.
KERAGAAN PRODUKSI TERNAK DOMBA PROLlFlK
Oleh
:
ISMETH INOUNU
PROGRAM PASCASARJANA
INSTITUT PERTANIAN BOGOR
PRODUCTION PERFORMANCE OF PROLIFIC SHEEP ABSTRACT
Production and economic performance of ewes from three different
genotypes of prolificacy was evaluated from f 980 to 1993. Non-carriers
(Fed'FecJ'), carriers (FecJF~ecJ') and homozygotes (Fec.JFFecJF) for a gene
affecting prolificacy, averaged 1 .I 9; 2.12 and 2.96 of ovulation rate, respectively.
This difference in turn resulted in variability of ewe productivity and management
level required for each genotype. Genotype and Management interaction effect
was significant source of variation of embryonic suwivd (DHE) and of litter weight
at weaning
(8s).
While parity did not significantly affect DHE but significantlyaffected
8s.
Ewe gestation gain (PKB) was negatively related to DHE, on thecontrary it was positively related to BS. Phenotypic and genetic correfations between weights at birth, at weaning, at mating and at lambing were generally
positive. This variability in the ewe production as a result of the differences in
genotype and management level practiced were economically evaluated. The
result shows that an increase of management practiced toward better
management was followed by an increase of ewe production (BS) per head.
However, these increases required higher input, which was expressed in the total
production cost. Ewes with ~ e d ~ F e c J + genotype gained the highest gross
margin if high level of management was practiced, followed by FecJFFec~F
genotype. On the low level of management, ewes carrying the F e d F gene did
not show their superiority since they gained lower gross margin compared with
INOUNU,
bimbingan Harimurti Martojo sebagai Ketua, Asikin Natasasmita, Ahmad Ansori
Mattjik. Subandriyo dan Tjeppy
D.
Soedjana masingmasing sebagai anggota).Domba-domba dari
P.
Jawa terkenal kerena kemampuannya untukrnenghasilkan anak banyak (prolifik). Dengan jarak kelahiran delapan bulan,
maka dalam setahun domba-domba dari
P.
Jawa rnampu menghasilkan 2 ekoranak per induk.
Di Stasiun Pemuliaan Balai Penelitian Ternak di Cicadas, Kabupaten Bogor
dari tahun 1980 sampai dengan tahun 1990 telah dilakukan penelitian untuk
mencari penyebab tingginya angka keragamsn jumlah anak sekelahiran pada
domba-domba di P. Jawa. Batas atas jumlah anak sekelahiran ditentukan oleh
jumlah ovum yang diovulasikan oleh seekor betina. Hasil-hasil sebelumnya
membuktikan bahwa sifat reproduksi ternak domba ini dipengaruhi oleh gen
tunggal FedF, yang bekeja secara aditif, sehingga keragaman jumlah ovum
yang dilepaskan oleh seekor induk tergantung dari genotipe induk tersebut.
Kehadiran gen FecJf pada populasi dornba di Indonesia, mengakibatkan ternak-
ternak ini dapat dibagi menjadi tiga kelompok yaitu: ternak dengan genotipe
FecJFFedF (prolifikasi tinggi), ~ e c . J ~ ~ e d ' (prolifikasi medium) dan Fed'FecJ' (prolifikasi rendah). Hal ini mengakibatkan pula terjadinya variasi dalam jumlah
anak yang dilahirkan. Akibat adanya interaksi dengan lingkungan, ternyata gen
juga secara tidak langsung terhadap bobot lahir, mortalitas, bobot sapih, dan
urnur kawin. Dengan rnengetahui sifat-sifat reproduksi dan produksi sejak dini
akan dapat rnembantu dalam seleksi individu-individu calon bibit yang akan
dikernbangkan lebih lanjut, dan rnernpercepat pengeluaran ternak-ternak sisa
seleksi untuk digunakan sebagai ternak bakalan pada usaha penggernukan.
Penelitian ini bertujuan untuk rnengamati sifat reproduksi dan produksi
domba dengan prolifikasi rendah, sedang dan tinggi yang dikembangkan di Balai
Penelitian Ternak, Ciawi, Bogor dan yang telah diidentifikasi sejak usia muda.
serta untuk rnengetahui kapan suatu genotipe tertentu berproduksi paling tinggi
dan pada lingkungan yang bagaimana produksi tinggi tersebut dicapai.
Selanjutnya rnengingat untuk rnencapai produksi tinggi tersebut diperlukan input
yang berbeda-beda pada masing-masing genotipe, rnaka penelitian ini juga
rnencari genotipe rnana yang paling rnenguntungkan ditinjau dari segi ekonorni.
Dari hasil penelitian ini dan dwi hasil pembahasan maka dapatlah ditarik
kesimputan sebagai berikut:
Rataan laju ovulasi (LO) adalah 1.96 buah per induk, keragarnannya
dipengaruhi oleh interaksi genotipe dan manajernen. Induk-induk dengan
genotipe FecJFFecJF lebih responsif terhadap perubahan msnajemen. Satu
duplikat gen F e d F dapat meningkatkan LO sebanyak 0.79-1 . I buah tergantung
rnanajernen yang diterapkan. Kenaikan bobot badan induk sebanyak satu kg.
diiringi dengan kenaikan LO sebanyak 0.03 buah.
Keragarnan daya hidup ernbrio (DHE) dipengaruhi oleh interaksi genotipe
manajemen. Peningkatan manajemen ke arah yang lebih baik menurunkan
angka DHE. Hal ini erat kaitannya dengan kapasitas tampung uterus yang
terbatas pada angka 3.06 buahhnduk.
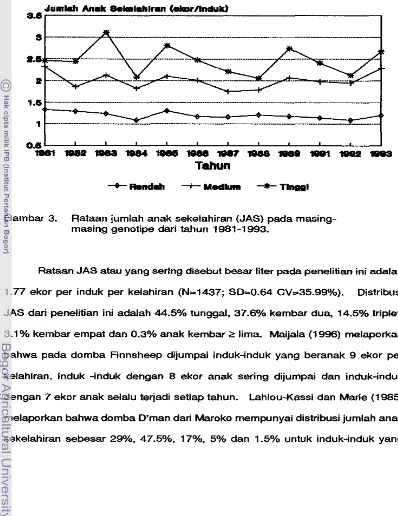
Rataan jumlah anak sekelahiran (JAS) yang didapatkan adalah sebesar 1.77 ekor per induk. Keragaman JAS dipengaruhi oieh genotipe dan interaksi manajemen dengan paritas induk serta pertambahan bobot badan induk (PKB).
Kehadiran satu duplikat gen F e d F dapat meningkatkan JAS sebanyak 0.8 ekor per induk. Pada indukinduk yang baru pertama kali beranak dihasilkan JAS 5.5% lebih rendah dibandingkan induk-induk yang tefah beranak dua kali.
Peningkatan satu kg bobot badan induk (PKB) diikuti dengan meningkatnya JAS
sebanyak 0.04 ekor.
Rataan total bobot badan anak saat lahir (BL) adalah 3.43 kg. Keragaman
B L dipengaruhi oleh genotipe dan interaksi rnanajemen dengan paritas induk. lndukinduk prirnipara (beranak pertama kali) menghasilkan E L lebih rendah dibandingkan indukinduk yang tefah beranak dua kdi.
Rataan daya hidup anak prasapih (DHA) didapatkan sebesar 73.9396,
keragamannya dipengaruhi oleh interaksi genotipe dan manajemen, paritas induk
dan pertambahan bobot badan induk. Pada paritas pertarna induk mempunyai
DHA 11% lebih rendah dibandingkan pada paritas kedua. Kenaikan pertambahan bobot badan induk sebesar 1 kg meningkatkan DHA sebesar 2.3 persen.
Keragaman totai bobot badan anak saat sapih
(6s)
dipengaruhi olehinduk (PKB). Raban
BS
yang didapat adalah13.12
kg.Bobot badan betina saat kawin (BK) adalah 23.49 kg. interaksi
manajemen dan genotipe di samping paritas induk mernpengaruhi keragarnan
BK. Bobot betina saat pertama kdi kawin adalah 20.55 kg. Flataan Bobot betina
saat beranak (BB) adalah 26.40 kg atau 11% lebih tinggi dibandingkan BK.
Bobot betina saat pertama kaIi beranak (88) adalah 24.41 kg. Keragarnan 85
dipengaruhi oleh interaksi rnanajemen dan paritas induk.
Makin dekatnya jarak umur antara sifat bobot badan yang diamati akan
diikuti dengan kian tingginya angka korelasi fenotipik. EL rnempunyai korelasi fenotipik dan genotipik yang tinggi dengan BS.
Peningkatan manajernen ke arah yang lebih baik diikuti dengan
peningkatan produksi bobot sapih per induk, narnun demikian perbaikan
rnanajemen ini menuntut peningkatan input yang tercerrnin dari kian
meningkatnya total biaya produksi. Induk-induk dengan genotipe Fec~~FecJ'
menghasilkan rnarjin kotor paling tinggi apabila manajemen tinggi diterapkan,
disusul ofeh induk-induk dengan genotipe FecJFFecJF. Pada manajemen rendah
induk-induk karier gen ~ e o J ~ tidak tampak keunggulannya karena rnernpunyai
rnarjin kotor yang lebih rendah dibandingkan induk-induk nonkarier.
Dari kesimpulan di atas dapatlah disarankan agar pernilihan induk-induk untuk pengembangan usaha disesuaikan dengan kondisi rnanajernen yang akan
diterapkan dan genotipe induk yang akan dikernbangkan, karena masing-masing
genoitpe induk rnernerlukan input yang berbeda untuk mencapai hasil yang
KERAGAAN PRODUKSI TERNAK
DOMBA
PROLlFlK
Oleh :
ISMET H lNOUNU
Disertasi sebagai
salah
satu syaat untuk memperoleh gelar Doktor pada ProgramPascasarjana, lnstihrt Pertanisn Bogor
PROGRAM PASCASARJANA fNSTlTUT PERTANIAN BOGOR
Narna Mahasiswa Nomor Pokok
Prof. Dr. Asikin Natasasrnita Anggots
1. Kornisi Pembimblng
4
Martolo
Ketua
Dr. Ir. Subandrlvo. M.Sc. Anggota
Keragaan Produksi Temak Domba Prolink
lsmeth rnounu 91513
Dr.
lr.
Ahmad Ansori Mattilk\
Awgo-Dr.
Ir.
T i e ~ p v 0. ~gediana. M.Sc. Anggota2. Ketua Bidang Keahiian
flmu
TernakKATA PENGANTAR
Penelitian mengenai keragaan produksi dan ekonomi ternak domba pada
tiga genotipe berbeda dilakuksn dilokasi Cicadas dan Bogor yang merupakan
stasiun pemuliaan Wai Penelitian Ternak. Ciawi-Bagor. Penelitian ini bertujuan
untuk mendapatkan informasi rnengenai potensi ternak domba yang
reproduksinya dipengaruhi oleh gen prolifik (gen F ~ c J ~ ) , dimana sebagian ternak
dapat beranak banyak dan sebagian lagi beranak tunggal, serta kondisi fluktuasi
manajemen yang rnengakibatkan terjadinya perbedaan produktivitas ternak dari
masingmasing genotipe ternak. Hal tersebut menjadi topik dari penulisan
disertasi ini.
Ide penulisan disertasi ini datang dari Prof. Dr. G.E. Bradford dari
Universitas California Davis di Arnerika Serikat, yang juga sekaligus mensponsori
biaya kutiah penulis (melalui program kerjasarna "SR-CRSP) di Program
Pascasarjana IPB, sejak dari program S2 sarnpai program S3 saat ini. Untuk itu penulis rnengucapkan banyak terima kasih atas segala bantuan materif maupun
moril, yang selama ini penulis rasakan. Beliau dengan tidak bosan-bosannya
rnendorong penulis untuk melanjutkan studi sarnpai kejenjang akadernik yang
paling tinggi.
Seorang bapak pemuliaan yang tidak hanya rnernuliakan ternak tetapi lebih
banyak mendidik manusia adalah Bapak Prof. Dr. Harirnurti Martojo. Beliaulah
yang menyambut penulis untuk hadir di Iingkungan IPB. Dimana pada lingkungan
kemampuan seseorang, walaupun penulis dalam keadaan patah semangat, beliau
mengarahkan penulis untuk dapat melanjutkan studi bahkan kejenjang yang tidak
pernah penulis irnpikan. Semoga yang Mahakuasa membaias segala kebaikan
beliau dan mernberi kekuatan kepada beliau untuk terus berkarya.
Penulisan disertasi ini diarahkan pula oleh Prof. Dr. Asikin Natasamita
sebagai seorang ahli dalarn bidang produksi ternak ruminansia kecil, beliaulah
yang mengarahkan penutisan desertasi ini sehingga bobot ilmiahnya semakin
terasa. Dr. A. A. Mattjik sangat berperan dalam pengarahan penulisan disertasi
ini terutarna dafam Mdang statistik. Dr. Subandriyo yang penulis kenal sejak
tahun 1981, membantu penulis tidak saja dari bidang pemuliaan tetapi juga dari
bidang statistik serta mernberi kemudahan untuk mendapatkan kepustakaan.
Penulisan ini menjadi lebih iengkap setelah Dr. T. D. Soedjana rnengarahkan
penulis untuk rnelakukan tinjauan ekonornis. Untuk itu penulis mengucapkan
banyak terima kasih atas bimibingan dan saran dari Sapak-bapak sekalian.
Tanpa izin dari Bapak Kepala Balai Penelitian Ternak, tidaklah mungkin
penulis dapat melanjutkan studi di IPB ini, untuk itu penulis rnengucapkan terima kasih atas kesempatan yang diberikan kepada penulis untuk menyelesaikan studi ini.
Terirna kasih penulis sampaikan kepada teman-teman Ir. Bambang Setiadi
xi
Pada kesempatan ini penulis sarnpaikan juga terima kasih kepada kedua
orang tua penulis yang telah mendidik penulis untuk rnerangkak, berjalan dan
beriari,
serta
tidak putus-putusnya mendorong penulis untuk berdiri kembalimanakala penulis jatuh terpuruk. Hanya yang Mahakuasalah yang dapat
memberi balasan kepadanya.
Kepada Nurhasanah Hidayafi, sebagai isteri yang mendampingi penulis
dalam kesulitan yang harus dilalui selama penulis rnenyelesaikan studi ini
diucapkan banyak terima kasih, juga kepada Aristogama dan Adiwicaksana
sernoga kalian dapat rnenyelesaikan studi kalian lebih baik dari penulis sebagai
bapakny a.
Akhir kata semoga karya tulis ini dapat menjadi bahan bacaan yang
berguna bagi mereka yang memeriukannya, serta sernoga dengan selesainya
studi penulis dijenjang akademik yang paling tinggi ini justru rnembuat penulis
semakin rendah hati dan lebih mengenal lagi akan kebesaran yang Mahakuasa
xii
Penulis dilahirkan di Bandung, pada tanggal 1 Januari 1955 sebagai anak
ketiga dari Bapak lsmu Athar dan Ibu 6. Zubaedah. Penulis rnenyelesaikan
Sekolah Dasar, Sekolah Menengah Pertama serta Sekolah Menengah Atas di
Bandung. Pada tahun 1980 penulis berhasil menyelesaikan pendidikan di
Fakultas Peternakan Universitas Padjadjaran (UNPAD), juga di Bandung. Di
Bandung pula penulis rnenikah dengan Ir. Nurhasanah Hidayati dan dikaruniai
dua orang anak lelaki, Aristogama yang lahir di Bogor pada bulan April 1982,
dan Adiwicaksana juga lahir di h g o r pada bulan September 1983. Penulis rnulai
bekerja sebagai pengajar tidak tetap go1 Ilb di Fakultas Peternakan UNPAD pada
tahun 1978. Setelah meyelesaikan studinya di S1 penulis bekerja di Balai
Penelitian Ternak Ciawi, sejak tahun 1981 hingga saat ini. Penulis mendapat
beasiswa dari "Small-Ruminant, Cotlaborative Research Support Project1*
Universitas California Davis dari Amerika Serikat pada tahun 1987 untuk
rnencapai gelar S2 di tPB, selanjutnya pada tahun 1991 penulis kembali
mendapatkan kesernpatan untuk rnelanjutkan studi kejenjang 53 di IPB Bogor
DAFTAR
I
S
1
Halaman
KATA PENGANTAR
. . .
ixRIWAYAT HIDUP
. . .
xiiDAFTARlSl
...
xiii. . .
DAFTARTABEL xv DAFTAR GAMBAR. . .
xix. . .
DAFTARLAMPIRAN xx PENDAHULUAN. . .
1. . .
TINJAUAN PUSTAKA 3. . .
Penemuan Gen Prolifik 3. . .
KeragaanReproduksi 7. . .
Kondisi Ternak Untuk Pasar Non Tradisional dan Ekspor 10. . .
Faktor-Faktor Yang Mempengaruhi Bobot Lahir 11. . .
Faktor-Faktor Yang Mempengaruhi Bobot Sapih 17 Pertumbuhan Pasca Sapih. . .
19Analisis Ekonorni
. . .
27MATERI DAN METODE
. . .
.
.
.
24Lokasi Penelitian
. . .
24Ternak
...
24Perkawinan Ternak
. . .
26Kelahiran Anak
. . .
27Tatalaksana Pemeli haraan
. . .
28Data
. . .
29Metode Analisis
. . .
31Ripitabilitas, Heritabilitas, Korelasi Fenotipik dan Genetik
...
33Metode Analisis Ekonomi
. . .
40HASIL DAN PEMBAHASAN
. . .
49Laju Ovulasi
. . .
49Daya Hidup Embrio
. . .
55Jumlah Anak seketahiran
. . .
6 7 Bobot Lahir Total Per lnduk. . .
68Daya Hidup Anak
. . .
74Bobot Sapih Total Per lnduk
. . .
83Bobot lnduk Pada Saat Kawin
. . .
88BobotBeranak
. . .
93Ripitabilitas, Heritabilitas, Korelasi Fenotipik dan Korelasi Genetik
.
.
98Ripitabilitas
dan
Heritabilitas. . .
98Uorelasi Fenotipik
. . .
100Korelasi Genetik 102 Fungsi Produksi 103 Biaya dan Penerirnaan
. . .
107...
xiv
. . .
KESIMPULAN DAN SARAN 114
DAFTARPUSTAKA
. . .
117Nomor Halaman
Teks
Domba-domba prolifik di dunia
. . .
Pertambahan bobot-badan harian (PBBH) pada domba Jawa dan target untuk mencapai bobot ternak komersil
. . .
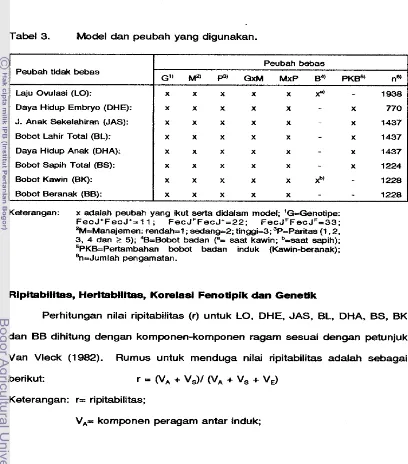
Model dan peubah yang digunakan
. . .
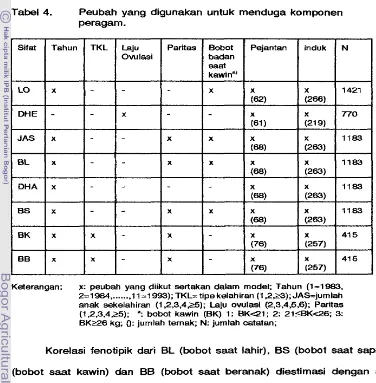
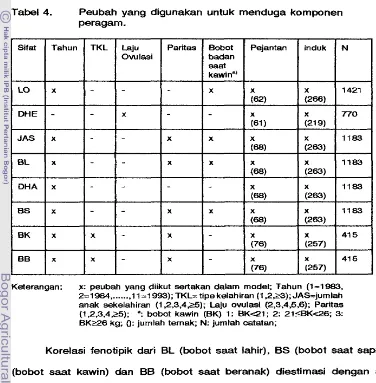
Peubah yang digunakan untuk menduga komponen peragam
. .
Analisis Peragam Saudara tiri seinduk
. . .
Jumlah konsumsi hijauan dan pakan tambahan pada tiga tipe kefahiran dan tingkat manajemen berbeda selama satu periode produksi (8 bulan)
. . .
Asumsi biaya dan penerimaan
. . .
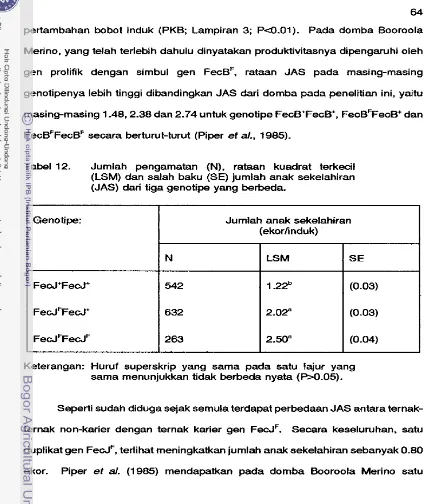
Jumlah pengamatan (N), rataan kuadrat terkecil (LSM) dan salah baku (SE) dari laju ovulasi (LO) pada tiga genotipe prolifikasi dan manajemen yang berbeda
. . .
Distribusi (46) anak kedalam kelompok genotipe berdasarkan kelasifikasi pejantan d m induk
. . .
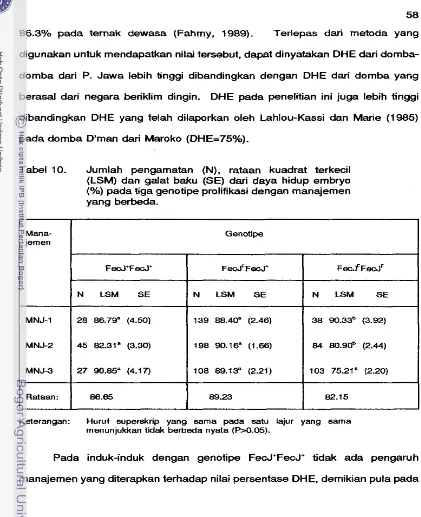
Jumlah pengamatan (N), Rataan kuadrat terkecil (LSM) dan Salah baku (SE) dari daya hidup embryo (%) pada tiga genotipe prolifikasi dengan manajemen yang berbeda
. . .
Daya hidup embryo
(DHE)
berdasarkan taju owlasi (LO) induk dan kapasitas tampung uterus (KTU). . .
Jurnlah pengamatan (N), rataan kuadrat terkecil (LSM) dan salah baku (SE) dari jurnlah anak sekelahiran (JAS) dari rnanajernen yang berbeda
. . .
Jumlah pengamatan (N), rataan kuadrat terkecil (LSM) dan salah baku (SE) dari jurnlah anak seketahiran (JAS) dornba pada
berbagai tingkat paritas
. . .
Jurnlah pengarnatan (N), rataan kuadrat terkecil (LSM) dan safah baku (SE) dari bobot lahir total (BL) pada tiga genotipe prolifikasi
berbeda
...
Jurnlah pengarnatan (N), rataan kuadrat terkecil (LSM) dan salah baku (SE) dari bobot lahir totaI (BL) pada rnanajernen berbeda
. .
Jurnlah pengamatan (N), rataan kuadrat terkecil (LSM) dan salah baku (SE) dari bobot lahir total (BL) domba pada berbagai tingkat
. . .
paritas
Jurnlah pengarnatan (N), rataan kuadrat terkecil (LSM) dan salah baku (SE) dari daya hidup anak (DHA) pada tiga genotipe prolifikasi dan rnanajemen yang berbeda
. . .
Jumlah pengamatan (N), rataan kuadrat terkecil (LSM) dan salah baku (SE) dari daya hidup anak (DHA) pada berbagai tingkat pantas
. . .
Jurnlah pengamatan
(N),
rataan kuadrat terkecil (LSM) dan salahbaku (SE) dari pertarnbahan bobot badan induk selama
kebuntingan (PKB) pada tiga genotipe prolifikasi dan rnanajernen yangberbeda
. . .
Persentase
(96)
total bobot lahir dari total produksi induk dari tiga...
genotipe prolifikasi pada rnanajernen yang betbeda
Jurnlah pengarnatan (N), rataan kuadrat terkecil (LSM) dan salah
baku (SE) dari bobot *h total (BS) pada tiga genotipe prolifikasi dan manajemen yang berbeda
. . .
Jurnlah pengarnatan (N), rataan kuadrat terkecil (LSM) dan salah baku dari bobot sapih total
(6s)
pada berbagai tingkat paritas. .
xvi
baku (SE) dari bobot kawin (BK) pada tiga genotipe prolifikasi dan manajemen yang berbeda
. . .
Jumlah pengamatan (N), rataan kuadrat terkecil (LSM) dan satah baku (SE) dari bobot kawin (BK) berdasarkan paritas induk
. . . .
Jurnlah pengamatan (N), rataan kuadrat terkecil (LSM) dan salah baku dari bobot saat beranak (BB) pada paritas induk dan manajernen yang berbeda
. . .
Nilai dugaan ripitabilitas (r) dan heritabilitas (h2) untuk sifat-sifat laju ovulasi (LO), daya hidup embrio (DHE), lama kebuntingan (LKB), jumlah anak sekelahiran (JAS), bobot lahir total (BL), daya hidup anak (DHA), bobot sapih total (BS), bobot kawin (BK) dan bobot
. . .
beranak (88)
Dugaan korelasi fenotipik (r,) antara bobot betina saat dilahirkan (BL), bobot saat sapih
(m),
bobot saat kawin (BK) dan bobot saat beranak (66). . .
Dugaan korelasi genetik (re) antara bobot betina saat dilahirkan
(BL), bobot saat sapih (BS), bobot saat kawin (BK) dan bobot saat beranak (65)
. . .
Nitai koefisien dugaan parameter yang diperoleh dari fungsi
. . .
produksi pada tingkat rnanajernen rendah (MNJ-1)
Nilai koefisien dugaan parameter yang diperoleh dari fungsi
. . .
produksi pada tingkat rnanajernen sedang (MNJ-2)
Nilai koefisien dugaan parameter yang diperoleh dari fungsi
. . .
produksi pada tingkat rnanajemen tinggi (MNJ-3)
Estirnasi marjin kotor usaha ternak dornba tipe kelahiran tunggal pada tiga tingkat rnanajernen
. . .
Estimasi marjin kotor usaha ternak domba tipe kelahiran kernbar
...
pada tiga tingkat manajemen
Estirnasi marjin kotor usaha ternak domba tipe kelahiran triplet pada
xviii
36. Distribusi
(96)
tipe kelahiran berdasarkan genotipe ternak. . . .
.
1 1 3 37. Majin kotor (Rp) berdasarkan genotipe induk dan tingkatNomor Halaman
Teks
1. Rataan laju ovulasi (LO) pada masing-masing genotipe dari bhun
1983-1993
. . .
492. Rataan daya hidup embrio {DHE) pada masing-masing genotipe
. . .
dari tahun 1983-1993 57
3. Rataan jumlah anak sekelahiran (JAS) pada masing-masing
. . .
genotipe dari tahun 1981 -1 993 62
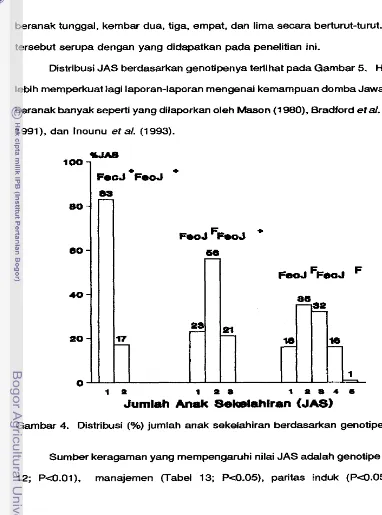
4. Distribusi (%) jumlah anak sekelahiran berdasarkan genotipe
. . .
635. Rataan total bobot lahii per induk (BL) pada masing-masing genotipe dari tahun 1981-1 993
. . .
696. Rataan daya hidup anak pra-sapih per induk (DHA) pada masing-
masing genotipe dari tahun 1981 -3 993
. . .
757.
Rataan total bobot sapih anak per induk (BS) pada masing-masing. . .
genotipe dari tahun 1981 -1 993
a4
8. Rataan bobot kawin induk (BK) pada masing-masing genotipe dari
tahun 1981 -1 993
. . .
889. Rataan bobot saat beranak (BB) pada masing-masing genotipe dari tahun 1981-1993
. . .
9410. Total bobot sapih per induk (kg) pada tiga manajemen (MNJ)
dengan tiga tipe kelahiran (TKL) berbeda
. . .
104Nomor Hataman
Teks
1. Jumlah pengamatan (N), Rataan nilai kuadrat terkecil (LSM) dan
Salah baku (SE) untuk Iaju ovulasi (LO).
. . .
1262. Jumlah pengamatan (N), Rataan nilai kuadrat terkecil (LSM) dan
Salah baku (SE) untuk Daya hidup embrio (DHE)
. . .
1283. Jurnlah pengamatan (N), Rataan nilai kuadrat terkecil (LSM) dan
Sdah baku (SE) untuk Jumlah anak sekelahiran (JAS)
. . .
3304. Jumlah pengamatan (N), Rataan nilai kuadrat terkecil (LSM) dan
Salah baku (SE) untuk Bobot lahir total (BL)
. . .
1325. Jumlah pengamatan (N), Rataan nilai kuadrat terkecil (LSM) dan
Salah baku (SE) untuk
Daya
hidup anak (DHA). . .
134 6. Jumlah pengamatan (N), Rataan nilai kuadrat terkecil (LSM) danSalah baku (SE) untuk Bobot sapih total (9s).
. . .
1367.
Jumlah pengamatan (N), Rataan nilai kuadrat terkecil (LSM) danSaiah baku (SE) untuk Bobot kawin (BK)
. . .
1388. Jumlah pengamatan (N). Rittaan nifai kuadrat terkecil fLSM) dan
Salah baku (SE) untuk Bobot beranak (66)
. . .
1409. Nilai usaha pembibitan dengan skala usaha 120 ekor induk
Domba-domba dari P. Jawa terkenal kerena kemampuannya untuk
menghasilkan anak banyak (prolifik). Mason (1 978) melaporkan prolifikasi domba
dari P. Jawa sebesar 170, 156 dan 136 persen. masing-masing untuk domba Priangan, domba ekor gemuk dan domba lokal ekor tipis. Dilaporkan pula bahwa
ternak-ternak ini mampu untuk beranak sepanjang tahun. Dengan jarak kelahiran
delapan butan, rnaka dalarn dua tahun seekor induk domba dari P. Jawa dengan
kelahiran tunggal mampu menghasilkan 3 ekor anak per induk atau 1.5 ekor anak
per induk per tahun.
Di Stasiun Pemuliaan Balai Penelitian Ternak. Cicadas. Kabupaten Bagor
dari tahun 1980 sampai dengan tahun 1990 telah dilakukan penelitian untuk
mencari penyebab tingginya angka keragaman jurnlah anak sekelahiran pada
domba-domba di P. Jawa. Saat ini penelitian masih dilanjutkan di Stasiun
Penelitian Ternak, lokasi Bogor. Batas atas jumlah anak sekelahiran ditentukan oleh jumlah ovum yang diovulasikan oieh seekor betina. Hasil analisa data
membuktikan bahwa sifat reproduksi ternak domba ini dipengaruhi ofeh gen
tunggal FecJF, yang bekerja secara aditif, sehingga keragaman jumlah ovum yang dilepaskan oleh seekor induk tergantung dari genotipe induk tersebut
(Bradford et a/.. 1991).
Kehadiran gen F e d F pada populasi domba di Indonesia, mengakibatkan
2
genotipe FecJF~ec.JF (prolifikasi tinggi), FedFFecJ+ (profifikasi sedang) dan
FecJ'FecJ' (prolifikasi rendah). Hal ini rnengakibatkan pula terjadinya variasi
dalarn jurnlah
anak
yang dilahirkan. Akibat adanya interaksi dengan lingkungan,ternyata gen F e d F ini tidak saja berpengaruh pada jurnlah ovulasi dan jurnlah
anak lahir; tetapi juga secara tidak langsung terhadap bobot tahir, mortalitas,
bobot sapih, dan umur kawin. Dengan rnengetahui sifat-sifat reproduksi dan
produksi sejak dini akan dapat rnernbantu dalarn seleksi individu-individu cdon
bibit yang akan dikernbangkan lebih fanjut, dan rnernpercepat pengeluaran ternak-
ternak sisa seleksi untuk dgunakan sebagai ternak bakalan pada usaha
penggernukan.
Penelitian ini bertujuan untuk mengamati sifat reproduksi dan produksi
dornba dengan prozifikasi rendah, sedang dan tinggi yang dikernbangkan di Balai
Penelitian Ternak. Ciawi Bogor dan yang telah diidentifikasi sejak usia rnuda,
serta untuk rnengetahui kapan suatu genotipe tertentu berproduksi paling tinggi
dan pada lingkungan yang bagairnana produksi tinggi tersebut dicapai.
Setanjutnya mengingat untuk mencapai produksi tinggi fersebut diperlukan input
yang berbeda-beda pada masing-masing genotipe, rnaka penelitian ini juga
Penemuan
Gen
ProlifikDornba-domba di
P.
Jawa dikenal rnernpunyai sifat keragarnan jurnlahanak sekelahiran yang tinggi. Banyak betina rnernpunyai anak satu atau dua
ekor saja, tetapi frekuensi betina-betina yang mernpunyai anak 3, 4 clan 5 ekor
lebih tinggi dari yang diharapkan untuk suatu populasi dengan rataan jurnlah anak
sekelahiran sekitar dua ekor (Bradford et a/., 1984). Laporan-iaporan dari peternak di pedesaan menyatakan bahwa pada induk-induk tertentu cendemng
untuk mempunyai anak banyak secara berulang. Catatan produksi dornba dari
Balitnak mernperkuat adanya pola beranak yang dernikian.
Kernatian anak dari tipe kelahiran 3 atau lebih sangat tinggi (fnounu et
al.,
1982), sehingga potensi keuntungan dari beiina dengan jurnlah anak banyak ini tidak terlihat. Betina-betina yang rnerawat anak banyak membutuhkan tingkat
nutrisi yang sangat baik pada fase sebelum dan sesudah kelahiran dan juga
rnernerlukan perawatan serta perhatian yang lebih banyak, jika persentase anak
hidup yang tinggi ingin dicapai. Apabila dalarn suatu kelompok ternak dapat
diprediksi jurnlah anak yang akan dilahirkan, hal tersebut akan sangat rnembantu
dalarn pengembangan rnanajernen yang efisien. Kelornpok ternak yang beranak
banyak akan lebih produktif pada kondisi pakan dan manajernen yang tinggi
sedangkan induk-induk beranak tunggal dapat beradaptasi pada kondisi pakan
4
berdasarkan jurnlah anak yang dilahirkannya menjadi sangat penting.
Pacia awalnya penelitian ternak domba di Bslitnak bertujuan untuk
mernpelajari potensi produksi ternak l o ~ 5 e s i a terutarna yang ada di
PJewa. Vntuk itu pada tahun
/
=
I
dibeli dornba d& Garut Jawa Barat- d
sebanyak 20 ekor betina dan 15 ekor pejantan, dari Sernarang Jawa Tengah 22
ekor betina dan 6 ekor pejantan, dan dari Grati Jawa Tirnur sebanyak 38 ekor
betina dan 5 ekor pqantan, sebagai tambahan terhadsp ternak yang telah ada
sejak tahun 1978 yaitu sebanyak
60
ekor betina dan 2 &or pejantan, yangsebelurnnya juga dibeli dari daerah Garut, Jawa Barat.
Catatan produksi sampai tahun 1982
&ri
ternak-ternak yang dibeli tersebutditambah catatan produksi dari ternak-ternak yang tefah ada di Stasiun
Percobaan sejak tahun 1978, rnenunjukkan adanya pola beranak yang tetap.
yaitu ada beberapa induk yang beranak banyak secara berulang dan ada pula
bebwapa induk yang selalu beranak tunggal. Hasil ini rnenarik perhatian Prof.
Bradford dari U.C. Davis yang berkunjung ke Balitnak pada saat itu. Beliau
meiihat ada kernungkinan reproduksi pada domba-domba ini dipengaruhi oleh gen
yang mirip dengan gen Booroola pada domba Booroola Merino. Sebagai
kelanjutannya pada bulan April-&lei 1983 difakukan pengarnatan terhadap Iaju
ovulasi ternak-ternak ini dengan teknik laparoskopi, sebanyak dua kali. Pada
saat itu jurnlah ternak yang tedibat adalah 90 ekor betina Garut. 37 ekor betina Ekor Gernuk dari Grati beserfa I 3 ekor anaknya, dan 18 ekor betina dari
tersebut diamati produksi sel telurnya (laju ovulasi) dengan
cara
menghitungjumlah "corpus luteum (CL)" yang dihasilkannya. Pengamatan sernentara menunjukkan bahwa dari 744 ekor betina yang dapat diamati produksi sel
telurnya sebanyak dua kali, ada 97 ekor betina yang rnenghasilkan jumlah CL
yang sama banyaknya antara birahi yang pertama dengan birahi yang kedua.
Dari penelusuran lebih lanjut, ternyata betina dengan CL yang tinggi juga berasal
&ti induk yang rnenghasilkan anak banyak, demikian pula betina dengan jurnlah
CL yang rendah berasal dari induk yang beranak sedikit. Berdasarkan hasit ini
Bradford
ef
a/.
(1 984) menduga adanya gen tipe Booroola yang mempengaruhilaju ovutasi pada domba-domba E3ooroola AAerino. Namun untuk membuktikannya
diperlukan tebih banyak data dan dengan garis keturunan yang jelas. Untuk itu
diiakukan perkawinan pada domba-dornba yang telah diiakukan klasifikasi
berdasarkan hasil laparoskopi (jumlah
CL)
dan catatan jumlah anak yang pernahdilahirkannya. Pada tahap ini klasifikasi pada k i n a baru dilakukan dengan cara
rnengelompokkan betinabetina yang mempunyai laju ovulasi .rendah (12) dan betina yang mempunyai laju ovulasi tinggi (23). Klasifikasi ini dilakukan pada
semua betina, baik yang berasal dari Garut, Semarang maupun yang berasai dari
Grati. Perkawinan dilakukan dengan mengawinkan kedua kelompok betina
tersebut dengan kelornpok pejantan dari Garut. Sedangkan klasifikasi pejantan
dilakukan dengan memperhatikan jumlah anak dari betina keturunannya, dan juga
dibeli beberapa pejantan lagi dari Garut dengan tipe kelahiran yang telah
6
ovuiasi dengan teknik yang sama dilakukan dua kali Iagi. Hasilnya lebih
menguatkan lagi dugaan akan adanya gen tunggal yang mempengaruhi
reproduktivitas pada domba-domba ini, dengan ripitabilitas untuk laju ovulasi
sebesar 0.80 d m untuk jurnlah anak sekelahiran 0.35 (Bradford et
aL,
1986)Pada tahun 1990 dalam suatu konferensi di Toulouse, Perancis, gen yang
mempengaruhi reproduktivitas pada domba dari Balitnak ini dinamakan gen
"Fecundity Javaw atau disingkat gen Fed; selanjutnya untuk gen pembawa sifat
jumlah an& banyak diberi notasi gen FedF, sedangkan untuk gen pembawa sifat
jumlah anak sedikit diberi notasi gen Fed' (Elsen et
a/.,
1991).Pada tahun 1993 Dr. J. Hetzel dari CSlRO Australia, tertarik untuk
mempetajari hubungan antara gen FecJ dengan gen FecB dari Booroola Merino, sehingga dilakukanlah kerjasama penelitian untuk mencari metoda yang tepat
untuk mendeteksi gen F e d ini. Hal ini dilakukan dengan cara mengawinkan
betina-betina dengan genotipe FecJ'FecJ* dengan pejantan FecJFFec4+ dan
betina-betina dengan genotipe ~ e c J ~ F e c ~ + dengan pejantan FecJ'FecJ',
kemudian diambil contoh darah dari hasii perkawinan ini dan juga dari tetuanya.
Penentuan genotipe keturunannya dilakukan dengan teknik laparoskopi sebanyak
empat kali, dengan kriteria yang sama seperti yang diajukan oleh Bradford
ef a/.
(1991). Hasil sementara dari analisis DNA dengan menggunakan metode yang
sama seperti yang dilakukan pada gen FecB, didapatkan adanya keterkaitan
antara penciri OarHH55 dengan FecJF, seperti halnya pada gen FecB
dalam tahap penyempurnaan metode yang tepat untuk mendeteksi gen F e d .
Apabila metode ini telah berhasil dengan baik maka untuk mendeteksi pqantan
karier gen FedF akan dapat dilakukan dengan mudah.
Keragaan Reproduksl
Batas atas keragaan reproduksi adalah jumlah sel telur yang dihasilkan
oleh seekor induk per satu siklus birahi, yang diamati dengan cara menghitung
jumlah "corpus luteum" (badan kuning) dari kedua indung tetur pada hari ke S t 0
setelah birahi. Faktor lingkungan merupakan faktor pendukung apakah batas
atas tersebut dapat dicapai.
Laju 0 W I ~ S I * dan JumIah Anak Sskelahlran: Laju ovulasi adatah rataan
jumlah sel tdur yang dihasilkan oleh seekor induk setiap siklus birahi. Di dunia
ini ada beberapa bangsa domba yang sangat prolifik, ditandai dengan laju owlasi
8 'Tabel 1 . Domba-domba proiifik di dunia'
1 : Bindon dan Piper (1 986).
i!.
Bradford ela/.
(1 986).Dari sekian banyak bangsa-bangsa dornba yang prolifik ini baru domba
Booroola Merino dan Domba Jawa (Priangan. Ekor Gemuk, dan lokal dari
Semarang) yang telah diteliti secara intensif penyebab tingginya laju ovulasi ini.
Pada dornba Booroola Merino dan Domba Jawa, temyata ada gen tunggal.
masing-masing gen FecB dan gen F e d F secara berturut-turut, yang
nlempengaruhi la@ ovulasi. Penemuan akan adanya gen tunggal ini
memudahkan kita untuk rneningkatkan penanganan ternak. Sehingga
perneliharan ternak dapat dilakukan berdasarkan genotipenya. Hal ini penting
karena pada ternak-ternak dengan prolifikasi tinggi diperlukan banyak pakan
tambahan untuk meningkatkan daya hidup anak. Apabila jurnlah pakan yang
siima diberikan pada domba non-karier, ha1 ini merupakan pernborosan yang
akan menurunkan keuntungan.
Dari beberapa pubtikasi (Hohenboken dan Clarke. 1981 ; Bradford, 1985) Jumlah anak
sekelahiran
Rataan Kisaran
2.6 1 -7
2.6
1-52.1
3-6
2.5 1 -7
2.6 1 -8
1.8 1-5
Bangsa
I
Laju OvulasiFinnsheep Romanov D'Man
Booroola Merino Cambridge Priangan2
Rataan Kisaran
3.5 1 -9
3.4 1-7
2.8 1 -8
4.2 1-1 1
4.0 1-13
diketahui bahwa peningkatan jumlah anak sekelahiran secara ekonomis
menguntungkan dibandingkan induk yang menghasitkan satu ekor anak saja
setiap kelahirannya. Bindon et
at.
(1984) mefaporkan bahwa kenaikan jumlahanak saat lahir 0.77 ekor per induk beranak, menghasilkan kelebihan keuntungan
sebesar 24%. Hal ini disebabkan total bobot liter anak saat sapih lebih tinggi
pada induk yang beranak kembar dibandingkan induk yang beranak tunggat.
lnounu et
d.
(I 993) melaporkan bahwa rataan bobot sapih dari induk yangmelahirkan tunggal, kembar dua dan tiga addah sebesar 10.5. 12.3 dan 13.1 kg
per induk secara berturut-turut pada kondisi lingkungan yang normal. Hal ini
belum memperfihatkan keunggulan dari induk-induk yang beranak kembar &lam
produksi anak sampai disapih. Namun, dengan peningkatan kondisi lingkungan
(peningkatan kualitas dan kuantitas pakan), bobot sapih tersebut meningkat
secara berturut-turut menjadi 15.2, 20.1 dan 24.4 kg per induk yang beranak
tunggal, kembar 2 dan 3. Pada kondisi dernikian baru terlihat keistirnewaan dari
induk-induk yang beranak banyak. Namun, keistimewaan dari domba Indonesia
tersebut belum banyak dipelajari dari segi ekonomi, sehingga belum merangsang
pengusaha untuk bergerak dalarn bidang usaha ternak domba. Penyebab lain
adalah rendahnya harga karkas, karena kuatitas karkas masih di bawah kualitas
karkas domba irnpor. Salah satu faktor yang mempengaruhi pertambahan bobot
badan dan kualitas karkas pada ternak domba adalah faktor genetik.
Untuk menangani ha1 tersebut di atas telah dilakukan persilangan betina
10
kualitas karkas yang tinggi sekaligus dengan jumlah anak yang banyak (Inounu
et al.. 1 995).
Kondlsi Temaic Untuk Pasar Non-Tradislonal Dan Ekspor
Pada Tabel 2 di bawah ini terlihat pertumbuhan domba Jawa (Inounu
et
al., 1993) dan target pertumbuhan untuk mencapai bobot ternak yang secara komersil diterima di pasar non-tradisional dan ekspor. Pada pasaran tersebut
yang dikehendaki adalah ternak domba dengan bobot badan 35 kg.
Tabef 2. Pertambahan bobot badan harian (PBBH) pada domba Jawa dan
target untuk rnencapai bobot ternak kornersil.
umber:
lnounu etat.
(1993). Bobot sapih pada umur 3 bulan. KarakterDari Tabei 2 di atas jelas terlihat bahwa untuk ternak-ternak yang
dilahirkan tunggal, tidak ada hambatan untuk mencapai target produksi tersebut,
karena pertambahan bobot badan harian (PBBH) sebesar itu banyak dilaporkan.
Masdah mulai timbul jika ternak dilahirkan lebih dari satu ekor per induk, karena Tipe Kebahiran
Tu nggal Kern bar Triplet
Bobot Lahir'
(Kg):
Bobot Sapih' (Kg): PBBH 0-3 bulan' (g):
Standar Bobot jual, 9 bulan (Kg):
I
Target PBBH 3-9 bulan(9):
2.60 1.83 1.43
15.20 10.50 8.1 0
140 96 74
35 35 35
pertumbuhan sebesar 134 dan 147 glhari untuk ternak kembar dan ternak triplet
akan sukar dicapai. Kalaupun dapat dicapai dengan pemberian pakan yang
bermutu tinggi akan sangat tidak ekonomis. Salah satu cara untuk pemecahan
masalah yang telah dilakukan adalah dengan mernperbaiki mutu genetiknya, yaitu
dengan
cross
breeding (terminalsire).
Faktor-faktor Yang Wmpengaruhi Bobot Lahir
Hansard dan Berry (1969) mengemukakan bahwa faktor-faktor yang
mempengaruhi bobot lahir seekor ternak adalah: intra-uterin (lingkungan fetus),
genotip induk, lingkungan induk, genotip anak, paritas, nutrisi, jenis kelamin, dan
umur induk.
Faknor
in-utetfn (lingkungan fetus> Pada spesiespofytocous
(beranakbanyak dafam satu kelahiran), peningkatan jumlah anak sekelahiran diiringi
dengan menurunnya laju pertumbuhan prenatal karena variasi dalam fungsi
plasenta dan lamanya kebuntingan. Keterlambatan pertumbuhan fetus pada
ternak yang berasal dari jumlah anak sekelahiran yang banyak semakin jelas
dengan bertambahnya umur kebuntingan. Untuk beberapa alasan yang tak jelas.
pengaruh jumlah anak sekelahiran pada bobot fetus tebih terfihat pada hewan
rodensia laboratorium dibandingkan pada babi. Lambatnya pertumbuhan pre-
natal tidak disebabkan ofeh pengaruh mekanik dari banyaknya fetus dalam satu
12
gizi darah dari induk. Namun, peningkatan konsumsi kalori pada induk tidak
mengakibatkan peningkatan yang nyata pada bobot fetus di atas batas normal
yang disebabkan oleh genotip fetus.
Pada tahap awal dari perkembangan fetus, pasokan nutrisi selalu cukup.
tetapi kemudian, saat fetus rnenjadi lebih besar dan tumbuh setiap hari, nutrisi
merupakan faktor utama yang sangat mempengaruhi pertumbuhan. Secara
umum dapat dikatakan bahwa jumlah pasokan darah induk dengan semua
kandungan gizinya yang mencapai fetus inilah yang menentukan laju
pertumbuhan fetus. Setelah lahir, bila makanan cukup, pengaruh genetik dari
bapak mulai tampak perannya, tetapi pada saat yang sama, karena lambatnya
laju pertumbuhan pada tahap akhir kebuntingan, nafsu makan akan diatur pada
tingkat yang sesuai dengan ukuran dan laju pertumbuhan seperti sebelum lahir
saat pusat nafsu makan (appetite centre) di hypothalamus sedang dikembangkan
(Widdowson dan McCane, 1975) dikutip oleh Widdowson (1980).
Plasenta adalah tempat fetus tumbuh dan berkembang sebelum
kelahirannya. Plasenta sering dihubungkan sebagai organ fetus. Karena semua
gizi yang rnencapai fetus diangkut oleh plasenta, sering diasumsikan bahwa
pertumbuhan prenatal ditentukan oleh plasenta, dan plasenta yang kecil
rnenghambat pertumbuhan prenatal. Namun mungkin pula bahwa fetus yang
kecil te jadi karena secara genetis mempunyai potensi pertumbuhan yang rendah
atau kemungkinan lain adalah pasakan makanan yang buruk dari induk akan
yang menjadi kecil karena buruknya pasokan makanan induk (Gruenwald, 1967).
Aherne (1966) melaporkan bahwa pada manusia ada hubungan alometrik
antara bobot plasenta dengan bobot fetus. Besar plasenta dapat dibatasi oleh
berbagai macam proses-proses maternal, dan kemudian keterbatasan plasenta
akan menghambat pertumbuhan fetus. Pengaruh maternal yang secara tidak
langsung ini diperantarai oleh plasenta. Pada spesies tertentu terdapat hubungan
yang erat antara ukuran pfasenta dengan bobot fetus. Pertumbuhan anak yang
kerdil, yang sering terlihat pada anak babi yang berasaf dari jumlah anak
sekelahiran yang tinggi, erat hubungannya dengan ukuran pfasenta yang kecil.
Secara anatomi plasenta bertindak sebagai sumber cadangan darah dalam
sirkulasi maternal. Aliran darah pada arteri uterin dapat rnenjadi rendah jika
terdapat banyak plasenta dalam satu tanduk uterin dibandingkan jika hanya satu
plasenta saja. Penurunan aliran darah &pat menyebabkan menurunnya pasokan
oksigen terutama pada piasenta-plasenta yang terletak lebih jauh jaraknya.
KeabnorrnaIan vaskuler plasenta tersebut seperti i n t r a ~ t e ~ n e transfusion
syndrome mempengaruhi secara nyata pertumbuhan prenatal.
Faktorgenerik Besarnya fetus banyak dipengaruhi ofeh genotip fetus dan
induk, dan fetus lain yang berada dalam uterus tersebut. Kontribusi maternal
terhadap keragaman ukuran fetus febih besar dibandingkan kontribusi paternal.
McCallum (1 898) dan Montgomery (1 962) yang dikutip oleh Widdowson
serat otot secara penuh. Pertumbuhan otot setelah lahir semata-mata
disebabkan oleh pembesaran (hypertrophy) dari serat-serat yang telah ada sejak
lahir. Penelitian yang serupa pada babi yang dilaporkan oleh Stickland,
Widdowson dan Goldspink (1975) yang dikutip oleh Widdowson (1980)
menunjukkan bahwa jumIah serat-serat dalam otot indikator pada kaki depan
(musculus flexor digiti minimi brevis) mempunyai jumlah yang sama antara
individu an&-anak sekelahiran, tetapi berbeda antara anggota anak sekelahiran
yang satu dengan anak sekdahiran lainnya. Hal ini menunjukkan bahwa jumlah
serat-serat otot ditentukan secara genetik, tetapi kita tahu pula bahwa ukuran dari
serat-serat otot tergantung dari makanan dan besar dari individu tersebut, dan
berapa banyak otot itu bekerja.
Pada domba ada kecenderungan bahwa pejantan tipe besar berasal dari
tipe kelahiran tunggal, sehingga secara genotip diduga domba tersebut termasuk
ternak non-karier (normaf) untuk gen prolifik. Apabila seleksi dilakukan hanya
berdasarkan bobot badan, ha1 ini akan berakibat menurunnya rataan jumlah anak
lahir pada peternakan tersebut. Untuk memecahkan masalah tersebut perlu
dilakukan tiga pola perkawinan, yaitu pada satu kelompok khusus untuk
mengembangkan ternak homozigot prolifik (FecdFFecJp, satu kelompok lagi khusus bagi pengembangan ternak hornozigot non-prolifik (FecJ'FecJ*), yang
ketiga adalah persilangannya, yakni ternak heterozigot ( F d F F e d + ) . Betina-
betina heterozigot inilah yang digunakan sebagai usaha ternak kornersil yakni
sehingga akan didapatkan jumlah anak seketahiran yang relatif banyak (2.1 1 ekor
per induk) dan mempunyai daya tumbuh yang tinggi, sehingga kombinasi dari
keduanya dapat mendukung tujuan usaha penggemukan (Bradford, 1993).
Selain itu, perturnbuhan prenatal yang besar erat hubungannya dengan
ukuran tubuh induk yang besar. lnounu et at. (1993) rnenganjurkan untuk
memilih calon induk dengan bobot minimal 23 kg pada
saat
perkawinan pertamauntuk domba ekor tipis. Sifat genetik dari bapaknya menentukan batas atas dari
ukuran anak saat lahir bila induknya besar, tetapi ukuran plasenta merupakan
faktor pembatas ukuran anak saat lahir apabila induknya kecil. Hal ini
disebabkan oleh nutrisi maternal dan bukan disebabkan oleh inherifan
sitoptasmik, ha1 ini terlihat dari penelitian dengan cara embrio transfer (Hafez,
1969~1). Hunter (1 956) mefakukan penelitian persilangan resiprokalantaradomba
bangsa besar Border Leicester dengan domba bangsa kecil Welsh. Hasilnya
terlihat adanya pengaruh maternal pada penyitangan resiprokal: anak domba
persilangan dengan induk besar fampak tebih berat dari anak domba persilangan
dan bangsa induk kecil. Perbandingan antara persilangan dan bangsa murninya
dari induk berbangsa besar dan kecil secara berturut-turut, menunjukkan bahwa
pejantan yang kecil sedikit pengaruhnya pada bobot lahir dibandingkan dari
pejantan yang besar yang dikawinkan dengan induk yang kecil.
Permanen atau tidaknya pengaruh maternal pada perbedaan ukuran-
ukuran tubuh anak dikemudian hari ditentukan oieh tingkat perkembangan fetus
16 secara penuh pada saat lahir, pengaruh maternal pada ukuran tubuh akan
rnenghiiang dengan bertarnbahnya urnur, walaupun perbedaan bobot masih
tarnpak sarnpai pada urnur beberapa bufan (Hunter, 1 956).
FBktor nutrlsl
nzaQemaI: Fetus sangat mengagumkan ditinjau dari segi nutrisi.Walaupun pada kondisi induk yang sangat kekurangan gizi, fetus tetap saja
turnbuh dan akan mencapai bobot fahir harnpir normal. Pada situasi yang
tertentu bobot fetus proporsional terhadap konsumsi kalori dari induk. Pada
dornba, kadar zat gizi pada akhir kebuntingan rnernpunyai pengaruh yang nyata
terhadap bobot lahir. Bila induk mengalami kekurangan gizi selarna sepetiiga
akhir kebuntingan, ia akan mernproduksi anak yang kerdil walaupun ia rnendapat
rnakanan yang baik selarna awal kebuntingannya. Sebaliknya kadar zat gizi yang tinggi pada akhir kebuntingan rnenghasilkan anak dengan ukuran yang normal
(Wallace, 1948). Pada sepertiga akhir kebuntingan variasi dalarn bobat fetus
rnenggarnbarkan perbedaan dari faktor genetik, jurnlah anak sekelahiran, status
gizi dan kesehatan dari induk. Bila induk rnendapatkan cukup rnakanan, bobot
tahir cenderung untuk mencapai batas atas dari kemarnpuan genetiknya. Ebbot
lahir dari anak tunggal dari induk domba yang diberi rnakanan tingkat tinggi pada
setengah akhir kebuntingan tidak lebih besar dari anak tunggal yang induknya
rnendapat kadar gizi yang sedang. Anak dornba jantan turnbuh lebih cepat
lebih tinggi (1 6% vs. 20%) pada sepertiga akhir kebuntingan rnenghasilkan anak
dengan bobot Iahir lebih tinggi terutama pada tipe kelahiran kernbar, demikian
pula dengan daya hidupnya. Gizi induk yang buruk selama akhir kebuntingan
akan rnenyebabkan rnenurunnya kandungan glikogen dalam otot fetus dan
terutarna hati dari fetus. Glikogen fetus yang disimpan secara normal &an
rneningkat pada akhir kebuntingan dan bertindak sebagai surnber energi segera
setelah lahir, sehingga gizi induk yang buruk dapat meningkatkan mortalitas
neonatal. Organ tubuh sangat rnudah terpengaruh oleh cekaman yang
diakibatkan oleh gizi saat laju pernbelahan sel rnencapai puncaknya. Organ yang
masak dini seperti otak dan sistern syaraf pusat (Central Nervous System) akan
sangat mudah terserang cekarnan gizi pada tahap awal kehidupan (Hafez, 1969).
lnounu
eta/.
(1 993) mdaporkan bahwa tidak ada keuntungan dari induk-induk yang melahirkan anak banyak, apabila tidak diiringi dengan peningkatan
kualitas dan kuantitas pakan. Dengan perbaikan pakan diharapkan bobot saat
lahir akan meningkat, yang pada gilirannya akan rneningkatkan daya hidup dan
dengan rneningkatnya daya hidup rnaka &an rneningkat pula produksi induk saat
sapih.
Faktor-faktor Yang Mempengaruhi Bobot Sapih
Bibit memegang peranan penting dalarn suatu usaha peternakan. Bibit
yang unggul secara genetik terkadang belurn tentu unggul pula secara ekonomis
18 yang diperlukan oleh suatu perusahaan peternakan adalah suatu program
pemuliaan yang mengarah kepada sifat-sifat yang secsra genetik unggul namun
tetap rnenguntungkan secara ekonornis. lnounu
ef
a/.
(1993) melaporkan adanyasifat unggul dalarn produksi anak pada domba yang berasal dari Pulau Jawa.
Dornba-dornba ini mempunyai sifat beranak &ngan tipe kdahiran yang berbeda.
Namun daiam kondisi normal. total produksi b o h t sapih anak per induk tidak
berbeda. Hal ini terutama disebabkan oleh kenaikan jumlah anak sekelahiran
yang selafu disertai dengan semakin rendahnya rataan bobot lahir per individu
anak (2.60, 1 -83 dan 1.50 kg secara berturut-turut untuk tipe kelahiran 1, 2 dan 3). Menurunnya bobot lahir anak menyebabkan lemahnya kondisi anak yang
diperburuk pula oleh kondisi tubuh induk yang buruk, pertambahan bobot badan
(PBB) induk dilaporkan kian rendah dengan meningkatnya jumlah anak
sekelahiran.
Menurunnya kondisi induk erat kaitannya dengan rendahnya produksi
susu, sehingga keunggulan induk yang prolifik tidak tampak karena secara urnum
produksi induk sampai saat sapih adalah sama untuk ketiga t i p kelahiran (sekitar
10 kg). Pertumbuhan pascanatal menurun karena terhambatnya perturnbuhan
prenatal, selanjutnya ternak yang mengalami keterlarnbatan perhrrnbuhan
prenatal mempunyai efisiensi penggunaan makanan yang buruk pada saat
pertu mbuhan pascanatal.
Hal ini menunjukkan perlunya manajemen pernberian pakan yang khusus
dan kuantitas pakan pada akhir kebuntingan, terutarna pada induk-induk yang
membawa anak kembar. Usaha ini telah menghasilkan anak dengan bobot lahir
yang lebih baik dengan daya hidup yang tinggi pula (Inounu et a/., 1993).
Kombinasi antara daya hidup dan bobot lahir inilah yang pada akhirnya akan
menentukan produksi akhir seekor induk pada saat sapih. Bagaimana
rnemperbaiki bobot tahir an& dari tipe kelahiran tinggi agar daya hidupnya tinggi beturn banyak dipelajari.
Pertumbuhan Pasca Sapih
Masalah yang dihadapi oleh usaha penggemukan adalah sangat
bervariasinya bobot ternak bakalan. Seperti telah diterangkan di atas,
bervariasinya bobot sapih ternak disebabkan oleh jumlah anak sekelahiran dan
bobot Iahir. Usaha untuk mendapatkan bobot akhir yang sama dari ternak
bakalan yang berumur sama namun berbeda tipe kelahiran dan bobot awalnya
telah dilakukan oleh Yulistiani et a/. (1 990) yang meneliti anak domba sapihan (90 hari) dari tip kelahiran tunggal disapih sebagai tunggal dan kembar dua disapih
sebagai kernbar dua. Dombadomba tersebut dikelornpokkan berdasarkan dua
golongan berat sapih, yaitu besar dan kecil. Dengan pemberian makanan secara
"ad libitum" didapatkan konsurnsi hijauan sebesar 16% dan konsentrat 84% dari
total bahan kering yang dikonsumsi dan dilaporkan bahwa perbedaan bobot
sapih yang terlihat pada awal percobaan juga terlihat pada akhir percobaan.
20 umumnya menghasilkan tubuh yang kecil saat dewasa, namun kurang gizi pada
fase lanjut dari periumbuhan yang kemudian diikuti dengan rehabilitasi dapat
menghasilkan tubuh yang normat. Hewan dan an*-anak yang kurang gizi
sesudah lewat periode awal yang kritis akan selalu lapar, mereka akan makan
dengan banyak apabila makanannya tersedia, bobot dan panjang tubuhnya akan
tumbuh dengan cepat sampai mendekati kurva perturnbuhan yang normal
sebelum fase kekurangan gizi. Kernudian mereka akan mengurangi makannya
dan laju pertumbuhannya mulai menurun. Ashworth (1969) tdah mdakukan studi
khusus tentang ini pada anakanak yang dipulihkan dari kekurangan protein-
kalori, demikian pula Harris (1 979) yang keduanya dikutip oleh Widdowson (1980)
melakukan hal yang sama pada tikus. Keduanya mendapatkan bahwa konsumsi
(intake) makanan yang sangat tinggi dan laju pertumbuhan selama periode
pemulihan berkorelasi tidak saja dengan perfumbuhan yang cepat tetapi juga
dengan peningkatan persentase Iemak tubuh di atas normal. Jika kekurangan
gizi ini demikian buruk dan berkepanjangan, seperti pada babi yang dipelihara
agar bobot tubuhnya tetap sekitar 5 1 0 kg untuk sdama 1.2 atau 3 tahun, maka
pemulihannya tidak lengkap dan apabila lebih lama lagi masa kurang gizi
tersebut, akan kian kurang komplit pemulihannya (Widdowson, 1973). Namun,
ditemukan pula bahwa makin panjang masa kurang gizi dan makin kurang komplit
pernulihan pertumbuhannya, ternak akan rnenjadi makin gemuk oleh lemak pada
saat pemulihannya.
telah ditentukan dalam empat organ-organ pada tikus hasil seleksi dan kontrol.
Ukuran dan jumlah sel meningkat pada gaiur b a r dan menurun pada gdur kecil
pada umur 6 minggu. Dengan menunda saat pengeluaran ternak ketahiran
kembar beberapa bulan lebih lama agar mencapai bobot tertentu yang s e r a
dengan ternak kelahiran tunggal, diduga kelebihan penerimaan dari biaya
produksi masih dapat terlihat. Hanya saja seberapa jauh hal ini masih menguntungkan bila ditinjau dari suatu sistem usaha ternak, masih perlu diuji di
lapangan, karena tujuan dari penggemukan tidak hanya untuk mendapatkan
bobot yang diinginkan, kuaIitas karkas tetapi juga perbedaan antara input dan
output. Konsumen tentu saja tidak menginginkan karkas yang mengandung
terlalu banyak lemak.
Analisis EkonomE
Dalam usaha peternakan domba ada beberapa faktor yang rnempengaruhi
keuntungan baik secara langsung maupun tidak langsung. Faktor alam yang
sesuai dengan habitat ternak yang dipelihara rnerupakan faktor yang secara tidak
langsung meningkatkan produktivitas ternak, yang pada gilirannya juga
merupakan faktor yang akan meningkatkan keuntungan. Bibit yang unggul dan
tidak terlalu mahal harganya juga akan menentukan besar keoilnya keuntungan
yang didapat. Faktor lain adalah ketersediaan serta harga pakan tambahan.
Apabila pakan tambahan sukar didapat dm mahal harganya maka sebaiknya
22
sejalan dengan hukurn ekonorni yang rnenuntut penggunaan input produksi
serninirnal rnungkin dengan pertarnbahan output yang rnaksirnal.
Anafisis ekonorni setiap usaha selatu rnemperhatikan parameter-parameter
yang terrnasuk di dalarn kelornpok penerirnaan dan pengetuaran. Dickerson
(1 970) rnenyatakan bahwa pengeluaran utama dari usaha peternakan sangat
tergantung dari tiga parameter biologis, yaitu produksi induk, reproduksi dan
perturnbuhan anak. Penerirnaan dari produksi induk per tahun salah satunya
dapat ditingkatkan rnelalui pernilihan bibit ternak yang tepat dengan lokasi usaha
atau dengan perbaikan mutu genetik ternak apabila pasar menuntut kualitas yang
tinggi. Dinyatakan bahwa produksi induk rnerupakan hasil multiplikasi dari jurnlah
induk, volume produksi per induk dan per unit nilai produk tersebut, sehingga,
penerirnaan ini dapat ditingkatkan melalui upaya peningkatan volume produksi per
induk sebagai akibat perbaikan rnutu genetik ternak.
Hubungan penerimaan dan pengeluaran rnerupakan suatu hubungan
antara ketuaran yang dihasilkan dengan suatu parameter faktor produksi. Sifat
hubungan antara satu penerirnaan dan pengeluaran ini bisa tetap, rneningkatkan
atau rnenurunkan produktivitas rnarjinal. Soedjana (1993) rnelaporkan bahwa
fungsi keuntungan usaha peternakan dornba dipengaruhi oleh suatu besaran
sifat-sifat biologis yang rnenentukan kuantitas bobot badan ternak seperti rataan
pertarnbahan bobot badan harian, bobot sapi h dan konsurnsi pakan. Dinyatakan
bahwa produktivitas rnarjinal rataan tarnbahan bobot badan harian ternak dornba
2.152,00 per ekor. Hal ini berkaitan dengan pengaturan panjang optimum
periode pemberian m a n . Produktivitas marjinal rataan botmt sapih yang
diperoleh adalah sebesar 19.46 kg, sehingga keuntungan maksimum yang
diperoleh adalah Rp.42.813.00. Hal ini rnenunjukkan bahwa penggunaan masing
masing tambahan unit pakan menghasilkan kenaikan bobot badan yang relatif
lebih kecil dibandingkan dengan kenaikan masukkan pakan sebslumnya.
Keberhasilan pengembangan peternakan domba di Indonesia akan sangat
ditentukan oleh potensi biologis dan ekonomi dari ternak itu sendiri, dan harus
ditunjang oleh faktor sumber daya alam yang memadai serta faktor sosial seperti
persepsi masyarakat terhadap usaha ternak yang bersangkutan. Hal ini erat
kaitannya dengan pengembangan konsep teknologi tepat guna untuk
MATERI DAN METODE
Lokasi Penelitian
Penelitian dilakukan di Stasiun Pemuliaan Ternak. Cicadas, Gunung Puteri,
28 km dari Bagor kearah Jakarta, dengan rataan suhu udara 33' C dan rataan
curah hujan 31 12 mm per tahun. Sarana yang tersedia adalah satu buah
kandang berukuran 8x30 mZ, dan dua buah kandang berukuran 8 x 15 m2,
dengan luas lahan 5.8 Ha, dengan fegetasi rumput gajah. Ternak
dikelornpokkan ke dalam ruang kandang berukuran 3x3 m2 dengan kepadatan
ternak 6-8 ekor per ruang. Sqak bulan Agustus 1990, lokasi ternak dipindahkan
ke Stasiun Penelitian Ternak Bogor, dengan rataan suhu udara 25" C dan rataan
curah hujan 4230 mm per tahun. Hal ini dilakukan karena sulitnya pengontrolan
ternak, adanya kendala jarak, serta sulitnya jalan masuk
ke
dalam lokasipenditian, yang berakibat seringnya terjadi ketelfambatan penyediaan pakan
tambahan. Pada lokasi Bogor, fasilitas yang tersedia relatif sama dengan Iokasi
Cicadas, dengan tuas kebun 1.8 Ha dengan vegetasi rumput raja dan lokasinya
sangat mudah dicapai, sehingga keterlambatan penyediaan pakan tambahan
dapat dikatakan tidak pernah terjadi.
Temak
Ternak
domba yang diamati pada penetitian ini bemsal dari Garut (Jawatahun 1981 di Stasiun Pemuiiaan Ternak Cicadas, Bogor, kemudian ternak-ternak
ini dipindahkan ke Bogor pada tahun t 990.
Sejak tahun 1 983, perkawinan diarahkan untuk pernbentukan galur
prolifikasi, dengan menggunakan pejantanpejantan dari Garut. Bradford et
a/.
(1991) mengklasifikasikan ternak-ternak ini kedalam kelompok genotipe
berdasarkan kriteria-kriteria sebagai berikut:
FecJ'FecJ' (prolifikasi rendeh): induk-induk yang tidak pernah mempunyai catatan menghasilkan
"corpus
lutwrn" (CL) atau jurnlah anak sekelahiran (JAS)> 2. Pada jumlah pengamatan 23 kali, rataan CL atau JAS I 1.7.
F ~ C J ~ F ~ ~ J ' (proliflkssi medium): induk-induk yang minimum mempunyai satu kali pengamatan menghasifkan CL atau JAS = 3 , atau mempunyai frekuensi
catatan CL atau JAS=2 yang tinggi. Pada jumlah pengarnatan 23 kaii, rataan CL
atau JAS s 1.7.
FecJFFec.JF Iproltflkasi tinggl): induk-induk yang minimum mempunyai satu kaJi pengamatan menghasiikan CL atau JAS = 4.
Kelompok ternak berdasarkan gaiur ini sering pula disebut sebagai
kelompok prolifikasi tinggi (FecJFFec8), sedang (FecJFFecJ+), dan rendah (Fd+FecJ*). Pencatatan jumlah CL dilakukan dengan teknik "taparoskopi"
secara periodik pacta hari ke 3
-
10 setelah induk menampakkan birahi.Berdasarkan kriteria tersebut di atas ternak dikelompokkan kedalam masing-
masing kelompok genotipe, untuk kemudian dikawinkan dalam masing-masing
26
Perkawinan ternak:
Sebelum tahun 1983 perkawinan ternak dilakukan secara kontinyu, untuk
mendapatkan sefang beranak 8 bulan, dengan demikian kelahiran ternak tersebar
dad bulan Januari sampai bulan Desember. Mulai pada tahun 1983, untuk
rnemudahkan pengumpulan data, dilakukan penyerentakan birahi dengan
rnenggunakan spons intra vaginal
Chronogesf
atau Repromap, yang dirnasukkanke dalam saluran vagina dan dibiarkan setama 14 hari. Pengecekan birahi
dimulai pada hari pencabutan spons. Ternak betina yang menunjukkan birahi
dapat diketahui dengan mempergunakan pejantan vasektomi. Setelah birahi
terdeteksi, ternak dikawinkan dengan pejantan yang telah ditetapkan, 3-1 0 hari
kemudian dilakukan pencatatan jurnlah CL dengan teknik laparoskopi untuk
mendapatkan angka laju ovulasi (LO). Setelah dilakukan perkawinan secara
individual selama dua rninggu, pejantan ditinggal di datam masing-masing
kandang selama dua minggu lagi untuk memberi kesempatan kawin bagi ternak-
ternak yang kembali birahi ataupun yang belum kawin. Ternak betina dijaga agar
tetap berada didalam kelompok yang sama seperti pada saat perkawinan sampai
beranak dan menyapih anaknya, ha1 ini dimaksudkan agar identifikasi nomor
pejantan tidak tertukar. Pada saat perkawinan ditakukan pencatatan nomor betina, nomor pejantan, tanggal kawin, dan bobot kawin (BK).
Untuk menghindari terjadinya perkawinan sekerabat {inbreeding)
perkawinan dilakukan dengan rnernperhatikan catatan tetua baik dari betina
komputer "R:BASEW, dengan program ini betina-betina yang mempunyai tetua yang sama dengan pejantan yang akan rnengawininya dapat dengan mudah
dideteksi, dengan demikian penempatan pejantan dapat dipertimbangkan kembali
agar tidak terjadi perkawinan sekerabat.
Sebelum dikawinkan ternak dicukur, dipotong kukunya dan diberi obat
cacing, ha1 yang sama dilakukan kembali pada saat kelahiran.
Kelahiran Anak
Pada saat kelahiran, induk dan anaknya dimasukkan kedalam sekat 1 x
1 m2, dan dibiarkan selama satu sampai tiga hari agar induk dan anak dapat
sding mengenal. Dalam kurun waktu tersebut, anak yang dilahirkan diidentifikasi
(diberi kalung nomor), dicatat polawarna tubuh, tipe telinga dan jenis kelaminnya,
serta ditimbang berat lahir individu yang kemudian dijumlahkan dengan bobot
lahir individu saudara sekelahirannya menjadi bobot lahir total (8L), dan dicatat juga berat induk (BB). lnformasi lain yang dicatat adalah jumlah anak lahir hidup (JAS) dan juga jumlah anak yang mati untuk mendapatkan daya hidup anak
(DHA). Ternak yang baru lahir diberi nomor identifikasi 5 digit, dua digit pertama menunjukkan tahun lahirnya, sdanjutnya merupakan nomor urut kelahirannya.
Sebagai contoh individu dengan nomor 96001 adalah ternak yang lahir pada
tahun 1996 dengan urutan lahir pertama Selanjutnya anak ditimbang setiap dua
minggu sekall, sampai urnur sapih dan dicaiat bobot sapih individu dan jumlah
Pada saat di sapih ternak-ternak ini diberi obat cacing.
Tatalaksana Pemelharaan
Di lokasi Stasiun Pemuliaan Ternak di Cicadas, 1 9 8 7 -1 989, ternak diberi
pakan konsentrat sebanyak 0-300 g/ekor/hari, dengan kualitas yang berbeda-
b?da (kisaran protein kasar 10-1 3%) tergantung dari ketersediaan dana pada saat
itu. Hal ini kemudian tercermin pada hasil penditian dari waktu ke waktu.
Hijauan rumput gajah diberikan sebanyak 2-3 kg/ekor/hari. Peningkatan jumlah
pakan penguat dilakukan pada saat rnengawinkan betina, dan saat akan beranak
sampai masa penyapihan (90 hari seteiah kelahiran), pada saat-saat demikian
jumlah pakan penguat yang diberikan adalah 400-500 g/ekor/hari.
Di tokasi Bogor. 1990-1 993, ternak mendapatkan hijauan rumput raja yang
telah dirajang, jumlah hijauan yang diberikan adalah 3 4 kglekorlhari, sedangkan
konsentrat komersial "GT 03" yang mengandung 16% protein kasar dan 68 %
TDN diberikan sebanyak 2.2
YO
dari total berat badan betina di dalamk~?lornpoknya. Perubahan jumlah konsentrat yang diberikan dilakokan pada saat
kebuntingan mencapai minggu ke 14, yaitu dengan meningkatkan sebanyak 110
gr/ekor/hari, berdasarkan asumsi induk akan turnbuh paling tidak sebanyak 5 kg
ssimpai dengan beranak. Pada minggu ke 4 setelah kelahiran, jumlah pakan kc~nsentrat induk ditingkatkan menjadi 2.5% dari b r a t badan. Haf ini dilakukan
urituk meningkatkan kondisi tubuh induk
agar
produksi susu tetap terjaga. PadaData
Data yang digunakan untuk penelitian ini diambil dari catatan produksi
mutai dari tahun 1987 sampai tahun 1993; dari lokasi Stasiun Pemuliaan Ternak
lokasi Cicadas, Kabupaten Bogor (1 981 -1 989) dan Stasiun Bogor (1 990-1 993).
Data tersebut meliputi data produktivitas induk mulai saat dikawinkan sampai
menyapih anak (90 hari setelah kelahiran), yang meliputi: laju owlasi (LO) yang
diamati dengan cara menghitung jumlah "corpus luteurn" dari kedua buah indung
telur dengan menggunakan teknik taparoskopi pada ternak-ternak betina sekitar
3-10 hari setelah siklus birahi dan Jumlah anak sekefahiran (JAS). Dari kedua
pengarnatan tadi dapat dilakukan perhitungan terhadap daya hidup embryo
(DHE). Daya hidup embryo (DHE) didefinisikan sebagai perbandingan jumlah
anak yang lahir dibagi dengan jumlah "corpus luteum" dari kedua indung telur
yang dicatat pada masa perkawinan dikalikan dengan 100 persen, dan hanya
terbatas pada induk-induk dengan laju owlasi febih besar dari satu. Hal ini
disebabkan pada ternak dengan LO satu buah tidak diketahui apakah pada saat
ternak melontarkan sel