REPRODUKSI SEKSUAL KARANG (Ordo Scleractinia):
PEMIJAHAN, PERKEMBANGAN LARVA
DAN METAMORFOSA
SYAFYUDIN YUSUF
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI
DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Reproduksi Seksual Karang (ordo Scleractinia): Pemijahan, Perkembangan Larva dan Metamorfosa adalah karya saya dengan arahan dari komisis pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain sudah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Juli 2012
ABSTRACT
SYAFYUDIN YUSUF, Sexual Reproduction of Coral (Ordo Scleractinia): Spawning, Larval Development and Metamorphosis. Supervised by: NEVIATY PUTRI ZAMANI, M. ZAIRIN JUNIOR and JAMALUDDIN JOMPA.
Coral reef damage was exacerbated in several parts of Indonesia and only had 70% in good and very good conditions remaining. Destructive fishing and exploitation of ornamental corals have added to the scarcity of coral species. As a result the distance between the colonies of the species can inhibit the further reproduction between the colonies. However, the information pattern and timing of reproduction in natural coral is still limited, and reproduction efforts in exsitu methods were still in the research scale. This research was carried out at two separate locations namely Spermonde Archipelago and the Great Barrier Reef and was aimed to : (a) compare the pattern and timing of coral reproduction in Spermonde Archipelago Indonesia and in Orpheus Island of Great Barrier Reef Australia, (b) know the response time of spawning and spawning behavior of coral polyps caused by induction of spawning, (c) compare the fertilization, embryo development, competence time of species, (e) determine the appropriate light intensity, substrate position, CCA and zooxanthellae induction for larvae settlement, juveniles attachment dan metamorphosis of planula larvae. The results were coral reproduction pattern of small polyp corals were performed during rainy season but the bigger polyps were in dry seasons in Spermonde, while in the GBR was held in November and December during early summer. Synchronous broadcast spawning took place at 18:00–19:00 in lunar period 0 – 2 days AFM, while the high tide in Spermonde. Otherwise, multispecies mass spawning in GBR took place at 19:00-21:.00 for 3-5 AFM in the low tides. The water flow and strong aeration provided for A. tenuis was spawned earlier more than one hour than natural spawning. The same pattern of inter-species fertilization rate, more than 95% fertilization were achieved in less than 4 hours. In addition, the development of embryos and larvae were significantly differed between two species, i.e. embryos and larvae A.millepora grew faster than A.tenuis while Ctenactis crassa was faster than Fungia concina. Furthermore, the competence time of Acroporidae was longer than Fungiidae. Planulae larvae and juveniles of A. tenuis much more settled and attached while in higher light intensities of 130-170 μmol/m2/s. However, light intensity did not affect the total deposition of larvae and juveniles settled. Juveniles prefer attached at the horizontal substrate were significantly much more than those at vertical substrate. Induction of zooxanthellae and CCA separately could affect metamorphosis significantly. Larval metamorphosis of A. tenuis showed the same response to different concentrations of zooxanthella.
RINGKASAN
SYAFYUDIN YUSUF, Reproduksi Seksual Karang (ordo Scleractinia):
Pemijahan, Perkembangan Larva dan Metamorfosa. Dibimbing oleh : NEVIATY PUTRI ZAMANI, M. ZAIRIN JUNIOR, JAMALUDDIN JOMPA.
Tekanan yang berlebih terhadap ekosistem terumbu karang menyebabkan ekosistem ini tidak mampu memulihkan daya dukungnya. Akibatnya, kerusakan terumbu karang yang makin parah di berbagai kawasan di Indonesia dan hanya tersisa 30 % dalam kondisi yang baik dan sangat baik. Disamping perikanan yang merusak (destructive fishing), ancaman eksploitasi karang hias telah menambah kelangkaan populasi jenis karang tertentu. Akibatnya jarak antara koloni spesies tertentu makin jauh sehingga menghambat komunikasi reproduksi antar koloni. Pemulihan spesies terumbu karang membutuhkan waktu yang lama dan upaya yang cukup besar. Pemulihan alami tergantung pada kemampuan reproduksi populasi karang dari berbagai daerah sumber larva. Namun demikian, informasi pola dan waktu reproduksi karang di alam masih sangat terbatas, sehingga upaya reproduksi secara terkontrol masih dalam skala riset.
Penelitian ini dilakukan secara terpisah pada dua lokasi yakni di Kepualan Spermonde dan di Great Barrier Reef dan berlangsung secara sekuensial dalam 4 tahap, yakni tahap (1) observasi pemijahan, (2) induksi pemijahan, (3) perkembangan larva dan (4) metamorfosis larva. Penelitian observasi bertujuan : (a) membandingkan pola reproduksi dan waktu puncak pemijahan beberapa spesies karang di Kepulauan Spermonde Indonesia dan di Orpheus Island Great Barrier Reef Australia. Penelitian eksperimen bertujuan : (c) mengetahui respon waktu pemijahan dan tingkahlaku pemijahan melalui metode induksi fisik, (d) membandingkan tingkat fertilisasi, perkembangan embrio, competense time antar spesies, (e) menentukan intensitas cahaya yang sesuai bagi pengendapan dan penempelan larva planula (e) menentukan posisi substrat (vertikal atau horizontal) untuk penempelan larva planula, dan (f) mengetahui pengaruh induksi zooxanthella dan CCA terhadap metamorfosis larva planula.
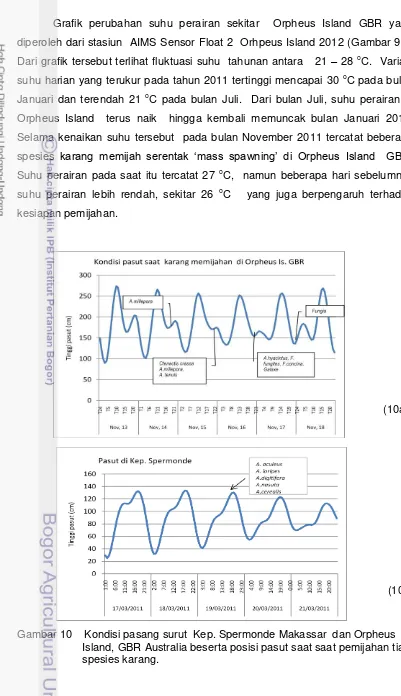
memijah pada suhu 27 oC, puncak dan lembah pasut kedua, dan curah hujan yang masih sedikit (awal musim hujan).
Penelitian tahap ke dua menitikberatkan pada ujicoba pemijahan secara induksi terhadap karang yang matang gonad. Hasil penelitian ini menunjukkan bahwa perlakuan induksi fisik pemijahan menggunakan air mengalir dan aerasi kuat mendapat respon yang positif dari karang Acropora tenuis. Dengan perlakuan tersebut, pemijahan karang lebih awal sekitar 2 jam sebelum terjadinya pemijahan secara alami atau normal baik di alam maupum di tempat penampungan induk laboratorium. Tingkahlaku pemijahan karang sebagai respon dari induksi pemijahan ini berlangsung normal, yakni fase setting, birth, glinding, upward dan brusting. Keseluruhan fase biologis pemijahan berlangsung selama 60 menit.
Penelitian tahap ke tiga difokuskan pengamatan proses biologi fertilisasi dan perkembangan embrio hingga larva planula. Hasil penelitian ini menunjukkan bahwa tingkat fertilisasi semua spesies memiliki pola yang sama hingga mendekati sempurna > 90% dalam kurun waktu 4 jam setelah pemijahan. Stadia sel blastula umumnya dicapai pada umur sel 10-17 jam. Perkembangan embrio dan larva karang Acropora tenuis dan A. millepora lebih cepat dibandingkan dengan kelompok Fungia concina dan Ctenactis crassa. Kelompok karang Acroporidae memiliki waktu berenang (competence time) lebih lama dibanding karang Fungiidae. Secara spesifik perkembangan embrio A. millepora lebih cepat dibanding A. tenuis, demikian halnya dengan Ctenactis crassa lebih cepat dibanding Fungia concina.
Penelitian tahap ke empat mengenai metamorfosa larva karang A. tenuis yang berlangsung di OIRS Jamescook University. Hasilnya memperlihatkan bahwa larva dan juvenil A. tenuis lebih banyak mengendap dan melekatan pada intensitas cahaya yang lebih terang dengan rentang 130-170 µmol/m2/s. Namun dari semua intensitas cahaya tidak mempengaruhi total pengendapan larva planula dan pelekatan juvenil. Preferensi penempelan juvenil lebih banyak di dalam substrat yang berposisi horizontal dan berbeda nyata dibanding substrat vertikal. Secara terpisah induksi CCA dan induksi zooxanthella mempengaruhi laju metamorfosis larva planula A. tenuis, namun kombinasi antara keduanya tidak berpengaruh. Metamorfosa larva A. tenuis menunjukkan respon yang sama terhadap perbedaan konsentrasi zooxanthella, tanpa zooxanthella dan CCA metamorfosa planula secara signifikan lebih rendah.
@ Hak Cipta milik IPB, tahun 2012
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruhnya karya tulis ini tanpa
mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk
kepentingan pendidikan, penelitian, penulisan karya ilmiah, penulisan
laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan
tersebut tidak merugikan kepentingan yang wajar IPB
REPRODUKSI SEKSUAL KARANG (Ordo Scleractinia):
PEMIJAHAN, PERKEMBANGAN LARVA
DAN METAMORFOSA
Syafyudin Yusuf
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor
pada Program Studi Ilmu Kelautan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji Luar Komisi Ujian Tertutup :
1. Prof. Dr. Dedi Soedharma, DEA. : Staf pengajar pada Fakultas
Perikanan dan Ilmu Kelautan Institut Pertanian Bogor.
2. Dr. Ir. Etty Riani, MS. Staf pengajar pada Fakultas Perikanan dan
Ilmu Kelautan Institut Pertanian Bogor.
Penguji Luar Komisi Ujian Terbuka :
1. Prof. Dr. Dedi Soedharma, DEA. : Staf pengajar pada Fakultas
Perikanan dan Ilmu Kelautan Institut Pertanian Bogor.
Judul
: Reproduksi Seksual Karang (ordo Scleractinia):
Pemijahan, Perkembangan Larva dan Metamorfosa
Nama
: Syafyudin Yusuf
NIM
: C 5610 700 41
Disetujui Komisi Pembimbing
Dr. Ir. Neviaty Putri Zamani, M.Sc. Ketua
Prof. Dr. M. Zairin Junior, M.Sc. Prof. Dr. Jamaluddin Jompa, M.Sc.
Anggota Anggota
Mengetahui
Ketua Program Studi DekanSekolah
Ilmu Kelautan Pascasarjana
Dr. Ir. Neviaty Putri Zamani, M.Sc. Dr. Ir. Dahrul Syah, MSc. Agr
PRAKATA
Gelar doktor merupakan gelar akademik tertinggi yang bisa diberikan oleh perguruan tinggi yang saya dapatkan saat ini di Institut Pertanian Bogor. Berbagai upaya dan doa telah dilakukan untuk menggapainya, sehingga dengan dengan kata bijak „tidak ada kata terlambat melainkan akan datang tepat pada waktu yang dijanjikanNya‟. Untuk itu saya patut bersyukur kepada Allah SWT yang telah menunjukkan jalan untuk sampai pada penyelesaian studi ini.
Disertasi ini berjudul Reproduksi seksual karang (Ordo: Scleractinia) : pemijahan, perkembangan larva dan metamorfosis. Salah satu subbagian yang akan dimuat dalam jurnal Indonesia Journal of Marine Science adalah „reproduction pattern and synchronous spawning of Acropora spp in Spermonde Archipelago, Makassar. Sementara tulisa ilmiah lain akan dimasukkan ke jurnal nasional dan internasional.
Dengan keberhasilan skala akademik ini, maka penulis inging menyampaikan ucapan terimakasih dan penghargaan yang setinggi-tingginya kepada :
Dr. Ir. Neviaty Putri Zamani, MSc., selaku ketua program studi Ilmu Kelautan dan sebagai komisi pembimbing yang telah memberikan perhatian, nasehat, motivasi, tenaga dan pikiran serta kesempatan kepada saya mulai dari rencana awal penelitian hingga akhir dari penulisan disertasi ini.
Prof. Dr. Ir. Muhammad Zairin Junior, MSc. dan Prof. Dr. Ir. Jamaluddin Jompa,MSc. selaku anggota komisis pembimbing yang telah memberikan arahan, bimbingan, motivasi dan pendampingan yang tulus sehingga mampu menjalani riset dan penyelesaian disertasi ini.
Prof. Bette Willis, ARC Center of Excellence Coral Reef Studies James Cook Australia, yang telah memberikan kesempatan kepada saya untuk bergabung dalam Coral Reproduction Project di Orpheus Island Research Station BGR Australia untuk menyelesaikan penelitian disertasi ini.
Rektor Institut Pertanian Bogor dan Dekan Sekolah Pascasarjana IPB yang telah memberikan kesempatan saya untuk mengenyam pendidikan program doktor dan program magister di Institut Pertanian Bogor. Rektor Universitas Hasanuddin dan Dirjen Dikti yang telah
Dekan dan Wakil Dekan Fakultas Perikanan dan Ilmu Kelautan Institut Pertanian Bogor yang telah memberi kesempatan kepada kami untuk belajar di Fakultas ini dan telah meberikan pelayanan terbaik khususnya dalam mempimpin ujian terbuka dan tertutup penyelesaian studi doktor. Prof. Dr. Dedi Soedharma, DEA., selaku penguji luar komisis
pembimbing pada ujian tertutup dan terbuka, Dr. Etty Riani, MSc. yang telah bersedia menguji saat ujian tertutup dan telah mengoreksi, mengarahkan, saran perbaikan disertasi ini.
Dr. Dirhamsyah, M.Sc. Pusat Penelitian Oseanologi yang telah bersedia diundang sebagai penguji tamu pada ujian terbuka promosi doktor. Rekan-rekan mahasiswa S2 dan S3 program studi Ilmu Kelautan IPB,
terkhusus rekan angkatan masuk 2007 IKL dan ITK, rekan-rekan pondokan PTD dan rekan asal unhas dan Sulawesi Selatan yang sama-sama merantau, rekan seorganisasi Wacana IPB, yang banyak saling membantu, memotivasi untuk bisa maju dan terus maju bersama hingga masuk finis meraih gelar doktor.
Orang tua (Yusuf Rahman, Alm.dan Hj. Mujijah) dan mertua (Abdurahman Karim dan Saadiah), adik-adik tercinta (Zainul Haris, Siti Nur Aini, SE, Suratman, Amd, Ssi, Surajiman, Amd) yang telah memberikan dukungan moril serta doa yang tak pernah putus untuk kesuksesan selama studi ini.
Istri tercinta Nunung Akhirany, SPt. MSi dan anakda tersayang Anugrah Samudra yang telah sabar menunggu selama masa studi dan banyak mengirimkan doa untuk papa selama pendidikan di Bogor dan di saat melakukan penelitian di laut. Terimakasih pula atas kesabaran dan pengorbanannya selama ini.
Disertasi ini akan ku persembahkan bagi keluarga, rekan, agama, bangsa dan negara dan siapa saja yang membacanya dan berterimakasih bila mendapat koreksi. Semoga karya ini memberikan inspirasi yang lebih besar untuk kemajuan ilmu pengetahuan kelautan Indonesia dan Internasional.
Bogor, 18 Juli 2012
RIWAYAT HIDUP
Penulis dilahirkan di Bima Nusa Tenggara Barat pada tanggal 19 Juli 1969 dari ayah Yusuf Rahman (Alm) dan ibunda Hj. Mujijah. Penulis adalah putra pertama dari 5 bersaudara. Pendidikan sarjana ditempuh di program studi Ilmu dan Teknologi Kelautan Universitas Hasanuddin Makassar dan tamat pada tahun 1995. Melanjutkan studi ke jenjang S2 pada tahun 2001 pada program studi Ilmu Kelautan Institut Pertanian Bogor, dan berhasil tamat pada tahun 2004. Pada tahun 2007 penulis kembali melanjutkan studi program doktor (S3) di IPB pada program studi yang sama atas biaya dari BPPS Dikti Kementrian Pendidikan nasional.
Penulis bekerja sebagai staf pengajar pada jurusan Ilmu Kelautan Universitas Hasanuddin dan sebagai peneliti pada Pusat Penelitian Terumbu Karang (Center for Coral Reef Research) Universitas Hasanuddin. Beberapa ekspedisi yang pernah diikuti antara lain, ekspedisi Karimunjawa bersama P3O-LIPI tahun 1994, ekspedisi Komodo tahun (1995), ekspedisi Taka Bonerate (1995) ekspedisi Togian –Banggai tahun 1998 dan 2000, hingga memimpin ekspedisi Wallacea 2006 di Teluk Tomini. Pada tahun 2011 yang lalu penulis berkesempatan mengikuti joint research ke Jamescook University Australia untuk meneliti reproduksi karang di Orpheus Island Research Station Great Barrier Reef.
DAFTAR ISI
Halaman
DAFTAR TABEL ...xxi
DAFTAR GAMBAR ...xxiii
DAFTAR LAMPIRAN ...xxv
PENDAHULUAN ………1
Latar Belakang ... 1
Rumusan Masalah ... 3
Tujuan Penelitian ... 5
Manfaat Penelitian ... 5
Ruang Lingkup Penelitian ... 5
Kebaruan Penelitian ... 9
Hipotesis penelitian... 10
METODOLOGI UMUM………11
Waktu dan Lokasi Penelitian ... 11
Metode Penelitian ... 13
POLA REPRODUKSI DAN SINKRONISASI PEMIJAHAN KARANG (Ordo Scleractinia) DI KEPULAUAN SPERMONDE INDONESIA DAN ORPHEUS ISLAND GREAT BARRIER REEF AUSTRALIA………..15
ABSTRAK ... 15
ABSTRACT ... 16
PENDAHULUAN ... 17
METODOLOGI PENELITIAN ... 18
HASIL PENELITIAN ... 24
PEMBAHASAN ... 39
SIMPULAN ... 42
SARAN ... 42
DAFTAR PUSTAKA ... 43
INDUKSI PEMIJAHAN KARANG Acropora tenuis UNTUK MENSTIMULASI RESPON POLIP MELEPASKAN GAMET………47
ABSTRAK ... 47
ABSTRACT ... 48
PENDAHULUAN ... 49
METODOLOGI PENELITIAN ... 50
PEMBAHASAN ... 64
SIMPULAN ... 68
SARAN ... 68
DAFTAR PUSTAKA ... 68
FERTILISASI DAN PERKEMBANGAN LARVA DALAM EMBRIOGENESIS KARANG (Ordo : Scleractinia) HASIL PEMIJAHAN EXSITU……….…..71
ABSTRAK ... 71
ABSTRACT ... 72
PENDAHULUAN ... 73
METODOLOGI PENELITIAN ... 74
PEMBAHASAN ... 96
SIMPULAN ... 101
SARAN ... 101
DAFTAR PUSTAKA ... 102
METAMORFOSIS LARVA KARANG Acropora tenuis (Dana 1846) DALAM KONDISI TERKONTROL………..105
ABSTRAK ... 105
ABSTRACT ... 106
PENDAHULUAN ... 107
MATERI DAN METODE ... 109
HASIL PENELITIAN ... 115
PEMBAHASAN ... 128
SIMPULAN ... 131
SARAN ... 131
DAFTAR PUSTAKA ... 132
PEMBAHASAN UMUM………134
KESIMPULAN UMUM………..146
DAFTAR PUSTAKA………147
DAFTAR TABEL
Halaman 1. Metode pengamatan gonad dan pemijahan spesies karang
di Kep. Spermonde Makassar dan Orpheus Island GBR... 20
2. Daftar spesies karang (tingkat klasifikasi) yang matang gonad dan yang memijah di Kepulauan Spermonde Makassar . ... 24
3. Pola musim dan bulan) kematangan gonad dan pemijahan karang di Kep.Spermonde dan Orpheus Island GBR. ... 26
4. Waktu pemijahan karang (masehi dan hijryah) di Kepulauan Spermonde Indonesia dan Orpheus Great Barrier Reef Australia. ... 28
5. Variabel lingkungan saat pemijahan dan matang gonad karang polip besar dan polip kecil di Kepulauan Spermonde Makassar ... 38
6. Respon waktu pada fase pemijahan pada perlakuan induksi pemijahan karang Acropora tenuis. ... 54
7. Analisis sidik ragan (satu arah) respon waktu pemijahan (menit) A. tenuis (General Linear model menggunakan Uji Tukey) ... 55
8. Respon waktu pada sitting dan glinding pasca sunset dari berbagai perlakuan induksi pemijahan dan pemijahan alami A. tenuis ... 55
9. Hasil pengamatan setiap fase pemijahan karang A. tenuis (15 November 2011) ... 58
10. Komparasi metode induksi pemijahan biota laut : kerang kima (Tridacnidae), kerang lola (Trochidae) dan karang (Scleractinia) ... 61
11. Metode stimulasi pemijahan karang (Scleractinia) dari berbagai sumber dan penelitian ini ... 63
12. Tingkat fertilisasi dari A. tenuis, A. millepora dan C. crassa ... 78
13. Analisis ragam tingkat fertilisasi karang spesies A. tenuis, A. millepora, Ctenactis crassa. ... 78
14. Tingkat pembelahan sel embrio karang A. millepora ... 86
15. Tingkat pembelahan sel embrio karang A.tenuis ... 87
16. Tingkat pembelahan sel embrio spesies Ctenactis crassa ... 89
17. Klasifikasi umur berdasarkan fase hidup embrio-larva-juvenil karang dan total competence time ... 90
18. Perbandingan umur larva dan competence time dari tiap spesies ... 92
19. Rata-rata dan rentang ukuran setiap fase embriogenesis masing-masing spesies karang ... 95
20. Analisis sidik ragam tingkhlaku pengendapan larva planula A. tenuis. ... 115
21. Proporsi pengendapan larva [rata-rata ± (SE)] pada intensitas cahaya dan selama waktu penelitian ... 117
23. Hasil analisis sidik ragam (ANOVA) perlakuan induksi CCA dan
Zooxanthella terhadap metamorfosa larva planula A. tenuis ... 123 24. Perbandingan laju metamorfosis larva planula A. tenuis antar
perlakuan Z3, Z6, Z9 ... 124 25. Perbandingan laju metamorfosis larva planula A. tenuis antar
DAFTAR GAMBAR
Halaman 1. Metode pengamatan gonad dan pemijahan spesies karang di
Kep. Spermonde Makassar dan Orpheus Island GBR ... 20 2. Daftar spesies karang (tingkat klasifikasi) yang matang gonad
dan yang memijah di Kepulauan Spermonde Makassar . ... 24 3. Pola musim dan bulan) kematangan gonad dan pemijahan karang
di Kep.Spermonde dan Orpheus Island GBR. ... 26 4. Waktu pemijahan karang (masehi dan hijryah) di Kepulauan Spermonde
Indonesia dan Orpheus Great Barrier Reef Australia. ... 28 5. Variabel lingkungan saat pemijahan dan matang gonad karang polip besar dan polip kecil di Kepulauan Spermonde Makassar ... 38 6. Respon waktu pada fase pemijahan pada perlakuan induksi
pemijahan karang Acropora tenuis. ... 54 7. Analisis sidik ragan (satu arah) respon waktu pemijahan (menit)
A. tenuis (General Linear model menggunakan Uji Tukey) ... 55 8. Respon waktu pada sitting dan glinding pasca sunset dari berbagai
perlakuan induksi pemijahan dan pemijahan alami A. tenuis ... 55 9. Hasil pengamatan setiap fase pemijahan karang A. tenuis
(15 November 2011) ... 58 10. Komparasi metode induksi pemijahan biota laut : kerang kima
(Tridacnidae), kerang lola (Trochidae) dan karang (Scleractinia) ... 61 11. Metode stimulasi pemijahan karang (Scleractinia) dari berbagai
sumber dan penelitian ini ... 63 12. Tingkat fertilisasi dari A. tenuis, A. millepora dan C. crassa ... 78 13. Analisis ragam tingkat fertilisasi karang spesies A. tenuis,
A. millepora, Ctenactis crassa. ... 78 14. Tingkat pembelahan sel embrio karang A. millepora ... 86 15. Tingkat pembelahan sel embrio karang A.tenuis ... 87 16. Tingkat pembelahan sel embrio spesies Ctenactis crassa ... 89 17. Klasifikasi umur berdasarkan fase hidup embrio-larva-juvenil
karang dan total competence time ... 90 18. Perbandingan umur larva dan competence time dari tiap spesies ... 92 19. Rata-rata dan rentang ukuran setiap fase embriogenesis
masing-masing spesies karang ... 95 20. Analisis sidik ragam tingkhlaku pengendapan larva planula A. tenuis. ... 115 21. Proporsi pengendapan larva [rata-rata ± (SE)] pada intensitas
cahaya dan selama waktu penelitian ... 117 22. Analisis sidik ragam pengaruh intensitas cahaya dan posisi substrat
23. Hasil analisis sidik ragam (ANOVA) perlakuan induksi CCA dan
Zooxanthella terhadap metamorfosa larva planula A. tenuis ... 123 24. Perbandingan laju metamorfosis larva planula A. tenuis antar perlakuan Z3,
Z6, Z9 ... 124 25. Perbandingan laju metamorfosis larva planula A. tenuis antar perlakuan
DAFTAR LAMPIRAN
Halaman 1. Kelompok karang berpolip kecil dan polip besar ... 153 2. Curah hujan terkini (2011) dan rata-rata setiap bulan di
Kepulauan Spermonde dan Orpheus island Great Barrier Reef. ... 156 3. Waktu yang dibutuhkan untuk fase pemijahan karang
A.tenuis dalam perlakuan induksi pemijahan. ... 156 3. Rancangan perlakuan induksi pemijahan karang di OIRS
Great Barrier Reef Australia (November 2011) ... 157 4. Hasil uji beda rata-rata (Tukey HSD) perlakuan induksi
pemijahan karang A.tenuis ... 159 5. Analisis sidik ragam respon waktu pemijahan fase setting
dan glinding karang A.tenuis ... 159 6. Hasil ujibeda nyata Tukey perlakuan induksi pemijahan ... 160 7. Tingkat fertilisasi A. tenuis dan A. millepora selama 6 jam
setelah pemijahan ... 161 8. Analisis varian tingkat fertilisasi pada setiap waktu antar spesies ... 161 9. Hasil uji beda nyata Tukey tingkat fertilisasi spesies karang
A. tenuis, A. millepora, Ctenactis crassa. ... 161 10. Diameter telur, embrio dan panjang larva karang rata-rata
setiap spesies. ... 162 11. Analisis varian ukuran telur, embrio dan larva karang antar spesies ... 163 12. Hasil Uji beda nyata Tukey perkembangan ukuran embrio,larva
planula antara spesies ... 163 13. Jumlah larva yang mengendap dan berenang bebas pengaruh
dari intensitas cahaya S1, S2, S3 dan S4 ... 164 14. Kombinasi percobaan induksi zooxanthella dan CCA dalam proses
metamorfosis juvenil karang ... 165 15. Hasil analisis varian metamorfosa juvenil karang yang diberi perlakuan
CCA/non CCA dan zoxanthella menurut waktu pengamatan ... 167 16. Hasil uji beda nyata Tukey metamorfosis harian juvenil karang ... 168 17. Hasil uji beda nyata Tukey HSD pengendapan tingkahlaku
larva setiap waktu pengamatan hari ke2, 4, dan 6 ... 168 18. Hasil analisis varian pengendapan larva karang berdasarkan hari,
intensitas cahaya dan kombinasi keduanya. ... 169 19. Hasil uji beda nyata Tukey metamorfosa larva planula yang
diinduksi zooxanthella ... 169 20. Hasil uji beda nyata tukey terhadap metamorfosis juvenil
karang yang diinduksi konsentrasi zooxanthella ... 170 21. Hasil uji beda nyata Tukey terhadap metamorfosis juvenil
PENDAHULUAN
Latar Belakang
Terumbu karang di Indonesia mengalami kerusakan akibat faktor antropogenik dan peristiwa alami. Sekitar 30 % terumbu karang masih dalam kondisi baik dan sangat baik, sebaliknya 70 % telah rusak dan rusak parah (Puslit Oseanografi-LIPI 2009). Wilayah Indonesia timur, perikanan yang merusak (destructive fishing) merupakan penyebab utama rusaknya terumbu karang. Di samping itu, perdagangan karang hidup untuk hiasan akuarium di seluruh dunia terus meningkat 12-30 % per tahun. Sementara Indonesia merupakan negara pengekspor terbesar setelah Filipina mengurangi jumlah kuota ekspornya (Yusuf et al. 2007). Eksploitasi karang hias dengan mengandalkan populasi di alam berdampak pada hilangnya beberapa spesies karang tertentu. Kelangkaan populasi karang tertentu dapat menyebabkan berkurangnya potensi reproduksi bagi sebagian besar spesies target. Akibatnya dari sisi fisiologi reproduksi, komunikasi secara biokimiawi reproduksi menjadi jarang sehingga beberapa spesies karang mengalami kegagalan reproduksi.
Pengendalian pengrusakan dan eksploitasi terumbu karang disamping melalui pendekatan sosial, juga diperlukan pendekatan melalui sains dan tehnologi. Upaya restorasi terumbu karang bisa dilakukan melalui metode transplantasi dan metode biorock. Namun tehnologi tersebut masih mengandalkan bibit hasil fragmentasi dari alam. Bila pengambilan induk karang dengan cara-cara yang tidak benar, maka terumbu juga akan terancam.
Barrier Reef, Australia (Willis et al. 1985). Hingga saat ini penelitian mengenai reproduksi karang secara alami di terumbu karang tropik Indonesia dilakukan oleh Bahtiar (1994), Munasik (2002), Rani (2004), Romawanti (2006), Kawaroe et al. (2007), Mustafa (2010) dan Patiung (2010). Penelitian masih sebatas histologi gonad untuk menentukan kematangan gonad dan periode reproduksi.
Jika dibanding dengan terumbu karang Indonesia yang sangat luas dan biodiversitas yang sangat beragam, informasi reproduksi beberapa spesies karang secara spasial masih belum cukup untuk menggambarkan pola reproduksi karang di Indonesia. Menurut Twan et al. (2006) faktor lingkungan berperan penting dalam menentukan waktu reproduksi dan pemijahan. Fluktuasi lingkungan yang kecil di daerah tropik seperti di Indonesia menjadi penyebab tersebarnya waktu reproduksi karang sepanjang tahun dan diduga tidak memijah secara sinkron antar spesies (Oliver et al. 1988; Richmond dan Hunter 1990).
Dengan adanya informasi waktu puncak reproduksi, maka pemijahan buatan (induksi reproduksi) karang menjadi lebih mudah untuk dilakukan. Teknologi induksi reproduksi merupakan langkah inisiasi dalam menemukan metode yang terbaik untuk memijahkan karang secara simultan, agar diperoleh sinkronisasi kebutuhan masing-masing sel gamet. Walaupun belum banyak informasi dan pengalaman mengenai pemijahan buatan karang sebelumnya, Seikai National Fisheries Research Institute Jepang telah mulai mengembangkan metode induksi pemijahan menggunakan hidrogen peroksida (H2O2) sebagai zat perangsang pemijahan gonad (Omori dan Fujiwara 2004). Metode ini masih menyisakan masalah, bila terjadi kesalahan konsentrasi H2O2 saat eksperimen, akan membawa dampak negatif bagi individu karang.
Hampir semua fase reproduksi invertebarata di laut termasuk hewan karang mengalami kondisi yang tidak terkontrol. Shimomura et al. (2000) dan Omori dan Fujiwara (2004) telah membuka jalan untuk memulai memelihara juvenil dari hasil induksi reproduksinya di Okinawa Jepang. Guest et al (2010) mengungkapkan metode terstruktur dan higienis penanganan reproduksi karang secara exsitu mulai dari pemijahan, fertilisasi dan pemeliharaan larva karang. Proses embriogenesis karang telah diteliti oleh Harrison dan Wallace (1990), khusus genus Acropora diteliti oleh Hayashibara et al. (1997), Fungia scutaria
Rumusan Masalah
Jika dibanding dengan luas dan keanekaragaman terumbu karang di Indonesia, maka informasi reproduksi karang masih sangat minim dan spasial. Metode penelitian reproduksi karang selama ini terbatas menggunakan metode histologi. Metode histologi membutuhkan perlakuan yang rumit dan lama sehingga informasi dan metode tersebut belum cukup untuk menggambarkan pola reproduksi karang di Indonesia.
Semua jenis karang keras yang berada di lintang tinggi memiliki kemampuan reproduksi masal dengan model pemijahan yang sinkron dalam jumlah yang besar (mass synchronous spawning). Sebaliknya, pada daerah lintang rendah atau mendekat ke daerah ekuator, sinkronisasi pemijahan antar koloni atau antar spesies jarang terjadi dan pola reproduksi sepanjang tahun (Oliver et al. 1988 dan Kenyon 1995 diacu Baird et al. 2004). Penelitian ini akan menjawab apakah terjadi sinkronisasi pemijahan karang di daerah tropis seperti di Indonesia ?
Ketika lingkungan perairan di sekitar daerah lintang tinggi Central Great Barrier Reef menunjukkan fluktuasi yang cukup besar, hal dapat memicu sinkronisasi pemijahan secara masal (Harrison dan Wallace 1990). Namun dengan fluktuasi faktor lingkungan yang relatif kecil atau hampir sama (Richmond dan Hunter 1990; Hayashibara et al. 1993), kapan dan dalam kondisi lingkungan yang bagaimana saat terjadinya pemijahan karang di Indonesia ?
Kondisi alamiah, pemijahan karang berlangsung pada saat bulan purnama atau bulan baru. Ketika itu terjadi pasang tertinggi dan surut terendah bulanan, efeknya kecepatan arus air yang lebih cepat juga membawa nutrien dan suhu yang relatif lebih dingin dibanding dengan saat pasang perbani. Berdasarkan prinsip fenomena osenografi tersebut, maka pemijahan buatan terhadap biota lola (Trochus nilotichus) dengan memanfaatkan arus air yang deras dan aerasi kuat mampu memberikan rangsangan terhadap pelepasan gamet atau pemijahan. Berdasarkan hal tersebut, apakah arus air deras dan aerasi kuat mampu memberikan respon kepada karang untuk melakukan pemijahan lebih awal dari jadwal alamiahnya ?
perkembangan embrio di luar tubuh mengalami banyak hambatan dan berbagai riset masih menyisakan pertanyaan. Untuk mempelajari embriogenesis bisa diawali dengan ujicoba dalam kondisi terkontrol laboratorium. Hampir semua spesies karang Acroporidae berjenis kelamin hermaprodit, sedangkan Fungiidae umumnya gonokhorik. Perbedaan morfologi keduanya lebih jelas dimana Acropora umumnya bercabang sementara Fungiidae semuanya berbentuk piring terbalik. Perbedaan morfologi dan model pemijahan antara keduanya, adakah perbedaan fertilisasi, embriogenesis dan competence time larva planulanya ?.
Metamorfosis dan pengendapan larva merupakan fase yang krusial dalam siklus hidup karang, karena pada fase ini larva mengalami perubahan morfologi tubuh dan berupaya menemukan substrat sebagai habitat baru untuk penempelan juvenilnya. Setiap spesies memiliki relung tersendiri untuk hidup dan beradaptasi sehingga secara genetik larva akan tetap memilih habitat seperti habitat induknya. Karang induk dan juga larva atau juvenilnya memiliki preferensi habitat pada kedalaman tertentu terkait dengan intensitas cahaya. Sehingga pertanyaan yang muncul adakah pengaruh cahaya terhadap pengendapan larva planula dan penempelan juvenil ?
Tujuan Penelitian
Tujuan umum dari penelitian reproduksi karang adalah untuk mengetahui pola dan waktu reproduksi karang di Kepulauan Spermonde Indonesia dan di Orpheus Island Central Great Barrier Reef dan membandingkan embriogenesis famili Acroporidae dan Pocilloporidae.
Secara khusus penelitian ini bertujuan :
1. Membandingkan pola reproduksi, sinkronisasi pemijahan karang tropis Indonesia dan subtropis Great Barrier Reef.
2. Mengetahui respon waktu pemijahan dan tingkahlaku pemijahan
Acropora tenuis melalui metode induksi fisik.
3. Membandingkan tingkat fertilisasi, perkembangan embrio, competense time karang dalam proses embriogenesis antar spesies dari Famili Acroporidae dan Fungiidae.
4. Mengetahui tingkahlaku pengendapan, metamorfosis dan penempelan larva berdasarkan intensitas cahaya, posisi substrat, induksi zooxanthella dan Crustose Coralline Algae (CCA).
Manfaat Penelitian
Hasil penelitian ini bermanfaat untuk mengembangkan metode reproduksi secara seksual ex-situ dalam kondisi terkontrol. Hasil pengembangbiakan secara masal ini bisa mensuplai kebutuhan juvenil karang untuk restorasi terumbu karang dan perbanyakan populasi karang hias skala besar.
Ruang Lingkup Penelitian
menentukan masa reproduksi karang di alam. Selain itu, habitat yang sangat bervariasi, diperkirakan bisa mempengaruhi masa reproduksi yang berbeda bagi setiap spesies karang. Asumsi perbedaan waktu reproduksi karang menurut wilayah akan memunculkan pertanyaan mengenai faktor lingkungan yang paling menentukan reproproduksi karang pada setiap kawasan. Kebanyakan peneliti menjelaskan perbedaan faktor oseanografi sangat mempengaruhi reproduksi karang secara independen. Menurut Kenyon (2008) di wilayah perairan Indo Pasifik Tengah isolasi reproduksi karang umumnya terjadi secara temporal.
Hingga saat ini penelitian reproduksi karang di Indonesia masih terbatas pada metode gametogenesis. Sementara metode pengecekan gonad yang matang secara insitu dan pemijahan secara exsitu belum banyak diterapkan. Mengingat wilayah terumbu karang Indonesia yang begitu luas dan sebagai kawasan hot spot diversity coral triangle, maka metode histologi untuk informasi reproduksi belumlah cukup untuk menggambarkan pola dan musim reproduksi karang di Indonesia.
Suatu tantangan dunia sains biologi karang, masih banyak spesies yang belum diketahui musim dan pola reproduksinya, disamping itu sinkronisasi pemijahannya pun masih belum jelas. Karang memijah membutuhkan waktu yang sinkron pada jam-jam tertentu (Veron 2000; Fukami et al. 2004; Omori dan Fujiwara 2004). Karang akan memijah pada jendela waktu 1 jam pada malam hari (Ananthasubramaniam et al. 2011).
Walaupun belum banyak informasi dan pengalaman mengenai pemijahan buatan karang sebelumnya, Seikai National Fisheries Research Institute Jepang telah mulai mengembangkan metode induksi pemijahan menggunakan hidrogen peroksida (H2O2) sebagai zat perangsang pemijahan gonad (Omori dan Fujiwara 2004). Prinsip dari induksi fisik pemijahan adalah merangsang polip karang untuk memberikan respon biologis pelepasan gamet. Metode induksi fisik menjadi alternatif untuk pemijahan biota yang dapat memecahkan masalah di atas. Yusuf et al. (2006) telah berhasil memijahkan kerang lola (Trochus
niloticus) menggunakan metode fisik (air mengalir, aerasi kuat, dan desikasi atau
penjemuran). Upaya pemijahan buatan merupakan langkah inisiasi dalam memecahkan masalah sinkronisasi pemijahan karang dalam laboratorium.
pada kolom air (Levitan et al. 1992; Lasker 2006). Dalam siklus hidup hewan karang, fertilisasi merupakan fase yang sangat ringkas (Lasker 2006). Kegagalan fertilisasi membawa dampak yang fatal bagi populasi karang. Sementara embriogenesis merupakan proses perkembangan embrio hingga menjadi juvenil. Embriogenesis telah dijelaskan oleh Harrison dan Wallace (1990), khusus genus Acropora oleh Hayashibara et al. (1997), Fungia scutaria
oleh Schwarz et al. (1999), Selain itu Ball et al. (2002), Okubo dan Motokawa (2007), Gleason dan Hoffman (2010) mereview beberapa aspek dalam embriogenesis karang. Selama fase planktonik, embrio-larva membutuhkan waktu yang berbeda hingga melekat di dasar atau disebut competence ime (Gleason dan Hoffmann 2011). Secara umum, competence time bagi larva karang sekitar 48 jam (Thamrin 2006), namun ada pula yang 96 jam tergantung genetik dan faktor lingkungan.
Gambar 1 Bagan alir kerangka penelitian reproduksi seksual karang yang berawal dari degradasi terumbu karang hingga solusi sains dan tehnis.
Reproduksi seksual di alam
Riset observasi reproduksi di alam eksperimen metode pengembangbiakan
Reproduksi seksual karang (ordo Scleractinia) : pemijahan, perkembangan larva dan metamorfosa
Pengembangbiakan terkontrol
- Pola reproduksi tahunan (T1) dan Sinkronisasi pemijahan
- Indikator lingkungan pemicu pemijahan (T3)
- Metode pemijahan yang efektif (T3)
- Perkembangan embrio-larva, competence time (T4). - Laju metamorfosis (T5,T6, T7)
Pengembangan sains bio-ekologi dan fisiologi reproduksi
Metode pengembangbiakan dan perbanyakan populasi karang,
Informasi reproduksi yang minim Ujicoba reproduksi yang jarang
Populasi karang menurun /degradasi terumbu karang
Pengrusakan habitat t.karang
Kegagalan reproduksi Eksploitasi karang hias/diperdagangkan
Sinkronisasi reproduksi Rekrutmen alami
dan restorasi
Tehnik reproduksi terkontrol
Kegunaan Hasil penelitian
Kebaruan Penelitian
Kebaharuan (novelty) penelitian ini yakni :
1. Penelitian ini mengungkap pola waktu reproduksi karang berpolip kecil dan berpolip besar dan pemijahan yang sinkron (menurut hari dan jam) beberapa spesies dari genus Acropora di perairan tropis Indonesia. Belum ada penelitian sebelumnya yang mengungkap pola reproduksi tahunan khusus karang tropis dan juga penelitian yang mengungkap sinkronisasi pemijahan karang di daerah tropis, mengingat kondisi lingkungan perairan yang cenderung konstan. Penelitian reproduksi karang oleh Bahtiar (2002), Rani dan Jompa (2006), Munasik dan Wijatmoko (2004); Romawanti (2006); Patiung (2010) dan Mustafa (2010) di Indonesia atau tropis lainnya hanya mengungkap gametogenesis secara spasial, tidak secara temporal tanpa menemukan sinkronisasi pemijahan. Sementara beberapa hasil riset mengungkapkan bahwa Pemijahan yang sinkron antar spesies dan koloni hanya terjadi pada terumbu karang lintang tinggi seperti di GBR, Red Sea dan Jepang (Willis et al. 1985; Hanafy et al. 2010; Hayashibara et al. 1993).
Hipotesis penelitian
Hipotesis yang diajukan untuk dianalisis dan dibuktikan adalah :
1. Induksi pemijahan berupa air mengalir dan aerasi kuat mampu memberikan rangsangan pemijahan karang lebih awal
2. Terdapat pengaruh intensitas cahaya terhadap pengendapan dan pelekatan juvenil karang Acropora tenuis.
3. Posisi substrat berpengaruh terhadap penempelan juvenil karang Acropora tenuis.
METODOLOGI UMUM
Waktu dan Lokasi Penelitian
Penelitian ini telah dilaksanakan selama dua tahun (2010-2012) yang bertempat di Kepulauan Spermonde Selat Makassar dan di Orpheus Island Research Station (OIRS) Central Great Barrier Reef Australia. Pengamatan pemijahan dan pengecekan gonad karang secara insitu di Kepulauan Spermonde dilakukan di terumbu karang Pulau Bone Tambu, Pulau Langkai, Pulau Badi. Pengamatan gonad dan pemijahan karang di alam secara insitu dilakukan di Cattle Bay, Pioneer Bay Orpheus Island, sementara pengamatan pemijahan karang exsitu dilakukan di laboratorium Orpheus Island Research Station (OIRS).
Kawasan Kepulauan Spermonde merupakan paparan yang tersusun dominan oleh terumbu karang yang memiliki 98 buah pulau dengan luas paparan 60.000 ha (Jompa et al. 2006) baik yang bertipe terumbu karang tepi (fringing reef), maupun patch reef yang membentuk terumbu pengahalang (barrier reef). Pada sisi luar sebelah barat, terumbu karang berupa gosong karang (patch reef) memanjang membentuk terumbu penghalang (barrier reef). Paparan terumbu karang dan perairan Spermonde lebarnya sekitar 40 km dari daratan Makassar yang terbagi menjadi empat zona berdasarkan jarak dan pengaruh daratan, yakni : zona satu atau zona pinggir dari pantai kearah laut lepas sejauh kira-kira 5 km atau hingga kedalaman 20 m. Pada zona dua atau zona dalam , mulai dari jarak 5 km kearah laut hingga 12,5 km, sedangkan zona tiga atau zona tengah dari jarak 12,5 km ke arah laut lepas hingga 30 km dengan kedalaman yang bervariasi antara 30 – 50 m. Sementara zona empat atau zona terluar atau barrier reef zone mulai dari jarak 30 km hingga 40 km batas terluar dari paparan Spermonde (Hoeksema 1990).
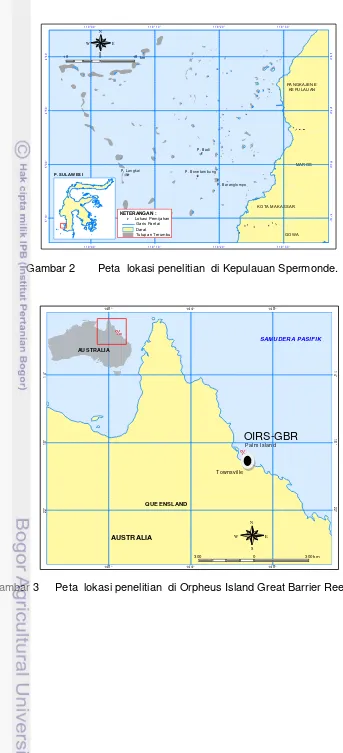
Gambar 2 Peta lokasi penelitian di Kepulauan Spermonde.
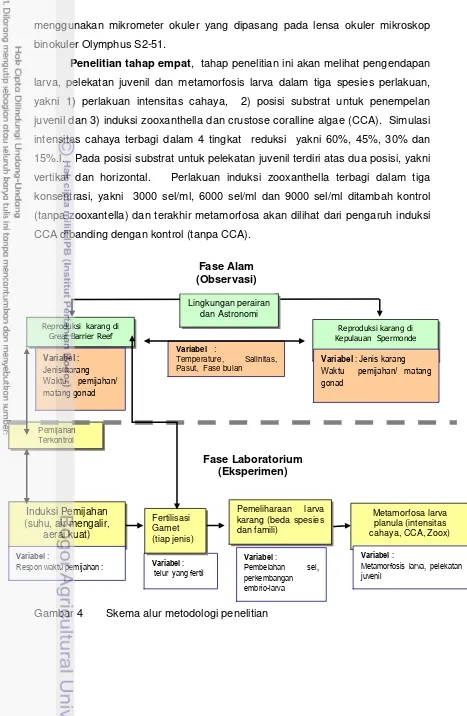
Gambar 3 Peta lokasi penelitian di Orpheus Island Great Barrier Reef. # Y # Y # Y # Y MAR OS
GO W A PA NG KA JE NE
KE PU LAU AN
KO T A M AK AS SA R P . B ad i
P . B ar a n gl o m po P . B on e ta m b u ng P . La n g ka i
5 ° 1 0 ' 5 ° 0 0 ' 4 ° 5 0 ' 4 ° 4 0 ' 5 ° 1 0 ' 5 ° 0 0 ' 4 ° 5 0 ' 4 ° 4 0 '
1 1 9°0 0 ' 1 1 9°1 0 ' 1 1 9°2 0 ' 1 1 9°3 0 ' 1 1 9°0 0 ' 1 1 9°1 0 ' 1 1 9°2 0 ' 1 1 9°3 0 '
N
E W
S
10 0 10 km
KETERANGAN :
Darat Tu tupa n T erumb u Ga ris Pan tai
#
Y Lok asi P emijahan P. S UL AW ES I
%
AUSTRALIA
QUE ENSL AND
Town s ville Pa lm Is lan d
SAM UDERA PAS IFIK
2 2 ° 1 8 ° 1 4 ° 2 2 ° 1 8 ° 1 4 °
14 0° 14 4° 14 8°
14 0° 14 4° 14 8°
%
AU S TR ALIA
N E W
S
300 0 300 km
Metode Penelitian
Secara umum, penelitian ini terbagi dalam empat bagian, yakni pola reproduksi, induksi pemijahan, perkembangan larva, dan metamorfosis. Penelitian tahap pertama, dengan melakukan pengecekan kematangan gonad dan pengamatan pemijahan secara insitu di alam. Disamping itu dilakukan pemijahan exsitu di laboratorium. Kemunculan gonad pada patahan karang bercabang menunjukkan kematangan gonad. Pengamatan kematangan gonad dan pemijahan karang dilakukan pada bulan terang (purnama) dan bulan gelap (perbani). Pencatatan waktu pemijahan dilakukan untuk menunjukkan sinkronisasi pemijahan karang. Untuk mendukung penelitian ini digunakan pula empat penelitian lain terhadap beberapa spesies karang Euphyllia glabrescen, Euphyllia ancora, Galacea fascicularis dan Acropora nobilis yang umumnya menggunakan metode histologi gonad.
Penelitian tahap dua, dalam wadah running water system percobaan induksi pemijahan menggunakan air mengalir, aerasi kuat dan kombinasi keduanya serta kontrol untuk membuktikan pengaruhnya terhadap waktu pemijahan. Pemberian rangsangan dalam induksi pemijahan karang berlangsung selama 3 jam (15: 00 – 18:00). Rangsangan fisik ini dihentikan untuk memberikan peluang hewan uji (Acropora tenuis) berelaksasi. Selama kurun waktu 30 menit dilakukan pengamatan respon tingkahlaku polip akibat perlakuan tersebut hingga saat pemijahan (pelepasan gonad) berlangsung. Rangkaian proses pemijahan gonad terbagi dalam 5 tahap, yakni setting stage atau kemunculan gonad sekitar mulut, birth stage atau proses pengeluaran gonad terjepit di mulut, gliding stage atau pelepasan buntelan dari mulut tapi tertahan oleh tentakel , upward stage atau pelepasan ke kolom air, dan bursting stage atau pemecahan buntelan telur.
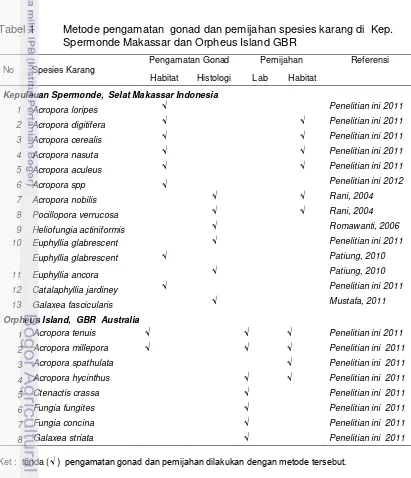
menggunakan mikrometer okuler yang dipasang pada lensa okuler mikroskop binokuler Olymphus S2-51.
[image:40.595.51.518.66.782.2]Penelitian tahap empat, tahap penelitian ini akan melihat pengendapan larva, pelekatan juvenil dan metamorfosis larva dalam tiga spesies perlakuan, yakni 1) perlakuan intensitas cahaya, 2) posisi substrat untuk penempelan juvenil dan 3) induksi zooxanthella dan crustose coralline algae (CCA). Simulasi intensitas cahaya terbagi dalam 4 tingkat reduksi yakni 60%, 45%, 30% dan 15%.l. Pada posisi substrat untuk pelekatan juvenil terdiri atas dua posisi, yakni vertikal dan horizontal. Perlakuan induksi zooxanthella terbagi dalam tiga konsentrasi, yakni 3000 sel/ml, 6000 sel/ml dan 9000 sel/ml ditambah kontrol (tanpa zooxantella) dan terakhir metamorfosa akan dilihat dari pengaruh induksi CCA dibanding dengan kontrol (tanpa CCA).
Gambar 4 Skema alur metodologi penelitian
Variabel : telur yang fertil
Metamorfosa larva planula (intensitas cahaya, CCA, Zoox)
Induksi Pemijahan (suhu, air mengalir,
aerai kuat)
Pemeliharaan larva karang (beda spesies dan famili)
Pemijahan Terkontrol
Variabel :
Pembelahan sel, perkembangan embrio-larva Fertilisasi Gamet (tiap jenis) Variabel :
Respon waktu pemijahan :
Variabel :
Metamorfosis larva, pelekatan juvenil
Fase Laboratorium (Eksperimen)
Reproduksi karang di Kepulauan Spermonde Reproduksi karang di
Great Barrier Reef
Variabel :
Temperature, Salinitas, Pasut, Fase bulan
Variabel: Jenis karang
Waktu pemijahan/ matang gonad
Variabel :
Jenis karang Waktu pemijahan/ matang gonad
Lingkungan perairan dan Astronomi
POLA REPRODUKSI DAN SINKRONISASI PEMIJAHAN KARANG
(Ordo
Scleractinia)
DI KEPULAUAN SPERMONDE INDONESIA DAN
ORPHEUS ISLAND GREAT BARRIER REEF AUSTRALIA
ABSTRAK
SYAFYUDIN YUSUF. Pola Reproduksi dan Sinkronisasi Pemijahan Karang (Ordo Scleractinia) Di Kepulauan Spermonde Indonesia dan Orpheus Island Great Barrier Reef Australia. Dibimbing oleh NEVIATY P. ZAMANI, M. ZAIRIN JUNIOR dan JAMALUDDIN JOMPA
Pola reproduksi seksual dan waktu pemijahan karang tergantung pada spesies, geografi, musim dan kondisi perairan baik lokal maupun regional. Penelitian ini bertujuan menentukan pola reproduksi sinkronisasi dan lingkungan yang berpengaruh terhadap pemijahan karang di Kepulauan Spermonde dibandingkan dengan di Great Barrier Reef (BGR) tengah. Pola reproduksi seksual bagi karang berpolip kecil umumnya terjadi pada musim hujan, akan tetapi karang berpolip besar matang gonad umumnya pada musim kemarau di kepulauan Spermonde dibanding dengan di GBR terjadi pada bulan November dan Desember pada awal musim panas. Pemijahan yang sinkron terjadi pada jam 18:00-19:00 pada 0-2 hari bulan purnama (BP) pada saat pasang tinggi, temperatur 30,3oC di kepulauan Spermonde. Sedangkan pemijahan yang multispesies di GBR berlangsung pada jam 19 : 00 – 21:00 selama 3-5 (BP) pada saat air laut surut dimana suhu 27oC. Perairan Kepulauan Spermonde memiliki variasi lingkungan yang kecil, sehingga waktu reproduksi karang menyebar sepanjang tahun mengikuti pola dua musim (hujan dan kemarau) dan periode bulan (purnama dan akhir atau awal), dibandingkan dengan pemijahan karang di Great Barrier Reef dimana terjadi pemijahan masal yang sinkron. Kondisi lingkungan pemijahan karang berpolip besar berbeda dengan lingkungan karang berpolip kecil.
ABSTRACT
SYAFYUDIN YUSUF. Reproduction Pattern and Synchronous Spawning of Coral (Ordo: Scleractinia) in Spermonde Archipelago and Great Barrier Reef Australia. Supervised by NEVIATY P. ZAMANI, M.ZAIRIN JUNIOR and JAMALUDDIN JOMPA
Pattern of sexual reproduction and spawning event of scleractinian corals are correlated with the species, geography, monsoon, local and regional oceanographic conditions. This study was aimed to determine the reproduction pattern, synchronization and the environmental cues of spawning in Spermonde Archipelago reefs compared to the Central Great Barrier Reef. Reproduction pattern of small polyp corals was commonly performed during rainy season but the bigger polyps were in dry seasons in Spermonde compared to the GBR that occurred during November and December of early summer. Synchronously broadcast spawning took place at 18:00–19:00 in lunar period 0–2 days after full moon, while the high tide level and the temperature was 30.3 oC in Spermonde. Otherwise, multispecies coral mass spawning in GBR took place at 19:00-21:.00 for 3-5 days AFM in the low tides while the temperature was 27 oC. Spermonde Archipelago waters have less environmental variability, so that coral spawning event extend for several months and occured during different lunar phases compared to the Great Barrier Reef that have synchronous mass spawning. Environmental condition for spawning event of the small polyps coral were different from the bigger polyps corals.
PENDAHULUAN
Karang keras (Filum Coelenterata; Kelas Anthozoa; Ordo Scleractinia) memiliki kemampuan reproduksi ganda yakni secara seksual dan aseksual (Sumich 1996, Richmond dan Hunter 1990, Thamrin 2006). Reproduksi seksual secara alami semua hewan karang bervariasi tergantung pada spesiesnya. Koloni karang mampu menghasilkan gamet jantan dan betina. Sekitar 75 persen dari semua spesies karang merupakan koloni yang hermaprodit. Sifat seksualitas dari karang baik yang hermaprodit atau yang gonokoristik tergantung pada spesies dan atau genera (Wallace 1999, Veron 2000).
Penelitian reproduksi karang dimulai sejak tahun 1937, sepuluh tahun kemudian Atoda 1947 meneliti perkembangan larva karang (Kojis dan Quinn, 1981), sebelumnya Kojis dan Quinn (1980) juga telah meneliti reproduksi karang Faviidae. Tahun 1980-an penelitian reproduksi karang berkembang di Great Barrier Reef Australia, Laut Merah dan Karibia (Willis et al. 1985, Richmond dan Hunter 1990, Wallace 1999). Perkembangan penelitian reproduksi karang sudah menemukan fenomena pemijahan yang diawali tahun 1980-an (Poinski 2004). Sebagian besar spesies karang keras (ordo Scleractinia) tergolong pemijah (spawner), yakni melepaskan gamet ke kolom air secara simultan (Richmond dan Hunter 1990, Baird et al. 2009, Willis et al. 1985).
Menurut Mendes dan Woodley (2002) bahwa reproduksi seksual karang ditentukan antara lain oleh spesies karang dan letak lintang, isolasi geografi, musim, kondisi oseanografi lokal maupun regional dimana karang hidup. Seperti terumbu karang di Indonesia yang terletak sekitar garis khatulistiwa kemungkinan memiliki pola reproduksi yang berbeda dengan daerah tropis lainnya dan juga subtropis. Iklim Indonesia yang terdiri dari musim hujan dan musim kemarau akan menentukan masa reproduksi karang di alam.
independen. Menurut Kenyon (2008) di wilayah perairan Indo Pasifik Tengah isolasi reproduksi karang umumnya terjadi secara temporal.
Willis et al. (1985), Harrison dan Wallace (1990) banyak menjelaskan mengenai reproduksi masal secara sinkron merupakan fenomena yang unik di terumbu karang Australia. Sekitar terumbu karang Great Barrier Reef lebih lebih dari 143 spesies memijah dalam minggu yang sama pada bulan purnama (Willis et al. 1985, Richmond dan Hunter 1990, Willis et al. 1997, Willis et al. 2006). Selain di Great Barrier Reef Australia fenomena pemijahan masal karang banyak tercatat di daerah subtropis seperti Jepang (Loya et al. 2009 ).dan Karibia (Willis et al. 1997, Secore 2009). Penelitian observasi reproduksi karang tersebut memberikan rujukan kepada para peneliti lain untuk melihat tipe dan pola reproduksi karang di wilayah terumbu karang lain seperti di kawasan hot spot diversity coral triangle, Indonesia. Hingga saat ini penelitian reproduksi karang masih terbatas pada gametogenesis seperti oleh Bahtiar (1994) di Selat Lombok, Munasik (2002) di Jepara, Rani (2004) dan Romawanti (2006) di Kep. Spermonde.
Penelitian reproduksi karang baik pola reproduksi, waktu kematangan gonad, maupun waktu pemijahan akan sangat bermanfaat sebagai langkah awal dalam mengembangkan reproduksi seksual karang di Indonesia. Penelitian ini bertujuan untuk membandingkan pola reproduksi dan waktu puncak pemijahan gamet beberapa spesies karang serta faktor lingkungan yang terdapat di Kepulauan Spermonde Indonesia dan di Orpheus Island Great Barrier Reef Australia.
METODOLOGI PENELITIAN
Waktu dan tempat
pemijahan karang exsitu dilakukan di laboratorium Orpheus Island Research Station (OIRS).
Kawasan Kepulauan Spermonde merupakan paparan yang tersusun dominan oleh terumbu karang yang memiliki 98 buah pulau dengan luas paparan 60.000 ha (Jompa et al. 2006) baik yang bertipe terumbu karang tepi fringing reef maupun patch reef yang membentuk barrier reef. Pada sisi luar sebelah barat, terumbu karang berupa gosong karang (patch reef) memanjang membentuk terumbu penghalang (barrier reef). Paparan terumbu karang dan perairan Spermonde lebarnya sekitar 40 km dari daratan Makassar yang terbagi menjadi empat zona berdasarkan jarak dan pengaruh daratan, yakni : zona satu atau zona pinggir dari pantai kearah laut lepas sejauh kira-kira 5 km atau hingga kedalaman 20 m. Pada zona dua atau zona alam , mulai dari jarak 5 km kearah laut hingga 12,5 km, sedangkan zona tiga atau “zona tengah” dari jarak 12,5 km ke arah laut lepas hingga 30 km dengan kedalaman yang bervariasi antara 30 – 50 m. Pada zona empat atau zona terluar atau barrier reef zone mulai dari jarak 30 km hingga 40 km batas terluar dari paparan Spermonde (Hoeksema 1990).
Orpheus Island adalah sebuah pulau dalam kawasan Palm Islands Marine Park yang terdapat dalam kawasan Great Barrier Reef. Pulau ini terletak memanjang utara-selatan dengan beberapa teluk yang menjadi kawasan konservasi dan riset, seperti Cattle Bay dan Pioneeer Bay dimana semuanya menghadap ke daratan utama (leeward). Lokasi ini masih mendapat pengaruh sedimentasi sehingga tingkat kekeruhan kolom air lebih tinggi dan jarak pandang yang pendek
Bahan dan Metode
Pengamatan insitu Kematangan Gonad Karang di Alam
[image:46.595.87.498.267.745.2]Pengamatan kematangan gonad karang dilakukan secara exsitu dan insitu (Willis et al. 1985) terhadap 13 spesies karang dari Kepulauan Spermonde dan 8 spesies di Orpheus Island. Daftar spesies karang dan metode pengamatannya tertera pada Tabel 1 di bawah. Secara exsitu, pengamatan gonad dil akukan dengan cara histologi dan pengamatan pemijahan terkontrol di laboratorium. Secara insitu kematangan gonad karang bisa terlihat dari warna sel gonad dari bekas patahan karang dengan cara mematahkan sebagian koloni karang Acropora (Harrison et al. 1984) langsung pada habitatnya.
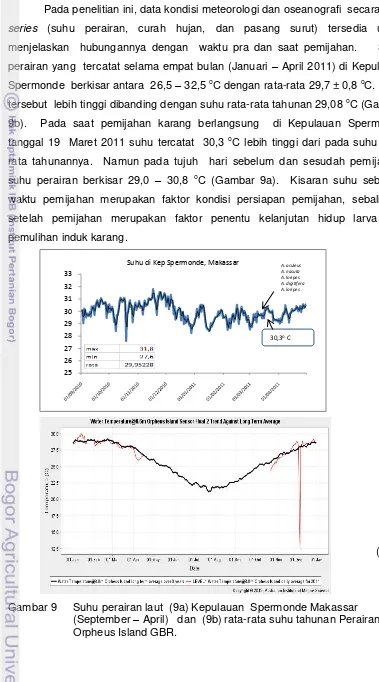
Tabel 1 Metode pengamatan gonad dan pemijahan spesies karang di Kep. Spermonde Makassar dan Orpheus Island GBR
No Spesies Karang
Pengamatan Gonad Pemijahan Referensi Habitat Histologi Lab Habitat
Kepulauan Spermonde, Selat Makassar Indonesia
1 Acropora loripes √ Penelitian ini 2011
2 Acropora digitifera √ √ Penelitian ini 2011
3 Acropora cerealis √ √ Penelitian ini 2011
4 Acropora nasuta √ √ Penelitian ini 2011
5 Acropora aculeus √ √ Penelitian ini 2011
6 Acropora spp √ Penelitian ini 2012
7 Acropora nobilis √ √ Rani, 2004
8 Pocillopora verrucosa √ √ Rani, 2004
9 Heliofungia actiniformis √ Romawanti, 2006
10 Euphyllia glabrescent √ Penelitian ini 2011
Euphyllia glabrescent √ Patiung, 2010
11 Euphyllia ancora √ Patiung, 2010
12 Catalaphyllia jardiney √ Penelitian ini 2011
13 Galaxea fascicularis √ Mustafa, 2011
Orpheus Island, GBR Australia
1 Acropora tenuis √ √ √ Penelitian ini 2011
2 Acropora millepora √ √ √ Penelitian ini 2011
3 Acropora spathulata √ Penelitian ini 2011
4 Acropora hycinthus √ √ Penelitian ini 2011
5 Ctenactis crassa √ Penelitian ini 2011
6 Fungia fungites √ Penelitian ini 2011
7 Fungia concina √ Penelitian ini 2011
8 Galaxea striata √ Penelitian ini 2011
Khusus karang yang berpolip besar Catalaphyllia jardiney, E. glabrescen dan E.ancora polipnya dibuka menggunakan betel dan palu untuk memecahkan kerangka kapurnya yang keras. Indikasi kematangan gonad dari hasil patahan cabang karang yakni bila terlihat buliran kuning atau oranye apabila specimen karang tersebut sudah matang gonad atau siap memijah (Harrison et al. 1984). Berikut pada Gambar 5 ditampilkan metode pengamatan reproduksi karang secara insitu dan exsitu.
(a) (b)
[image:47.595.105.493.147.759.2](c) (d)
Gambar 5 Pengecekan kematangan gonad secara insitu (a,b), pemijahan secara exsitu (c,d).
Pengamatan Pemijahan Karang insitu dan exsitu
Secara exsitu pengamatan pemijahan karang dilakukan dalam laboratorium Orpheus Island Research Station (OIRS) GBR (Gambar 1) dan Marine Station Pulau Barrang Lompo Makassar. Beberapa koloni karang target yang telah diketahui matang gonad diangkat dari habitatnya kemudian disimpan dalam bak-bak penampungan. Bak-bak tersebut berisi air tersaring 10-50 mµ dan terus diberi aerasi. Setiap koloni karang diberi alas agar mendapat sirkulasi air yang sama pada semua bagian koloni. Pengamatan mulai dilakukan sesaat setelah matahari terbenam, suasana lingkungan dalam kondisi gelap. Setiap koloni diamati kondisi pemijahannya menggunakan senter cahaya infra red.
Variabel Pemijahan Karang
Variabel pemijahan karang dalam penelitian ini terbagi dua, yakni variabel tergantung (dependent / internal karang) dan variabel bebas (independen / lingkungan). Variabel dependent terdiri dari tipe pemijahan : apakah karang brooder atau spawner dan jenis kelamin spesies : apakah hermaprodit atau gonokhorik (Richmond dan Hunter 1990; Wallace 1999, Thamrin 2006) . Variabel waktu pemijahan yang perlu dicatat yakni bulan, tanggal, dan jam pemijahan (setelah tenggelam matahari) (Willis et al. 1985). Tipe karang brooder, yakni kelompok karang yang melakukan pembuahan (fertilisasi) secara internal dalam rongga tubuh sehingga bakal individu keluar dalam bentuk planula. Tipe karang spawner, yakni kelompok karang yang melepaskan sel telur dan sel sperma ke kolom air sehingga melakukan fertilisasi di luar tubuh. Sementara karang yang hermaprodit dimana ovari dan sperma berkembang dalam mesenteri yang sama, atau dalam polip yang sama, atau berbeda polip tapi dalam koloni yang sama, melepaskan gamet baik secara bersamaan ataupun berbeda waktu. Sebaliknya, karang yang gonokhorik hanya memiliki satu spesies gamet (sperma atau ovari) dan hanya melepaskan salah satunya (Richmond dan Hunter 1990, Guest et al. 2010).
Variabel Lingkungan Reproduksi Karang
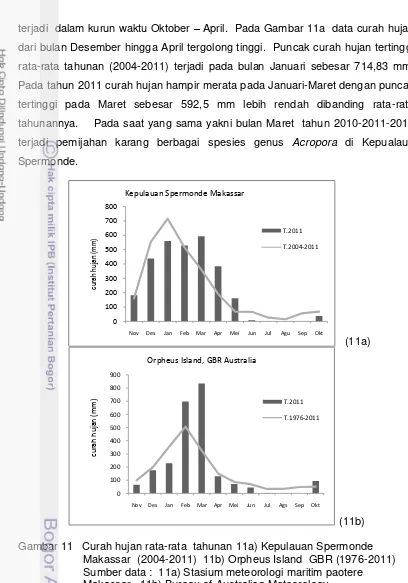
et al. 2009 ), dan curah hujan (Mendes dan Woodley 2002). Dalam penelitian ini variabel lingkungan (variabel independen) diperlukan adalah suhu perairan, pasang surut, fase bulan, dan curah hujan.
Pengukuran suhu perairan laut dilakukan secara manual menggunakan digital temperature di Pulau Badi setiap 3 kali sehari (pagi 06:00-07:00, siang 12:00-13:00 dan sore 17:00-18:00) dari tanggal 1 September 2009 hingga 30 April 2011. Data suhu perairan Orpheus Island tahun 2011 diperoleh dari AIMS data stasiun pengamatan Sensor Float 2 yang terpsang di Orpheus Island pada tahun 2012 (Australian Institute of Marine Science, 2012). Selanjutnya data curah hujan (2004-2011) di Kepulauan Spermonde diperoleh dari Stasiun Meteorologi Maritim Paotere Makassar, dan untuk data curah hujan Orpheus Island diperoleh dari Station Palm Island Water Treat Lat: 18.71° S Lon: 146.59° E (Bureau Meteorology of Queensland Australia 2012).
Analisis Data
Analisis multivariat Euclidian Distance (Krebbs 1999; Supranto 2010) terhadap variabel lingkungan yang berpengaruh terhadap pemijahan digunakan untuk mengklasifikasikan kelompok karang (polip besar dan polip kecil) berdasarkan kebutuhan lingkungan pemijahannya.
HASIL PENELITIAN
Spesies karang yang matang gonad dan memijah
Dari bebeberapa penelitian reproduksi karang tercatat 13 spesies karang yang diketahui waktu kematangan gonad dan puncak reproduksinya di Kepulauan Spermonde dan 8 spesies dari sekitar 30 spesies karang yang memijah secara sinkron sekitar Orpheus Island Great Barrier Reef (Tabel 2).
[image:50.595.71.493.31.805.2]Semua spesies karang pada Tabel 2 di bawah, dibagi menjadi dua kelompok yakni a) kelompok spesies polip kecil dan b) kelompok spesies polip besar. Karang berpolip kecil adalah spesies karang yang memiliki ukuran polip ≤ 5 mm, sedangkan karang berpolip besar ukuran polipnya > 5 mm hingga puluhan sentimeter (Veron, 2000).
Tabel 2 Daftar spesies karang (tingkat klasifikasi) yang matang gonad dan yang memijah di Kepulauan Spermonde Makassar .
Filum Coelenterata
Kelas Anthozoa
Ordo Scleractinia
Suku Acroporidae Genus Spesies Lokasi
Acropora
A.loripes* Spermonde
A.digitifera* Spermonde
A.cerelais* Spermonde
A.nasuta* Spermonde
A.aculeus* Spermonde
A.nobilis* Spermonde
Acropora spp Spermonde
A. tenuis* Orpheus Is
A. millepora* Orpheus Is
A. spathullata Orpheus Is
A. cytherea* Orpheus Is Pocilloporidae Pocillopora P. verrucosa* Spermonde Oculinidae Galaxea G.fascicularis** Spermonde
Euphyllidae Euphyllia E.ancora** Spermonde
E.glabrescen** Spermonde
Fungiidae Fungia F.fungites** Orpheus Is
F.concina** Orpheus Is
Ctenactis crassa** Orpheus Is
Yang termasuk karang polip kecil dalam Tabel 2 di atas yakni dari genera Acropora : A.loripes, A. digitifera, A. cerealis, A.nasuta, A.aculeus, Acropora nobilis dan Pocillopora verrucosa. Sedangkan yang termasuk karang berpolip besar yakni : Euphyllia glabrescen, Heliofungia actiniformis, Catalaphyllia jardiney, Galaxea fascicularis, Euphyllia ancora dan Euphyllia glabrescen. Spesies-spesies karang dari hasil penelitian lain di Kepulauan Spermonde : A. nobilis dan Pocillopora verrucosa (Rani 2004), Euphyllia ancora dan Euphyllia glabrescen (Patiung 2011), Heliofungia actiniformis (Romawanti 2006), dan Galaxea fascicularis (Mustafa 2011).
Pola Reproduksi Karang menurut Bulan dan Musim
Perbedaan letak geografis antara wilayah kepulauan Spermonde dan GBR Australia memberikan gambaran perbedaan waktu kematangan gonad (gametogenesis) dan waktu puncak pemijahan karang (spawing). Pada daerah dengan kondisi lingkungan yang cenderung konstan seperti daerah tropis Indonesia, periode pemijahan melebar sampai beberapa bulan dan pelepasan gamet terjadi fase bulan yang berbeda (Shlesinger dan Loya 1985, Szmant 1986, Richmond dan Hunter 1990, Hayashibara et al. 1983). Karakteristik perairan sekitar Orpheus Island Great Barrier Reef yang mengenal empat musim memiliki informasi reproduksi yang dan waktu puncak pemijahan berbagai spesies karang. Berbeda dengan perairan tropis seperti di Indonesia, misalnya di Kepulauan Spermonde, dimana perbedaan suhu perairan tidak terlalu menyolok menyebabkan waktu reproduksi karang menyebar sepanjang tahun.
Pada penelitian ini tercatat beberapa spesies karang yang matang gonad dan memijah di Kepulauan Spermonde Selat Makassar. Jika dilihat dari periode musim, maka kematangan gonad dan atau pemijahan gamet karang di Kepulauan Spermonde mengikuti pola musim, yakni ada yang memijah pada musim hujan dan ada pula pada musim kemarau. Secara normal, musim hujan berlangsung dari bulan Oktober – April, sementara musim kemarau berlangsung dalam bulan April –Oktober (Nontji 2007).
semuanya dari satu genera Acropora yakni A. loripes, A. digitifera, A cerealis, A. nasuta, dan A. aculeus.
Delapan spesies karang yang diteliti di Orpheus Island GBR semuanya memijah pada bulan November 2011. Setiap tahun pada lokasi yang sama dan spesies yang sama di Orpheus Island GBR berlaku ketepatan waktu pemijahan (synchronous spawning) yang sudah diprediksi sebelumnya. Namun demikian, hasil penelitian lain pada tahun 1980-an dan 1990-an memperlihatkan pemijahan karang di beberapa lokasi dan spesies yang sama di GBR sedikit bergeser dari Bulan November, misalnya pada Bulan Oktober dan Desember. Acropora tenuis yang memijah bulan November 2011 lalu, pernah tercatat memijah pada bulan Oktober di Magnetic Island dan bulan Desember di lokasi lain di GBR (Wallace 1999; Willis et al. 1985). Demikian halnya dengan Acropora millepora cukup banyak ditemukan memijah pada bulan November, namun pernah tercatat memijah pada bulan Desember (Wallace 1999; Richmond dan Hunter 1990).
Tabel 3 Pola musim dan bulan) kematangan gonad dan pemijahan karang di Kep.Spermonde dan Orpheus Island GBR.
No Spesies Bulan
Pemijahan
Musim Lokasi
SPE 1 Euphyllia ancora* E. glabrescen*
Desember Awal Hujan Spermonde
SPE 2 Catalaphyllia jardiney* Acropora .nobilis Pocillopora verrucosa
Februari Hujan Spermonde
SPE 3 A. loripes, A. digitifera A. cerealis, A. nasuta A. aculeus, Acropora spp
Maret Akhir Hujan Spermonde
SPE 4 Heliofungia actiniformis* E.glabrescen*
Agustus – September
Kemarau Spermonde
GBR 1 Acropora tenuis, A. millepora, A.hyacinthus, A. spathullata Galaxea sp* Fungia fungites* F. concina* Ctenactis crassa*
November Akhir Semi-Awal Panas
Great Barrier Reef
GBR 2 A. nasuta, A. listeri A.muricata, A. illepora, A.longicyanthus
Desember Awal Musim Panas
Great Barrier Reef
Keterangan : *karang berpolip besar
Data pemijahan bulan Desember diacu dari Wallace (1999)
kemarau seperti Heliofungia actiniformis dan Euphyllia glabrescen. Khusus spesies E.glabrescen tercatat matang gonad dua musim, yakni musim kemarau dan musim hujan tepatnya pada Bulan Agustus dan Desember. Karang polip besar lainnya adalah Catalaphyllia jardiney hanya ditemukan matang gonad pada akhir musim hujan tepatnya pada bulan Februari.
Sebaliknya, beberapa spesies karang di Orpheus Island GBR memijah p