh tanaman dari tiga genotipe padi elit yang lebih toleran terhadap kekeringan dibandingkan kontrol induknya. Ketiga genotipe tersebut adalah Batutegi, Code, dan Konawe. Silang balik antara tanaman F1 padi elit transgenik dengan padi elit diperoleh lima tanaman padi yang berasal dari tiga genotipe elit transgenik. Hasil analisis PCR menggunakan primer HPTII menunjukkan bahwa kelima tanaman tersebut mengandung pita DNA berukuran 500 bp. Hal ini menunjukkan bahwa gen tersebut dapat dipindah dan diturunkan pada padi elit melalui persilangan. Berdasarkan penelitian ini, diperoleh tiga genotipe padi elit transgenik yang lebih toleran terhadap kekeringan yaitu Batutegi, Code, dan Konawe transgenik yang mungkin berguna untuk program pemuliaan perbaikan toleransi kekeringan pada padi di Indonesia.
Hak Cipta milik IPB, tahun 2012
Hak Cipta dilindungi Undang-undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber.
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah.
b. Pengutipan tidak merugikan kepentingan yang wajar bagi IPB
OVER-EKSPRESI
GEN OsDREB1A GUNA PERBAIKAN
TOLERANSI CEKAMAN KEKERINGAN PADA PADI
BUDI SANTOSA
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Disertasi : Over-ekspresi Gen OsDREB1A guna Perbaikan Toleransi Cekaman Kekeringan pada Padi Nama Mahasiswa : BUDI SANTOSA
Nomor Pokok :A263070081
Disetujui Komisi Pembimbing
Dr. Ir. Sobir, MSi Prof. Dr. Ir. Sriani Sujiprihati, MS (Alm)
Ketua Anggota
Dr. Kurniawan Rudi Trijatmiko. SP, MSi Prof. Dr. Sudirman Yahya MSc Anggota Anggota
Diketahui,
Ketua Program Studi Dekan Sekolah Pascasarjana Pemuliaan dan Bioteknologi Tanaman
Dr. Ir. Trikoesoemaningtyas, MSi Dr. Ir. Dahrul Syah, MSc. Agr
PRAKATA
Puji syukur penulis panjatkan kepada Tuhan Yesus Kristus, atas hikmatNya hingga penulis dapat menyelesaikan desertasi ini. Judul desertasi ini ialah Over-ekspresi Gen OsDREB1A guna Perbaikan Toleransi Cekaman Kekeringan pada Padi.
Pada kesempatan ini penulis menyampaikan ucapan terimakasih kepada Dr. Ir. Sobir, MSi., Prof. Dr. Ir. Sriani Sujiprihati, MS.(Alm), Dr. Kurniawan Rudi Trijatmiko, MSi., dan Prof. Dr. Ir. Sudirman Yahya, MSc. Selaku pembimbing atas segala saran, bimbingan dan kritik selama penelitian dan penulisan desertasi. Terima kasih kepada Kepala Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian, Bogor. Kepala Badan Penelitian dan Pengembangan Pertanian, atas kesempatan yang diberikan untuk studi dan beasiswa yang penulis terima.
Ucapakan terima kasih disampaikan kepada rekan-rekan staf peneliti dan teknisi di kelompok peneliti Biologi Molekuler maupun kelompok penelitian lainnya serta staf administrasi di B B Biogen atas bantuan, dukungan dan pengertiannya.
Terima kasih kepada rekan-rekan Program Studi Ilmu Teknologi Benih, Agronomi dan Hortikultura, program lainnya serta khususnya program Studi Pemuliaan dan Bioteknologi Tanaman atas persaudaraannya. Terima kasih untuk Ir. Sutono (BBSDLP, Bogor), sdr Asep Wahyudi dan Endang Ibrahim yang telah banyak membantu selama pelaksanaan penelitian.
Ungkapan terima kasih kepada Bapak (Alm.) dan Ibu serta seluruh keluarga atas dukungan dan doanya. Terimakasih juga kepada sahabat-sahabatku yang tidak dapat penulis sebutkan satu persatu atas doa dan dukungan dalam menyeselesaikan pendidikkan ini.
Kiranya Tuhan Yesus Kristus memberkati semua pihak yang telah membantu dan mendukung dalam penelitian dan penulisan desertasi ini. Semoga karya ilmiah dapat bermanfaat bagi kita semua.
Bogor, Juli 12012
RIWAYAT HIDUP
Penulis dilahirkan di Solo, Jawa Tengah, pada tanggal 12 Juni 1961 sebagai anak kedua dari lima bersaudara dari pasangan Soeharto (Alm) dan Sri Soemarni. Penulis memperoleh gelas Sarjana Pertanian di Fakultas Pertanian, Universitas Sebelas Maret Surakarta pada tahun 1986 Jurusan Budidaya Pertanian. Tahun 2002 memperoleh gelar Magister Pertanian dari Program Studi Ilmu Tanaman, Bidang Kajian Utama Ilmu Pemuliaan Tanaman, Program Pascasarjana, Universitas Padjadjaran, Bandung melalui beasiswa pendidikan dan dana penelitian proyek ARMPII, Badan Penelitian dan Pengembangan Pertanian, Kementrian Pertanian. Pada tahun 2007 melanjutkan studi Program Doktor (S3) dengan biaya APBN Badan Penelitian dan Pengembangan Pertanian, Kementrian Pertanian pada Program Studi Pemuliaan dan Bioteknolgi Tanaman, Departemen Agronomi dan Hortikultura, Institut Pertanian Bogor.
DAFTAR ISI
TRANSFORMASI GEN OsDREB1A DENGAN Agrobacterium tumefaciens DAN REGENERASI TANAMAN PADI Abstrak ... 13
UJI TOLERANSI KEKERINGAN TANAMAN PADI TRANSGENIK NIPPONBARE 35S::OsDREB1A
A. Seleksi Tanaman Transgenik Hasil Transformasi terhadap
Kekeringan ... B. Studi Fisiologi Gen OsDREB1A ...
Hasil Penelitian 34
...
Pembahasan 40 A. Seleksi tanaman transgenik hasil transformasi terhadap kekeringan . B. Studi fisiologi gen OsDREB1A ...
Simpulan ... 43
Daftar pustaka ... 43
PEMINDAHAN GEN 35S::OsDREB1A DARI PADI TRANSGENIK KE VARIETAS ELIT PADI MELALUI PERSILANGAN Abstrak ... 45
Abstract ... 45
Pendahuluan ... 46
Tujuan Penelitian ... 47
Metodologi Penelitian ... 47
Hasil Penelitian ... 49
Pembahasan ... 54
Simpulan ... 57
Daftar Pustaka ... 57
PEMBAHASAN UMUM ... 59
SIMPULAN DAN SARAN ... 63
DAFTAR TABEL
1. Hasil transformasi dan regenerasi tanaman padi varietas Nipponbare putatif transgenik (T0) dengan menggunakan 35S::OsDREB1A ...
18 2. Klasifikasi tanggap tanaman terhadap kekeringan (daun menggulung
pada fase vegetatif) ... 27 3. Klasifikasi tanggap tanaman terhadap kekeringan (daun mengering
pada fase vegetatif) ... 28 4. Klasifikasi tanggap kesembuhan tanaman setelah
perlakuan pengeringan ... 28 5. Skor hasil pengujian kekeringan tanaman padi putatif transgenik
(T1) dan tanaman non-transgenik pada fase vegetatif di rumah kaca
Fasilitas Uji Terbatas (IRRI 1996) ... 35 6. Hasil analisis PCR enam genotipe dan hibridisasi Southern lima
genotipe padi transgenik ... 36 7. Rerata kerapatan stomata daun per mm2 dan evapotranspirasi ... 37 8. Persentase tanaman hidup dan rerata daun hijau pada waktu pemulihan 38 9. Bobot basah dan bobot tajuk, bobot basak dan kering akar, panjang
akar, nisba tajuk:akar, dan bobot tajuk kering/evapotranspirasi ... 39 10. Klasifikasi tanggap tanaman terhadap kekeringan (daun menggulung
pada fase vegetatif) ... 48 11. Klasifikasi tanggap tanaman terhadap kekeringan (daun mengering
pada fase vegetatif) ... 48 12. Klasifikasi tanggap kesembuhan tanaman setelah
perlakuan pengeringan ... 48 13. Skor hasil pengujian kekeringan pada fase vegetatif dari genotipe
DAFTAR GAMBAR
1. Kerangka alur penelitian tanaman padi trangenik yang mengandung
gen OsDREB1A untuk perbaikan toleransi terhadap kekeringan ... 5
2. Mekanisme toleransi gen DREB1A terhadap kekeringan ... 8
3. Proses transformasi dengan Agrobacterium tumefaciens ... 10
4. Segregasi tanaman transgenik dengan satu salinan transgen ... 12
5. Profil plasmid pCambia 1310 (11 kb) dan pr35S::OsDREB1A::T35S (1,6 kb) yang digunakan untuk transformasi ... 17
6. Hasil amplifikasi PCR DNA 14 genotipe tanaman padi Nipponbare putatif transgenik (T0) dengan menggunakan primer HPTII ... 18
7. Distribusi frekuensi tinggi tanaman padi transgenik (T0) ... 18
8. Distribusi frekuensi rerata panjang malai per rumpun dan jumlah malai per rumpun tanaman padi transgenik (T0) ... 19
9. Distribusi frekuensi jumlah gabah isi per rumpun dan persentase gabah hampa per rumpun tanaman padi transgenik ... 19
10. Distribusi frekuensi jumlah gabah total per rumpun tanaman transgenik (T0) ... 19
11. Induksi dan seleksi kalus padi Nipponbare ... 20
12. Regenerasi kalus padi Nipponbare ... 20
13. Hasil analisis PCR dan Southern hibridisasi sampel DNA padi Nipponbare transgenik ... 35 14. Penampilan tanaman padi Nipponbare transgenik generasi T1 setelah uji kekeringan ... 36
15. Evapotranspirasi selama perlakuan pengeringan ... 37
16. Evapotranspirasi selama perlakuan pemulihan ... 37
17. Hasil analisis PCR sampel DNA tanaman padi transgenik, Nipponbare, Singkarak dan Kalimutu dengan menggunakan primer HPTII ... 39
18. Proses persilangan untuk memindahkan gen OsDREB1A ke genotipe padi elit ... 50
19. Penampilan tanaman F1 transgenik setelah perlakuan kekeringan (10 hsp) pada fase vegetatif ... 51
20. Penampilan tanaman F1 padi elit transgenik ... 51
21. Amplifikasi PCR tujuh tanaman padi F1 elit transgenik ... 51
22. Jumlah malai per rumpun F1 Batutegi transgenik dan non-transgenik .. 52
23. Jumlah gabah isi per rumpun dan persentase gabah hampa per rumpun F1 Batutegi transgenik dan non-transgenik ... 52
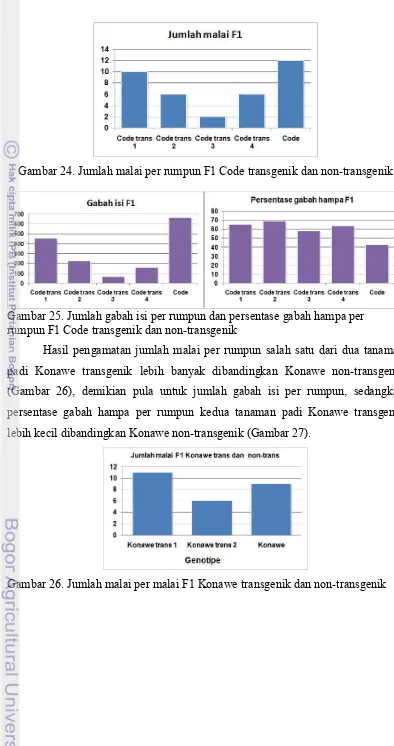
24. Jumlah malai per rumpun F1 Code transgenik dan non-transgenik ... 53
25. Jumlah gabah isi per rumpun dan persentase gabah hampa per rumpun F1 Code transgenik dan non-transgenik ... 53
26. Jumlah malai per rumpun F1 Konawe transgenik dan non-transgenik ... 53
27. Jumlah gabah isi per rumpun dan persentase gabah hampa per rumpun F1 Konawe transgenik dan non-transgenik ... 54
DAFTAR LAMPIRAN
1. Diskripsi varietas padi unggul Indonesia yang digunakan dalam
penelitian ... 70
2. Komposisi media pertumbuhan bakteri ... 76
3. Komposisi media transformasi dan regenerasi padi ... 77
4. Komposisi larutan untuk hibridisasi southern ... 80
ABSTRAK
Kekeringan merupakan salah satu faktor yang berdampak pada penurunan produksi tanaman padi. Salah satu usaha untuk menanggulangi masalah kekeringan adalah menanam varietas toleran. Perakitan varietas padi toleran terhadap cekaman kekeringan dapat dilakukan menggunakan teknik rekayasa genetik dengan mengintroduksikan gen ke genom padi melalui Agrobacterium tumefaciens. Penelitian ini bertujuan untuk mendapat tanaman padi putatif transgenik yang mengandung gen OsDREB1A yang ekspresinya dikendalikan oleh promoter 35S. Kalus yang berasal dari biji padi varietas Nipponbare ditransformasi dengan konstruk 35S::OsDREB1A melalui Agrobacterium, kemudian diregenerasikan dan diaklimatisasi hingga menghasilkan tanaman padi putatif transgenik. Hasil penelitian menunjukkan bahwa konstruk 35S::OsDREB1A dapat ditransformasikan ke genom padi Japonica Nipponbare dengan metode A. tumefaciens dan seleksi higromisin. Hasil analisis PCR menggunakan primer HPTII memperlihatkan bahwa semua tanaman padi yang dianalisis positif mengandung transgen. Terdapat variasi morfologi pada tanaman padi putatif transgenik generasi T0.
Kata kunci : Transformasi, regenerasi, padi, gen OsDREB1A
ABSTRACT
Drought is a critical factor affecting the reduction in rice production. One of the efforts to overcome the problem of drought is to grow tolerant varieties. Creating of rice varieties tolerant to drought stress can be made using genetic engineering techniques by introducing genes into rice genome by Agrobacterium tumefaciens. The aim of this study was to obtain putative transgenic rice plants containing OsDREB1A gene under the promoter 35S. Calli derived from seeds of rice variety Nipponbare were transformed using 35S::OsDREB1A construct mediated by Agrobacterium, then regenerated and acclimatized to generate putative transgenic rice plants. The results showed the 35::OsDREB1A construct could be transformed into Japonica rice cv. Nipponbare genome with A. tumefaciens method and hygromycine selection. PCR analysisi using the primers HPTII showed that all analyzed rice plants contained transgene. There were variation on the morphology of putative transgenic rice T0 plants.
Pendahuluan
Perakitan varietas padi toleran kekeringan dapat dilakukan melalui pemuliaan klasik maupun menggunakan teknologi transformasi genetik. Sifat toleransi kekeringan pada padi dikendalikan oleh banyak gen (Kasuga et al. 1999, Shinozaki dan Yamaguchi-Shinozaki 2004), sehingga sulit untuk merakit varietas toleran bila menggunakan pemuliaan klasik. Perakitan varietas padi dengan menggunakan teknik transformasi genetik diharapkan dapat menghasilkan varietas padi unggul dengan sifat-sifat yang dikehendaki. Transfomasi genetik dengan perantaraan Agrobacterium tumefaciens merupakan salah satu teknik transformasi yang banyak dilakukan oleh para peneliti dan telah berhasil dengan baik. Agrobacterium merupakan bakteri yang berperanan untuk membantu menyisipkan gen ke dalam genom tanaman, misalnya pada transformasi tanaman kentang dengan gen RB untuk meningkatkan ketahanan terhadap penyakit hawar daun ( Song et al. 2003), tanaman padi dengan gen CBF3/DREB1A dan ABF3 dari Arabidobsis untuk meningkatkan toleransi terhadap cekaman abiotik (Oh et al. 2005).
Gen DREB1A termasuk gen faktor transkripsi yang berperan dalam meregulasi sejumlah gen lain yang berhubungan dengan karakter kekeringan ( Liu et al. 1998). Hasil penelitian Dubouzet et al. (2003) menunjukkan bahwa over ekspresi dari gen OsDREB1A yang berasal dari genom padi pada tanaman Arabidopsis transgenik dapat menginduksi over ekspresi gen target DREB1A, dan menghasilkan tanaman Arabidopsis yang lebih toleran terhadap cekaman kekeringan, kadar garam tinggi, dan suhu rendah. Gen-gen yang berbasis DREB (Dehydration Response Element Binding) yang telah diisolasi oleh Dubouzet et al. (2003) dari genom padi adalah OsDREB1A, OsDREB1B, OsDREB1C, OsDREB1D, dan OsDREB2A.
Tanaman padi subgroup japonica yang ditransfomasi dengan 35S::OsDREB1A memperlihatkan lebih toleran terhadap kekeringan (Ito et al. 2006). Namun, efek over-ekspresi gen OsDREB1A pada toleransi kekeringan taanaman padi subgroup indica belum diketahui.
Tujuan Penelitian
Untuk mendapatkan tanaman padi cv. Nipponbare yang mengandung konstruk 35S::OsDREB1A.
Metodologi Penelitian
a. Bahan Genetik
Gen 35S::OsDREB1A yang dikonstruk oleh Mulyana (2010) ditransformasi ke genom padi Nipponbare melalui Agrobacterium tumefaciens. b. Bahan tanaman
Benih padi genotipe Nipponbare yang digunakan dalam penelitian ini diperoleh dari International Rice Research Institute.
c. Pelaksanaan Penelitian
Penelitian ini dilaksanakan di Laboratorium Biologi Molekuler dan Rumah kaca Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumber Daya Genetik Pertanian, Bogor, Februari 2008 – Maret 2010.
Transformasi
(Media dasar R2 + 30 g/l glukosa (pH 6,0) + 3g/l phytagel + 2,5 mg/l 2.4-D + 400 mg/l cefotaxime + 100 mg/l vancomycine + 50 mg/l hygromycine), kemudian diinkubasikan dalam ruang gelap (28oC) selama dua minggu dan disubkultur pada media dan kondisi tumbuh yang sama selama 2 minggu. Kalus yang embriogenik dipindahkan ke dalam media induksi embrio (Media dasar LS + 30 g/l sukrosa (pH 5,8) + 100 ml/l air kelapa + 3g/l phytagel + 2,5 mg/l 2.4-D + 400 mg/l cefotaxime + 100 mg/l vancomycine + 50 mg/l hygromycine) dan diinkubasi dalam ruang gelap (28oC) selama dua minggu.
Regenerasi Tanaman. Kalus embriogenik yang terseleksi dipindahkan pada media regenerasi embrio (Media dasar LS + 40 g/l sukrosa (pH 5,8) + 3g/l phytagel + 0,5 mg/l IAA + 0,3 mg/l BAP + 400 mg/l cefotaxime + 100 mg/l vancomisin + 50 mg/l higromisin) dan diinkubasikan dalam ruang kultur dengan cahaya kontinyu selama dua minggu pada suhu 25oC. Kalus yang ada bercak hijau disubkultur pada media yang sama. Kalus yang sudah tumbuh daun/tunas sepanjang minimal sekitar 1 cm disubkultur pada media perakaran (Media dasar LS + 30 g/l sukrosa (pH 5,8) + 3 g/l phytagel, diautoklaf + 40 g/l higromisin). Planlet disimpan dalam ruang kultur dengan cahaya kontinyu pada suhu ruang (280C).
Aklimatisasi Tanaman. Planlet (tunas) yang berakar dan tinggi tajuk minimal 5 cm dipindahkan ke media air selama satu minggu dan disimpan pada suhu ruang. Planlet selanjutnya dipindahkan ke media tanah dalam bak plastik untuk pembibitan selama dua sampai tiga minggu. Bibit tanaman padi dipindahkan pada media tanah dalam ember dirumah kaca dan dipelihara sampai menghasilkan biji. Pengamatan dilakukan terhadap tinggi tanaman, jumlah anakan, panjang malai, jumlah gabah.
Analisis Molekuler dengan Teknik PCR
DNA) diamplifikasi menggunakan mesin PCR. Profil suhu yang digunakan adalah predenaturasi 94 oC selama 5 menit, dilanjutkan dengan 35 siklus meliputi pemisahan (denaturation) pada suhu 94oC selama 1 menit, penempelan primer (annealing) pada suhu 55oC selama 1 menit, pemanjangan primer (extension) pada suhu 72 oC selama 2 menit. Pada tahap akhir PCR dilakukan pemanjangan akhir pada 72 oC selama 5 menit. Produk amplifikasi diseparasi pada elektrofloresis gel agarose. Fragmen-fragmen DNA pada gel agarose direndam dalam larutan ethidium bromida dan divisualisasi menggunakan Chemidoc gel system.
Hasil Penelitian
Plasmid pCambia 1301 yang mengandung 35S::OsDREB1A berukuran sekitar 12,6 kb. Pemotongan plasmid tersebut menggunakan enzim EcoRI dihasilkan dua fragmen yang berukuran 11 kb dan 1,6 kb, yang masing-masing adalah vektor biner bakcbone pCambia 1301 dan kaset pr35S:OsDREB1A::T35S ( Gambar 6). Konfirmasi ini bertujuan untuk memastikan bahwa plasmid yang akan digunakan untuk transformasi sudah benar.
Gambar 5. Profil plasmid pCambia 1301 (11 kb) dan pr35S::OsDREB1A::T35S (1,6 kb) yang digunakan untuk transformasi
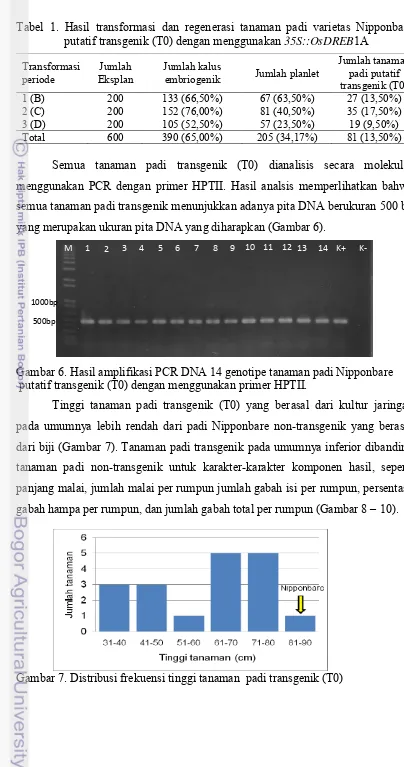
Tabel 1. Hasil transformasi dan regenerasi tanaman padi varietas Nipponbare putatif transgenik (T0) dengan menggunakan 35S::OsDREB1A
Transformasi periode
Jumlah Eksplan
Jumlah kalus
embriogenik Jumlah planlet
Jumlah tanaman padi putatif transgenik (T0)
1 (B) 200 133 (66,50%) 67 (63,50%) 27 (13,50%)
2 (C) 200 152 (76,00%) 81 (40,50%) 35 (17,50%)
3 (D) 200 105 (52,50%) 57 (23,50%) 19 (9,50%)
Total 600 390 (65,00%) 205 (34,17%) 81 (13,50%)
Semua tanaman padi transgenik (T0) dianalisis secara molekuler menggunakan PCR dengan primer HPTII. Hasil analsis memperlihatkan bahwa semua tanaman padi transgenik menunjukkan adanya pita DNA berukuran 500 bp yang merupakan ukuran pita DNA yang diharapkan (Gambar 6).
Gambar 6. Hasil amplifikasi PCR DNA 14 genotipe tanaman padi Nipponbare putatif transgenik (T0) dengan menggunakan primer HPTII.
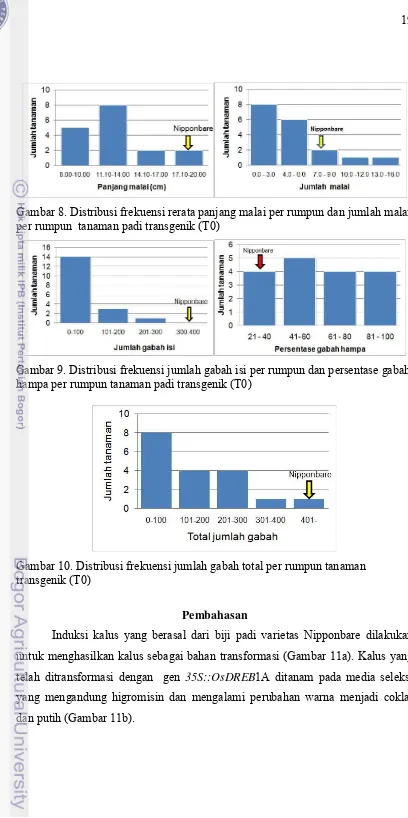
Tinggi tanaman padi transgenik (T0) yang berasal dari kultur jaringan pada umumnya lebih rendah dari padi Nipponbare non-transgenik yang berasal dari biji (Gambar 7). Tanaman padi transgenik pada umumnya inferior dibanding tanaman padi non-transgenik untuk karakter-karakter komponen hasil, seperti panjang malai, jumlah malai per rumpun jumlah gabah isi per rumpun, persentase gabah hampa per rumpun, dan jumlah gabah total per rumpun (Gambar 8 – 10).
Gambar 7. Distribusi frekuensi tinggi tanaman padi transgenik (T0)
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 K+ K‐
Gambar 8. Distribusi frekuensi rerata panjang malai per rumpun dan jumlah malai per rumpun tanaman padi transgenik (T0)
Gambar 9. Distribusi frekuensi jumlah gabah isi per rumpun dan persentase gabah hampa per rumpun tanaman padi transgenik (T0)
Gambar 10. Distribusi frekuensi jumlah gabah total per rumpun tanaman transgenik (T0)
Pembahasan
Gambar 11. Induksi dan seleksi kalus padi Nipponbare. a. Induksi kalus padi Nipponbare
b. Kalus padi pada media seleksi
Kalus yang berwarna putih berkembang semakin membesar, kemudian dipindahkan pada media regenerasi dan berkembang menjadi planlet yang memiliki tunas dan akar (Gambar 12a). Planlet selanjutnya dipindah ke media perakaran yang mengandung 40 mg/l higromisin dengan tujuan untuk memperkuat perakaran dan sebagai tahap akhir seleksi. Planlet yang akarnya berwarna putih pada media ini menunjukkan bahwa planlet tersebut benar-benar transgenik, sedangkan berwarna coklat merupakan planlet non transgenik yang lolos seleksi (Gambar 12b).
Gambar 12. Regenerasi kalus padi Nipponbare. a. Kalus padi dalam media regenerasi
b. Planlet padi dalam media perakaran
Jalur regenerasi tanaman yang digunakan dalam penelitian ini adalah embriogenesis somatik yang memiliki keuntungan bahwa tanaman transgenik yang dihasilkan berasal dari satu sel tunggal yang tertransformasi, sehingga mencegah terjadinya khimera. Hal ini berbeda dengan jalur organogenesis, dimana tanaman transgenik berasal dari kumpulan sel yang sebagian tertransformasi dan sebagian tidak.
a b
Jumlah kalus embriogenik yang dihasilkan untuk setiap periode transformasi berbeda, demikian juga untuk jumlah planlet maupun tanaman putatif transgenik. Kalus embriogenik yang dapat menjadi plantlet dan akhirnya menjadi tanaman transgenik pada semua periode transformasi cenderung menurun. Jumlah tanaman transgenik yang dihasilkan berkisar 9,5 – 17,5% (Tabel 1), variasi ini mungkin disebabkan oleh kualitas eksplan yang digunakan.
Hasil amplifikasi PCR semua sampel DNA padi transgenik yang dianalisis menggunakan primer HPTII (Gambar 6) menunjukkan bahwa semua sampel yang dianalisis menghasilkan fragmen atau pita DNA berukuran 500 bp. Kaset pr35S::HPTIIT35S terletak lebih dekat dengan Left Border dibanding kaset pr35S::OsDREB1A::T35S pada plasmid biner yang ditransformasi ke kalus padi. Karena integrasi T-DNA dimulai dari Right Border dan diakhiri pada Left Border, apabila kaset pr35S::HPTII::T35S terintegrasi dalam genom padi maka kemungkinan besar kaset pr35S::OsDREB1A::T35S juga sudah terintegrasi dalam genom, kecuali terjadi delesi yang cukup panjang mulai dari Right Border sampai kaset pr35S::OsDREB1A::T35S yang kemungkinannya sangat kecil. Pada tanaman transgenik yang memberikan hasil amplifikasi yang diharapkan pada PCR menggunakan primer HPTII kemungkinan besar juga mengandung 35S::OsDREB1A.
Simpulan
1. Gen 35S::OsDREB1A dapat ditransformasikan ke genom padi Japonica Nipponbare dengan A. tumefaciens dan seleksi higromisin.
2. Diperoleh 81 genotipe padi Nipponbare transgenik yang mengandung gen OsDREB1A berdasarkan analisis PCR menggunakan primer HPTII
3. Secara umum tanaman padi putatif transgenik generasi T0 menunjukkan kekerdilan dan keragaan yang lebih jelek untuk karakter-karakter komponen hasil dibanding tanaman padi Nipponbare non-transgenik.
Daftar Pustaka
Dubouzet JG, Sakuma YZY, Ito YY, Kasuga M, Dubouzet EG, Miura ZS, Seki M, Shinozaki K, and Shinozaki KY. 2003. OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high-salt- and cold-responsive gene expression. Plant 33: 751–763
Gilmour SJ, Zarka DG, Stockinger EJ, Salazar MP, Houghton JM, and Thomashow MF. 1998. Low temperature regulation of the Arabidopsis CBF family of AP2 transcriptional activators as an early step in cold-induced COR gene expression. Plant J. 16: 433–442
Greco R, Ouwerkerk PBF, Taal AJC, Favalli C, Beguiristan T, Puigdomenech P, Colombo L, Hoge JHC, Pereira A. 2001. Early and multiple A transposition in rice suitable for efficient insertion mutagenesis. Plant Molecular Biology. 67(1): 16-37.
Hsieh TT, Lee J-t, Chang Y-y, and Chan MT. 2002. Tomato Plants Ectopically Expressing Arabidopsis CBF1 Show Enhanced Resistance to Water Deficit Stress.Plant Physiology 130: 618–626.
Ito Y, Katsura K, Maruyama K, Taji T, Kobayashi M, Seki M, Shinozaki K, and Yamaguchi-Shinozaki K. 2006. Functional Analysis of Rice DREB 1/CBF-type Transcription Factors Involved in Cold-responsive Gene Expression in Transgenic Rice. Plant Cell Physiol. 47(1):141-163
Liu Q, Kasuga M, Sakuma Y, Abe H, Miura S, Yamaguchi-Shinozaki K, Shinozaki K. 1998. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transcription pathways in drought- and low-temperature-responsive gene expression, recspectively, in arabidopsis. Plant Cell 10: 1931-1406.
Mulyana NS. 2010. Over-ekkspresi gen OsDREB1A (Oryza sativa Dehydration-Responsive Element Binding protein 1A) untuk perbaikan sifat toleran kekeringan pada tanaman tembakau (Nicotiana tabacum L.) (skripsi). Universutas Gajah Mada.
Oh, S-J, Song SI, Kim YS, Jang H-J, Kim SY, Kim M, Kim Y-K, Nahm BH, and Kim J-K. 2005. Arabidopsis CBF3/DREB1A and ABF3 in transgenic Rice increased tolerance to abiotic stress without stunting growth. Plant Physiology Preview 11 p.
Pellegrineschi A, Reynolds M, Pacheco M, Brito RM, Almeraya R, Yamaguchi-Shinozaki K, and Hoisington D. 2004. Stress-induced expression in wheat of the Arabidopsis thaliana DREB1A gene delays water stress symptoms under greenhouse conditions. Genome 47:493-500.
Shinozaki K, Yamaguchi-Shinozaki K. 2004. Plant response to stress: regulation of plant gene expression to drought. Encyclopedia of Plant and Crop Science. DOI:10.1081/E-EPCS 120010660, Marcel Dekker, Inc. New York. Pp 99-101.
Shure M, Wessler S, and Fedoroff N. 1983. Molecular identification and isolation of the Waxy locus in Maize. Cell 35:225-233.
Abstrak
Tanaman padi transgenik toleran terhadap cekaman kekeringan merupakan salah satu alternatif yang diharapkan dapat mengatasi penurunan produksi padi. Keberhasilan transformasi menggunakan gen OsDREB1A pada tanaman padi genotipe Nipponbare memerlukan konfirmasi lebih lanjut menyangkut toleransi terhadap cekaman kekeringan. Penelitian ini bertujuan untuk mengevaluasi toleransi padi transgenik cv. Nipponbare 35S::OsDREB1A terhadap cekaman kekeringan. Evaluasi toleransi kekeringan menggunakan metode Standard System Evaluation for Rice (IRRI 1996). Hasil evaluasi diperoleh lima genotipe transgenik yang toleran terhadap kekeringan. Hasil karakterisasi molekuler menggunakan hibridisasi Southern menunjukkan bahwa lima genotipe transgenik memiliki 2 – 4 kopi transgen. Studi fisiologi mengindikasikan bahwa daya adaptasi tanaman transgenik 35S::OsDREB1A kemungkinan melibatkan mekanisme penghindaran kekeringan atau toleran kekeringan.
Kata kunci: evaluasi, padi transgenik, gen OsDEB1A, southern hibridisasi
Abstract
Transgenic rice plants tolerant to drought stress is one alternative which is expected to overcome the decrease in rice production. The success of the transformation using OsDREB1A gene in rice genotype Nipponbare requires further confirmation of tolerance to drought stress. The aim of this study was to evaluate the tolerance of the transgenic rice cv. Nipponbare 35S::OsDREB1A to drought stress. The evaluation of droug ht tolerance was performed using Standard System Evaluation for Rice method (IRRI 1996). Five transgenic genotypes tolerant to drought were selected from the evaluation. Molecular characterization using Southern hybridization showed that the five genotipes had 2-4 copies of transgene. Physiological analysis of the transgenic 35S::OsDREB1A plants indicated that the adaptability to drought of the transgenic plants might involve drought avoidance mechanisms.
Pendahuluan
Kekeringan hampir terjadi setiap tahun di daerah pertanian padi, baik di lahan sawah maupun di lahan tadah hujan. Hal ini disebabkan oleh pendeknya musim hujan, penanaman yang terlambat, dan pengairan tanaman yang bergantung pada hujan. Kekeringan dapat menghambat pertumbuhan dan menurunkan hasil padi, bahkan sampai menyebabkan puso. Kekeringan adalah suatu kondisi kekurangan air pada tanaman pada stadia tertentu yang dapat mempengaruhi pertumbuhan dan perkembangan tanaman (Suardi, 1988). Tanaman yang mengalami cekaman kekeringan menunjukkan pertumbuhan yang lebih lambat dari tanaman kontrol (Degenkolbe et al 2009). Keterlambatan saat berbunga dari genotipe yang tumbuh di lingkungan tercekam kekeringan akan memperpendek fase pengisian biji. Cekaman kekeringan pada pertanaman padi gogo yang terjadi pada saat pembungaan akan meningkatkan persentase kehampaan malai dan menurunkan hasil gabah. Di rumah kasa pada lingkungan tercekam kekeringan, hasil padi menurun 52,3% dibanding lingkungan normal (Samaullah dan Darajat, 2001).
Mekanisme adaptasi terhadap kekeringan ada tiga yaitu: 1. Drought escape ialah kemampuan tanaman untuk menyelesaikan satu siklus hidup sebelum tanaman mengalami kekeringan, 2. Drought avoidance ialah kemampuan tanaman untuk memelihara potensial air tetap tinggi dengan menyerap air terus kepucuk, atau kemampuan mempertahankan kehilangan air sekecil-kecilnya misalnya akar menjadi lebih panjang, stomata menutup, 3. Drought tolerance ialah kemampuan tanaman untuk bertahan hidup pada kondisi kekurangan air dengan jaringan potensial air yang rendah (Mitra 2001).
kekeringan, kadar garam tinggi dan suhu rendah (Dubouzet et al 2003; Ito et al. 2006).
Evaluasi untuk cekaman kekeringan pada tanaman padi dapat dilakukan di lapangan dalam rumah kasa atau kaca, dan laboratorium, ketika tanaman berada pada fase vegetatif, generatif, maupun keduanya. Sammaullah dan Darajat (2001) melakukan evaluasi beberapa genotipe padi di lapangan maupun rumah kasa menggunakan pot plastik berisi 10 kg tanah. Oh et al. (2005) mengevaluasi padi transgenik yang mengandung gen Ubi:CBF3 dan Ubi:ABF3 dengan menggunakan pot berukuran 5cm x 5cm x 6 cm di rumah kaca. Ai et al. (2010) menggunakan media cair PEG di laboratorium pada fase perkecambahan padi ketika melakukan evaluasi indikator toleransi cekaman kekeringan
Materi tanaman yang digunakan dalam penelitian ini adalah genotipe putatif transgenik yang mengandung gen OsDREB1A dengan promotor 35S. Gen ini merupakan gen regulator yang responsif terhadap suhu rendah dan over-ekspresinya dapat meningkatkan toleransi cekaman kekeringan dan atau suhu rendah (Kasuga et al. 2004).
Tujuan Penelitian
Untuk mengevaluasi toleransi padi transgenik cv. Nipponbare 35S::OsDREB1A terhadap cekaman kekeringan.
Metodologi Penelitian
A. Seleksi Tanaman Transgenik Hasil Transformasi terhadap Cekaman Kekeringan
Bahan Tanaman
Genotipe padi transgenik Nipponbare 35S::OsDREB1A generasi T1 yaitu B9, B11, B14, C3, C4, C9, C10, C12, dan C14. Genotipe-genotipe tersebut merupakan hasil seleksi pada media 100 mg/ l higromisin. Genotipe kontrol yaitu Kalimutu (toleran), Singkarak (peka) (Hermanto et al. 2009), dan Nipponbare. Pelaksanaan Penelitian
Juli sampai dengan September 2010. Rancangan percobaan sederhana setiap pot berisi lima tanaman padi.
Evaluasi terhadap Cekaman Kekeringan
Benih padi transgenik dari genotipe-genotipe yang terpilih, Nipponbare, Kalimutu dan Singkarak dikecambahkan pada petridis yang telah dilapisi kertas saring dan diberi air secukupnya. Benih padi transgenik dan Nipponbare yang telah pecah kulit dipindahkan ke media cair yang mengandung 100 mg/l higromisin selama 10 hari dengan tujuan mendapatkan bibit tanaman padi yang tahan terhadap higromisin. Bibit dipindahkan pada media air selama satu hari sebelum ditanam pada media tanah. sebanyak 5 tanaman ditanam untuk tiap pot. Pot (diameter 13 cm, tinggi 11 cm) diisi media tanam 700 g dengan komposisi : tanah + pasir + pupuk kompos dengan perbandingan= 1 : 1 : 1. Penyiraman dilakukan satu atau dua hari sekali sebanyak 750 ml / 10 kg tanah (Samaullah dan Daradjat 2001) sampai tanaman siap untuk uji kekeringan yaitu pada umur sekitar tiga - empat minggu. Perlakuan cekaman kekeringan diberikan dengan cara menghentikan penyiraman sampai tanaman peka (Singkarak) menunjukkan gejala layu dan dilakukan pengamatan pertama (daun menggulung pada fase vegetatif). Pengamatan selanjutkan dilakukan pada saat tanaman kontrol peka (Singkarak) mati atau semua daun mengering (daun mengering pada fase vegetatif) . Setelah selesai pengamatan kemudian dilakukan pemulihan tanaman dengan cara penyiraman. Tanaman yang masih tumbuh kemudian dilakukan penilaian kesembuhan. Penilaian skor penggulungan daun, pengeringan daun, dan kesembuhan tanaman dilakukan dengan berdasarkan skor pengamatan seperti yang dijelaskan pada Standard Evaluation System for Rice (IRRI 1996, Silitonga et al. 2003) seperti tertera pada Tabel 2–4 :
Tabel 2. Klasifikasi tanggap tanaman terhadap kekeringan (daun menggulung pada fase vegetatif)
Skor/skala Kriteria
0 Daun-daun sehat
Tabel 3. Klasifikasi tanggap tanaman terhadap kekeringan (daun mengering pada fase vegetatif)
Skor/skala Keterangan
0 Tidak ada gejala
1 Ujung daun sedikit mengering
3 Ujung daun mengering sampai ¼ panjang pada hampir semua daun
5 ¼ sampai ½ dari semua daun betul-betul kering
7 Lebih dari ⅔ dari semua daun betul-betul kering
9 Tanaman mati
Tabel 4. Klasifikasi tanggap kesembuhan tanaman setelah perlakuan pemulihan
Skor/skala Keterangan
1 90-100% tanaman sembuh
3 70-89% tanaman sembuh
5 40-69% tanaman sembuh
7 20-39% tanaman sembuh
9 0-19% tanaman sembuh
Isolasi DNA
Metode Dellaporta et al. (1983) yang sudah dimodifikasi digunakan untuk isolasi DNA. Daun padi yang masih muda dipotong kecil-kecil dimasukkan ke dalam ependorf dua ml dan diberi gotri (pelor), kemudian dimasukkan ke dalam nitrogen cair dan divorteks sampai halus. Serbuk daun diberi larutan buffer ekstraksi DNA yang sudah ditambah Na-disulfit (0,38 g/100ml) sebanyak 700 µl. Selanjutnya tabung dipanaskan dalam water bath pada suhu 65°C selama lima menit. Untuk memisahkan asam nukleat dengan komponen lain ditambahkan kloroform : isoamyl alcohol (Chisam 24:1) dengan perbandingan 1 : 1, kemudian digoyang atau dikocok selama lebih kurang 50 kocokan, selanjutnya disentrifuge pada 12,000 rpm selama lima menit. Supernatan dipindahkan ke tabung ependorf 2 ml yang baru, kemudian ditambahkan ethanol absolut (98%) dengan perbandingan supernatant : etanol absolut = 1 : 2 dan dicampur dengan membalik-balikkan tabung. Selanjutnya tabung disentrifuse selama lima menit. Endapan DNA dicuci dengan etanol 70% dua kali, kemudian dikeringkan dalam pompa vakum. DNA dilarutkan dalam Tris-EDTA (TE) buffer sesuai dengan besar kecilnya endapan.
Amplifikasi PCR
dimasukkan ke dalam tabung PCR, kemudian diamplifikasi dengan mesin PCR pada program Hyg-60. Profil suhu yang digunakan adalah predenaturasi 94 oC
selama 5 menit, dilanjutkan dengan 35 siklus meliputi pemisahan (denaturation) pada suhu 94oC selama 1 menit, penempelan primer (annealing) pada suhu 55oC selama 1 menit, pemanjangan primer (extension) pada suhu 72 oC selama 2 menit. Pada tahap akhir PCR dilakukan pemanjangan akhir pada 72 oC selama 5 menit. Hasil amplifikasi dicek dengan elektrofloresis gel agarose 0,8%. Delapan µl DNA + 1 µl blue juice dicampur kemudian dimasukkan ke dalam sumur gel. Separasi dilakukan pada voltase 80 – 100 V selama sekitar satu jam. Gel direndam dalam larutan 0,5 g/ml Ethidium Bromide dan pita DNA divisualisasikan menggunakan Chemidoc.
Hhibridisasi Southern
Analisis hibridisasi southern yang digunakan dalam penelitian ini berdasarkan metode Trijatmiko et al. (2011) dengan prosedur pelaksanaan sebagai berikut.
Pemotongan DNA. DNA sampel transgenik generasi T1 dan kontrol non-transgenik dipotong menggunakan enzim EcoRI yang memotong sekali di dalam T-DNA. Total volume untuk masing-masing reaksi adalah 150 μl dalam ependrof steril 1.5 ml (10 ug DNA, menggunakan 3 μl enzim restriksi 10 U/ μl) dan reaksi diinkubasi semalam pada suhu 37oC. Pada hari berikutnya, setiap sampel diambil 5 µl dan di elekrofloresis pada gel untuk memastikan bahwa DNA telah terpotong sempurna. Jika pemotongan telah baik akan terlihat smer yang homogen pada jalur. Jika DNA tidak terpotong sempurna, ditambahkan enzim lagi dan dilanjutkan pemotongan.
menit. Pelet dilarutkan dalam 15 µl 0,1 x TE dan 3 µl 6x ficoll loading dye dan dinkubasi pada suhu 65oC selama 15 menit. Tabung disentil-sentil setiap lima menit untuk melarutkan DNA.
Pemotongan Kontrol Positif. Pemotongan juga dilakukan pada 200 pg plasmid biner sebagai kontrol positif.
Elektroporesis gel agarose. Ditambahkan 343 ml ddH2O dalam botol 500 ml dan 2,45 agarose selanjutnya dipanaskan dalam microwave sampai larut. Ditambahkan 7 ml 50x TAE dalam botol dan dicampur dengan stirre. Setelah cukup dingin gel dituangkan ke dalam cetakan. Sampel (18 µl) dan MW markers (5 µl) diloading, selanjutkan di elektroforesis pada 30 volt semalam. Pada hari berikutnya voltase diturunkan menjadi 15 volt.
Prosesing gel. Gel dipotong untuk blotting dengan ukuran ( 20 cm x 10,5 cm ). Gel direndam pada larutan depurinasi dan digoyang selama 10 menit pada suhu ruang. Gel dicuci dengan air distilat ganda, selanjutknya gel direndam pada larutan denaturasi 2 x 15 menit pada suhu ruang dengan digoyang secara perlahan-lahan. Gel kemudian dicuci dengan air dobel distilat dan gel direndam pada larutan neutralisasi 2 x 15 menit pada suhu ruang. Selanjutnya gel direndam dalam 20 x SSC kurang lebih 10 menit.
permukaan kertas Whatman di atas kaca yang tidak tertutupi gel, begitu juga bagian bak yang terbuka, ditutup dengan plastik pembungkus. Kemudian ditambahkan kertas tissue, dan gelas pemberat sekitar 200-500 g. Transfer blot dilakukan semalam di dalam transfer buffer (20x SSC). Pada hari berikutnya, blot dibongkar dan membran secara hati-hati diambil selanjutnya direndam selama 5 menit dalam larutan 2x SSC. Membran dikeringkan di atas kertas tissue selama 3 menit. Membran dibungkus dengan plastik pembungkus dan DNA difiksasi dengan sinar UV selama 5 menit pada 302nm dalam transilluminator. Membran bisa langsung digunakan untuk prehibridisasi atau disimpan di dalam tempat lembab diantara ( 2 kertas whatman 3MM dan dibungkus dengan plastik pembungkus ) pada suhu – 20oC.
Labelling Menggunakan Polymerase Chain Reaction
Reagen yang diperlukan untuk setiap reaksi labelling (total volume 50 µl) adalah 29,25 µl dd H2O; 5 µl bufer PCR dengan MgCl2, 10 x konsentrasi; 5 µl PCR DIG mix, 10 konsentrasi; 2,5 µl primer 1 (5 µM); 2,5 µl primer 2 (5 µM); 0,75 µl enzim mix, expand high fidelity, 5 µl plasmid (20 pg/ µl). Disiapkan juga reaksi untuk amplifikasi (regular PCR menggunakan dNTPs) sebagai kontrol. Profil siklus suhu
1 siklus : 95oC selama 2 menit. 30 siklus : 95oC selama 30 detik, 60oC selama 30 detik, 72oC selama 40 detik. 1 siklus : 72oC selama 7 menit.
Setiap reaksi dielektroforesis ( 5 µl ) pada gel agarose bersama dengan marker DNA. Jika labelling berhasil, akan terlihat probe yang sudah berlabel akan bermigrasi lebih lambat (bobot molekul lebih besar). dibandingkan probe kontrol (disebabkan adanya DIG). Intensitas DNA yang sudah berlabel akan sama atau agak berkurang dibandingkan DNA kontrol yang tidak berlabel ( disebabkan adanya DIG).
Hibridisasi
dalam es selama 5 menit. Satu menit sebelum inkubasi probe dalam es selesai, 50 mL larutan DIG Easy Hyb dipindahkan dari bak plastik ke tabung 50 mL. Kemudian probe yang sudah didenaturasi ditambahkan ke tabung yang berisi larutan DIG Easy Hyb tersebut dan dicampur dengan baik untuk membentuk larutan hibridisasi. Selanjutnya larutan hibridisasi dimasukkan ke bak plastik dan dilakukan hibridisasi semalam pada suhu 42oC dengan penggoyangan.
Deteksi Menggunakan Chemiluminescent Reaction
Membran dicuci 2 x 5 menit dengan 200 ml washing 1 pada suhu kamar, selanjutnya 2 x 15 menit dengan 200 ml washing 2 pada suhu 68oC. Untuk langkah selanjutnya dilakukan pada suhu ruang. Membran diseimbangkan selama 2 menit dengan 200 ml buffer 1 + Tween. Kemudian membran diinkubasi selama 30 menit sampai 3 jam dengan 200 ml buffer 2. Selanjutnya menggunakan bak plastik baru, membran dicuci untuk membuang antibodi yang berlebihan dengan diinkubasi 2 x 15 menit pada 200 ml buffer 1 + Tween. Selanjutnya membran diseimbangkan selama 3 menit dengan larutan 40 ml bufer 3. Membran diletakkan (bagian DNA menghadap ke atas) di atas plastik. Tiga ml Ready-to-use CDP-Star diteteskan pada permukaan blot sampai seluruh permukaan basah. Kemudian membran dibungkus dengan plastik pembungkus. Membran diinkubasi selama 5 menit pada suhu ruang. Cairan yang berlebih dalam plastik wrap dibuang. Selanjutnya membran dimasukkan dalam kaset dan film X-ray dimasukkan ke dalam kaset di ruang gelap. Film X-ray direndam selama 1 menit di dalam larutan developer dengan digoyang-goyang, 30 detik di dalam air, dan 2 menit dalam larutan fiksasi. Selanjutnya film X-ray dibilas dengan air ledeng dan dikering-anginkan.
B. Studi Fisiologi Gen OsDREB1A Bahan Tanaman
Genotipe padi Nipponbare transgenik B11, C10, Nipponbare, Kalimutu (kontrol toleran) dan Singkarak (kontrol peka).
Pelaksanaan Penelitian
Pertanian, Bogor. Penelitian dilaksanakan dari bulan November 2011 sampai dengan Februari 2012.
Studi Fisiologi
Biji padi dikecambahkan pada petridis yang telah diberi air. Benih yang telah berkecambah ditanam pada botol yang berisi air sampai menjadi bibit tanaman kemudian ditanam pada pot (diameter 23 cm dengan tinggi 11 cm) yang berisi tanah + pasir + pupuk kompos (2:1:1) sebanyak 4,5 kg. Setiap pot berisi 3 tanaman padi dan disiram setiap hari sebanyak 750 ml/10 kg media tanam (Samaullah dan Daradjat 2001). Pada waktu dilakukan pengujian kekeringan setiap pot berisi 1 tanaman padi. Pengeringan dilakukan pada umur 40 hari setelah tanam. Setiap hari, ditimbang bobot total setiap pot untuk mengetahui besarnya evapotranspirasi sampai ada tanaman kontrol peka (Singkarak) yang mati. Pada pagi hari berikutnya dilakukan penimbangan bobot total setiap pot untuk mengetahui bobot awal sebelum dilakukan penyiraman lagi. Kemudian dilakukan penyiraman kembali untuk pemulihan. Setelah mengalami evapotranspirasi, bobot total setiap pot ditimbang dan tanaman disiram kembali sesuai dengan banyaknya air yang hilang selama evapotranspirasi. Peubah yang diamati : jumlah stomata, bobot tajuk, bobot akar, daun hijau (%).
Pengamatan stomata dilakukan dengan cara mengolesi permukaan daun padi dengan kutek/ cat kuku yang transparan. Setelah kutek kering kemudian diambil dengan selotip dan ditempelkan pada gelas preparat selanjutnya diamati di bawah mikroskop dengan perbesaran 40 x dan dihitung jumlah stomata tiap bidang pandang. Penghitungan dilakukan pada 3 bidang pandang yang berbeda. Berikut rumus penghitungan stomata:
Kerapatan stomata
Diameter bidang pandang = 0,5 mm.
Luas bidang pandang = d2
Isolasi DNA
nitrogen cair dan divorteks sampai halus. Serbuk daun diberi larutan buffer ekstraksi DNA yang sudah ditambah Na-disulfit (0,38 g/100ml) sebanyak 700 µl. Selanjutnya tabungdipanaskan dalam penangas air pada suhu 65°C selama lima menit. Untuk memisahkan asam nukleat dengan komponen lain ditambahkan kloroform : isoamyl alcohol (Chisam 24:1) dengan perbandingan 1 : 1, kemudian digoyang atau dikocok selama lebih kurang 50 kocokan, selanjutnya disentrifuge pada 12,000 rpm selama lima menit. Supernatan dipindahkan ke tabung ependorf 2 ml yang baru, kemudian ditambahkan ethanol absolut (98%) dengan perbandingan supernatant : etanol absolut = 1 : 2 dan dicampur dengan membalik-balikkan tabung. Selanjutnya tabung disentrifuse selama lima menit. Endapan DNA dicuci dengan etanol 70% dua kali, kemudian dikeringkan di dalam pompa vakum. DNA dilarutkan di dalam Tris-EDTA (TE) buffer sesuai dengan besar kecilnya endapan.
Amplifikasi PCR
Campuran reaksi (2 µl buffer 10 x PCR;1,2 µl MgCl; 0,4 µl DNTPs; 2 µl Primer HPTII; 0,16 µl Taq DNA; 9,24 µl ddH2O; dan 5 µl sampel DNA) dimasukkan di dalam tabung PCR, kemudian diamplifikasi dengan mesin PCR pada program Hyg-60. Profil suhu yang digunakan adalah predenaturasi 94 oC selama 5 menit, dilanjutkan dengan 35 siklus meliputi pemisahan (denaturation) pada suhu 94oC selama 1 menit, penempelan primer (annealing) pada suhu 60oC selama 1 menit, pemanjangan primer (extension) pada suhu 72 oC selama 2 menit. Pada tahap akhir PCR dilakukan pemanjangan akhir pada 72 oC selama 5 menit. Hasil amplifikasi dicek dengan elektrofloresis gel agarose 0,8%. Delapan µl DNA + 1 µl blue juice dicampur kemudian dimasukkan ke dalam sumur gel. Separasi dilakukan pada voltase 80 – 100 V selama sekitar satu jam. Gel direndam dalam larutan 0,5 g/ml Ethidium Bromide dan pita DNA divisualisasikan menggunakan Chemidoc.
Hasil Penelitian
A. Evaluasi Toleransi Tanaman Transgenik terhadap Kekeringan
kontrol (Nipponbare, Batutegi dan Singkarak) memiliki skor 9 yaitu mati semua. Tanaman padi transgenik untuk genotipe yang lain memiliki skor kesembuhan berkisar antara 3 – 7, berarti tanaman masih ada yang hidup dan dapat tumbuh dengan baik. Tanaman padi transgenik generasi T1 ini masih mengalami segregasi yang menyebabkan nilai skor daun menggulung, dan daun mengering bervariasi.
Tabel 5. Skor hasil pengujian kekeringan tanaman padi transgenik (T1) dan tanaman non-transgenik pada fase vegetatif di rumah kaca Fasilitas Uji Daun menggulung Daun mengering
B9 5 9 9 9
*= dua pot. skor 3 = 70-89% tanaman sembuh, skor 5 = 40-69% tanaman sembuh, skor 7 = 20-39% tanaman sembuh, skor 9 = 0-19% tananam sembuh.
Gambar 13. Hasil analisis PCR dan hibridisasi Southern sampel DNA padi Nipponbare transgenik
Kontrol= genotipe Nipponbare tipe liar
a. Hasil amplifikasi PCR enam genotipe padi transgenik menggunakan primer HPTII b. Profil hibridisasi Southern lima genotipe padi transgenik
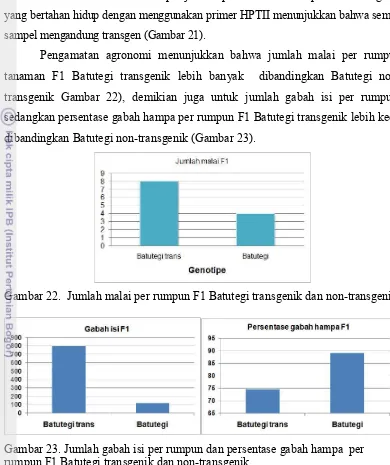
Dari delapan genotipe transgenik yang lebih toleran terhadap kekeringan
dibanding Nipponbare non-transgenik ini dipilih 6 genotipe yang memiliki jumlah
B11, B14, C10, C14, C12, dan C3. Genotipe-genotipe tersebut kemudian dianalisis
menggunakan PCR. Hasil analisis menunjukkan bahwa lima genotipe menghasilkan
pita dengan ukuran yang diharapkan, sementara satu genotipe yaitu C12 tidak
menghasilkan pita DNA (Gambar 13a dan Tabel 6). Lima genotipe tersebut
selanjutnya dianalisis menggunakan hibridisasi Southern. Hasil analisis hibridisasi
terhadap lima sampel DNA menunjukkan bahwa kelima genotipe memiliki 2 – 4
salinan transgen (Gambar 13b dan Tabel 6).
Tabel 6. Hasil analisis PCR enam genotipe dan hibridisasi Southern lima genotipe padi transgenik
Genotipe
PCR Southern hibridisasi
+/- Jumlah pita B14 + + 2 B11 + + 2 C14 + + 4 C10 + + 2 C12* -
C3 + + 4
*tidak dilakukan analisis hibridisasi Southern
Gambar14. Penampilan tanaman padi Nipponbare transgenik generasi T1 setelah uji kekeringan
B. Studi Fisiologi Gen OsDREB1A
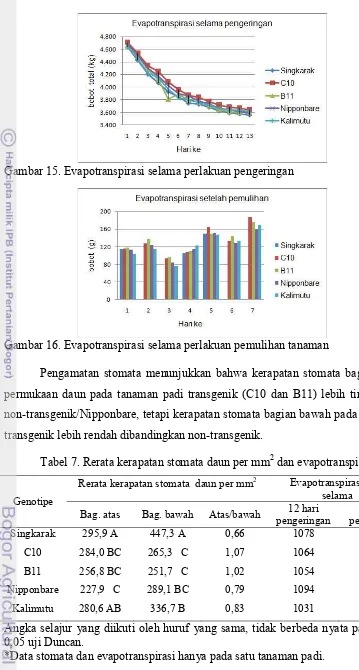
Gambar 15. Evapotranspirasi selama perlakuan pengeringan
Gambar 16. Evapotranspirasi selama perlakuan pemulihan tanaman
Pengamatan stomata menunjukkan bahwa kerapatan stomata bagian atas permukaan daun pada tanaman padi transgenik (C10 dan B11) lebih tinggi dari non-transgenik/Nipponbare, tetapi kerapatan stomata bagian bawah pada tanaman transgenik lebih rendah dibandingkan non-transgenik.
Tabel 7. Rerata kerapatan stomata daun per mm2 dan evapotranspirasi
Genotipe
Rerata kerapatan stomata daun per mm2 Evapotranspirasi (ml) selama
Bag. atas Bag. bawah Atas/bawah 12 hari
pengeringan
7 hari pemulihan
Singkarak 295,9 A 447,3 A 0,66 1078 1055
C10 284,0 BC 265,3 C 1,07 1064 1085
B11 256,8 BC 251,7 C 1,02 1054 920
Nipponbare 227,9 C 289,1 BC 0,79 1094 835
Kalimutu 280,6 AB 336,7 B 0,83 1031 1060
Angka selajur yang diikuti oleh huruf yang sama, tidak berbeda nyata pada taraf 0,05 uji Duncan.
Secara statistik rerata kerapatan stomata daun per mm2 pada permukaan atas dan bawah daun antara genotipe transgenik (C10 dan B11) dengan non-transgenik (Nipponbare) tidak berbeda nyata, namun berbeda nyata dengan Singkarak (kontrol peka). Evapotranspirasi selama perlakuan kekeringan dan selama pemulihan tanaman tidak berbeda nyata untuk semua genotipe (Tabel 7). Berdasarkan data kerapatan stomata mekanisme toleransi genotipe transgenik belum dapat disimpulkan.
Uji korelasi antara kerapatan stomata dengan evapotranspirasi selama pemulihan menunjukkan bahwa koefisien korelasi antara kerapatan stomata daun pada permukaan atas = 0,95602, sedangkan koefisien korelasi koefisien korelasi antara kerapatan stomata pada permukaan bawah dengan evapotranspirasi setelah pemulihan mempunyai koefisien korelasi (r2) = 0,40234.
Tabel 8. Persentase tanaman hidup dan rerata daun hijau pada waktu pemulihan Genotipe Tanaman hidup (%) Rerata Daun hijau (%)
Singkarak 80 34,44 B
C10 100 57,92 A
B11 100 58,20 A
Nipponbare 100 51,29 AB
Kalimutu 100 46,90 AB
Angka selajur yang diikuti oleh huruf yang sama, tidak berbeda nyata pada taraf 0,05 uji Duncan
Setelah pemulihan sebanyak 80% dari tanaman genotipe kontrol peka (Singkarak) yang hidup, sementara untuk genotipe yang lain tanaman yang hidup mencapai 100%. Rerata daun hijau tanaman padi transgenik (C10 dan B11) paling tinggi namun tidak berbeda nyata dengan Nipponbare dan Kalimutu, tetapi berbeda dengan Singkarak (Tabel 8).
mana genotipe C10 memiliki nilai yang kecil dan berbeda nyata dengan Nipponbare non-transgenik dan genotipe transgenik B11.
Tabel 9. Bobot basah dan kering tajuk, bobot basah dan kering akar, panjang akar, nisbah tajuk:akar, dan bobot kering tajuk/evapotranspirasi
Angka selajur yang diikuti oleh huruf yang sama, tidak berbeda nyata pada taraf 0,05 uji Duncan.
Gambar 17. Hasil analisis PCR sampel DNA tanaman padi transgenik Nipponbare, Singkarak dan Kalimutu dengan menggunakan primer HPTII. M= Marker, K- = Kontrol negatif, K+ = kontrol positif, I = genotipe C10, J = genotipe B11, Np = Nipponbare, Si = Singkarak, Kl = Kalimutu.
Hasil analisis PCR dengan primer HPTII pada generasi T1 menunjukkan bahwa pada genotipe padi transgenik B11 terdapat 1 tanaman T1 yang non-transgenik (J5) sementara 4 tanaman lainnya non-transgenik (J1-J4, Gambar 7). Hal ini menunjukkan bahwa masih terjadi segregasi pada genotipe B11.
Pembahasan
A. Seleksi Tanaman Transgenik Hasil Transformasi terhadap Kekeringan
Hasil pengamatan daun menggulung pada fase vegetatif pada hari ke 6 (Tabel 5) menunjukkan bahwa genotipe Singkarak (peka) dan genotipe B9 memiliki skor 9 (daun menggulung ketat), sedangkan Kalimutu (toleran) dan Nipponbare memiliki skor 7 (ujung-ujung daun bersentuhan). Delapan genotipe padi transgenik memiliki skor 0-9 (daun sehat – daun menggulung ketat). Hasil pengamatan daun mengering pada fase vegetatif pada hari ke 10 (Tabel 5) menunjukkan bahwa delapan genotipe transgenik memiliki skor 0-9 (tidak ada gejala – tanaman mati), sedang genotipe B9 dan semua tanaman kontrol memiliki skor 9. Tanaman padi transgenik genotipe B9 pada uji kekeringan mati semua walaupun telah lolos seleksi higromisin. Hal ini mungkin disebabkan gen OsDREB1A yang masuk dalam genom padi tidak pada posisi yang tepat atau tidak sempurna. Tanaman padi transgenik tersebut mungkin mengandung banyak gen sehingga terjadi pembungkaman gen. Variasi skor yang terjadi pada masing-masing dari delapan genotipe tanaman padi transgenik, kemungkinan disebabkan karena adanya segregasi gen OsDREB1A.
Hasil analisis PCR terhadap enam genotipe yang lolos uji kekeringan menunjukkan satu genotipe tidak terdapat pita DNA dan lima genotipe terdapat pita DNA (Gambar 13a, Tabel 6) berarti lima genotipe padi tersebut mengandung gen OsDREB1A yaitu B11, B14, C10, C14, dan C3. Genotipe C12 yang lolos uji kekeringan tetapi negatif PCR kemungkinan disebabkan oleh kekurang tegasan dalam mengambil keputusan selama seleksi perkecambahan dengan higromisin, di mana tanaman yang berkecambah namun memiliki akar coklat tetap diikutkan dalam uji toleransi kekeringan, padahal akar coklat merupakan salah satu tanda penting dari tanaman non-transgenik.
tidak dijumpai hubungan yang sistematis antara jumlah transgen yang dimiliki dengan tingkat toleransi kekeringan pada tanaman.
B. Studi Fisiologi Gen OsDREB1A
Pengamatan pada evapotranspirasi selama perlakuan kekeringan, menunjukkan bahwa rerata bobot total untuk semua genotipe mengalami penurunan yang besarnya hampir sama pada suhu 23-36oC dengan kelembaban 46-54% (Gambar 15). Evapotranspirasi pada waktu pemulihan berdasarkan pengukuran harian untuk semua genotipe hampir sama pada suhu 25-35oC dengan kelembaban 47-54%, seperti terlihat pada Gambar 16. Kadar air berdasarkan analisis fisik tanah dengan pF 4.2 adalah 18,84% (titik layu permanen), sedangkan rerata kadar air tanah pada waktu akhir pengeringan untuk genotipe Singkarak, C10, B11, Nipponbare, dan Kalimutu masing-masing adalah 14,78%, 14,86%, 17%, 16,48%, dan 14,26%. Jadi data kadar air tanah tersebut menunjukkan bahwa semua tanaman sudah berada dalam kondisi cekaman kekeringan. Berdasarkan pengamatan pada hasil evapotranspirasi pada saat pengeringan maupun pemulihan setelah perlakuan pengeringan, mekanisme toleransi dari gen OsDREB1A belum dapat diketahui.
Pada uji korelasi antara jumlah stomata dengan evapotranspirasi selama pemulihan, jumlah stomata daun pada permukaan atas mempunyai koefisien korelasi (r2) = 0,95602, sedangkan untuk jumlah stomata permukaan bawah mempunyai koefisien korelasi (r2) = 0,40234. Hal ini menunjukkan bahwa diduga jumlah stomata bagian atas lebih berpengaruh terhadap evapotranspirasi dibandingkan bagian bawah.
Pengamatan tanaman setelah perlakuan kekeringan menunjukkan bahwa 80% dari total tanaman genotipe Singkarak (peka) dapat pulih dengan rerata daun hijau sebanyak 34,44%, sedangkan genotipe lainnya pulih 100%. Genotipe padi transgenik C10 dan B11 mempunyai rerata daun hijau lebih tinggi dibandingkan genotipe kontrol (Nipponbare tipe liar), meskipun tidak berbeda nyata secara statistik
(Tabel 8). Rerata daun hijau yang lebih tinggi ini mungkin terkait dengan kemampuan
tanaman untuk melakukan proteksi terhadap sel dan jaringan dalam menghadapi produksi
yang berlebihan dari senyawa-senyawa yang merusak seperti reactive oxygen species
(ROS) yang terbentuk selama cekaman kekeringan. Hal ini mungkin mengindikasikan
keterlibatan mekanisme toleransi kekeringan (drought tolerance) pada tanaman
transgenik 35S::OsDREB1A.
Hasil pengamatan terhadap beberapa karakter agronomi tanaman (Tabel 9) terdapat perbedaan terhadap bobot basah tajuk, bobot kering akar dan nisba tajuk : akar. Tanaman transgenik genotipe C10 memiliki nilai nisba tajuk:akar yang secara nyata lebih kecil dari tanaman transgenik B11 dan Nipponbare non-transgenik. Nilai nisba tajuk:akar yang kecil ini biasanya terkait dengan mekanisme penghindaran kekeringan yang lebih baik.
Simpulan
1. Dihasilkan lima genotipe padi Nipponbare transgenik yang lebih toleran terhadap kekeringan dibandingkan genotipe Nipponbare non-transgenik. 2. Lima genotipe padi Nipponbare transgenik memiliki 2 – 4 salinan
transgen.
3. Daya adaptasi terhadap kekeringan dari genotipe Nipponbare transgenik yang mengandung gen 35S::OsDREB1A diduga melibatkan mekanisme penghindaran kekeringan atau toleran kekeringan.
Daftar Pustaka
Ai NS, Tondais SM, Butarbutar R. 2010. Evaluasi indikator toleransi cekaman kekeringan pada fase perkecambahan padi (Oryza sativa L.). J. Biologi XIV(1): 50-54
Degenkolbe T, Do PT, Zulther E, Repsiber D, Walther D, Hincha DK, Kohl KI. 2009. Expression profilling of rice cultiars differing in their tolerabce to long-term drought stress. Plant Mol Biol 69:133-153.
Dellaporta SL, Wood J, Hicks JB. 1983. A plant DNA minipreparation: version II. Plant Molecular Biology Reporter 1(4): 19-21.
Dubouzet JG, Sakuma Y, Ito Y, Kasuga M, Dubouzet EG, Miura S, Seki M, Dubouzet KSh, Miura S, Seki M, Shinozaki K, and Yamaguchi-Shinozaki K. 2003. OsDREB genes in rice, Oryza sativa L., encode transcription activators that functiion in drought-, high-salt-and cold-responsive gene expression. The Plant J. 33: 751-763
Hermanto, Sadikin DW, dan Hikmat E. 2009. Deskripsi Varietas Unggul Padi 1943-2009. Pusat Penelitian dan Pengembangan Tanaman Pangan. 220 h.
International Rice Research Institute. 1996. Standard evaluation system for rice. 4th ed, IRTO, IRRI. Los Banos, Philippines. 54p.
Ito Y, Katsura K, Maruyama K, Taji T, Kobayashi M, Seki M, Shinozaki K, and Yamaguchy-Shinozaki K. 2006. Functional Analysis of Rice Gene Expression in Transgenic Rice. Plant Cell l 47(1):141-153
Mitra J. 2001. Genetics and genetic improvement of drought resistance in crop plants. Current Sci. 80(6): 758-762
Oh, S-J, Song SI, Kim YS, Jang H-J, Kim SY, Kim M, Kim Y-K, Nahm BH, and Kim J-K. 2005. Arabidopsis CBF3/DREB1A and ABF3 in transgenic Rice increased tolerance to abiotic stress without stunting growth. Plant Physiology Preview 11 p.
Sammaullah MJ, dan Darajat AA. 2001. Toleransi beberapa Genotipee Padi Gogo terhadap Cekaman Kekeringan. Penelitian Pertanian Tanaman Pangan 20(1):17-23.
Silitonga TS, IH Somantri, AA Daradjat, H Kurniawan. 2003. Panduan Sistem Karakterisasi dan Evaluasi Tanaman Padi. Departemen Pertanian. Badan Penelitian dan Pengembangan Pertanian. Komisi Nasional Plasma Nutfah. 58h.
Suardi D. 1988. Ketahanan tanaman padi terhadap kekeringan. Bulletin Penelitian Balai Penelitian Tanaman Pangan Bogor. 29-37
Suardi D, TS Silitonga, B Abdullah, dan T Suhartini. 2004. Evaluasi Spesies Padi Liar Toleran Kekeringan. Penelitian Pertanian Tanaman Pangan 23(1): 23-27.
PEMINDAHAN GEN 35S::OsDREB1A DARI PADI TRANSGENIK KE VARIETAS ELIT PADI MELALUI PERSILANGAN
Abstrak
Genotipe padi toleran terhadap cekaman kekeringan sangat diperlukan untuk daerah-daerah yang mengalami kekeringan agar tetap dapat berproduksi. Telah dihasilkan tanaman padi Nipponbare transgenik yang mengandung gen 35S::OsDREB1A dan toleran terhadap cekaman kekeringan. Namun gen tersebut belum diketahui efeknya terhadap toleransi kekeringan ketika dipindahkan ke varietas padi elit. Tujuan penelitian ini adalah untuk mengetahui efek gen 35S::OsDREB1A terhadap toleransi kekeringan pada varietas elit padi Indonesia setelah dipindahkan melalui persilangan. Bahan persilangan yang menggunakan adalah genotipe padi Nipponbare transgenik sebagai tetua donor, dan varietas elit Ciherang, Batutegi, Code dan Konawe sebagai tetua penerima. Hasil penelitian menunjukkan bahwa genotipe padi F1 elit transgenik yang diturunkan dari Batutegi, Code, dan Konawe yang toleran terhadap cekaman kekeringan dan positif mengandung transgen berdasarkan analisis PCR. Tanaman BC1F1 turunan dari tiga varietas elit tadi juga positif mengandung transgen berdasarkan analisis PCR. Hasil ini menunjukkan bahwa gen 35S::OsDREB1A dapat dipindahkan dari Nipponbare transgenik ke tanaman padi elit melalui persilangan.
Kata kunci: pemindahan, gen OsDREB1A, padi elit, silangbalik
Abstract
Rice genotypes tolerant to drought stress are require for areas that experiencing drought in order to maintain the production. Transgenic rice cv. Nipponbare containing 35S::OsDREB1A and tolerant to drought stress have been generated. However, the effect of the gene had not yet been tested in the Indonesian elite rice genotypes. The purpose of this research was to assess the effect of 35S::OsDREB1A gene in the Indonesian elite varieties after being transferred through crossbreeding. The 35S::OsDREB1A transgenic rice cv. Nipponbare was crossed with rice cv. Ciherang, Batutegi, Code and Konawe . The results showed that F1 plants derived from Batutegi, Code, and Konawe were tolerant to drought stress and contained the transgen as shown by PCR analysis. The BC1F1 plants derived from those three varieties were also contained transgen as shown by PCR analysis. These result showed that 35S::OsDREB1A gene can be transferred into elite rice genotypes through crossbreeding.
Pendahuluan
Padi adalah tanaman sereal yang bernilai sosial, politik dan ekonomi, karena merupakan bahan makanan pokok bagi lebih dari setengah penduduk dunia. Untuk mempertahankan kesimbangan antara permintaan dan produksi padi, maka hampir semua negara penghasil padi berupaya menstabilkan laju kenaikan produksi melalui peningkatan produktivitas tanaman (Darajat et al. 2008). Usaha untuk menstabilkan laju kenaikan produksi merupakan usaha yang tidak mudah karena adanya pengaruh antara lain faktor cekaman kekeringan, yang berakibat pada tertekannya pertumbuhan tanaman dan penurunan produksi. Salah satu cara untuk menanggulangi masalah ini adalah menanam varietas toleran kekeringan. Upaya perbaikan sifat toleran kekeringan pada padi, baik padi lokal maupun padi unggul masih sangat terbatas (Suardi et al. 2004).
Transformasi genetika tanaman merupakan alternatif untuk memecahkan masalah yang sulit diatasi dengan teknik pemuliaan tanaman konvensional. Teknik tersebut mempunyai keuntungan, antara lain dapat memindahkan suatu gen ke kutivar target tanpa melalui banyak persilangan, dan gen tersebut dapat ditransfer tanpa menyertakan banyak gen lain yang tidak dikehendaki (Usyati et al. 2010). Padi transgenik yang sudah dihasilkan ada yang toleran terhadap cekaman biotik dan toleran terhadap kondisi lingkungan yang ekstrim (Bajaj dan Mohanty 2005). Beberapa hasil penelitian yang telah menghasilkan padi transgenik yang toleran terhadap cekaman biotik, yaitu padi tahan penggerek batang menggunakan gen cry1AB ( Ye et al. 2001), padi tahan penggerek batang padi kuning menggunakan gen cry1Ac (Nayak et al. 1997), padi tahan wereng coklat menggunakan gen GNA (Rao et al. 1998), padi tahan blast dengan gen Pi-d2 (De-xi et al. 2010), padi toleran terhadap cekaman abiotik dengan gen DREB1A (Oh et al. 2005), padi tolerans terhadap defisit air pada kondisi suhu rendah dengan gen OsDREB1A (JIRCAS 2005).
Tujuan Penelitian
Untuk mengetahui pengaruh gen 35S::OsDREB1A terhadap tanaman padi varietas elit setelah dipindahkan melalui persilangan.
Metodologi Penelitian
Bahan tanaman
Lima genotipe transgenik digunakan sebagai donor atau tetua jantan yaitu: B14, B11, C14, C10 , dan C3. Empat genotipe elit digunakan sebagai tetua betina yaitu: Ciherang, Batutegi, Code, dan Konawe.
Pelaksanaan penelitian
Penelitian dilakukan di Laboratorium Kultur Jaringan dan Rumah Kaca Fasilitas Uji Terbatas, Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian, Bogor. Penelitian dilaksanakan dari September 2010 – Mei 2011.
Persilangan
Persilangan dilakukan berdasarkan metode Jennings et al. (1979). Malai yang belum membuka padi yang berfungsi sebagai tetua betina yaitu varietas elit dikastrasi dan diserbuki dengan serbuk sari masak dari tanaman padi transgenik, kemudian malai disungkup dengan kertas minyak.
Uji kekeringan
kemudian dilakukan penilaian kesembuhan. Penilaina skor penggulungan daun, pengeringan daun dan kesembuhan tanaman dilakukan dengan berdasarkan skor pengamatan seperti yang dijelaskan pada Standard Evaluation System for Rice (IRRI 1996, Silitonga et al. 2003) yaitu :
Tabel 10. Klasifikasi tanggap tanaman terhadap kekeringan (daun menggulung pada fase vegetatif)
Skor/skala Kriteria
0 Daun-daun sehat
3 Daun-daun melipat (sangat berbentuk huruf V) 5 Daun betul-betul kuncup (membentuk huruf U) 7 Ujung-ujung daun bersentuhan (bentuk O) 9 Daun menggulung ketat
Tabel 11. Klasifikasi tanggap tanaman terhadap kekeringan (daun mengering pada fase vegetatif)
Skor/skala Keterangan
0 Tidak ada gejala
1 Ujung daun sedikit mengering
3 Ujung daun mengering sampai ¼ panjang pada hampir semua daun
5 ¼ sampai ½ dari semua daun betul-betul kering
7 Lebih dari ⅔ dari semua daun betul-betul kering
9 Tanaman mati
Tabel 12. Klasifikasi tanggap kesembuhan tanaman setelah perlakuan pemulihan
Skor/skala Keterangan
1 90-100% tanaman sembuh
3 70-89% tanaman sembuh
5 40-69% tanaman sembuh
7 20-39% tanaman sembuh
9 0-19% tanaman sembuh
Silang balik
Tanaman padi transgenik yang lolos seleksi terhadap kekeringan ditanam pada ember yang berisi media tanah untuk dijadikan tetua jantan (donor) dan disilangkan dengan tetua betina (non transgenik). Biji hasil silang balik ditanam untuk dianalisis secara molekuler dengan teknik PCR.
Isolasi DNA