Comm. Appl. Biol. Sci, Ghent University, 77/4, 2012 635
INVESTIGATION ON SOME BIOLOGICAL ASPECTS
OF CHRYSOPERLA LUCASINA (CHRYSOPIDAE:
NEUROPTERA) ON BEMISIA TABACI
IN LABORATORY CONDITIONS
A. BAGHDADI¹, F. SHARIFI² and A. MIRMOAYEDI² 1 Department of Agricultural Science, Payame Noor University
PO BOX 19395-3697, Tehran- IRAN
2 Department of Plant protection, Agricultural Faculty, Razi University, Kermanshah-IRAN Corresponding author E-mail: [email protected]
SUMMARY
Bemisia tabaci is one of the most important key pests of many types of cultivated plants. Lacewings (Chrysopidae: Neuroptera) are predatory insects, widely used in biological control programs. Between them green lacewing is a promising biological control agent of pests in green houses and crop fields. In this study, gravid females of the green lacewing Chrysoperla lucasina (Lacroix) were captured from Sarepolzahab ( altitude 540m, latitude 34°,14´ N 46°,9´ E) in western part of Iran. Collected insects were reared in a growth chamber, under experimental conditions (25±1°C, 70±5% RH and a photoperiod of 16:8 L: D).
Different diets were offered to larvae which consisted of a whitefly species B.tabaci, an aphid Myzus persica and also lyophilized powder of drone honeybee (Apis melifera). As different foods were used to nurish larvae, so for each diet, mean larval period were calculated, and finally means were compared to each other. Anova in MSTAT-C was used for analysis of variance, and Duncan multiple range test (DMRT) to compare between means.
The results showed that larvae had maximum duration of 27±0.33 days when fed on honeybee lyophi-lized powder and the minimum value was 17.9±0.3 days for B. tabaci. 25±0.27 day recorded for M. persicae.
Food preference of the 3rd instar larvae of green lacewing was surveyed, they showed a food preference to M. persicae, to compare with B. tabaci, as the former has a bigger body size, so more easily to be captured by the predator larvae.
The 3rd instar larvae of lacewing were more voracious on preys, than the 1st or the 2nd instar larvae. Statistically speaking, there were a significantly difference when mean of different preys consumed by predator larvae were compared. We found, that when the predator larvae have fed on B.tabaci, their development time was shorter, and when arrived to adult stage, the adults showed, an improved fertili-ty.
The results indicated that the suitable prey not only can increase the rate of through accelerating devel-opmental stages of the predator and by means of an increase in its pupal body weight consequently promoting the fecundity of resulting adults, but also can alter predators population density in relation to own production numbers.
Key words: Chrysoperla lucasina, Bemisia tabaci, Biological control
INTRODUCTION
Bemisia tabaci is a plant sap sucking insect belonging to the family Aleyrodidae. It is broadly polyphagous, feeding on an estimated 500 plants species (Greathead, 1986) from 60 plant families (Mound and Halsey, 1978, Jiang et al., 1999, Hilje et al., 2001). Since the early 1980’s
hon-636
eydew on which sooty moulds grow, reducing the photosynthetic capabilities of the plant, resulting in defoliation and stunting. B.tabaci is also a vector of 111 plant viruses, some of which are of high economic importance. Resistance to approximately 35 active ingredients has been reported for B.tabaci in at least 20 countries worldwide (Roditakis, 2005).
Chrysopids are polyphagous predators that can suppress population of many pest species (Hydron, 1971) such as whiteflies, thrips and leafhoppers (Plincipi and Canard, 1984). Their efficacy in biological control of whiteflies and aphids as well as other arthropod pests has been recognized for more than 250 years (Pappas, et al. 2011). These polyphagous predators can be easily reared on artificial diets and used for controlling agricultural insect pest (Cohen and Smith, 1998).
For every biological control, to be successful, primarily, it’s of outmost importance, to
evalu-ate the potential of the predator for targeted preys ( Al-Zyoud and Sengonca, 2004). In this study different diets offered to larvae and longevity of life for each larval instars and food preference were evaluated. Analysis of variation was done by using ANOVA and com-paring between means by Duncans multiple range test (DMRT), utilizing MSTAT-C software.
MATERIALS AND METHODS
Adult insects of C. lucasina were collected by netting from fields of sare-polezahab (altitude 540m, 34°, 14´N46°.9´E) in western part of Iran. Rearing was carried out in UPVC pipes
(height 20, diameter 15 cm) after species discrimination. 10% honey was used as diet for ovipositing adults, the eggs were collected on paper sheets.
Deposited eggs were collected daily and transported to cold chamber (5-6°C). Experiments started with transport of egg to growth chamber (25±1°C, 16L: 8D and 70±5% RH). First instar larvae hatched after an incubation period and reared on artificial diet after 1-12 hours. In rearing, different diets offered to larvae consisted of a whitefly species B. tabaci, an aphid M. persica and also lyophilized powder of drone honeybee (Apis melifera).
Consumption rate of different diets used in this study were carried out in 5 replications in each 2 treatment based on completely randomized design. Nymphs of B. tabaci and M. persicae were used as live. Larval development and nymph consumption rate was counted each 12 hours with considering different foods used. Longevity for each larval instar was evaluated. Analysis of variation and comparing between means carried by use ANOVA and duncan multiple range test (DMRT) respectively.
RESULTS AND DISCUSSION
The biology and behavior of C. lucasina larvae is dependent on the quality of food which they fed on. Consumption rate of different larval instars of green lacewing (C. lucasina) on B. tabaci and M. persicae showed maximum larval consumption. Analysis of variation showed maximum larval differences between preys at level of 1% (table 1a and 1b). The results of life history experiments showed that C.lucasina was able to develop and reach the adult stage by feeding on mean 444.2 and 281.8 third instars of B.tabaci and M. persicae, respec-tively.
Consumption rate of different larval instars is showed in Table 1a. The result shows signifi-cant difference between larval instars at 5% level. As it is seen from Table 1a, the highest feeding rate of the lacewing belonged to the 3rd instar larvae
Comm. Appl. Biol. Sci, Ghent University, 77/4, 2012 637
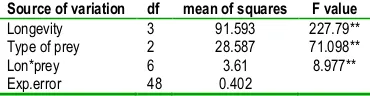
diets at level 1% ( table 2a and 2b). Longevity of larvae had maximum duration 27±0.33 days when fed by honeybee lyophilized powder and minimum 17.9± 0.3 days was recorded for B. tabaci. 25±0.27 days recorded from M. persicae. Alikhan and Wan (2004) reported 18.6 days for larval duration C. pallens feeding on B. tabaci.
Food preference of 3rd instars green lacewing larvae between live preys was investigated.
Results showed 70±1.7 and 27.6±2.86 for M.persicae and B.tabaci respectively.
When fertility of adults emerging from those larvae which fed on different diets was com-pared, those females who were fed on B.tabaci had a higher fertility rate, so we could con-sider, that for rearing in experimental purpose, it to be a more suitable host for lacewing larvae than other preys.
This study showed, that using larvae of green lacewing C. lucasina, is a very good way to control B. tabaci, although we used laboratory conditions, but potentially, it could be used in greenhouses and even in crop fields to control this pest.
Table 1a. Mean and standard error of consumption of different larval instars (C.lucasina) on live prey
larval instar B. tabaci M. persica
Instar1 66.2±4.78 26.2±1.24 Instar2 132±10.44 70.6±7.53
Instar3 246±5.33 185±6.94
sum of mean (x±SE) 444.2±6.8 281.8±5.2
Table 1b. Analysis of variance of consumption rate of different larval instar C.lucasina when were fed on live food
source of variation df mean of squares F. value
larval instars 2 74566.9 403.536** type of prey 1 21978.133 118.94** t.prey×l.instar 2 376.633 2.027n.s experimental error 24 184.783
Table 2a. Mean and standard error of longevity of different larval instars of C.lucasina in feeding on different diets.
Longevity of different stage of C. lucasina
B.tabaci M. persicae Honeybee lyophilized powder
Larval Instar1 2.7±0.12 4.3±0.19 4.8±0.35 Larval Instar2 3.3±0.19 4.5±0.22 5±0.31 Larval Instar3 7± 0.31 7.2±0.37 3.1±0.24
Pupa 8.8±0.68 9±0.31 10.2±0.35
Sum of mean 17.9±0.3 25±0.27 27±0.33
Table 2b. Analysis of variance of longevity of C.lucasina
Source of variation df mean of squares F value
Longevity 3 91.593 227.79**
Type of prey 2 28.587 71.098**
Lon*prey 6 3.61 8.977**
Exp.error 48 0.402
638
Table3a. Analysis of variation of prey type on fecundity of C. lucasina
Source of variation df mean of squares F value
type of prey 2 1288.8 52.718**
exp. error 12 195.167
** Significant difference at level 1%, c.v. 5.42%
Table3b. Mean and standard error of deposited egg of C. lucasina in feeding on different diets.
Mean of egg/female (20 days)
B. tabaci M.persicae Honeybee lyophilized powder
x-±S.E 291.2±6.47 275.6±4.56 206±7.37
ACKNOWLEDGEMENT
The authors thank Dr. Bahraminejad, of Dep. of Plant breeding, College of Agriculture, Razi University of Kermanshah for his assistance in statistical analysis, of data obtained in our study.
REFERENCES
Alikhan I., and Wan F.H. (2008). Life history of Chrysopa pallens (Rambur) Neuroptera, Chrysopidae) on Bemisia tabaci (Homoptera, Aleyrodidae) biotype B as prey. Sarhad journal of Agriculture. 24(4):635-639.
Al-Zyoud F. and Sengonca C. (2004). Prey consumption preference of Serangium parcesetosum sicard (Col: Coccinellidae) for different prey stages, species and parasitized prey. Journal of pest sci-ence.77:197-204.
Cohen A.C. and Smith L.K. (1998). New concept in artificial diets for Chrysopa rufilabris.The efficacy of solid diets. Biological content.13:49-54
Greathead A.H. (1986). Host plants. Chapter 3, pp.17-25.In: Bemisa tabaci-a literature survey on the cotton whitefly with an annotated bibliography (Ed.M.J.W.Cock).CAB International Institute of Bio-logical Control, Ascot, UK. 121 pages.
Hilje L., Costa H.S. and Stansly. (2001). Cultural practices for managing Bemisiatabaciand associated viral disease. Crop protection. 20:801-812.
Hydron S.B. (1971). Food preference of Chrysopa rufilabris in north central florida. M.S. Thesis, un-published, university of Florida Tallahassee, Florida.79 p.
Jiang Y.X., Lei H., Collat J.L., Martin B., Muniz M. and Fereres A. (1999). Probing and feeding behavior of two distinct biotypes of Bemisia tabaci (Hom: Aleyrodidae) on tomato plants. Journals of Economic Entomology. 92(2): 357-366.
Mound L.A. and Halsey S.H. (1978). Bemisia tabaci (Gennadius)In: Whitefly of the world, A Systematic Catalog of the Aleyrodidae (Homoptera)with Host plant and Natural enemy Data. British Museum (Natural History)and John Wiley and Sons, Chichester, New york, Brisbane,Toronto.pp.118-124. Pappas M.L., Broufas G.D. and Koveos D.S. ( 2011). Chrysopid predators and their role in biological
control. Journal of Entomology. 8(3):301-326.
Principi M.M. and Canard M. (1984). Feeding habits. In: Biology of chrysopidae, canard, M., Semeria Y. and T.R. New(Ed). Dr.w.Junk publishers, the Hague,The Netherlands.pp.76-92.
Roditakis E., Roditakis N. and Tsagkarakou A. (2005). Insecticide resistance in Bemisia tabaci (Homop-tera: Aleurodidae) populations from Crete. Pest Management Sciens. 61: 577-582.
Tauber M.J. Tauber C.A.andGardescuS. (1993). Prolonged storage of Chrysopa carnea(Neur.Chrysopidae) .Environmetal Entomology. 22(4):843-848.
Tauber M.J., Tauber C.A., Daane K.M. and Hagen K.S. (2000). Advances in commercialization of green lacewing (Neur.Chrysopidae). part2: field Applied