OPTIMASI UKURAN SUBCONTOH MELALUI
BOOTSTRAP
DAN
JACKKNIFE
UNTUK PENDUGAAN STATISTIK DEMOGRAFI
HAMA
Aphis glycines
LENI MARLENA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul “Optimasi Ukuran Subcontoh Melalui Bootstrap dan Jackknife untuk Pendugaan Statistik Demografi Hama Aphis glycines” adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Agustus 2014
Leni Marlena
RINGKASAN
LENI MARLENA. Optimasi Ukuran Subcontoh Melalui Bootstrap dan Jackknife
untuk Pendugaan Statistik Demografi Hama Aphis glycines. Dibimbing oleh BUDI SUSETYO dan HERMANU TRIWIDODO.
Pendugaan statistik demografi diperlukan untuk dapat menduga pertumbuhan suatu populasi. Pertumbuhan suatu populasi dapat dihitung berdasarkan pertumbuhan betina dalam menghasilkan keturunan. Statistik demografi yang penting untuk mengetahui tingkat kepadatan populasi hama yaitu laju reproduksi kotor (GRR), laju reproduksi bersih (R0), laju pertumbuhan intrinsik (r), dan masa generasi (T).
Metode pengumpulan data dalam menghitung statistik demografi ini yaitu dengan cara memelihara beberapa hama kemudian perkembangan setiap individunya dicatat setiap hari. Data hasil pengamatan ini kemudian disebut sebagai data kohort. Dari data kohort yang ada kemudian dibuat sebuah Tabel Kehidupan (Life Table), kemudian dihitung dugaan GRR, R0, r, dan T. Berdasarkan hasil penelitian biologi ini hanya akan diperoleh satu nilai statistik demografi tersebut. Dengan demikian data ini tidak memiliki keragaman. Padahal keragaman penting dalam statistika yaitu dalam menduga suatu parameter. Apabila peneliti ingin mendapatkan keragaman dari statistik demografi tersebutmaka penelitian biologi hama harus dilakukan beberapa kali. Akan tetapi, cara seperti ini akan membutuhkan biaya yang cukup besar dan waktu yang cukup lama.
Dalam statistika dikenal suatu metode resampling yaitu bootstrap dan
jackknife. Denganmenggunakan kedua metode ini maka dari satu gugus data kohort hasil penelitian biologi hama akan dapat dihasilkan beberapa gugus data baru sehingga dapat diperoleh beberapa nilai statistik demografi. Selain itu, dengan menggunakan metode bootstrap akan ditentukan ukuran subcontohyang optimum agar dapat dijadikan rekomendasi bagi peneliti hama dalam menduga statistik demografi hama A. glycines. Pada penelitian ini juga dilakukan suatu uji-t untuk mengetahui pemberian PGPR dapat menghambat laju pertumbuhan intrinsik hama
A. glycines atau tidak.
Berdasarkan hasil penelitian ini, dapat disimpulkan bahwa ukuran sebesar 19 atau 20 adalah ukuran subcontoh yang optimum dalam menduga statistik demografi hama A. glycines. Kemudian, hasil uji-t pada taraf nyata 5% menunjukkan bahwa pemberian PGPR berpengaruh dalam menurunkan laju pertumbuhan intrinsik hama
A. glycines. Pada penelitian ini juga ditunjukkan bahwa pendugaan laju pertumbuhan intrinsik melalui matriks Leslie dan persamaan Lotka-Euler menghasilkan nilai dugaan yang tepat sama. Pendugaan laju pertumbuhan intrinsik melalui matriks Leslie jauh lebih mudah dan cepat dibandingkan dengan melalui persamaan Lotka-Euler.
SUMMARY
LENI MARLENA. Optimization Subsample Size by Bootstrap and Jackknife to Estimate Demographic Statistic of Pest Aphis glycines. Supervised by BUDI SUSETYO and HERMANU TRIWIDODO.
Estimation of demographic statistic is needed to estimate the growth of a population. The population growth can be calculated based on the growth of the female in reproducing the offsprings. Gross reproductive rate (GRR), net reproductive rate (R0), intrinsic rate of increase (r), and generation time (T) are the demographic statistic which are important in determining the level of pest population density.
The method to collect the data of which was used to obtain the statistic, was done by preserving some pests to then each pest was observed on daily basis and any growth to be noted. The observation result is then to be called as cohort data. Based on the existing cohort data, a life table was constructed, following the estimation of GRR, R0, r, and T. Based on this biological observation, only one value of those statistic was obtained. It means that the data does not have variability, whereas variability is needed on parameter estimation. If a researcher would like to obtain the variability of those demographic statistic, so the biological observation ought to be repeated many times. However, to conduct such method, much cost and time will be needed.
In statistics, there are known resampling methods such as bootstrap and
jackknife. Using the two methods, new set cohort data can be generated from the original data set so that some demographic statistic could be obtained. Besides, by using bootstrap method, the optimum subsample size could be determined. In this research, t-test was applied to determine whether or not the PGPR can be used to decrease the intrinsic rate of increase of A. glycines.
From this research we can conclude that 19 or 20 is the optimum subsample size in estimating demographic statistic of A. glycines. Using 5% level of significance, the t-test shows that PGPR can be used to decrease the intrinsic rate of increase of A. glycines. Two approximations used to estimate the intrinsic rate of increase (Leslie matrix and Lotka-Euler equation) gave the same results. In this research, the estimation of the instrinsic rate of increase using Leslie matrix is much easier and faster than Lotka-Euler equation.
© Hak Cipta Milik IPB, Tahun 2014
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah, dan pengutipan tersebut tidak merugikan kepentingan IPB
i
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Statistika
OPTIMASI UKURAN SUBCONTOH MELALUI
BOOTSTRAP
DAN
JACKKNIFE
UNTUK PENDUGAAN STATISTIK
DEMOGRAFI HAMA
Aphis glycines
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2014
ii
iii
Judul Tesis : Optimasi Ukuran Subcontoh Melalui Bootstrap dan Jackknife untuk Pendugaan Statistik Demografi Hama Aphis glycines
Nama : Leni Marlena NIM : G151120081
Disetujui oleh Komisi Pembimbing
Dr Ir Budi Susetyo, MS Ketua
Dr Ir Hermanu Triwidodo, MSc Anggota
Diketahui oleh
Ketua Program Studi Statistika
Dr Ir Anik Djuraidah, MS
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
iv
PRAKATA
Puji syukur penulis panjatkan kehadirat Allah SWTatas limpahan rahmat dan ridho-Nya, kesempatan, dan kesehatan yang dikaruniakan-Nya sehingga tesis yang berjudul “Optimasi Ukuran Subcontoh Melalui Bootstrap dan Jackknife untuk Pendugaan Statistik Demografi Hama Aphis glycines” ini dapat terselesaikan.
Terima kasih penulis ucapkan kepada Bapak Dr Ir Budi Susteyo, MS dan Bapak Dr Ir Hermanu Triwidodo, MSc selaku pembimbing atas kesediaan dan kesabarannya dalam membimbing dan membagi ilmunya kepada penulis selama penyusunan tesis ini. Ucapan terima kasih juga penulis sampaikan sebesar-besarnya kepada seluruh Dosen Departemen Statistika IPB yang telah mengasuh dan mendidik penulis selama di perkuliahan hingga berhasil menyelesaikan studi, serta seluruh staf Departemen Statistika IPB atas bantuan, pelayanan, dan kerjasamanya selama ini.
Ucapan terima kasih dan penghargaan yang tak terhingga juga penulis ucapkan kepada kedua orang tua tercinta dan adik – adik tersayang atas doa dan dukungannya. Terakhir tak lupa penulis juga menyampaikan terima kasih kepada teman – teman mahasiswa Pascasarjana Departemen Statistika atas segala bantuan dan kebersamaannya selama menghadapi masa – masa terindah maupun tersulit dalam menuntut ilmu, serta semua pihak yang telah banyak membantu dan tak sempat penulis sebutkan satu per satu.
Semoga tesis ini dapat bermanfaat.
Bogor, Agustus 2014
v
DAFTAR ISI
DAFTAR GAMBAR vi
DAFTAR TABEL vi
DAFTAR LAMPIRAN vi
1 PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
2 TINJAUAN PUSTAKA 2
Model Pertumbuhan Populasi Eksponensial 2
Tabel Kehidupan (Life Table) 3
Pendugaan Laju Pertumbuhan Intrinsik 4
Model Matriks Pertumbuhan Populasi 5
Metode Jackknife 6
Metode Bootstrap 7
3 DATA DAN METODE 8
Data 8
Prosedur Pendugaan dengan Menggunakan Jackknife dan Bootstrap 9
4 HASIL DAN PEMBAHASAN 13
Pendugaan Statistik Demografi A. glycines Kedelai Kontrol 14 Pendugaan Statistik Demografi A. glycines Kedelai Aplikasi PGPR 16 Pembuktian Terhadap Pendugaan Laju Pertumbuhan Intrinsik (r) 19
5 SIMPULAN 21
vi
DAFTAR GAMBAR
1 Contoh tabel kehidupan 4
2 Bagan alir algoritma pendugaan r secara iteratif 10
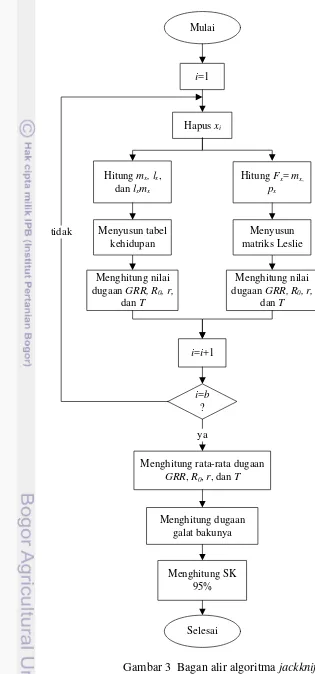
3 Bagan alir algoritma jackknife 11
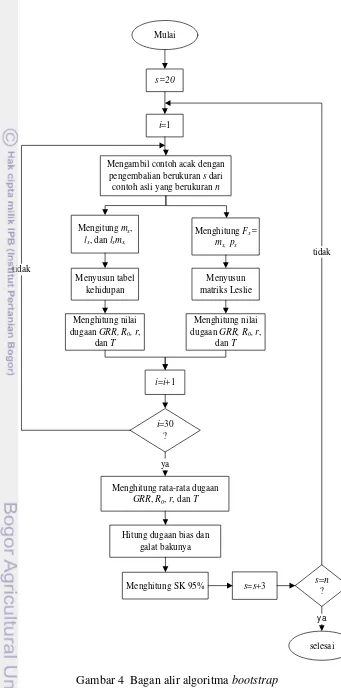
4 Bagan alir algoritma bootstrap 12
DAFTAR TABEL
1 Statistik demografi A. glycines data contoh asli 13 2 SK 95% bagi GRRA. glycines kedelai kontrol hasil resampling bootstrap dan
jackknife 14
3 SK 95% bagi r dan T A. glycines kedelai kontrol hasil resampling bootstrap
dan jackknife 15
4 SK 95% bagi GRR dan R0 A. glycines kedelai Aplikasi PGPR hasil resampling
bootstrap dan jackknife 16
5 SK 95% bagi r dan T A. glycines kedelai aplikasi PGPR hasil resampling
bootstrap dan jackknife 17
6 Uji-t efektivitas pemberian aplikasi PGPR terhadap laju pertumbuhan intrinsik
A. glycines 18
7 Rataan waktu komputasi pendugaan statistik demografi A. glycines hasil metode bootstrap dan jackknife pada berbagai jenis perhitungan 18
DAFTAR LAMPIRAN
1 Contoh Data Kohort A. glycines kedelai kontrol 23 2 Contoh Data Kohort A. glycines kedelai aplikasi PGPR 23
3 Tabel kehidupan A. glycines kedelai kontrol 24
4 Tabel kehidupan A. glycines kedelai aplikasi PGPR 25 5 Contoh sintaks SAS 9.2 prosedur jackknife untuk menghitung statistik
demografi berdasarkan tabel kehidupan 25
6 Contoh sintaks SAS 9.2 prosedur jackknife untuk menghitung statistik
demografi berdasarkan matriks Leslie 27
7 Contoh sintaks SAS 9.2 pembangkitan data bootstrap 28 8 Contoh sintaks SAS 9.2 untuk menghitung statistik demografi (r iteratif)
berdasarkan hasil bangkitan bootstrap 28
9 Contoh sintaks SAS 9.2 untuk menghitung statistik demografi (r matriks
Leslie) berdasarkan hasil bangkitan bootstrap 30
1
PENDAHULUAN
Latar Belakang
Pendugaan statistik demografi diperlukan untuk dapat menduga pertumbuhan suatu populasi. Pertumbuhan populasi dapat dihitung berdasarkan pertumbuhan betina dalam menghasilkan keturunan. Statistik demografi serangga oleh Zeng et al. (1983) diartikan sebagai analisis secara kuantitatif populasi serangga dalam hubungannya dengan kelangsungan hidup, keperidian, dan pola pertumbuhan populasi.
Statistik demografi yang penting untuk mengetahui tingkat kepadatan populasi hama yaitu laju reproduksi kotor (GRR), laju reproduksi bersih (R0), laju pertumbuhan intrinsik (r), dan masa generasi (T). Laju reproduksi kotor menunjukkan jumlah keturunan betina yang dihasilkan oleh semua serangga betina sepanjang generasi kohort (Poole 1974). Laju reproduksi bersih menunjukkan rata-rata jumlah keturunan yang dihasilkan oleh setiap individu sepanjang generasi kohort (Poole 1974). Laju pertumbuhan intrinsik (r) menggambarkan laju peningkatan populasi pada populasi yang berkembang dengan sumber daya yang tidak terbatas (Poole 1974). Kemudian masa generasi (T)merupakan waktu yang dibutuhkan sejak nimfa dilahirkan sampai menjadi imago dan melahirkan nimfa untuk pertama kalinya (Price 1997; Begon et al. 2008).
Metode pengumpulan data untuk dapat menduga statistik demografi tersebut yaitu dengan cara memelihara beberapa hama kemudian dicatat perkembangan setiap individunya. Pengamatan dilakukan setiap hari, mulai dari instar 1 sampai serangga tersebut mati. Data hasil pengamatan seperti ini kemudian disebut sebagai data kohort. Dari data kohort yang ada kemudian dibuat sebuah tabel kehidupan (life table). Selanjutnya, dari tabel kehidupan tersebut dihitung nilai GRR, R0, r, dan
T. Dari satu penelitian biologi hama hanya akan dihasilkan satu nilai GRR, R0, r, dan T, sehingga nilai statistik demografi yang dihasilkan melalui satu penelitian tersebut tidak memiliki keragaman. Padahal keragaman penting dalam pendugaan suatu parameter. Agar diperoleh beberapa nilai untuk setiap statistik demografi, seharusnya penelitian biologi hama ini dilakukan beberapa kali. Akan tetapi, hal ini tentu sangat menyulitkan, membutuhkan waktu yang lama dan biaya yang cukup besar. Oleh sebab itu, penelitian untuk mengetahui statistik demografi dari suatu populasi tidak dapat dilakukan dengan penelitian biologi saja tetapi juga harus dilakukan pendekatan statistika agar dapat diperoleh ukuran keragaman dan ukuran pemusatannya.
Dalam statistika, kita mengenal istilah resampling yaitu pengambilan contoh acak secara berulang-ulang. Metode resampling yang umum dipelajari yaitu metode jackknife dan metode bootstrap. Dengan menggunakan metode resampling
2
statistik demografi di kemudian hari. Berdasarkan ukuran subcontoh yang diperoleh, diharapkan kedepannya para peneliti hama cukup menggunakan ukuran subcontoh tersebut pada saat proses resampling bootstrap.
Laju pertumbuhan intrinsik (r) pada umumnya dihitung melalui dua pendekatan yaitu dengan cara mengoptimasi persamaan Lotka-Euler dan dengan cara menghitung nilai akar ciri dari matriks Leslie. Pengoptimasian persamaan Lotka-Euler dilakukan secara iteratif sehingga memerlukan teknik komputasi yang cukup tinggi dan membutuhkan waktu komputasi yang lama. Sedangkan pendugaan r dengan mencari nilai akar ciri matriks Leslie, secara komputasi akan lebih mudah dan cepat. Pada penelitian ini akan ditunjukkan bahwa hasil dugaan r
melalui persamaan Lotka-Euler dan melalui matriks Leslie adalah sama dan menduga r melalui matriks Leslie adalah cara yang paling mudah dan cepat dibandingkan melalui persamaan Lotka-Euler. Sehingga untuk pendugaan laju pertumbuhan intrinsik selanjutnya direkomendasikan untuk menggunakan matriks Leslie saja dibandingkan menggunakan teknik optimasi persamaan Lotka-Euler.
Pada penelitian ini digunakan dua data kohort, yaitu data kohort A. glycines
kedelai kontrol dan A. glycines kedelai aplikasi PGPR (Plant Growth Promoting Rhizobacteria). PGPR merupakan kelompok bakteri yang dapat dimanfaatkan sebagai pupuk hayati untuk membantu tanaman menyediakan unsur hara dan memperkuat tanaman terhadap serangan hama maupun penyakit tanaman (Soesanto 2008 dalam Agustini 2013). Efektivitas pemberian PGPR dilihat dengan cara membandingkan nilai dugaan laju pertumbuhan intrinsik (r) hama A. glycines pada kedelai yang diberi PGPR dengan kedelai kontrol.
Tujuan Penelitian
Tujuan penelitian ini adalah:
1. Menentukan ukuran subcontoh yang optimum untuk pendugaan parameter
GRR, R0, r, dan T dengan menggunakan metode bootstrap;
2. Mengetahui efektivitas penggunaan PGPR dalam menghambat laju pertumbuhan intrinsik (r) hama A. glycines;
3. Membuktikan bahwa nilai dugaan laju pertumbuhan intrinsik (r) yang dihasilkan melalui matriks Leslie dan melalui optimasi persamaan Lotka-Euler adalah sama;
4. Menunjukkan bahwa pendugaan laju pertumbuhan intrinsik (r) dengan menggunakan matriks Leslie lebih mudah dan efisien dibandingkan dengan menggunakan optimasi persamaan Lotka-Euler.
2
TINJAUAN PUSTAKA
Model Pertumbuhan Populasi Eksponensial
Jenis – jenis pertumbuhan populasi yang umum dikenal yaitu pertumbuhan geometrik, pertumbuhan eksponensial, dan pertumbuhan Malthus. Hasibuan (1987) menyatakan bahwa jenis pertumbuhan populasi terjadi pada populasi yang memenuhi asumsi – asumsi berikut:
3 2. Tidak ada persaingan di antara individu – individu di dalam populasi;
3. Selalu ada ruang dan makanan yang cukup untuk mendukung populasi.
Berdasarkan asumsi – asumsi di atas, maka jika t adalah waktu, Nt adalah ukuran populasi pada waktu ke-t, dan jika ukuran populasi pada periode waktu ke-t adalah
λ kali ukuran populasi pada satu unit waktu sebelumnya, maka ukuran populasi pada waktu ke-t adalah (Hasibuan 1987):
Nt = N0 λt
Persamaan (1) dapat juga dituliskan dalam suatu persamaan yang memuat bilangan Euler (e), karena λ = er. Konstanta r, di dalam ekologi dikenal sebagai laju pertumbuhan intrinsik sedangkan λadalah laju pertumbuhan terbatas (finite rate of increase). Dengan demikian persamaan (1) merupakan model eksponensial apabila ditulis menjadi (Poole 1974):
Nt = ertN0
Selanjutnya, persamaan (1) dapat juga ditulis menjadi:
��
� = λt
Dengan demikian, konstanta λ dikenal juga sebagai laju reproduksi, atau nisbah antara ukuran populasi suatu waktu dengan ukuran populasi satu unit waktu sebelumnya λ = Nt/Nt-1.
Pendekatan lain model eksponensial dapat dilakukan sebagai berikut: berdasarkan asumsi 1, maka laju pertumbuhan populasi, r, adalah konstan (Hasibuan 1987). Pernyataan ini dalam bahasa matematika dapat ditulis sebagai:
d��
��dt= r
Ruas kiri persamaan (4) merupakan laju pertumbuhan perkapita. Persamaan ini juga dapat ditulis sebagai:
d�� dt = r��
Persamaan (5) mengindikasikan bahwa jumlah individu dalam suatu populasi akan berubah seiring berubahnya waktu. Hal ini mengakibatkan (dNt/dt) semakin menurun dan berhenti bertumbuh (r=0) pada saat daya dukung tercapai. Nilai r
konstan pada kondisi lingkungan yang konstan. Jika kondisi berubah, r juga akan berubah. Kondisi tertentu mungkin lebih kondusif untuk reproduksi atau menyebabkan mortalitas lebih rendah pada spesies tertentu dibandingkan lainnya (Poole 1974).
Tabel Kehidupan (Life Table)
Informasi dasar yang dibutuhkan untuk studi perubahan kepadatan dan laju pertumbuhan atau penurunan populasi terkandung dalam sebuah tabel kehidupan. Tabel kehidupan memuat beberapa statistik yang sangat penting seperti peluang suatu individu pada umur tertentu mati, atau sebaliknya. Tabel kehidupan kohort merupakan tabel kehidupan yang mengikuti perkembangan kohort dimulai dari kemunculan individu pertama (kelahiran) sampai kematian individu terakhir yang bertahan hidup (Begon et al. 2008). Tabel kehidupan pada serangga sangat penting dikaji untuk mengetahui perkembangan, distribusi, dan kepadatan populasi serangga (Andrewartha dan Birch 1982).
(1)
(3)
(4) (2)
4
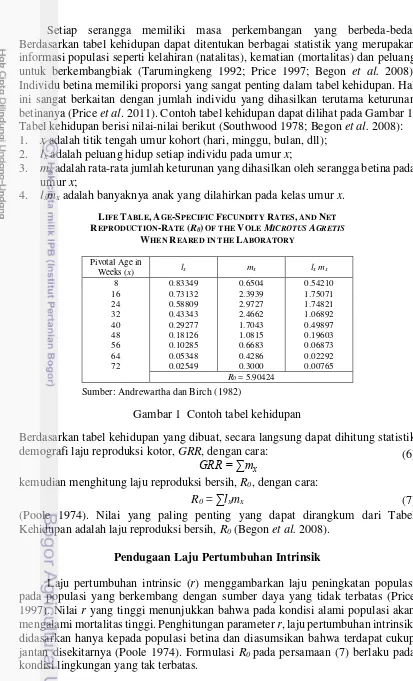
Setiap serangga memiliki masa perkembangan yang berbeda-beda. Berdasarkan tabel kehidupan dapat ditentukan berbagai statistik yang merupakan informasi populasi seperti kelahiran (natalitas), kematian (mortalitas) dan peluang untuk berkembangbiak (Tarumingkeng 1992; Price 1997; Begon et al. 2008). Individu betina memiliki proporsi yang sangat penting dalam tabel kehidupan. Hal ini sangat berkaitan dengan jumlah individu yang dihasilkan terutama keturunan betinanya (Price et al.2011). Contoh tabel kehidupan dapat dilihat pada Gambar 1. Tabel kehidupan berisi nilai-nilai berikut (Southwood 1978; Begon et al. 2008): 1. x adalah titik tengah umur kohort (hari, minggu, bulan, dll);
2. lx adalah peluang hidup setiap individu pada umur x;
3. mx adalah rata-rata jumlah keturunan yang dihasilkan oleh serangga betina pada umur x;
4. lxmx adalah banyaknya anak yang dilahirkan pada kelas umur x.
Berdasarkan tabel kehidupan yang dibuat, secara langsung dapat dihitung statistik demografi laju reproduksi kotor, GRR, dengan cara:
GRR = ∑mx
kemudian menghitung laju reproduksi bersih, R0, dengan cara:
R0 = ∑lxmx
(Poole 1974). Nilai yang paling penting yang dapat dirangkum dari Tabel Kehidupan adalah laju reproduksi bersih, R0 (Begon et al. 2008).
Pendugaan Laju Pertumbuhan Intrinsik
Laju pertumbuhan intrinsic (r) menggambarkan laju peningkatan populasi pada populasi yang berkembang dengan sumber daya yang tidak terbatas (Price 1997). Nilai r yang tinggi menunjukkan bahwa pada kondisi alami populasi akan mengalami mortalitas tinggi. Penghitungan parameter r, laju pertumbuhan intrinsik, didasarkan hanya kepada populasi betina dan diasumsikan bahwa terdapat cukup jantan disekitarnya (Poole 1974). Formulasi R0pada persamaan (7) berlaku pada kondisi lingkungan yang tak terbatas.
(7) (6)
LIFE TABLE, AGE-SPECIFIC FECUNDITY RATES, AND NET REPRODUCTION-RATE (R0) OF THE VOLEMICROTUS AGRETIS
WHEN REARED IN THE LABORATORY
Pivotal Age in
Weeks (x) lx mx lx mx
8 0.83349 0.6504 0.54210
16 0.73132 2.3939 1.75071
24 0.58809 2.9727 1.74821
32 0.43343 2.4662 1.06892
40 0.29277 1.7043 0.49897
48 0.18126 1.0815 0.19603
56 0.10285 0.6683 0.06873
64 0.05348 0.4286 0.02292
72 0.02549 0.3000 0.00765
R0= 5.90424 Sumber: Andrewartha dan Birch (1982)
5 Laju reproduksi bersih, R0, analogi dengan λ, laju pertumbuhan terbatas, kecuali apabila λ dibatasi untuk selang waktu sama dengan 1, dan R0pada suatu jangka waktu sama dengan panjang rata – rata dari suatu generasi T (Poole 1974). Jika selang waktu t sama dengan masa generasi T, R0 sama dengan λ, maka secara umum:
R0 = erT.
Dengan demikian, masa generasi dapat diduga dengan membagi logaritma natural (ln) dari R0 dengan laju pertumbuhan intrinsik r,
T=lnR0 r .
Jika λ mendekati 1, maka persamaan (8) dapat didekati dengan (Dublin dan Lotka 1925 dalam Poole 1974):
� ≈ ∑∑xllxmx xmx
atau jumlah dari kolom xlxmx(misal x adalah titik tengah umur dalam hari) dibagi dengan R0. Jika T diketahui, maka kira – kira dugaan bagi r adalah:
r =lnR0 T .
Hal ini hanya perkiraan kasar dan akan akurat jika λ mendekati 1.
Poole (1974) menyatakan bahwa jika angka kematian dan kelahiran tetap atau sama dan populasinya memiliki sebaran usia yang stabil, maka nilai dari r dapat diduga dengan persamaan: Persamaan (11) disebut juga sebagai persamaan Lotka-Euler. Berdasarkan persamaan (11) tersebut, maka nilai r diperoleh melalui proses iterasi. Nilai yang mungkin dari r, disubstitusikan ke dalam sisi kiri persamaan (11), dengan dugaan awal r dihitung dari persamaan (10). Iterasi dilakukan sampai sisi kiri persamaan (11) sama dengan 1.
Model Matriks Pertumbuhan Populasi
Matriks Leslie digunakan sebagai model pertumbuhan populasi untuk mengetahui prediksi jumlah dan prediksi laju pertumbuhan dari suatu populasi untuk tahun berikutnya. Model Leslie merupakan alat yang sangat berguna untuk menentukan pertumbuhan populasi berdasarkan sebaran umur dalam populasi sepanjang waktu.
Matriks Leslie memiliki bentuk yang unik yaitu matriks Leslie berbentuk matriks persegi dengan elemen baris pertama dari matriks Leslie terdiri dari tingkat kesuburan betina, sub diagonalnya berisi tingkat ketahanan hidup betina dan elemen yang lain bernilai nol. Matriks Leslie ditemukan oleh seorang pakar Ekologi bernama P. H Leslie pada tahun 1945. Matriks Leslie dikembangkan dari matriks sederhana yang dikemukakan pertama kali oleh Lotka pada tahun 1920-an.
Populasi yang digunakan pada perhitungan dengan matriks Leslie adalah populasi betina dari populasi tersebut. Tiga statistik dasar yang diperlukan untuk menyusun matriks Leslie adalah (Poole 1974):
(8)
(9)
(10)
6
Persamaan di atas dapat ditulis dalam bentuk matriks sebagai berikut:
atau ditulis:
��+ = ���,
Vektor nt menunjukkan ukuran populasi berdasarkan kategori umurnya yang dihitung pada waktu ke-t, dan M adalah matriks Leslie.
Matriks Leslie dan tabel kehidupan adalah model demografi yang umum digunakan untuk mengevaluasi kemampuan hidup suatu populasi. Matriks Leslie maupun tabel kehidupan dapat digunakan untuk menduga laju pertumbuhan intrinsik. Matriks Leslie memiliki paling banyak satu nilai akar ciri real positif yang nilainya akan selalu lebih besar dari nilai akar ciri yang lainya (nilai akar ciri lainnya adalah bilangan negatif atau kompleks). Nilai akar ciri positif tersebut dinamakan nilai akar ciri dominan. Jika dimisalkan 1 merupakan nilai akar ciri positif dominan dari matriks Leslie, maka nilai 1 akan sama dengan laju pertumbuhan terbatas, λ. Jika laju pertumbuhan terbatasnya ada, maka populasi akan mencapai sebaran umur stabil (Poole 1974).
Pratama et al. (2013) menyatakan bahwa jika diketahui 1 < 1, maka populasi pada semua kelas umur cenderung menurun. Kemudian jika diketahui 1 = 1 , maka populasi pada semua kelas umur cenderung tetap. Sedangkan apabila diketahui 1 > 1, maka populasi pada semua kelas umur cenderung meningkat. Dengan demikian untuk memprediksi laju pertumbuhan suatu populasi dapat menggunakan nilai akar ciri dominan dari matriks Leslie.
Metode Jackknife
Istilah “Jackknife” muncul dari analogi pisau lipat Swiss yang mudah dibawa
kemana-mana. Dengan analogi tersebut, Tukey (1958) menciptakan istilah
7 selang kepercayaan. Jackknife digunakan dalam statistika inferensia untuk menduga bias dan galat baku suatu statistik.
Contoh acak dari pengamatan digunakan untuk menghitung bias dan galat baku. Ide dasar dibalik penduga jackknife adalah melakukan penghitungan dengan menghilangkan satu atau beberapa pengamatan dari contoh yang ditetapkan. Kemudian berdasarkan gugus data baru tersebut dihitung bias dan ragam statistiknya.
Proses resampling dilakukan dengan membuang satu pengamatan secara berulang-ulang. Misalkan x = ( x1, x2, x3, … , xn ) adalah satu gugus data contoh acak sebanyak n pengamatan dan didefinisikan x(i) adalah gugus data hasil
resampling dengan mengeluarkan pengamatan ke-i. Artinya proses resampling
dilakukan sebanyak n kali, dengan setiap gugus data yang baru mempunyai ukuran contoh sebesar m = n – 1, sehingga untuk setiap resampling ke-i diperoleh gugus data x(i) = ( x1, x2, …, xi-1, xi+1, …, xn ).
Jika �̂ didefinisikan sebagai penduga suatu parameter dengan fungsi �̂ = s(x),
maka untuk menghitung statistik masing – masing replikasi pada resampling jackknife dapat menggunakan formula berikut:
�̂ =s(x(i)), untuk i=1,2, …,n. Misal
�̂• = ∑ �̂ �
= maka dugaan bias jackknife adalah:
Biaŝ jack= n-1 (θ̂ • -θ̂n)
sehingga dugaan jackknife,�̂�� dari θ adalah: θ
̂jack= θ̂n-Biaŝ =n θ̂n- n-1 θ̂ • .
Dugaan galat baku jackknife adalah:
sêjack= {n-n ∑(θ̂ i -θ̂• ) n
i=
}
Metode Bootstrap
Pada tahun 1979, seorang professor statistika di Stanford, Bradley Efron, memperkenalkan istilah Bootstrap dan ditulis dalam bentuk buku oleh Efron dan Tibshirani (1993) dan beberapa penulis lain sesudahnya. Bootstrap berasal dari kata “pull oneself up one’s Bootstrap” yang berarti berpijak diatas kaki sendiri, dalam pengertian bahwa data yang sedikit dan data yang tidak diketahui sebaran populasinya (Efron dan Tibshirani 1993).
Bootstrap merupakan suatu metode statistik yang dapat digunakan untuk menduga sebaran penarikan contoh (sampling distribution) dari suatu penduga.
Bootstrap dikenal sebagai suatu metode pengambilan contoh secara acak dengan pengembalian (resampling with replacement). Bootstrap merupakan suatu metode statistik yang bersifat nonparametrik. Suatu populasi yang tidak diketahui sebarannya atau populasi nonparametrik dapat menggunakan bootstrap untuk melakukan pendugaan parameter. Misalkan x = ( x1, x2, x3, … , xn) adalah satu (15)
(16)
8
gugus data contoh acak berukuran n, kemudian �̂ didefinisikan sebagai penduga suatu parameter dengan fungsi �̂ = � � maka pembangkitan contoh acak
Bootstrap nonparametrik adalah :
1. Mengambil contoh acak berukuran m dengan pemulihan dari gugus data
x = ( x1, x2, x3, … , xn) berukuran n. Misalkan diperoleh gugus data baru x*=(x1*,x2*,…..,xm*) kemudian diulang sebanyak B kali.
2. Berdasarkan setiap penarikan contoh acak dilakukan pendugaan θ sehingga diperoleh gugus data �̂ , �̂ , �̂ , … , �̂�.
3. Dari gugus nilai �̂ , �̂ , �̂ , … , �̂� diperoleh nilai rata-rata, �̂• , yang merupakan nilai penduga bootstrap untuk
.Dengan demikian nilai penduga bootstrap yaitu:
�̂• = �∑�̂�. �
=
Selanjutnya bias bootstrap dihitung dengan formula sebagai berikut:
Biaŝ boot= �̂• − �̂
Kemudian galat baku sêbootdiduga dengan dengan menggunakan simpangan baku contoh dari B replikasi,
sêboot={∑(θ
̂B-θ̂(•))2
B-1
B
i=1
} 1 2
3
DATA DAN METODE
Data
Data yang akan digunakan dalam penelitian ini adalah data kohort kutu daun (A. glycines) pada tanaman kedelai kontrol dan tanaman kedelai aplikasi PGPR hasil penelitian mahasiswa S1 Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor. Penelitian tersebut dilakukan pada bulan Januari sampai April 2013. Pengamatan dilakukan setiap hari pada masing-masing A. glycines.
Siklus hidup A. glycines dihitung sejak nimfa instar 1 diinokulasi ke tanaman perlakuan sampai menjadi imago.
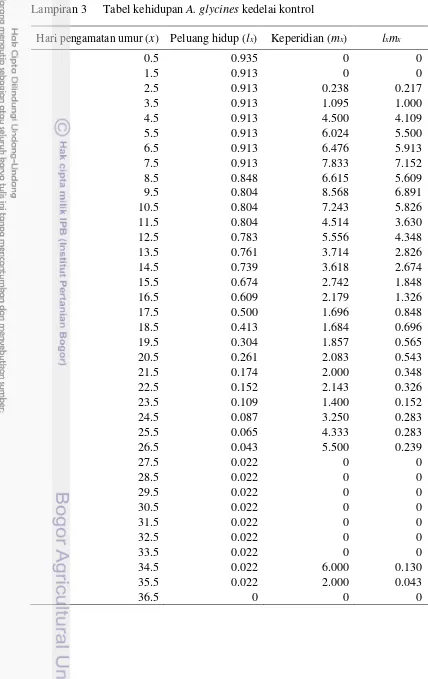
Selama perkembangan menjadi imago A. glycines akan mengalami fase nimfa sebanyak 4 stadia instar, perpindahan antar instar dibedakan melalui pergantian kulit. Pengamatan lama hidup dilakukan mulai dari A. glycines menjadi imago hingga imago tersebut mati. Kemudian pengamatan keperidian didapat dari hasil jumlah nimfa yang dilahirkan oleh setiap imago A. glycines selama hidupnya. Contoh data kohort A. glycines pada tanaman kedelai kontrol dan kedelai aplikasi PGPR dapat dilihat pada Lampiran 1 dan 2. Berdasarkan hasil pengamatan harian tersebut, maka disusun tabel kehidupan A. glycines. Tabel kehidupan A. glycines
pada tanaman kedelai kontrol dan kedelai aplikasi PGPR dapat dilihat pada Lampiran 3 dan 4.
(18)
(19)
9 Jumlah individu yang masih hidup dihitung setiap hari, sehingga diperoleh data peluang hidup (lx) A. glycines pada berbagai umur. Pengamatan keperidian harian (mx) dilakukan dengan merata-ratakan jumlah nimfa A. glycines yang dilahirkan oleh setiap imago untuk berbagai umur (x). Data kohort A. glycinespada tanaman kedelai kontrol terdiri dari 46 individu dengan panjang hari pengamatan yaitu 37 hari. Sedangkan data kohort A. glycines pada tanaman kedelai aplikasi PGPR terdiri dari 41 individu dengan panjang hari pengamatan yaitu 26 hari.
Prosedur Pendugaan dengan Menggunakan Jackknife dan Bootstrap
Pada setiap data kohort A. glycinestanaman kedelai kontrol dan tanaman kedelai aplikasi PGPR diterapkan metode Jackknife dan Bootstrap. Berikut ini adalah langkah – langkah pengerjaan untuk masing – masing metode.
A. Metode Jackknife
Berikut ini adalah langkah – langkah untuk menghitung statistik demografi hama A. glycines dengan menggunakan metode jackknife:
1. Membuang baris ke – i data kohort asli sehingga terbentuk data baru yang berukuran k= b-1, dengan i = 1, 2, ..., b;
2. a. Menghitung nilai mx, lx, dan lxmx; b. Menghitung nilai Fx = mx dan px;
3. a. Menyusun tabel kehidupan dari nilai yang diperoleh pada langkah 2a; b. Menyusun matriks Leslie dari nilai yang diperoleh pada langkah 2b;
4. Menghitung nilai dugaan GRR, R0, r, dan T. Statistik demografi r dan T
dihitungdua kali. Pertama dihitung berdasarkan tabel kehidupan (3a) dan yang kedua dihitung berdasarkan matriks Leslie (3b);
5. Mengulang langkah 1, 2, 3, dan 4 sampai baris yang dibuang adalah baris ke –
n data kohort contoh asli;
6. Menghitung dugaan GRR, R0, r, dan T hasil resamplingjackknife; 7. Menghitung dugaan galat baku untuk setiap statistik demografi tersebut; 8. Membuat selang kepercayaan (SK) 95% bagi GRR, R0, r, dan T.
B. Metode Bootstrap
Ukuran subcontoh (s) yang akan dicobakan pada metode bootstrap untuk data kohort A. glycines pada tanaman kedelai kontrol adalah 19, 22, 25, 28, 31, 34, 37, 40, 43, dan 46. Kemudian ukuran subcontoh (s) yang akan dicobakan pada data kohort A. glycinespada tanaman kedelai aplikasi PGPR adalah 20, 23, 26, 29, 32, 35, 38, dan 41. Berikut ini adalah langkah – langkah untuk menghitung statistik demografi hama A. glycines dengan menggunakan metode bootstrap:
1. Melakukan pengambilan contoh secara acak dengan pengembalian berukuran s
dari data contoh asli yang berukuran nx*=(x1*,x2*, ...,xs*); 2. a. Menghitung nilai mx, lx, dan lxmx;
b. Menghitung nilai Fx = mx dan px;
3. a. Menyusun tabel kehidupan dari nilai yang diperoleh pada langkah 2a; b. Menyusun matriks Leslie dari nilai yang diperoleh pada langkah 2b;
4. Menghitung nilai dugaan GRR, R0, r, dan T. Statistik demografi r dan T dihitung dua kali. Pertama dihitung berdasarkan tabel kehidupan (3a) dan yang kedua dihitung berdasarkan matriks Leslie (3b);
10
6. Menghitung rata-rata dugaan GRR, R0, r, dan T hasil resamplingbootstrap; 7. Menghitung dugaan galat baku untuk setiap statistik demografi tersebut; 8. Membuat selang kepercayaan (SK) 95% bagi GRR, R0, r, dan T;
9. Mengulang langkah 1 sampai 8 untuk berbagai ukuran subcontoh s.
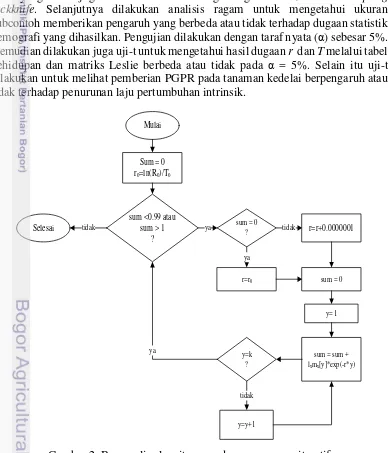
Bagan alir algoritma laju pertumbuhan intrinsik baik melalui metode
bootstrap maupun jackknife dapat dilihat pada Gambar 2. Kemudian bagan alir algoritma pendugaan statistik demografi berdasarkan hasil resampling bootstrap
dan jackknife dapat dilihat pada Gambar 3 dan Gambar 4. Selanjutnya, dibuatlah sebuah program dalam SAS 9.3 (sintax terdapat pada Lampiran 5 – 9).
Misal θboot adalah penduga bagi θ hasil resampling bootstrap, maka SK 95% dihitung dengan menggunakan rumus berikut:
θboot ± zα/2seboot.
Cara yang sama juga dilakukan untuk menghitung SK 95% hasil resampling jackknife. Selanjutnya dilakukan analisis ragam untuk mengetahui ukuran subcontoh memberikan pengaruh yang berbeda atau tidak terhadap dugaan statistik demografi yang dihasilkan. Pengujian dilakukan dengan taraf nyata (α) sebesar 5%. Kemudian dilakukan juga uji-t untuk mengetahui hasil dugaan r dan T melalui tabel kehidupan dan matriks Leslie berbeda atau tidak pada α = 5%. Selain itu uji-t dilakukan untuk melihat pemberian PGPR pada tanaman kedelai berpengaruh atau tidak terhadap penurunan laju pertumbuhan intrinsik.
Mulai
Sum = 0 r0=ln(R0)/T0
sum <0.99 atau sum > 1
?
r=r0 sum = 0
y= 1
sum = sum + lxmx[y]*exp(-r*y)
y=y+1 y=k
?
tidak sum = 0
?
ya r=r+0.0000001
ya
tidak
ya
Selesai tidak
11
i=1
Hapus xi
i=i+1
i=b ? tidak
Menghitung rata-rata dugaan GRR, R0, r, dan T
ya
Menghitung dugaan galat bakunya Hitung mx, lx,
dan lxmx
Menghitung nilai dugaan GRR, R0, r,
dan T Menyusun tabel
kehidupan
Hitung Fx= mx,
px
Menghitung nilai dugaan GRR, R0, r,
dan T Menyusun matriks Leslie
Menghitung SK 95% Mulai
Selesai
12
s=20
i=1
Mengambil contoh acak dengan pengembalian berukuran s dari
contoh asli yang berukuran n
Mengitung mx,
lx, dan lxmx,
Menghitung nilai dugaan GRR, R0, r,
dan T
i=i+1
i=30 ? tidak
ya
Hitung dugaan bias dan galat bakunya
s=s+3 s=n
? tidak
Menyusun tabel kehidupan
Menghitung Fx=
mx, px
Menghitung nilai dugaan GRR, R0, r,
dan T
Menyusun matriks Leslie Mulai
selesai Menghitung rata-rata dugaan
GRR, R0, r, dan T
ya
Menghitung SK 95%
13
4
HASIL DAN PEMBAHASAN
Statistik demografi hama berdasarkan data kohort A. glycines asli yang dihitung menggunakan tabel kehidupan maupun matriks Leslie ditunjukkan oleh Tabel 1. Berdasarkan data kohort asli kedelai kontrol, nilai laju reproduksi kotor A. glycines (GRR) yaitu 104.86, artinya jumlah keturunan betina yang dihasilkan oleh semua A. glycines betina sepanjang generasi kohort adalah sebanyak 104.861 individu/generasi. Nilai laju reproduksi bersih (R0) yaitu 63.326 individu/generasi, dapat diartikan bahwa populasi A. glycines memiliki kapasitas untuk berkembang biak 63.326 kali pada setiap generasi dengan kondisi lingkungan tertentu dan dalam lingkungan tak terbatas (Poole 1974). Besarnya laju pertumbuhan intrinsik (r) untuk
A. glycines kedelai kontrol yaitu 0.537 individu/hari, artinya jumlah individu yang bertambah pada populasi A. glycines kedelai kontrol adalah 0.537 individu/hari (Andrewartha dan Birch 1982). Selanjutnya maka masa generasinya adalah 8.958 hari.
Pada kedelai aplikasi PGPR, nilai laju reproduksi kotor A. glycines (GRR) sebesar 71.834, artinya jumlah keturunan betina yang dihasilkan oleh semua A. glycines betina sepanjang generasi kohort adalah sebanyak 71.834 individu/generasi. Nilai laju reproduksi bersih (R0) A. glycines kedelai aplikasi PGPR yaitu 57.78 individu/generasi, dapat diartikan bahwa populasi A. glycines
memiliki kapasitas untuk berkembang biak 57.78 kali pada setiap generasi dengan kondisi lingkungan tertentu dan dalam lingkungan tak terbatas (Poole 1974). Kemudian nilai laju pertumbuhan intrinsik (r) untuk A. glycines kedelai aplikasi PGPR yaitu 0.513 individu/hari, artinya jumlah individu yang bertambah pada populasi A. glycines kedelai aplikasi PGPR adalah 0.513 individu/hari (Andrewartha dan Birch 1982). Kemudian masa generasi (T) pada A. glycines
kedelai aplikasi PGPR yaitu 7.911 hari.
Efektivitas pemberian PGPR pada tanaman kedelai dapat dilihat dari penurunan nilai laju pertumbuhan intrinsik (r) hama A. glycines. Dengan demikian perlu dilakukan uji-t satu populasi untuk mengetahui pemberian PGPR berpengaruh atau tidak dalam menghambat laju pertumbuhan intrinsik (r) hama A. glycines. Berdasarkan hasil yang diperoleh pada Tabel 1, pengujian tidak dapat dilakukan karena masing – masing data kohort hanya memiliki satu nilai r. Oleh sebab itu, tahapan selanjutnya adalah melakukan resampling bootstrap dan jackknife untuk dapat memperoleh keragaman data. Pada metode bootstrap, setiap data direplikasi sebanyak 30 kali, sehingga diperoleh 30 nilai dugaan bagi masing – masing statistik demografi. Sedangkan pada metode jackknife akan dihasilkan 46 nilai dugaan bagi masing – masing statistik demografi A. glycines kedelai kontrol dan 41 nilai bagi masing – masing statistik demografi. A. glycines kedelai aplikasi PGPR.
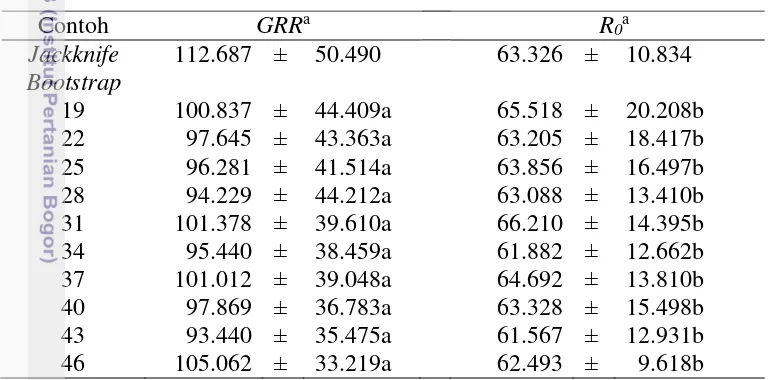
Tabel 1 Statistik demografi A. glycines data contoh asli
Kohort Metode GRR R0 r T
A. glycines kedelai
kontrol Tabel kehidupan 104.861 63.326 0.537 7.724
A. glycines kedelai
14
Pendugaan Statistik Demografi A. glycines Kedelai Kontrol
Tabel 2 memperlihatkan SK 95% bagi GRR dan R0 berdasarkan contoh
jackknife dan contoh bootstrap pada berbagai ukuran subcontoh. Nilai dugaan GRR
yang diperoleh melalui contoh bootstrap cenderung tidak konsisten. Selain itu, nilai dugaan galat baku yang dihasilkan masih sangat besar. Berbeda dengan hasil dugaan GRR, nilai dugaan R0 relatif konsisten meskipun nilai dugaan galat bakunya masih cukup besar. Hasil analisis ragam menunjukkan bahwa pada taraf nyata 5% ukuran subcontoh bootstrap tidak memberikan pengaruh yang berbeda terhadap hasil dugaan GRR dan R0 A. glycines kedelai kontrol. Dengan demikian dapat dinyatakan bahwa untuk pendugaan GRR dan R0 melalui metode bootstrap dapat menggunakan ukuran subcontoh yang minimum apabila ukuran ini dianggap dapat memudahkan pendugaan statistik demografi hama A. glycines.
Pada Tabel 2 juga terlihat bahwa SK 95% bagi GRR contoh jackknife lebih lebar dari SK 95% bagi GRR contoh bootstrap pada berbagai ukuran subcontoh. Dengan demikian, pada pendugaan GRR, nilai dugaan galat baku yang diperoleh melalui contoh jackknife lebih besar dari nilai dugaan galat baku yang diperoleh dari contoh bootstrap dan galat baku minimum yaitu saat ukuran subcontoh
bootstrap-nya sama dengan 46. Sedangkan pada pendugaan R0, umumnya nilai dugaan galat baku contoh jackknife lebih kecil dari nilai dugaan galat baku contoh
bootstrap, kecuali saat ukuran subcontoh bootstrap-nya sama dengan 46. Jadi, berdasarkan hasil pendugaan GRR dan R0, metode bootstrap akan menghasilkan nilai dugaan yang lebih baik dari jackknife apabila digunakan ukuran subcontoh yang sesuai dengan ukuran contoh data aslinya.
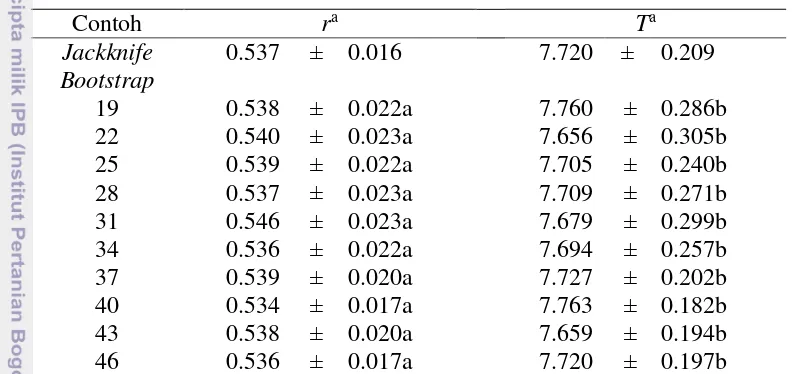
Nilai dugaan r yang dihitung melalui persamaan Euler-Lotka maupun matriks Leslie menghasilkan nilai yang tepat sama baik berdasarkan data contoh bootstrap
maupun data contoh jackknife.Tabel 3, memperlihatkan bahwa nilai dugaan rA. glycines kedelai kontrol konsisten dengan dugaan galat baku yang cukup kecil. Tabel 2 SK 95% bagi GRR A. glycines kedelai kontrol hasil resampling
bootstrap dan jackknife
Contoh GRRa R
0a
Jackknife 112.687 ± 50.490 63.326 ± 10.834
Bootstrap
19 100.837 ± 44.409a 65.518 ± 20.208b
22 97.645 ± 43.363a 63.205 ± 18.417b
25 96.281 ± 41.514a 63.856 ± 16.497b
28 94.229 ± 44.212a 63.088 ± 13.410b
31 101.378 ± 39.610a 66.210 ± 14.395b
34 95.440 ± 38.459a 61.882 ± 12.662b
37 101.012 ± 39.048a 64.692 ± 13.810b
40 97.869 ± 36.783a 63.328 ± 15.498b
43 93.440 ± 35.475a 61.567 ± 12.931b
46 105.062 ± 33.219a 62.493 ± 9.618b
aAngka – angka pada kolom yang sama diikuti dengan huruf yang sama tidak berbeda nyata
15 Secara umum, berdasarkan nilai dugaan r yang diperoleh ini, maka apabila dihitung nilai laju pertumbuhan terbatasnya (), maka akan dihasilkan < 1. Hasil ini menunjukkan bahwa populasi A. glycines kedelai kontrol pada semua kelas umur cenderung meningkat. Hasil analisis ragam menunjukkan bahwa pada taraf nyata 5% ukuran subcontoh bootstrap tidak memberikan pengaruh yang berbeda terhadap dugaan r A. glycines kedelai kontrol. Dengan demikian dapat dinyatakan bahwa untuk pendugaan r baik menggunakan persamaan Lotka-Euler maupun matriks Leslie dapat menggunakan ukuran subcontoh yang minimum apabila ukuran ini dapat memudahkan peneliti hama dalam menduga statistik demografi hama.
Berdasarkan Tabel 3, contoh jackknife menghasilkan SK 95% bagi r yang lebih sempit dari contoh bootstrap pada berbagai ukuran subcontoh. Hal ini mengindikasikan bahwa nilai dugaan galat baku contoh jackknife lebih kecil dari nilai dugaan galat baku contoh bootstrap. Dengan demikian untuk pendugaan r, metode resampling yang sebaiknya digunakan adalah metode jackknife. Akan tetapi, jika ingin menggunakan metode bootstrap maka gunakan ukuran subcontoh yang sesuai dengan ukuran data aslinya, karena pada saat ukuran subcontohnya sama dengan ukuran data aslinya dihasilkan nilai dugaan galat baku yang minimum.
SK 95% bagi T hasil resampling boostrap dan jackknife juga ditunjukkan oleh Tabel 3. Nilai dugaan T hasil resampling bootstrap dan jackknife memberikan hasil dugaan yang konsisten dengan dugaan galat baku yang cukup kecil. Hasil analisis ragam menunjukkan bahwa pada taraf nyata 5% ukuran subcontoh bootstrap tidak memberikan pengaruh yang berbeda terhadap dugaan masa generasi A. glycines
kedelai kontrol. Dengan demikian dapat dinyatakan bahwa untuk pendugaan masa generasi dapat menggunakan ukuran subcontoh yang minimum apabila ukuran tersebut dinilai dapat memudahkan penelitian hama dalam menduga statistik demografi hama. Kemudian, berdasarkan Tabel 3, juga dapat dilihat bahwa nilai dugaan galat baku contoh bootstrap pada saat ukuran subcontohnya sama dengan 46 lebih kecil daripada nilai dugaan galat baku contoh jackknife. Dengan demikian, untuk pendugaan mendapatkan penduga T yang lebih baik hasil resampling
Tabel 3 SK 95% bagi r dan T A. glycines kedelai kontrol hasil resampling bootstrap dan jackknife
Contoh ra Ta
Jackknife 0.537 ± 0.016 7.720 ± 0.209
Bootstrap
19 0.538 ± 0.022a 7.760 ± 0.286b
22 0.540 ± 0.023a 7.656 ± 0.305b
25 0.539 ± 0.022a 7.705 ± 0.240b
28 0.537 ± 0.023a 7.709 ± 0.271b
31 0.546 ± 0.023a 7.679 ± 0.299b
34 0.536 ± 0.022a 7.694 ± 0.257b
37 0.539 ± 0.020a 7.727 ± 0.202b
40 0.534 ± 0.017a 7.763 ± 0.182b
43 0.538 ± 0.020a 7.659 ± 0.194b
46 0.536 ± 0.017a 7.720 ± 0.197b
aAngka – angka pada kolom yang sama yang diikuti dengan huruf yang sama tidak berbeda nyata
16
boostrap dan jackknife yaitu dapat menggunakan metode bootstrap dengan ukuran subcontoh sama dengan ukuran subcontoh aslinya.
Pendugaan Statistik Demografi A. glycines Kedelai Aplikasi PGPR
Tabel 4 memperlihatkan bahwa SK 95% bagi GRR dan R0 berdasarkan contoh
jackknife dan bootstrap pada berbagai ukuran subcontoh. Nilai dugaan GRR contoh
bootstrap konsisten dengan dugaan galat baku yang cukup kecil. Pada tabel tersebut juga ditunjukkan bahwa nilai dugaan R0 pada SK 95% relatif konsisten tetapi dugaan galat bakunya masih cukup besar. Hasil analisis ragam menunjukkan bahwa pada taraf nyata 5% ukuran subcontoh bootstrap tidak memberikan pengaruh yang berbeda terhadap hasil dugaan GRR dan R0 A. glycines kedelai aplikasi PGPR. Dengan demikian dapat dinyatakan bahwa untuk pendugaan GRR dan R0 melalui metode bootstrap dapat menggunakan ukuran subcontoh yang minimum apabila ukuran subcontoh minimum ini dinilai dapat memudahkan penelitian hama dalam menduga statistik demografinya.
Pada Tabel 4 juga terlihat bahwa untuk pendugaan GRR, nilai dugaan galat baku contoh bootstrap saat ukuran subcontoh sama dengan 41 lebih kecil dari nilai dugaan galat baku contoh jackknife. Sedangkan pada pendugaan R0, nilai dugaan galat baku minimum dihasilkan dari contoh jackknife. Jadi, berdasarkan hasil pendugaan GRR, metode bootstrap akan menghasilkan nilai dugaan yang lebih baik dari jackknife apabila digunakan ukuran subcontoh yang sesuai dengan ukuran contoh data aslinya. Sedangkan untuk pendugaan R0, secara umum metode
jackknife menghasilkan dugaan yang lebih baik dari metode bootstrap.
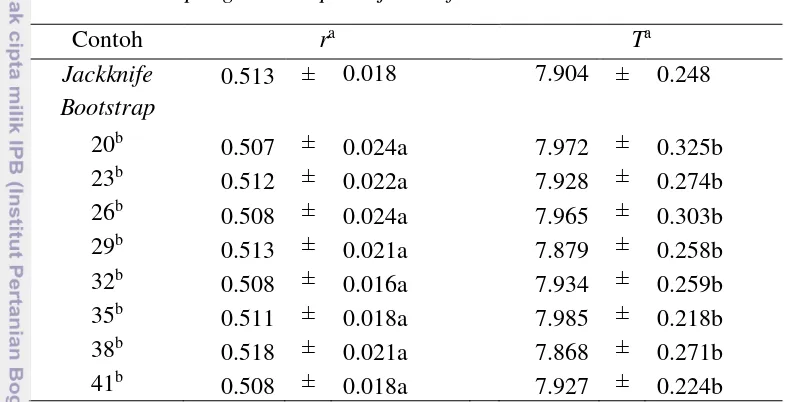
SK 95% bagi r hasil resampling bootstrap dan jackknife yang ditunjukkan oleh Tabel 5. Sama seperti pada nilai dugaan r A. glycines kedelai kontrol, nilai dugaan r A. glycines kedelai aplikasi PGPR yang dihitung melalui optimasi persamaan Lotka-Euler juga menghasilkan nilai dugaan yang sama dengan melalui matriks Leslie. Nilai dugaan r hasil resampling bootstrap dan jackknife
memberikan hasil dugaan yang konsisten dengan dugaan galat baku yang sangat kecil. Nilai dugaan galat baku contoh jackknife sama dengan dari nilai dugaan galat Tabel 4 SK 95% bagi GRR dan R0 A. glycines kedelai Aplikasi PGPR hasil
resampling bootstrap dan jackknife
Contoh GRRa R0a
Jackknife 71.834 ± 6.551 57.780 ± 8.998
Bootstrap
20 71.965 ± 8.549a 56.713 ± 9.579b
23 72.264 ± 7.108a 57.996 ± 12.030b
26 72.129 ± 6.772a 57.283 ± 10.557b
29 70.303 ± 5.642a 56.703 ± 9.166b
32 71.477 ± 7.960a 56.407 ± 10.285b
35 72.267 ± 6.366a 59.182 ± 9.284b
38 71.915 ± 7.109a 58.728 ± 10.754b
41 70.581 ± 5.192a 56.371 ± 10.203b
aAngka – angka pada kolom yang sama diikuti dengan huruf yang sama tidak berbeda nyata
17 baku contoh bootstrap saat ukuran subcontohnya 41. Akan tetapi hasil dugaan r
contoh jackknife menghasilkan bias yang lebih kecil daripada hasil dugaan r contoh
bootstrap. Hasil analisis ragam menunjukkan bahwa pada taraf nyata 5% ukuran subcontoh bootstrap tidak memberikan pengaruh yang berbeda terhadap dugaan laju pertumbuhan intrinsik A. glycines kedelai aplikasi PGPR. Dengan demikian dapat dinyatakan bahwa untuk pendugaan laju pertumbuhan intrinsik dapat menggunakan ukuran subcontoh yang minimum apabila ukuran subcontoh ini memudahkan peneliti hama dalam menduga statistik demografinya.
Tabel 5 memperlihatkan bahwa nilai dugaan T contoh bootstrap memberikan hasil dugaan yang konsisten dengan dugaan galat baku yang cukup kecil. Nilai dugaan galat baku contoh bootstrap pada saat ukuran subcontohnya sama dengan 35 lebih kecil daripada nilai dugaan galat baku contoh jackknife. Dengan demikian, untuk pendugaan T dapat digunakan metode bootstrap dengan ukuran subcontoh sama dengan 35. Akan tetapi, seandainya digunakan ukuran subcontoh bootstrap
yang maksimum (41), nilai dugaan galat bakunya juga tetap lebih kecil dari nilai dugaan galat baku contoh jackknife. Hasil analisis ragam menunjukkan bahwa pada taraf nyata 5% ukuran subcontoh bootstrap tidak memberikan pengaruh yang berbeda terhadap dugaan masa generasi A. glycines kedelai aplikasi PGPR. Dengan demikian dapat dinyatakan bahwa untuk pendugaan masa generasi dapat menggunakan ukuran subcontoh yang minimum apabila ukuran ini bisa memudahkan peneliti hama dalam menduga statistik demografi suatu hama.
Setelah dilakukan metode jackknife dan bootstrap terhadap masing – masing data kohort A. glycines kedelai kontrol dan kedelai aplikasi PGPR, maka dilakukan uji-t terhadap kedua data tersebut. Uji-t ini bertujuan untuk mengetahui laju pertumbuhan intrinsik A. glycines kedelai aplikasi PGPR lebih kecil dari laju pertumbuhan A. glycines kedelai kontrol atau tidak. Hasil uji-t untuk masing – masing r hasil resamplingjackknife dan bootstrap dapat dilihat pada Tabel 6. Hasil uji-t pada Tabel 6 menunjukkan bahwa pada taraf nyata 5%, laju pertumbuhan intrinsik A. glycines kedelai kontrol lebih besar dari laju pertumbuhan intrinsik A.
Tabel 5 SK 95% bagi r dan T A. glycines kedelai aplikasi PGPR hasil
resampling bootstrap dan jackknife
Contoh ra Ta
Jackknife 0.513 ± 0.018 7.904 ± 0.248
Bootstrap
20b
0.507 ± 0.024a 7.972 ± 0.325b
23b 0.512 ± 0.022a 7.928 ± 0.274b
26b 0.508 ± 0.024a 7.965 ± 0.303b 29b 0.513 ± 0.021a 7.879 ± 0.258b
32b 0.508 ± 0.016a 7.934 ± 0.259b
35b 0.511 ± 0.018a 7.985 ± 0.218b
38b 0.518 ± 0.021a 7.868 ± 0.271b
41b 0.508 ± 0.018a 7.927 ± 0.224b
aAngka – angka pada kolom yang sama yang diikuti dengan huruf yang sama tidak berbeda nyata
18
glycines kedelai aplikasi PGPR. Berdasarkan hasil uji-t ini maka dapat dinyatakan bahwa pemberian PGPR pada tanaman kedelei efektif menghambat laju pertumbuhan intrinsik hama A. glycines.
Tabel 7 menunjukkan waktu komputasi yang dibutuhkan untuk melakukan pendugaan statistik demografi A. glycines kedelai kontrol dan kedelai aplikasi PGPR dengan menggunakan metode bootstrap dan jackknife. Waktu komputasi diketahui berdasarkan keluaran pada jendela log di SAS 9.3. Berdasarkanhasil yang diperoleh pada Tabel 7, metode bootstrap membutuhkan waktu komputasi yang lebih cepat dibanding metode jackknife. Hal ini disebabkan replikasi dilakukan dalam metode bootstrap hanya 30 kali, sedangkan pada metode jackknife proses replikasi dilakukan sebanyak 46 kali untuk gugus data A. glycines kedelai kontrol dan 41 kali untuk gugus data A. glycines kedelai aplikasi PGPR.
Selain itu, pada Tabel 7 juga terlihat bahwa baik untuk metode bootstrap
maupun jackknife, waktu komputasi yang dibutuhkan pada proses pendugaan statistik demografi hama A. glycines kedelai aplikasi PGPR lebih cepat daripada waktu komputasi yang dibutuhkan pada proses pendugaan statistik demografi hama
A. glycines kedelai kontrol. Hal ini dapat disebabkan oleh perbedaan ukuran contoh kedua data tersebut. Ukuran contoh pada data kohort A. glycines kedelai aplikasi PGPR lebih sedikit dibandingkan data kohort A. glycines kedelai kontrol. Dengan demikian dapat dikatakan bahwa semakin kecil ukuran contoh datanya akan semakin cepat proses komputasinya.
Sedangkan jika dilihat dari teknik penghitungan dugaan statistik demografi hama A. glycines-nya, pendugaan melalui persamaan Lotka-Euler membutuhkan waktu komputasi yang lebih lama dibandingkan dengan pendugaan melalui matriks Tabel 7 Rataan waktu komputasi pendugaan statistik demografi A. glycines
hasil metode bootstrap dan jackknife pada berbagai jenis perhitungan
Data Metode Penghitungan Waktu (detik)
A. glycines kedelai kontrol
Jackknife Lotka-Euler 1639.580
Matriks Leslie 0.280
Bootstrap Lotka-Euler 1191.367
Matriks Leslie 0.236
A. glycines kedelai aplikasi PGPR
Jackknife Lotka-Euler 801.630
Matriks Leslie 0.180
Bootstrap Lotka-Euler 568.730
Matriks Leslie 0.161 Tabel 6 Uji-t efektivitas pemberian aplikasi PGPR terhadap laju pertumbuhan
intrinsik A. glycines
Metode H0 H1 Nilai-p
Jackknife rA≤rB rA > rB 0.000*
19 leslie. Hal ini disebabkan pada pendugaan melalui persamaan Lotka-Euler terdapat proses iterasi di dalam iterasi. Jadi, pada saat melakukan proses iterasi untuk metode
bootstrap ataupun jackknife dilakukan juga proses iterasi untuk mengoptimasi persamaan Lotka-Euler.
Pembuktian Terhadap Pendugaan Laju Pertumbuhan Intrinsik (r)
Berdasarkan subbab Model Matriks Pertumbuhan Populasi diketahui bahwa
M = Untuk k bilangan bulat positif ganjil, maka
|M- λI| = 1 -1 -2
Kemudian untuk k bilangan bulat positif genap, maka
20
Persamaan (21) disebut juga sebagai persamaan ciri dari matriks M. Sisi kiri dari persamaan tersebut hanya sekali berubah tanda, dan menurut “Aturan Tanda” dari Descartes, persamaan seperti ini paling banyak memiliki satu akar ciri positif yang riil. Selanjutnya Persamaan (21) juga dapat dituliskan sebagai berikut:
1
-Kemudian bagi kedua sisinya dengan λk+1 sehingga diperoleh:
-( 1)
Apabila Persamaan (22) dijabarkan maka diperoleh:
-1 -2 - - -1
0 0 1 1 -1 -1 1
k k
k k k k
F l F l F l F l .
Selanjutnya akan dijabarkan persamaan Lotka-Euler (Persamaan (11)) dengan mensubstitusi λ = er sehingga diperoleh:
Pada Persamaan (23) dan (24) terlihat bahwa pada λ yang sama terdapat koefisien – koefisien yang berbeda. Apabila koefisien tersebut diekuivalenkan maka akan diperoleh:
Secara umum dapat dituliskan menjadi:
dapat juga dituliskan sebagai berikut:
1
Persamaan di atas ini merupakan persamaan Lotka-Euler seperti yang dijelaskan pada bab Tinjauan Pustaka. Jadi, dengan p = lx (x+1/ )lx dan Fx p mx x 1
pada matriks Leslie kemudian dicari akar ciri dominannya (λ) maka diperoleh nilai dugaan r yang sama seperti menghitung dugaan r melalui pengoptimasian persamaan Lotka-Euler. Mencari akar ciri dari matriks M tentu akan sangat sulit jika dilakukan secara manual karena dimensi dari matriks tersebut sangat besar. (21)
(22)
(23)
(24)
21 Akan tetapi dengan bantuan perangkat lunak dalam komputer, hal ini akan menjadi sangat mudah dan sangat cepat bahkan jauh lebih mudah daripada mengoptimasi persamaan Lotka-Euler. Dengan hasil dugaan yang tepat sama, maka sangat direkomendasikan untuk menduga statistik demografi hama menggunakan matriks Leslie saja.
5
SIMPULAN
Berdasarkan hasil pendugaan statistik demografi A. glycines kedelai kontrol dan kedelai aplikasi PGPR melalui metode bootstrap dan jackknife dapat disimpulkan bahwa:
1. Perbedaan ukuran subcontoh pada resampling bootstrap tidak memberikan pengaruh yang berbeda terhadap nilai dugaan statistik demografi A. glycines. Ukuran subcontoh minimum yang digunakan pada penelitian ini adalah sebesar 19 dan 20. Untuk penelitian hama selanjutnya, para peneliti hama dapat menggunakan ukuran subcontoh ini untuk menduga statistik demografi hama melalui metode bootstrap.
2. Penggunaan PGPR pada tanaman kedelai dapat menurunkan laju pertumbuhan intrinsik hama A. glycines.
3. Pendugaan parameter laju pertumbuhan intrinsik (r) melalui teknik optimasi persamaan Lotka-Euler dan melalui matriks Leslie baik berdasarkan contoh
bootstrap maupun jackknife memberikan hasil dugaan yang sama.
4. Waktu komputasi yang dibutuhkan untuk menduga statistik demografi A. glycines dipengaruhi oleh beberapa hal, yaitu: teknik pendugaan, ukuran contoh data asli, dan jumlah replikasi pada metode resampling yang digunakan. Pendugaan statistik demografi A. glycines melalui matriks Leslie lebih cepat daripada melalui persamaan Lotka-Euler. Kemudian, Pendugaan statistik demografi A. glycines kedelai aplikasi PGPR lebih cepat daripada kedelai kontrol, karena ukuran contoh kedelai kontrol lebih banyak yaitu 46 individu sedangkan kedelai aplikasi PGPR hanya 41 individu. Selanjutnya, pendugaan statistik demografi A. glycines melalui metode bootstrap dengan jumlah replikasi sama dengan 30 lebih cepat dibandingkan dengan melalui metode
jackknife dengan replikasi sebesar 46 untuk data A. glycines kedelai kontrol dan 41 untuk data A. glycines kedelai aplikasi PGPR.
DAFTAR PUSTAKA
Agustini A. 2013. Pengaruh Plant Growth Promoting Rhizobacteria Terhadap Biologi dan Statistik Demografi Aphis glycines Matsumura (Hemiptera: Aphididae) pada Tanaman Kedelai [skripsi]. Bogor (ID): Institut Pertanian Bogor.
22
Begon M, Townsend CR, dan Herper JL. 2008. Ecology: From Individuals to Ecosystems. 4th edition. Oxford (UK): Blackwell Puslishing.
Chernick MR. 2008. Bootstrap Methods: A Guide for Practitioners and Researchers. Second Edition. New York (USA): John Wiley & Sons.
Efron B. dan Tibshirani RJ. 1993. An Introduction to the Bootstrap. London (UK): Chapman & Hall.
Efron B. 1982. The jackknife, the bootstrap, and other resampling plans. Philadelphia (PA): SIAM.
Hasibuan KM. 1987. Pemodelan Matematika di dalam Biologi Populasi Dinamika Populasi. Bogor (ID): Institut Pertanian Bogor.
Iskandar R, Mara MN, Satyahadewi N. 2013. Perbandingan Metode Bootstrap dan
Jackknife dalam Menaksir Parameter Regresi untuk Mengatasi Multikolinearitas. Buletin Ilmiah Matematika, Statistika dan Terapannya (Bimaster) 2(2):137–146.
Poole RW. 1974. An Introduction to Quantitative Ecology. USA: McGraw-Hill. Pratama Y, Prihandono B, Kusumastuti N. 2013. Aplikasi Matriks Leslie untuk
Memprediksi Jumlah dan Laju Pertumbuhan Suatu Populasi. Buletin Ilmiah Matematika, Statistika dan Terapannya (Bimaster) 2(3):163-172.
Price PW. 1997. Insect Ecology. 3th ed. New York (USA): John Wiley & Sons. Price PW, Denno RF, Eubank MD, Finke DI, Kaplan I. 2011. Insect Ecology
Behavior, Population and Communities. Cambridge (UK): Cambridge University Press.
Southwood TRE. 2000. Ecological Methods. 3th ed. New York (USA): Wiley-Blackwell.
Tarumingkeng RC. 1992. Dinamika Pertumbuhan Populasi Serangga. Bogor (ID): Institut Pertanian Bogor.
_______________. 1994. Dinamika Populasi Kajian Ekologi Kuantitatif. Jakarta (ID): Universitas Kristen Krida Wacana.
Untung K. 1993. Pengantar Pengelolaan hama Terpadu. Yogyakarta (ID): Andi Offset.
Zeng F, Pederson G, Ellsbury M, dan Davis F. 1983. Demographic statistic for pea aphids (Homoptera: Aphididae) on resistant and susceptible red clovers.
23 Lampiran 1 Contoh Data Kohort A. glycines kedelai kontrol
Individu\
Lampiran 2 Contoh Data Kohort A. glycines kedelai aplikasi PGPR
24
Lampiran 3 Tabel kehidupan A. glycines kedelai kontrol
Hari pengamatan umur (x) Peluang hidup (lx) Keperidian (mx) lxmx
0.5 0.935 0 0
1.5 0.913 0 0
2.5 0.913 0.238 0.217
3.5 0.913 1.095 1.000
4.5 0.913 4.500 4.109
5.5 0.913 6.024 5.500
6.5 0.913 6.476 5.913
7.5 0.913 7.833 7.152
8.5 0.848 6.615 5.609
9.5 0.804 8.568 6.891
10.5 0.804 7.243 5.826
11.5 0.804 4.514 3.630
12.5 0.783 5.556 4.348
13.5 0.761 3.714 2.826
14.5 0.739 3.618 2.674
15.5 0.674 2.742 1.848
16.5 0.609 2.179 1.326
17.5 0.500 1.696 0.848
18.5 0.413 1.684 0.696
19.5 0.304 1.857 0.565
20.5 0.261 2.083 0.543
21.5 0.174 2.000 0.348
22.5 0.152 2.143 0.326
23.5 0.109 1.400 0.152
24.5 0.087 3.250 0.283
25.5 0.065 4.333 0.283
26.5 0.043 5.500 0.239
27.5 0.022 0 0
28.5 0.022 0 0
29.5 0.022 0 0
30.5 0.022 0 0
31.5 0.022 0 0
32.5 0.022 0 0
33.5 0.022 0 0
34.5 0.022 6.000 0.130
35.5 0.022 2.000 0.043
25 Lampiran 4 Tabel kehidupan A. glycines kedelai aplikasi PGPR
Hari pengamatan umur (x) Peluang hidup (lx) Keperidian (mx) lxmx
0.5 0.951 0 0
1.5 0.951 0 0
2.5 0.927 0.211 0.195
3.5 0.902 0.622 0.561
4.5 0.902 3.568 3.220
5.5 0.854 5.743 4.902
6.5 0.854 7.029 6.000
7.5 0.854 8.029 6.854
8.5 0.854 6.857 5.854
9.5 0.854 6.543 5.585
10.5 0.854 7.257 6.195
11.5 0.829 6.147 5.098
12.5 0.805 4.788 3.854
13.5 0.732 4.467 3.268
14.5 0.707 3.138 2.220
15.5 0.683 2.429 1.659
16.5 0.634 1.538 0.976
17.5 0.610 1.080 0.659
18.5 0.463 0.737 0.341
19.5 0.268 0.636 0.171
20.5 0.195 0.625 0.122
21.5 0.171 0.143 0.024
22.5 0.098 0.250 0.024
23.5 0.024 0 0
24.5 0.024 0 0
25.5 0 0 0
Lampiran 5 Contoh sintaks SAS 9.2 prosedur jackknife untuk menghitung statistik demografi berdasarkan persamaan Lotka-Euler
proc iml;
use tesis.kontrol;
read all var {D1 D2 D3 D4 D5 D6 D7 D8 D9 D10 D11 D12 D13 D14 D15 D16 D17 D18 D19 D20 D21 D22 D23 D24 D25 D26 D27 D28 D29 D30 D31 D32 D33 D34 D35 D36
} into data; b=nrow(data); c=ncol(data);
start delrow(x,d);
26
print GRRbar R0bar rbar Tbar;
27 Lampiran 6 Contoh sintaks SAS 9.2 prosedur jackknife untuk menghitung
28
P=diag(px);
M=(F||0)//(P||nol); lamda=eigval(M); r[i]=log(lamda[<>]); T[i]=log(R0[i])/r[i];
end;
GRRbar=GRR[:]; R0bar=R0[:]; rbar=r[:]; Tbar=T[:];
print GRR;
print R0;
print r;
print T;
print GRRbar R0bar rbar Tbar;
quit;
Lampiran 7 Contoh sintaks SAS 9.2 pembangkitan data bootstrap
libname tesis 'B:\Kuliah\S2\TESIS\SAS';
data tesis.kontrol;
input ind D1-D37;
cards;
... ... ... ;
%macro bootstrap;
%do i=19 %to 46 %BY 3 ;
proc surveyselect DATA=tesis.kontrol METHOD=urs n=&i OUTHITS REP=30
out=boot30.kontrol_m&i;
%end;
%mend bootstrap; %bootstrap; quit;
Lampiran 8 Contoh sintaks SAS 9.2 untuk menghitung statistik demografi (r
iteratif) berdasarkan hasil bangkitan bootstrap
*Program untuk menghitung statistik demografi data contoh bootstrap dengan s=19;
proc iml;
use boot30.kontrol_m19;
read all var{D1 D2 D3 D4 D5 D6 D7 D8 D9 D10 D11 D12 D13 D14 D15 D16 D17 D18 D19 D20 D21 D22 D23 D24 D25 D26 D27 D28 D29 D30 D31 D32 D33 D34 D35 D36
29
print GRRbar R0bar rbar Tbar;