ANALISIS GENETIK DAN EFISIENSI SELEKSI
MENGGUNAKAN
SINGLE SEED DESCENT
PADA KEDELAI
[
Glycine max
(L.) Merr.] UNTUK ADAPTASI TANAH MASAM
ARVITA NETTI SIHALOHO
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa disertasi „Analisis Genetik dan Efisiensi Seleksi Menggunakan Single Seed Descent pada Kedelai [Glycine max
(L.) Merril] untuk Adaptasi Tanah Masam‟ adalah karya saya sendiri dengan arahan Komisi Pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau kutipan dari karya yang diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor,Maret 2015
RINGKASAN
ARVITA NETTI SIHALOHO. Analisis Genetik dan Efisiensi Seleksi Menggunakan Single Seed Descent pada Kedelai [Glycine max (L.) Merr] untuk Adaptasi Tanah Masam. Dibimbing oleh TRIKOESOEMANINGTYAS, DIDY SOPANDIE dan DESTA WIRNAS.
Persilangan merupakan salah satu metode untuk menggabungkan sifat-sifat yang diinginkan. Hasil persilangan akan mempermudah usaha dalam menyeleksi tanaman untuk mendapatkan sifat yang diinginkan.Diharapkan dari penelitian ini dapat diseleksi galur-galur yang toleran tanah masam dan berdaya hasil tinggi di tanah masam dari persilangan Argomulyo denganTanggamus.
Penelitian ini bertujuan untuk (1) memperoleh informasi tentang aksi gen dan pewarisan sifat karakter agronomi kedelai terhadap cekaman aluminium di tanah masam dan di kultur hara; (2) memperoleh marka RAPD terpaut toleransi cekaman aluminium melalui metode bulk segregrant analisis; (3) memperoleh informasi tentang keragaman genetik karakter pertumbuhan, komponen hasil dan hasil pada F3 dan F4 dengan metode single seed descent; (4) memperoleh galur kedelai yang toleran tanah masam melalui seleksi daya hasil dan marka RAPD; (5) memperoleh informasi tentang keragaan pertumbuhan dan daya hasil galur-galur F5 terseleksi berdasarkan daya hasil dan marka RAPD terpaut toleransi cekaman Al.
Studi pewarisan sifat toleransi kedelai terhadap tanah masam dilaksanakan di dua lokasi, yaitu lahan di Jasinga, Kabupaten Bogor mulai bulan Mei sampai Agustus 2012 dan di Rumah Kaca University Farm IPB Cikabayan Bogor bulan Maret 2013. Studi keragaman genetik generasi F3 dan F4 dengan metode single seed
descent dilaksanakan di Kebun Percobaan BB Biogen, Cimanggu mulai bulan
Oktober 2012 sampai Mei 2013. Seleksi marka RAPD terpaut sifat toleransi aluminium menggunakan metode bulk segregant analysis dilaksanakan di Laboratorium Biomolekuler, Departemen Agronomi dan Hortikultura IPB mulai bulan April 2013 sampai Juni 2013. Perbandingan keragaan galur-galur F5 hasil seleksi karakter agronomi dan marka molekuler di tanah masam dilaksanakan di UPTD lahan kering Tenjo, Kabupaten Bogor pada bulan Oktober 2013 sampai Januari 2014. Rancangan yang digunakan yaitu Augmented Design.
komplementer tetapi bobot kering akar dikendalikan oleh sedikit gen dengan aksi gen aditif dan epistasis komplementer.
Studi keragaman karakter agronomi pada populasi kedelai yang dikembangkan dengan metode single seed descent memperlihatkan bahwa populasi F4 memiliki nilai tengah dan ragam genetik tetap tinggi untuk karakter agronomi yang diamati sama dengan populasi F3. Populasi F3 memiliki nilai heritabilitas dan KKG tergolong tinggi untuk karakter-karakter agronomi kedelai sama dengan populasi F4, kecuali karakter bobot 100 biji tergolong sedang.
Seleksi marka molekuler terpaut toleransi aluminium dengan metode bulk
segregant analysis (BSA) menghasilkan satu marka OPH-12 pada lokus 1200
yang terpaut sifat toleransi aluminium pada kelompok dan individu yang toleran di kultur hara. Marka OPH-12-1200 ini digunakan sebagai marker assisted
selection (MAS) pada generasi F4. Hasil seleksi populasi F4 berdasarkan daya
hasil per tanaman menunjukkan bahwa populasi F4 yang mempunyai bobot biji per tanaman yang tertinggi ada sebanyak 50 genotipe. Seleksi lanjutan menggunakan MAS pada 50 genotipe menghasilkan 20 genotipe yang membawa marka OPH-12-1200 untuk sifat toleransi tanah masam. Seleksi menggunakan marka OPH-12-1200 memberikan perbaikan karakter dan respon seleksi tertinggi terhadap karakter bobot biji per tanaman pada populasi F4. Karakter bobot biji per tanaman memiliki nilai diferensial seleksi sebesar 28.2% dan dugaan respon seleksi sebesar 18.33%.
Hasil penelitian perbandingan keragaan galur-galur F5 hasil seleksi karakter agronomi dan marka molekuler di tanah masam memperlihatkan bahwa keragaan karakter agronomi galur-galur F5 membawa marka OPH-12-1200 lebih baik dibandingkan dengan galur-galur F5 yang tidak membawa marka OPH-12-1200 pada kondisi tanah masam. Nilai tengah galur-galur F5 hasil seleksi dengan marka OPH-12-1200 berada di antara kedua tetua Argomulyo dengan Tanggamus untuk semua karakter agronomi, tetapi lebih tinggi dari nilai tengah varietas pembanding Anjasmoro, Willis, dan Pangrango. Galur AT/SSD/429, AT/SSD/1, AT/SSD/476, AT/SSD/503, AT/SSD/8 dan AT/SSD/423 merupakan galur-galur F5 hasil seleksi marka OPH-12-1200 yang mempunyai bobot biji per tanaman dan bobot 100 biji yang lebih tinggi dari tetua toleran tanah masam Tanggamus.
SUMMARY
ARVITA NETTI SIHALOHO. Genetic Analysis and Efficient Selection Using Single Seed Descent on Soybean for Adaptation of Acid Soil. Supervised by TRIKOESOEMANINGTYAS, DIDY SOPANDIE dan DESTA WIRNAS.
Hybridization is one of the methods for combining desired character and increasing soybean genetic variability. The result of hybridization would improve the probability to select for desired character From this research is expected to be selected lines tolerance to acid soil and yield potential in acid soil through crossing of Argomulyo with Tanggamus.
This research aimed (1) to obtain information about gene action and inheritance agronomy character soybean tolerance to aluminum (Al) stress in the acid soil and in nutrient culture; (2) to obtain RAPD markers linked to tolerance to Al stress through Bulk Segregrant analysis method; (3) to obtain information about the genetic diversity of character growth, yield components and yield in the F3 and F4 with single seed descent method; (4) to obtain F4 genotypes tolerant to acid soil based on selection on seed weight plant-1 and RAPD markers; (5) to obtain information about growth performance and yield components of F5 genotypes selected based on yield and RAPD markers linked to the Al stress tolerance.
The experiment for trait inheritance study was carried out in two locations, field in Jasinga, Bogor started in May until August 2012, and in a green house of University Farm, Bogor Agricultural University in March 2013. The study of genetic diversity in F3 and F4 generation with single seed descent method was carried out at experimental field of the Agency for Biotecnology and Genetic Rerouce, Bogor from October 2012 to May 2013. Selection of RAPD markers linked to aluminum tolerance using bulk segregant analysis was carried out in the Laboratory of Biomoleculer, Department of Agronomy and Horticulture IPB from April 2013 until June 2013. Performance comparison of F5 lines selected based on agronomy character and molecular markers in the acid soil, was carried out in acid soil UPTD Tenjo, Bogor, in October 2013 to January 2014 using an Augmented Design.
complementary epistasis but root dry weight is controlled by a few genes with additive gene action and complementary epistasis.
Study diversity of agronomy character in the soybean populations using single seed descent method showed that the mean agronomic character and genetic diversity of F4 population from single seed descent remains high as with the F3 population. The pattern of distribution of the phenotypes shows that there is transgessive segregant in F3 and F4 population for agronomic characters. The agronomic character of F3 population have heritability and genetic diversity coefficient was high and similar to the population of F4, except for the 100 seed weight had moderate value. The total number of pods correlated with grain weight per plant. Selection conducted directly on the character of grain weights per plant resulted in selection differential value higher than indirect selection using the character of number of total pods.
The selection of molecular markers linked to tolerance aluminum by bulk segregant analysis (BSA) resulted in only one consistent markers OPH-12 on locus 1200 that linked to tolerance aluminum on bulk and individuals tolerance in nutrient culture. Marker OPH-12-1200 was used as marker assisted selection (MAS) in F4 generation. The results of selection on F4 generation based on yield per plant showed that F4 generation had the highest seed weight per plant there as many as 50 genotypes. The next selection using markers OPH-12-1200 as MAS in 50 genotypes resulted in 20 genotypes that carry markers OPH-12-1200 linked to acid soil tolerant. Selection using OPH-12-1200 markers provide improved agronomic character and the selection response highest seed weight per plant in the F4 population. Character weights of seed per plant has a value of 28.2% selection differential and selection response alleged by 18.33%.
The results of performance of agronomic character under acid soil condition of F5 lines carrying marker OPH-12-1200 better than F5 lines that do not carry marker OPH-12-1200. The phenotypic values of the F5 lines selected by
OPH-12-1200 markers were between the two parent Argomulyo and Tanggamus for all agronomic characters, but higher than the mean varieties Anjasmoro, Willis and Pangrango. The F5 lines AT/SSD/429, AT/SSD/1, AT/SSD/476, AT/SSD/503, AT/SSD/8 dan AT/SSD/423 selected marker that OPH-12-1200 has a weight of seeds per plant and 100 seeds weight higher than the acid soil tolerant parental line Tanggamus.
© Hak Cipta Milik Institut Pertanian Bogor, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
ANALISIS GENETIK DAN EFISIENSI SELEKSI
MENGGUNAKAN
SINGLE SEED DESCENT
PADA KEDELAI
[
Glycine max
(L.) Merr.] UNTUK ADAPTASI TANAH MASAM
ARVITA NETTI SIHALOHO
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor
pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji luar komisi pada ujian tertutup :
1. Dr Willy B Suwarno, SP MSi 2. Dr Ir Maya Melati, MS MSc
Penguji luar komisi pada ujian terbuka :
1. Prof Dr Sumarno MSc
2. Dr Ir Hajrial Aswidinnoor MSc
Judul Disertasi : Analisis Genetik dan Efisiensi Seleksi Menggunakan Single
Seed Descent pada Kedelai [Glycine max (L.) Merr.] untuk
Adaptasi Tanah Masam Nama : Arvita Netti Sihaloho
NIM : A263100131
Disetujui oleh Komisi Pembimbing
Dr Ir Trikoesoemaningtyas, MSc Ketua
Prof Dr Ir Didy Sopandie, MAgr Dr Desta Wirnas, SP MSi
Anggota Anggota
Diketahui oleh
Ketua Program Studi Dekan Sekolah Pascasarjana
Pemuliaan dan Bioteknologi Tanaman
Dr Ir Yudiwanti Wahyu EK, MS Dr Ir Dahrul Syah, MScAgr
PRAKATA
Puji syukur penulis panjatkan ke hadirat Tuhan Yang Maha Esa, karena dengan rahmat dan karuniaNya, penulis dapat menyelesaikan penelitian dan
penulisan disertasi yang berjudul “Analisis Genetik dan Efisiensi Seleksi
Menggunakan Single Seed Descent pada Kedelai [Glycine max (L.) Merr.] untuk Adaptasi Tanah Masam”.
Penelitian dan penulisan disertasi ini berlangsung di bawah bimbingan Dr Ir Trikoesoemaningtyas, MSc selaku ketua komisi pembimbing, Prof Dr Ir Didy Sopandie, MAgr dan Dr Desta Wirnas, SP MSi selaku anggota komisi pembimbing. Pada kesempatan ini penulis menyampaikan terima kasih dan penghargaan yang tulus atas ilmu yang bermanfaat, motivasi, nasihat, kesabaran dan waktu yang telah diluangkan dalam mengarahkan dan membimbing penulis mulai dari perencanaan, penyusunan dan penyelesaian disertasi..
Ucapan terima kasih juga penulis sampaikan kepada Prof Dr Ir Munif Ghulamahdi, MS dan Prof Dr Muhammad Syukur, MSi masing-masing sebagai penguji pada Ujian Pra Kualifikasi Doktor atas saran-saran yang memberi bekal bagi penulis untuk melakukan penelitian ini. Ucapan terima kasih yang tulus juga penulis sampaikan kepada penguji luar ujian tertutup Dr Willy B Suwarno, SP MSi dan Dr Ir Maya Melati, MS MSi yang telah banyak memberikan saran dan masukan untuk perbaikan isi disertasi.Kepada Prof (R) Dr Sumarno, MSc danDr Ir Hajrial Aswindinnoor, MSc, penulis ucapkan terima kasih atas kesediaan menjadi penguji luar ujian terbuka. Saran dan masukan sangat diharapkan untuk penyempurnaan tulisan ini.
Penghargaan yang setinggi-tingginya serta rasa terima kasih yang tulus, penulis sampaikan kepada Rektor Universitas Sisingamangaraja XII Tapanuli dan Kepala KOPERTIS WIL.I SUMUT ACEH yang telah memberikan izin belajar, Dirjen DIKTI yang telah memberikan beasiswa BPPS, Rektor Institut Pertanian Bogor serta Dekan Pascasarjana IPB dan Ketua Program Studi Pemuliaan dan Bioteknologi Tanaman Sekolah Pascasarjana IPB yang telah menerima penulis untuk melanjutkan studi di Institut Pertanian Bogor, Kepala BB. Biogen Bogor beserta staf yang telah memfasilitasi lahan percobaan, Kerjasama IPB-IMHERE atas pembiayaan penelitian disertasi ini, Kepala University Farm, Institut Pertanian Bogor beserta staf di rumah kaca, Tehnisi Laboratorium Biomolekuler Departemen Agronomi dan Hortikultura yang telah banyak membantu, Kepala Kebun Percobaan Pengembangan Teknologi Lahan Kering, Jasinga-Bogor dan staf yang telah memfasilitasi lahan percobaan.
Penghargaan yang tulus penulis sampaikan kepada seluruh staf pengajar pada Departemen Agronomi dan Hortikultura, khususnya program studi Pemuliaan dan Bioteknologi Tanaman atas ilmu yang telah diberikan selama penulis mengikuti studi di sekolah Pascasarjana IPB. Terima kasih juga penulis sampaikan kepada teman-teman seperjuangan PBT angkatan 2010 dan sahabatku Tri Lestari, SP MSi dan Yuli Sulistyowati, SP MSi untuk persahabatan dan kebersamaan dalam berbagi ilmu, berbagi suka maupun duka. Semoga segala kebaikan dibalas berlipat oleh Tuhan Yang Maha Esa.
Sonia Febiola Hutabarat atas kasih sayang, doa dan suportnya bagi penulis untuk terus melangkah dan berjuang menyelesaikan studi. Semoga tulisan ini membawa manfaat dalam bidang pemuliaan tanaman dan menambah rasa syukur penulis kepada sang Pencipta.
DAFTAR ISI
Hal
DAFTAR TABEL ix
DAFTAR GAMBAR xi
DAFTAR LAMPIRAN xiii
PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 3
Tujuan Penelitian 4
Kebaruan Penelitian 4
Ruang Lingkup Penelitian 5
TINJAUAN PUSTAKA 7
Morfologi dan Fisiologi Tanaman Kedelai 7
Karakteristik Tanah Masam 8
Mekanisme Toleransi Tanaman terhadap Cekaman Al. 9 Keragaman dan Seleksi pada Tanaman Kedelai 10 Pola Pewarisan sifat Karakter Kualitatif dan Kuantitatif 13
Marka Molekuler 14
STUDI PEWARISAN SIFAT TOLERANSI KEDELAI TERHADAP
TANAH MASAM 17
Abstrak 17
Abstract 17
Pendahuluan 18
Metode Penelitian 19
Hasil dan Pembahasan 22
Kesimpulan
KEMAJUAN SELEKSI KEDELAI UNTUK TOLERANSI TANAH MASAM BERDASARKAN KARAKTER AGRONOMI
Abstrak
Abstract 37
Pendahuluan 38
Metode Penelitian 39
Hasil dan Pembahasan 40
Kesimpulan 5
KEMAJUAN SELEKSI KEDELAI UNTUK TOLERANSI ALUMINIUM
BERDASARKAN MARKA MOLEKULER 53
Abstrak 53
Abstract 53
Pendahuluan 54
Metode Penelitian 55
Hasil dan Pembahasan 57
Kesimpulan 63 PERBANDINGAN KERAGAAN GALUR-GALUR F5 HASIL SELEKSI
AGRONOMI DAN MARKA MOLEKULER 64
Abstrak 64
Abstract 64
Pendahuluan 65
Metode Penelitian 66
Hasil dan Pembahasan 67
Kesimpulan 73
PEMBAHASAN UMUM 74
KESIMPULAN 78
DAFTAR PUSTAKA 79
DAFTAR TABEL
Hal
1. Metode seleksi untuk tanaman menyerbuk sendiri 11 2. Nilai tengah karakter agronomi tetua Argomulyo (P1),
Tanggamus (P2) dan F2di tanah masam 23
3. Pendugaan aksi gen karakter agronomi populasi F2 (Argomulyo x
Tanggamus) di tanah masam 26
4. Pendugaan jumlah gen karakter agronomi populasi (Argomulyo x
Tanggamus) di tanah masam 26
5. Nilai pendugaan komponen ragam dan heritabilitas arti luas
populasi F2 pada tanah masam 27
6. Nilai tengah karakter akar tetua Argomulyo (P1), Tanggamus
(P2) dan F2 pada cekaman aluminium di kultur hara 28 7. Pendugaan aksi gen karakter pertumbuhan populasi F2
(Argomulyo x Tanggamus) pada cekaman aluminium
di kultur hara 30
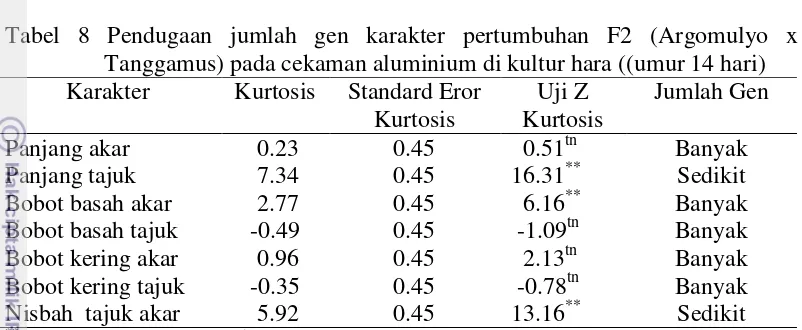
8. Pendugaan jumlah gen karakter pertumbuhan populasi F2 (Argomulyo x Tanggamus) pada cekaman aluminium
di kultur hara 31
9. Nilai pendugaan komponen ragam dan heritabilitas arti luas populasi
F2 pada cekaman aluminium di kultur hara 32
10.Nilai tengah karakter akar tetua Argomulyo (P1), Tanggamus (P2)
dan F2 pada cekaman aluminium di root re-growth 33 11.Pendugaan aksi gen karakter akar F2 hasil (Argomulyo x
Tanggamus) pada cekaman aluminium di rootre-growth 34 12.Pendugaan jumlah gen karakter akar F2 hasil (Argomulyo x
Tanggamus) pada cekaman aluminium di root re-growth 35 13.Nilai pendugaan komponen ragam dan heritabilitas arti luas populasi
F2 pada cekaman aluminium di root re-growth 35
14.Hasil analisis nilai tengah karakter agronomi pada populasi F3
(Argomulyo x Tanggamus) pada kondisi tidak tercekam 42 15.Hasil analisis nilai tengah karakter agronomi pada populasi F4
(Argomulyo x Tanggamus) pada kondisi tidak tercekam 42 16.Hasil analisis komponen ragam, koefisien keragaman genetik dan
heritabilitas arti luas karakter agronomi pada populasi F3 dan F4
(Argomulyo x Tanggamus) pada kondisi tidak tercekam 43 17.Matriks korelasi karakter agronomi pada populasi F4 hasil
persilangan Argomulyo dan Tanggamus 49
18.Perbaikan karakter agronomi berdasarkan seleksi langsung populasi
F4 (Argomulyo x Tanggamus) pada kondisi tidak tercekam 49 19.Perbaikan karakter agronomi berdasarkan seleksi tidak langsung
populasi F4 (Argomulyo x Tanggamus) pada kondisi tidak tercekam 50 20.Keragaan bobot biji per tanaman 30 genotipe hasil seleksi
berdasarkan karakter bobot biji per tanaman 51
21.Nilai fenotipe panjang akar populasi F2 tercekam aluminium
22.Rekapitulasi hasil validasi primer pada tetua peka (Argomulyo) dan
tetua toleran (Tanggamus) 58
23.Rekapitulasi hasil validasi primer pada bulk toleran dan bulk peka
genotipe F2 (Argomulyo x Tanggamus) 59
24.Keragaan bobot biji per tanaman 20 genotipe F4 hasil seleksi
berdasarkan karakter bobot biji per tanaman dan marka molekuler 61 25.Diferensial seleksi berdasarkan marka molekuler toleran aluminium
populasi F4 (Argomulyo x Tanggamus) pada kondisi tidak tercekam 62
26.Sidik ragam augmented design 66
27.Rekapitulasi sidik ragam karakter agronomi genotipe F5 dan kontrol
pada tanah masam 67
28.Keragaan nilai tengah karakter agronomi 30 galur F5 terseleksi marka OPH-12-1100 dengan lima varietas pembanding di tanah masam 68 29.Keragaan karakter pertumbuhan 30 galur F5 hasil seleksi marka dan
kedua tetua di tanah masam 68
30.Keragaan karakter pertumbuhan 30 galur F5 hasil seleksi marka dan
kedua tetua di tanah masam 70
31.Nilai tengah karakter agronomi dan komponen hasil galur F5 ada
DAFTAR GAMBAR
Hal
1. Bagan alir penelitian 6
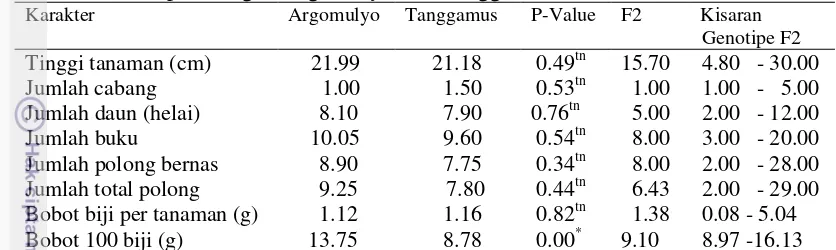
2. Pola sebaran data tinggi tanaman dan jumlah daun populasi F2
(Argomulyo x Tanggamus) pada tanah masam 24
3. Pola sebaran data bobot 100 biji dan jumlah cabang produktif
populasi F2 (Argomulyo x Tanggamus) pada tanah masam 24 4. Pola sebaran data jumlah polong bernas dan jumlah buku populasi
F2 (Argomulyo x Tanggamus) pada tanah masam 25 5. Pola sebaran data bobot biji per tanaman dan jumlah total polong
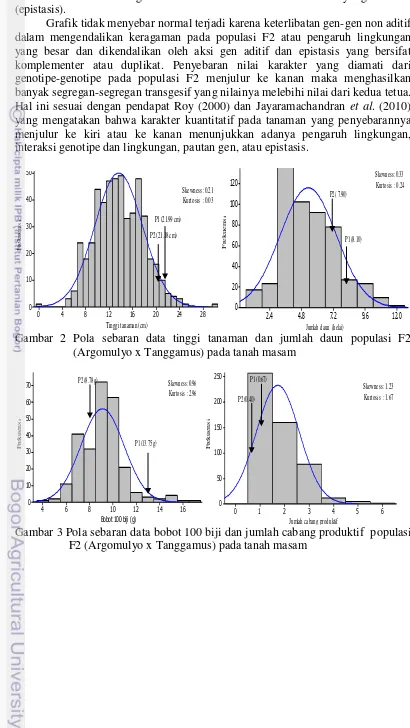
populasi F2 (Argomulyo x Tanggamus) pada tanah masam 25 6. Pola sebaran data panjang akar dan panjang tajuk populasi
F2 (Argomulyo x Tanggamus) tercekam aluminium pada stadia bibit
di kultur hara 29
7. Pola sebaran data nisbah panjang tajuk akar populasi F2
(Argomulyo x Tanggamus) dalam keadaan tercekam aluminium
pada stadia bibit di kultur hara 29
8. Pola sebaran data berat basah dan berat kering tajuk
populasi F2 (Argomulyo x Tanggamus) dalam keadaan tercekam
aluminium pada stadia bibit di kultur hara 29
9. Pola sebaran data berat basah dan berat kering akar populasi F2 (Argomulyo x Tanggamus) tercekam aluminium pada stadia bibit
di kultur hara 30
10.Pola sebaran data berat basah dan berat kering akar populasi F2
(Argomulyo x Tanggamus) setelah tercekam aluminium pada stadia bibit 33 11.Pola sebaran data panjang akar populasi F2 (Argomulyo x
Tanggamus) keadaan setelah tercekam dan setelah recovery 34 12.Pola sebaran data root re-growth populasi F2
(Argomulyo x Tanggamus pada stadia bibit 36
13.Pola sebaran data tinggi tanaman populasi F3 dan F4
(Argomulyo x Tanggamus pada kondisi tidak tercekam 44 14.Pola sebaran data jumlah cabang produktif populasi F3 dan F4
(Argomulyo x Tanggamus) pada kondisi tidak tercekam 44 15.Pola sebaran data jumlah buku produktif populasi F3 dan F4
(Argomulyo x Tanggamus) pada kondisi tidak tercekam 45 16.Pola sebaran data jumlah total polong populasi F3 dan F4
(Argomulyo x Tanggamus) pada kondisi tidak tercekam 45 17.Pola sebaran data jumlah polong bernas populasi F3 dan F4
(Argomulyo x Tanggamus) pada kondisi tidak tercekam 45 18.Pola sebaran data jumlah polong hampa populasi F3 dan F4
(Argomulyo x Tanggamus) pada kondisi tidak tercekam 46 19.Pola sebaran data bobot biji per tanaman populasi F3 dan F4
(Argomulyo x Tanggamus) pada kondisi tidak tercekam 46 20.Pola sebaran data bobot 100 biji populasi F3 dan F4
(Argomulyo x Tanggamus) pada kondisi tidak tercekam 46 21.Pola sebaran data sink size populasi F3 dan F4
22.Keragaan tanaman kedelai generasi F3 dan F4
(Argomulyo x Tanggamus) pada kondisi tidak tercekam 47 23.Keragaan akar dan tajuk genotipe peka (S) dan toleran (T) kedelai
populasi F2 (Argomulyo x Tanggamus) pada stadia
bibit di kultur hara dengan perlakuan 1,5 mM Al 57 24.Pola sebaran panjang akar sebelum dan setelah distandarisasi pada
populasi F2 (Argomulyo x Tanggamus) 57
25.Validasi primer RAPD yang polimorfik pada bulk toleran dan peka
genotipe F2 (Argomulyo x Tanggamus) 59
26.Hasil amplifikasi DNA marka OPH-12 konsisten terpaut
pada genotipe toleran F2 (Argomulyo x Tanggamus) 60 27.Hasil amplifikasi DNA marka OPH-12 pada populasi F4
(Argomulyo x Tanggamus) 61
28.Sebaran galur-galur F5 Argomulyo x Tanggamus untuk karakter
DAFTAR LAMPIRAN
Hal 1. Sifat kimia contoh tanah pada lokasi penelitian di tanah masam
milik petani Jasinga, Kabupaten Bogor Barat 88
2. Sifat kimia contoh tanah pada lokasi penelitian di kebun percobaan
Balai Besar Biogen Kabupaten Bogor 89
3. Sifat kimia contoh tanah pada lokasi penelitian di tanah masam kebun percobaan UPT Pengembangan Teknologi Lahan Kering
wilayah Tenjo Kabupaten Bogor Barat 90
4. Data iklim di Jasinga tahun 2012 91
5. Data iklim di BB Biogen, Cimanggu tahun 2012 92 6. Data iklim di BB Biogen, Cimanggu tahun 2013 93 7. Data iklim di percobaan UPT Pengembangan Teknologi Lahan Kering
PENDAHULUAN
Latar Belakang
Kedelai (Glycine max (L.) Merr.) merupakan komoditi pangan strategis di Indonesia yang kebutuhannya terus meningkat sejalan pertumbuhan jumlah penduduk. Kedelai adalah salah satu sumber protein utama dan banyak disukai masyarakat. Biji kedelai mengandung protein tinggi, yaitu sekitar 40-46 g per 100 g bahan kering (Muhidin 2004).
Produksi kedelai di Indonesia pada tahun 2013 hanya mencapai 892.602 ton. Jumlah ini belum memenuhi target pemerintah yakni 2.2 juta ton. Usaha pemerintah untuk memenuhi kebutuhan kedelai, selain mengimpor sebesar 85% adalah dengan meningkatkan produktivitas dan menambah luas areal sebesar 100 ribu ha per tahun (BPS 2013). Upaya perluasan areal tanam kedelai menghasilkan peningkatan produksi kedelai nasional. Tahun 2013 luas areal tanam kedelai hanya mencapai 550.793 ha dengan produksi 779.992 ton, meningkat menjadi 601.237 ha dengan produksi mencapai 892.602 ton pada tahun 2014 (ARAM I BPS).
Salah satu usaha untuk meningkatkan produksi kedelai nasional adalah memperluas areal tanam dengan memanfaatkan lahan-lahan marginal yang potensinya sangat besar. Menurut Mulyani et al. (2011), luas lahan kering di Indonesia mencapai 148 juta hektar dan diperkirakan 102.8 juta hektar di antaranya berupa tanah masam. Diantara 102.8 juta hektar tanah masam tersebut, hanya 6 juta hektar berpotensi untuk areal tanaman pangan (Dakhyar et al. 2012).
Perluasan areal panen kedelai di Indonesia untuk mencapai swasembada produksi, satu-satunya adalah memanfaatkan tanah masam. Akan tetapi tanah masam kurang sesuai untuk tanaman kedelai karena pH nya yang rendah, terjadinya keracunan Al dan ketersediaan unsur hara makro yang rendah. Selain upaya ameliorasi, tanah masam agar menjadi sesuai untuk tanaman kedelai perlu, disediakan varietas kedelai yang toleran tanah masam dan memiliki mutu biji yang bagus, ukuran biji cukup besar supaya diterima oleh pasar.
Faktor pembatas produksi tanaman kedelai pada tanah masam adalah kesuburan tanah yang rendah karena toksisitas aluminium (Al) dan defisiensi hara-hara makro di antaranya fosfor (P), kalsium (Ca) dan magnesium (Mg) (Marschner 1995). Keracunan Al merupakan faktor pembatas utama karena dapat menyebabkan terganggunya pertumbuhan akar sehingga tanaman mengalami hambatan dalam penyerapan air dan hara yang akhirnya berpengaruh terhadap pertumbuhan dan produktivitas tanaman (Kochian et al. 2004).
bobot kering akar sebesar 40 g, mengalami penurunan bobot kering akar sekitar 46.67%.
Pendekatan pemuliaan yang dapat dilakukan untuk mendapatkan produktivitas yang tinggi di tanah masam adalah dengan menghasilkan varietas kedelai toleran terhadap cekaman Al. Kementerian Pertanian (2012) telah melepas 74 varietas kedelai, beberapa di antaranya toleran terhadap tanah masam seperti Sibayak, Ratai, Nanti, dan Tanggamus. Umumnya varietas toleran tanah masam yang telah dilepas tersebut mempunyai biji ukuran kecil.
Adie dan Krisnawati (2007) melakukan pengelompokan biji kedelai berdasarkan ukuran biji dan menyatakan bahwa varietas Sibayak, Ratai, Nanti, dan Tanggamus termasuk kelompok berbiji kecil (< 10 g/100 biji). Saat ini konsumen lebih menyukai kedelai berbiji besar. Oleh karena itu, perlu dilakukan pengembangan varietas kedelai berbiji besar untuk kesesuaian budidaya di tanah masam. Keragaman genetik kedelai untuk sifat berbiji besar dan toleran aluminium masih rendah sehingga perlu dilakukan persilangan antara varietas toleran tanah masam seperti Tanggamus dengan varietas berbiji besar (≥ 14 g/100 biji), seperti Argomulyo untuk memperoleh galur yang toleran tanah masam berbiji besar.
Persilangan diharapkan akan menghasilkan rekombinasi dan keragaman genetik tinggi, sehingga tersedia materi genetik untuk dilakukan seleksi. Keberhasilan seleksi atau efisiensi seleksi untuk meningkatkan daya hasil dan toleransi tanah masam ditunjukkan oleh perolehan kemajuan genetik dari karakter yang diseleksi. Kemajuan genetik yang diperoleh sangat tergantung kepada variabilitas genetik dan heritabilitas (Roy 2000). Perolehan kemajuan genetik dapat dimaksimalkan dengan memilih karakter seleksi yang sesuai, yaitu yang memiliki nilai heritabilitas tinggi dan berkorelasi dengan hasil, untuk itu perlu dilakukan studi genetik untuk memperoleh pola pewarisan dari karakter yang ingin diperbaiki.
Seleksi untuk adaptasi terhadap lingkungan bercekaman harus dilakukan di lingkungan target dengan tujuan untuk dapat memaksimalkan ekspresi gen-gen yang mengendalikan daya hasil maupun daya adaptasi (Ceccareli et al. 2007). Seleksi untuk adaptasi terhadap lingkungan bercekaman dapat dilakukan berdasarkan fenotipe dan gabungan antara fenotipe dengan marka molekuler. Seleksi berdasarkan fenotipe menjadi sulit karena nilai heritabilitas yang rendah dan adanya interaksi antara genotipe dan lingkungan sehingga untuk meningkatkan efisiensi seleksi dapat dilakukan seleksi berdasarkan fenotipe dan marka molekuler (Bernando 2002).
Seleksi menggunakan karakter agronomi lebih praktis dan cepat untuk karakter-karakter yang memiliki nilai heritabilitas dan korelasi yang tinggi karena pengamatan dapat dilakukan secara langsung, tetapi karakter agronomi umumnya bersifat kuantitatif dan ekspresinya dipengaruhi oleh kondisi lingkungan.
dan ketahanan tanaman padi terhadap kekeringan (Youssef et al. 2010). Marka molekuler yang dapat dipergunakan sebagai marka seleksi harus terlebih dahulu diseleksi, sehingga marka molekuler yang terpaut sifat yang diinginkan yang dipergunakan sebagai marka seleksi.
Salah satu metode seleksi marka molekuler adalah bulk segregrant
analysis. Metode bulk segregant analysis (BSA) dapat digunakan untuk
menyeleksi marka yang terpaut karakter yang diinginkan. Metode bulk segregant
analysis (BSA) merupakan metode skrining marka yang didasarkan atas
pengelompokan DNA dari individu-individu tanaman pada populasi segregan. Metode ini dapat menyeleksi marka-marka molekuler yang terpaut erat dengan lokus yang dituju secara lebih cepat dan tepat (Zhang et al. 2008).
Penelitian ini dilakukan untuk mempelajari pola pewarisan sifat kedelai yang akan digunakan sebagai karakter seleksi untuk menghasilkan genotipe atau galur kedelai yang toleran tanah masam dan berbiji besar. Penelitian ini menjadi penting dilakukan karena masih terbatasnya informasi mengenai studi pewarisan sifat toleran kedelai di tanah masam, dan penggunaan marka RAPD sebagai alat bantu seleksi kedelai pada populasi bersegregasi untuk adaptasi di tanah masam juga masih perlu dilakukan.
Populasi hasil persilangan antara Argomulyo dengan Tanggamus akan diseleksi pada generasi lanjut (F5) karena penelitian ini menggunakan metode seleksi single seed descent untuk pembentukan galur. Metode seleksi single seed
descent ini digunakan untuk karakter yang memiliki nilai heritabilitas rendah.
Single seed descent atau turunan biji tunggal merupakan metode
pembentukan galur yang ditujukan untuk tetap menjaga keragaman genetik selama populasi bersegregasi dari satu generasi ke generasi berikutnya dengan tidak menambah ruang dan area pertanaman serta tanpa seleksi. Pemilihan genotipe-genotipe harapan dilakukan ketika telah terjadi fiksasi dari alel-alel karakter yang diinginkan pada generasi lanjut. Metode pembentukan galur secara
single seed descent ini dapat juga digunakan untuk studi genetik terkait dengan
pemetaan lokus karakter kuantitatif (Falconer dan Mackay 1996).
Perumusan Masalah
Luas areal tanam untuk pertanaman kedelai terus menurun dari tahun ke tahun sehingga perlu dilakukan ekstensifikasi pertanaman di areal tanah-tanah marginal yang potensial, misalnya tanah masam. Masalah yang dihadapi dalam pengembangan kedelai di tanah masam adalah belum tersedianya varietas toleran tanah masam berbiji besar yang lebih disukai oleh petani, sehingga perlu pengembangan varietas toleran tanah masam berbiji besar untuk meningkatkan produktivitas.
Seleksi karakter kuantitatif dapat ditingkatkan akurasi seleksinya dengan menggunakan marka molekuler, misalnya marka RAPD dengan metode bulk
segregant analyisis (BSA). Penggunaan marka molekuler RAPD dalam
melakukan seleksi merupakan salah satu cara mengembangkan perbaikan metode seleksi pada tanah masam karena seleksi menggunakan marka molekuler tidak dipengaruhi lingkungan dan memiliki nilai heritabilitas yang tinggi.
Tujuan Penelitian
Tujuan umum penelitian ini adalah memperoleh informasi tentang dasar genetik dan karakter seleksi untuk perbaikan daya adaptasi dan potensi hasil di tanah masam. Secara khusus penelitian ini bertujuan untuk:
1. Memperoleh informasi tentang aksi gen dan pewarisan sifat toleransi karakter agronomi kedelai terhadap cekaman aluminium di tanah masam dan di kultur hara.
2. Memperoleh marka RAPD terpaut toleransi cekaman aluminium melalui metode bulk segregrant analysis.
3. Memperoleh informasi tentang keragaman genetik karakter pertumbuhan, komponen hasil dan hasil pada F3 dan F4 dengan metode single seed descent.
4. Memperoleh galur kedelai yang toleran tanah masam melalui seleksi daya hasil dan marka RAPD.
5. Memperoleh informasi tentang keragaan pertumbuhan dan daya hasil serta membandingkan kemajuan genetik galur F5 terseleksi berdasarkan daya hasil dan marka RAPD terpaut toleransi cekaman Al.
Kebaruan Penelitian
Penelitian ini dilakukan untuk memperoleh informasi yang sesuai dengan tujuan penelitian dan belum pernah dilakukan di Indonesia. Kriteria seleksi toleran tanah masam yang telah dilakukan selama ini menggunakan kriteria keragaan pertumbuhan agronomi dan berdasarkan gejala keracunan Al. Oleh karena itu, penelitian yang telah dilakukan melalui berbagai percobaan, menghasilkan:
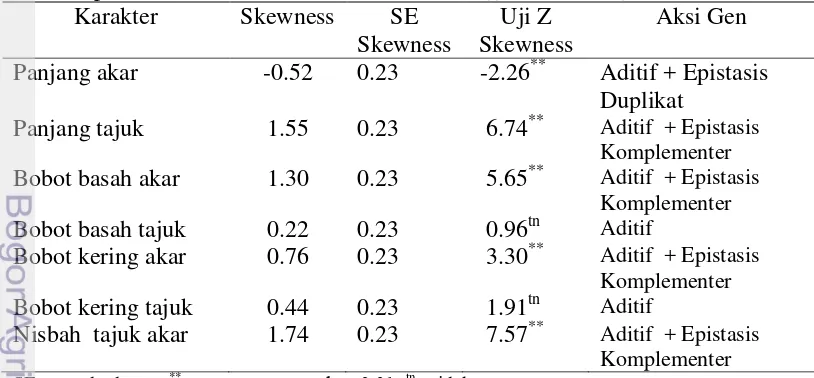
1. Informasi bahwa pewarisan sifat toleransi kedelai terhadap cekaman aluminium dan tanah masam dikendalikan banyak gen dengan aksi gen aditif epistasis, didapatkan informasi bahwa karakter seleksi panjang akar dikendalikan banyak gen dengan aksi gen aditif epistasis.
2. Satu marka RAPD terpaut sifat toleran cekaman aluminum pada tanaman kedelai, yaitu marka OPH-12-1200 yang diperoleh pada populasi F2 dengan menggunakan metode bulk segregant analyisis (BSA) dan marka OPH-12-1200 dapat digunakan sebagai MAS (marker assisted selection) untuk meningkatkan akurasi seleksi pada generasi lanjut.
Ruang Lingkup Penelitian
Penelitian ini mencakup empat tahap, yaitu studi pewarisan sifat toleran cekaman aluminium di tanah masam dan kultur hara pada generasi F2, penggaluran populasi bersegregasi dengan metode single seed descent (SSD) sampai generasi F4 dan seleksi generasi F4 berdasarkan karakter daya hasil, seleksi marka RAPD terpaut sifat toleran cekaman aluminium. Marka yang diperoleh akan dipergunakan dalam seleksi berbasis marka molekuler (marker
assisted selection) pada populasi F4 untuk menghasilkan galur-galur harapan
Gambar 1 Kerangka penelitian untuk pewarisan sifat toleransi dan seleksi kedelai secara karakter hasil dan marka pada tanah masam
SSD F4
Studi Pewarisan
Seleksi Penggaluran
SSD
-Parameter Genetik -Karakter Akar SSD F3
Galur-Galur F5 Toleran Masam Berdasarkan Hasil dan Marka Molekuler
Kultur Hara Tanah Masam
Persilangan
F1
F2
-Parameter Genetik -Karakter Agronomi
Karakter Seleksi Hasil dan Marka Molekuler Seleksi Marka
TINJAUAN PUSTAKA
Morfologi dan Fisiologi Tanaman Kedelai
Kedelai [Glycine max (L.) Merr] bukan tanaman asli Indonesia, diduga tanaman kedelai berasal dari daratan tengah dan utara Cina. Hal ini didasarkan pada adanya penyebaran Glycine usseriensis spesies yang diduga sebagai tetua
Glyine max, sebab bukti sitogenetika menunjukkan bahwa Glycine max dan
Glycine usseriensis tergolong spesies yang sama (Sharma et al.1993).
Glycine max merupakan tanaman semusim, warna bunga putih atau ungu,
memiliki bentuk daun bulat dan lancip. Bentuk biji bervariasi tergantung pada varietas tanaman, yaitu bulat, agak gepeng dan bulat telur. Setiap biji kedelai mempunyai ukuran bervariasi, mulai dari kecil (sekitar 7-9 g/100 biji), sedang (10-13 g/100 biji) dan besar (> 13 g/100 biji).
Spesies Glycine max memiliki jumlah kromosom somatik 2n=2x=40, sedangkan subgenus Glycine lain seperti Glycine bracteata memiliki kromosom 2n=2x=44 dan Glycine canescens yang tidak dibudidayakan memiliki kromosm 2n=4x=80 (Burton 1997).
Pertumbuhan tanaman kedelai dibagi dalam dua fase yaitu fase vegetatif dan fase generatif (reproduksi). Fase vegetatif diawali saat tanaman muncul dari tanah dan kotiledon belum membuka (Ve-Vc), V1-V6 berdasarkan jumlah daun trifoliat telah berkembang penuh dan fase ini diakhiri setelah mulai terbentuknya bunga sebagai organ reproduktif. Fase reproduksi dikelompokkan ke dalam tiga fase yaitu fase pembungaan, pembentukan polong dan pematangan biji (R1 –R8). Fase pembungaan (R1-R2) dicirikan oleh munculnya dua atau lebih bunga mekar, sedangkan fase pembentukan polong (R3-R6) dicirikan oleh mulai terbentuk polong berukuran 5 mm pada batang utama dan berakhir dengan terbentuknya polong berukuran 2 cm dan biji dalam polong telah mengisi penuh rongga polong. Fase pematangan polong (R7-R8) diawali adanya satu polong yang telah berwarna kuning (matang), fase ini sering disebut sebagai fase masak fisiologis. Fase pematangan biji berakhir jika 90% polong telah berwarna coklat (matang) dan siap untuk dipanen (Fehr et al. 1971).
Komponen lingkungan yang menjadi penentu keberhasilan usaha produksi kedelai adalah faktor iklim dan tanah. Faktor iklim yang menentukan pertumbuhan kedelai adalah panjang hari, suhu, kelembaban udara dan curah hujan, sedangkan faktor tanah yang menentukan pertumbuhan kedelai adalah pH tanah.
Kedelai termasuk tanaman hari pendek, yaitu tidak mampu berbunga bila panjang hari atau lama penyinaran melebihi 16 jam dan mempercepat pembungaan bila lama penyinaran kurang dari 12 jam. Tanaman hari pendek pada kedelai bermakna bahwa lama penyinaran yang semakin pendek akan merangsang pembungaan lebih cepat (Sumarno dan Manshuri 2007).
Kelembaban udara yang optimal bagi tanaman kedelai berkisar antara 75-90% selama periode tanaman tumbuh hingga stadia pengisian polong dan kelembaban udara 60-75% pada periode pematangan polong hingga panen. Pengaruh langsung kelembaban udara terhadap pertumbuhan dan perkembangan tanaman kedelai tidak terlalu besar, tetapi secara tidak langsung berpengaruh terhadap perkembangan hama dan penyakit (Sarwanto 2007).
Tanaman kedelai sangat efektif dalam memanfaatkan air yang berasal dari kelembaban udara. Curah hujan yang dibutuhkan tanaman kedelai tergantung dari kemampuan tanah menyimpan air, besar penguapan dan kedalaman lapisan olah tanah. Tanaman kedelai dapat tumbuh dengan baik pada curah hujan 120-150 mm per bulan tetapi pada curah hujan lebih dari 300 mm per bulan, pertumbuhan dan produksi kedelai sangat baik bila pengelolaan air dilakukan dengan baik (Agus et al. 2005)
Tanaman kedelai dapat tumbuh baik pada tanah yang sedikit masam sampai mendekati netral, yaitu pH 5.5-6.5 karena pada kisaran pH tersebut hara makro dan mikro tersedia bagi tanaman kedelai. Pada tanah yang bereaksi masam atau pH kurang dari 5.5 hara makro seperti P dan Ca tidak mudah tersedia, sedangkan pada tanah yang bereaksi basa atau pH lebih besar dari 7, tanaman kedelai mengalami gejala klorosis karena unsur Fe menjadi tidak tersedia bagi tanaman (Wijaya 2008).
Sekitar 74 varietas unggul kedelai telah dilepas pemerintah tetapi varietas unggul toleran tanah masam masih sedikit, yaitu Tanggamus, Ratai, Nanti, Sibayak dan Rajabasa. Pada penelitian ini varietas Tanggamus dan Argomulyo yang digunakan sebagai tetua. Varietas Tanggamus merupakan varietas yang toleran tanah masam, dapat tumbuh dengan baik pada pH < 5, mempunyai tinggi tanaman 67 cm, umur berbunga 35 HST, umur panen 85 HST, jumlah cabang produktif 3-5, jumlah polong 90, biji sedang 10-11 g100 biji-1 dan produksi 2.5 ton ha-1.
Varietas Argomulyo merupakan varietas yang direkomendasikan untuk tanah sawah, tidak dapat tumbuh dengan baik pada tanah masam, mempunyai tinggi tanaman 40 cm, umur berbunga 35 HST, umur panen 80 HST, jumlah cabang 3-4, jumlah polong 45, biji besar14 g100 biji-1 dan produksi 1.5-2.0 ton ha-1.
Penelitian mengenai ketahanan berbagai varietas kedelai terhadap tanah masam telah banyak dilakukan, seperti Delhaize dan Ryan (1995) serta Sopandie
et al. (2003).
Karakteristik Tanah Masam
Bentuk-bentuk Al di dalam tanah dapat berupa ion trivalen yaitu Al3+, bentuk hidroksida seperti Al(OH)2+, Al(OH)2+, Al(OH)3, Al(OH)4- atau
berasosiasi dengan berbagai senyawa organik dan anorganik seperti SO43-, SO42-,
asam organik, protein dan lipid (Delhaize dan Ryan, 1995). Ion Al3+ merupakan bentuk yang paling toksik dan mendominasi di tanah masam terutama pada pH <5 (Matsumoto 2000).
Tanah-tanah yang masih banyak tersedia untuk pertanian di Indonesia adalah tanah-tanah yang bereaksi masam yang memiliki nilai pH kurang dari 5.5 dan miskin unsur hara, seperti tanah ultisol. Tanah ultisol mempunyai potensi yang besar untuk dijadikan lahan pertanian. Luas tanah ultisol di Indonesia mencapai 45.8 juta ha atau 25% luas tanah Indonesia (Subagyo et al. 2004). Menurut Subandi (2007) tanah ultisol umumnya mempunyai pH rendah dengan kelarutan Al, Fe, dan Mn tinggi sehingga dapat meracuni tanaman. Jenis tanah ini biasanya miskin unsur hara esensial makro seperti N, P, K, Ca, dan Mg, unsur hara mikro Zn, Mo, Cu, dan B, serta bahan organik tanah seperti humus. Secara umum tanah ultisol atau podsolik merah kuning mempunyai kejenuhan Al yaitu sebesar 33-86%, namun hasil penelitian menunjukkan bahwa beberapa contoh tanah ultisol mengandung kejenuhan Al yaitu sekitar 16-75% (Prasetyo et al. 2006)
Penyebaran tanah masam di Indonesia cukup luas terutama pada wilayah beriklim basah dan pulau Sumatera adalah pulau dengan luasan tanah masam sekitar 9.5 juta hektar (Subagyo et al. 2004). Tanah-tanah masam itu ditemukan baik pada lahan kering maupun lahan basah (rawa/pasang surut) dan dapat berasal dari bahan/batuan mineral maupun bahan non mineral (organik) yaitu hasil pelapukan makhluk hidup. Luas tanah masam lahan kering ditemukan sekitar 45.8 juta hektar lebih luas dibandingkan tanah masam lahan basah (Mulyani et al.
2011).
Cekaman Al terhadap tanaman terutama menekan pertumbuhan akar sehingga akar memendek, tebal dan rapuh. Terhambatnya pertumbuhan akar tersebut karena pada inti sel, Al berasosiasi dengan DNA dan menghentikan proses pembelahan sel meristem apikal. Bagian yang paling peka terhadap keracunan Al terutama pada bagian ujung akar (tudung akar, meristem dan zona pemanjangan), hal ini karena bagian tersebut mengakumulasi Al lebih banyak (Maschner 1995).
Mekanisme Toleransi Tanaman Terhadap Cekaman Tanah
Masam
Aluminium (Al) merupakan ion yang menghambat pertumbuhan dan produktivitas tanaman di tanah mineral masam. Walaupun Al menghambat proses metabolisme dan pertumbuhan tanaman, akan tetapi sampai batas kritis keracunan sekitar 1.33 me Al 100 g-1 tanah Al-dapat ditukar, pengaruh Al dapat ditoleransi oleh tanaman kedelai yang toleran (Kasim 2000).
Ion Al yang telah terikat pada dinding sel dapat masuk ke plasma membran melalui ion channel. Al yang ada pada pada membran akan terikat pada protein dan fosfolipid sehingga membran menjadi kaku dan fluiditasnya menurun. Terikatnya Al pada protein membran, juga menyebabkan terganggunya sirkulasi berbagai ion hara penting (Kochian et al. 2005).
Sebagian Al yang tidak terikat di plasma membran masuk ke sitoplasma melalui transport aktif sehingga Al masuk ke inti sel dan menghalangi metabolisme asam nukleat dengan cara Al mengikat sangat kuat fosfat yang ada pada kedua utas DNA yang mengakibatkan gagalnya pemisahan DNA utas ganda saat proses replikasi. Hasil penelitian Silva et al. (2000) memperlihatkan terhambatnya pembelahan sel di daerah meristematis ujung akar kedelai disebabkan oleh akumulasi Al pada inti sel.
Gejala yang umum dijumpai akibat cekaman Al adalah terjadinya defisiensi nutrisi akibat sebagian besar unsur hara makro dan mikro tidak tersedia karena pH tanah rendah dan terikat dengan Al sehingga tanaman menjadi kerdil (Taiz dan Zeiger 2002). Pengaruh cekaman Al3+ tidak sama pada setiap spesies, bahkan pada tanaman dalam satu spesies.
Marschner (1995) dan Blum (1996) mengatakan bahwa tanaman yang mampu beradaptasi terhadap cekaman Al adalah tanaman yang memiliki suatu mekanisme eksternal atau internal sehingga tanaman tersebut memiliki kemampuan untuk menekan pengaruh buruk keracunan Al dengan cara mengurangi absropsi Al dan immobilisasi Al di permukaan dinding sel, yaitu Al akan terikat pada senyawa pektin atau komponen dinding sel yang bermuatan negatif pada jaringan epidermis dan tudung akar.
Menurut Utama (2004) dan Li et al. (2000) tanaman yang toleran Al adalah tanaman yang memiliki berbagai cara untuk menetralkan toksik Al setelah diserap tanaman dengan cara kelatisasi Al oleh asam organik, protein atau ligand organik lainnya di sitoplasma, serta sanggup menciptakan keadaan tidak masam di daerah perakaran dengan cara mengeluarkan asam organik melalui akar untuk mengikat Al3+ sehingga sistem yang ada di dalam dan ruang antar sel terhindar dari pengaruh Al. Selain itu, menurut Prasetiyono dan Taslih (2003) tanaman yang toleran Al dapat mentranslokasi ion Al ke bagian atas tanaman sedikit, karena sebagian besar ditoleransi di akar, dengan cara diakumulasi di akar terutama di korteks dan epidermis. Al bisa lolos ke tajuk melalui jaringan meristem akar dan pembuluh akar (stele) dan tanaman memiliki mekanisme anion yang diserap lebih banyak dari pada kation sehingga total anion yang diserap lebih banyak dari total kation menyebabkan ekskresi OH- sehingga pH media meningkat, akibatnya ion aluminium tidak sanggup menghambat serapan Ca, Mg dan K.
Hasil penelitian pada tanaman kedelai toleran memperlihatkan kemampuannya mengubah pH media 2-2.5 kali lebih besar dibandingkan tanaman peka, demikian juga dengan kemampuan eksudasi asam sitrat dan malat, yaitu 2-3 kali lebih besar dibandingkan spesies peka (Utama et al. 2005).
Keragaman dan Seleksi pada Tanaman Kedelai
mutasi (Poehlman dan Sleper 2006). Rekombinasi gen dapat terjadi melalui hibridisasi yaitu persilangan antara tanaman yang berbeda secara genetik. Melalui hibridisasi, pemulia tanaman dapat menggabungkan sifat-sifat yang diinginkan dari tetua ke keturunannya.
Persilangan diharapkan akan menghasilkan keragaman genetik yang tinggi. Oleh karena itu dapat dilakukan seleksi untuk mendapatkan genotipe-genotipe dengan karakter yang diinginkan. Seleksi merupakan salah satu kegiatan utama dalam pemuliaan tanaman. Seleksi adalah prosedur memilih sejumlah individu dari suatu populasi dan membiarkannya membentuk generasi baru. Seleksi merupakan salah satu upaya mengubah frekuensi gen dengan mengambil yang diinginkan dan membuang yang tidak diinginkan (Falconer dan Mackay 1996).
[image:30.595.109.509.295.659.2]Seleksi pada tanaman menyerbuk sendiri dapat dibagi menjadi beberapa metode, yaitu seleksi silsilah (pedigree), Seleksi bulk dan seleksi turunan biji tunggal (single seed descent, SSD) (Poehlman dan Sleper 2006; Chahal dan Gosal 2003).
Tabel 1. Metode seleksi untuk tanaman menyerbuk sendiri
No Metode Seleksi Prinsip Seleksi Syarat Seleksi
1. Pedigree Dilaksanakan pada generasi awal
dengan tingkat segregasi tinggi (F2 atau F3), berulang dilakukan terhadap individu terbaik dari famili terbaik sampai tercapai tingkat homozigositas tinggi dan silsilah setiap galur di catat
Karakter kuantitatif memiliki nilai heritabilitas tinggi
2. Bulk Generasi awal ditanam rapat dan
dipanen secara gabungan, tidak dilakukan seleksi pada generasi awal, seleksi baru dilakukan setelah tercapai tingkat homozigositas yang tinggi (F5/F6)
Karakter kuantitatif memiliki nilai heritabilitas rendah sampai sedang
3. Single seed
descent
Generasi awal ditanam hanya satu benih per tanaman, tidak dilakukan seleksi pada generasi awal, penggaluran baru dilakukan setelah mencapai tingkat homozigositas tinggi (F5/F6)
Karakter kuantitatif memiliki nilai heritabilitas rendah sampai sedang
tandem) atau berdasarkan beberapa karakter (seleksi indeks dan independent
culling level) (Roy 2000).
Metode single seed descent telah banyak digunakan, seperti pada tanaman kedelai (Miladinovic et al. 2011) untuk mengetahui metode seleksi yang lebih efisien antara metode bulk, pedigree dan single seed descent dan Janwan et al. (2013) pada tanaman padi menggunakan metode single seed descent untuk mendapatkan galur yang berproduksi tinggi.
Parameter yang harus diperhatikan dalam pelaksanaan seleksi adalah keragaman genetik dan heritabilitas. Efektivitas seleksi untuk memperoleh genotipe unggul ditentukan oleh keragaman genetik pada suatu populasi dan seberapa besar sifat unggul yang diinginkan dapat diturunkan pada generasi selanjutnya (Poehlman dan Sleper 2006). Keragaman suatu populasi dapat diketahui dari keragaman fenotipe dan keragaman genotipenya. Keragaman fenotipe merupakan keragaman yang dapat diukur atau dilihat langsung pada karakter yang diamati. Keragaman genetik tidak dapat dilihat atau diukur secara langsung, melainkan dapat diduga melalui analisis ragam.
Heritabilitas merupakan suatu parameter yang biasa digunakan oleh pemulia tanaman untuk menduga seberapa besar ragam fenotipe yang teramati ditentukan oleh ragam genetik. Heritabilitas yang tinggi menunjukkan bahwa ragam dari sifat-sifat yang diamati akan diwariskan (Kuswantoro et al. 2011). Heritabilitas merupakan salah satu parameter genetik yang digunakan untuk menduga kemajuan dalam perbaikan suatu karakter tanaman. Heritabilitas dapat digolongkan menjadi dua macam, yaitu heritabilitas arti luas dan heritabilitas arti sempit (Fehr 1987).
Heritabilitas arti luas merupakan nisbah total ragam genetik dengan lingkungan, sedangkan heritabilitas arti sempit mempertimbangkan keragaman yang disebabkan oleh peranan gen aditif sebagai bagian dari keragaman genetik total. Secara umum, heritabilitas arti sempit mendapat perhatian khusus karena pewarisan sifat dari tetua kepada keturunannya merupakan pengaruh aditif dari gen sehingga fenotipe tidak tergantung dari adanya interaksi antar alel. Nilai duga heritabilitas memiliki beberapa kegunaan, antara lain untuk mengetahui respon karakter yang diamati terhadap tekanan seleksi sehingga dapat diduga respon seleksinya. Semakin tinggi nilai heritabilitas maka semakin tinggi respon seleksi dan semakin efektif seleksi yang dilakukan (Lalic et al. 2003).
Banyak metode yang dapat digunakan untuk menduga nilai heritabilitas dan komponen ragam. Heritabilitas dapat diduga dengan menggunakan perhitungan ragam turunan, dengan regresi parent-offspring, perhitungan komponen ragam dari analisis ragam dan dengan rancangan hibridisasi (Koumber and El-Gammaal 2012). Nilai heritabilitas tergolong rendah jika kurang dari 20%, cukup tinggi jika 20-50% dan tinggi jika lebih dari 50% (Stanfield 1983). Nilai-nilai tersebut sangat tergantung dari metode dan populasi yang digunakan (Nasir 2005).
Seleksi yang dilakukan terhadap suatu populasi tanaman diharapkan dapat menghasilkan keturunan yang lebih baik. Seleksi diharapkan dapat menghasilkan kemajuan genetik. Secara konsep kemajuan genetik didasarkan pada perubahan dalam rata-rata penampilan yang dicapai suatu populasi dalam setiap siklus seleksi (Baihaki 2000).
heritabilitasnya besar maka kemajuan seleksi yang diperoleh akan semakin baik. Kemajuan seleksi juga dapat diukur menggunakan diferensial seleksi. Diferensial seleksi merupakan komponen persamaan respon seleksi. Diferensial seleksi dugaan merupakan ukuran baku selisih antara rata-rata populasi terseleksi dengan rata-rata populasi awal pada generasi yang sama (Barman dan Borah 2012).
Pola Pewarisan Sifat Karakter Kualitatif dan Kuantitatif
Karakter-karakter yang diekspresikan tanaman dibedakan atas karakter kualitatif dan karakter kuantitatif. Karakter kualitatif seperti warna dan bentuk bunga, bentuk dan warna biji, penampilannya sangat sedikit dipengaruhi oleh faktor lingkungan. Karakter-karakter ini akan mempunyai penampilan yang tetap pada berbagai lingkungan yang berbeda (Stoskopf 1993). Hal ini dapat terjadi karena karakter-karakter ini dikendalikan oleh gen-gen mayor yang mempunyai pengaruh yang sangat besar terhadap penampilan/fenotipe sehingga pengaruh lingkungan terhadap karakter tersebut kecil. Karakter kualitatif memiliki keragaan yang dapat dengan mudah dikelaskan (Singh dan Chaudhary 1979) yaitu menurut Hukum Segregasi, dimana alel-alel dari pasangan gen bersegregasi (berpisah) satu dengan lainnya ke dalam gamet dan Hukum Perpaduan Bebas yaitu pada waktu pembentukan gamet, salah satu pasangan gen berpadu secara bebas dengan pasangan gen lainnya.
Keragaman karakter kualitatif dapat dibedakan berdasarkan aksi gen yang mengendalikan karakter tersebut (Stoskopf 1993; Chahal dan Gosal 2002). Karakter kualitatif dapat dikendalikan oleh satu gen yang mempunyai aksi dominan, over dominan atau kodominan, yang merupakan bentuk interaksi alel dalam lokus/gen yang sama. Dominansi adalah bentuk interaksi antar alel yang menyebabkan alel pasangannya dalam lokus yang sama tertekan ekspresinya. Alel yang terekspresi disebut alel dominan, sedangkan alel yang tertekan ekspresinya disebut alel resesif. Jika dominansi bersifat penuh, maka ekspresi aksi gen dominan akan menyebabkan genotipe dengan alel heterozygot mempunyai fenotipe yang sama dengan bentuk homozigot dominannya. Jika dominansi tidak bersifat penuh, maka genotipe heterozigot akan mempunyai fenotipe yang berada di antara fenotipe genotipe homozigot dominan dengan homozigot resesif. Aksi gen dominan dapat terjadi dalam bentuk overdominansi, yaitu genotipe heterozigot akan mempunyai fenotipe yang melebihi genotipe homozigot dominan. Fenomena ini umumnya terlihat pada karakter yang berhubungan dengan ukuran dan viabilitas biji. Epistasis terjadi jika dua gen berinteraksi mengatur karakter yang sama dan interaksi tersebut melibatkan alel dari lokus yang berbeda. Epistasis dapat menyebabkan timbulnya fenotipe yang berbeda dari fenotipe yang disebabkan oleh interaksi antar alel dalam lokus yang sama, sehingga menyebabkan terjadinya perubahan dalam nisbah fenotipe di generasi berikutnya.
demikian disebut gen minor. Aksi gen minor ditentukan oleh bentuk interaksi yang terjadi baik interaksi antar alel pada lokus yang sama. Untuk karakter kuantitatif maka interaksi antar alel dapat terjadi dalam bentuk interaksi aditif dan dominan maupun interaksi antar alel pada lokus yang berbeda (epistasis). Keragaman fenotipe dari karakter kuantitatif tidak dapat dikelaskan dengan jelas dan cenderung membentuk sebaran yang kontinyu karena dikendalikan oleh banyak gen. Seleksi terhadap karakter kuantitatif tidak didasarkan pada visual tetapi pada hasil analisis statistik.
Beberapa hasil penelitian tentang pola pewarisan sifat toleransi terhadap cekaman aluminium sudah banyak dilaporkan, antara lain pada tanaman gandum (Nava et al. 2006) dan sorghum (Ringo et al. 2010), namun penelitian pola pewarisan sifat toleransi terhadap tanah masam belum banyak dilaporkan. Menurut hasil penelitian Wulandari (2003) pada tanaman padi bahwa pola pewarisan sifat toleransi tanah masam pada karakter panjang akar relatif dikendalikan aksi gen dominan resesif, sedangkan hasil penelitian Wirnas (2007) pada tanaman kedelai toleran naungan bahwa jumlah polong isi, jumlah polong total dan bobot biji per tanaman dikendalikan aksi gen dominan parsial.
Marka Molekuler
Keberhasilan program pemuliaan ditentukan dengan pemilihan metode seleksi dan karakter seleksi yang tepat. Seleksi dapat dilakukan secara langsung atau tidak langsung. Seleksi langsung hanya efisien jika karakter yang ingin diperbaiki memiliki nilai heritabilitas yang tinggi, namun jika karakter yang ingin diperbaiki mempunyai nilai heritabilitas yang rendah maka seleksi tidak langsung menggunakan marka molekuler lebih efisien.
Marka molekuler memiliki kelebihan dibandingkan marka morfologi, diantaranya meningkatkan efisiensi seleksi dalam pemuliaan tanaman dengan cara seleksi tidak langsung terhadap karakter yang dituju dengan menyeleksi sekuen marka yang terdapat pada tanaman, mempercepat deteksi introgesi gen asing atau bagian-bagian kromosom dari spesies liar ke dalam kultivar budidaya melalui silang balik. Selain itu marka molekuler tidak dipengaruhi oleh lingkungan sehingga tidak dipengaruhi oleh kondisi dimana tumbuhan berada dan dapat terdeteksi pada semua tahap perkembangan tanaman (Mohan et al. 1997). Di samping nilai heritabilitas yanng tinggi, keuntungan menggunakan marka molekuler sebagai kriteria seleksi adalah jumlahnya tidak terbatas (Gupta et al.
2002)
Random amplified polymorphic DNA (RAPD) adalah teknik untuk
mengamplifikasi DNA dengan mesin PCR menggunakan primer tunggal berukuran 10 nukleotida. Primer yang digunakan merupakan primer acak yang mengamplifikasi genom target secara acak. Primer acak yang dicampurkan dengan reaksi amplifikasi akan berikatan dengan sekuens komplemen di sepanjang genom target. Selanjutnya sekuens target akan teramplifikasi, yang dapat divisualisasikan pada gel agarose yang diwarnai dengan ethidium bromida (Sharma et al. 2008)
mudah, murah, dan menganalisis dalam tingkat DNA, maka seleksi dini dapat dilakukan terhadap sifat-sifat ketahanan terhadap cekaman biotik dan abiotik.
Tehnik RAPD lebih sederhana dan mempunyai beberapa keuntungan dibandingkan tehnik lain, yaitu tidak memerlukan informasi tentang genom, membutuhkan sampel DNA yang lebih sedikit (10-25 ng), tidak bersifat radioaktif, pelaksanaannya relatif lebih mudah karena tidak perlu pelabelan dengan radioisotop, DNA tidak perlu di potong dengan enzim restriksi dan menghasilkan estimasi yang lebih tinggi untuk kesamaan interspesifik serta jumlah lokus yang diketahui lebih banyak yaitu 1-10. Tehnik RAPD juga mempunyai beberapa keterbatasan, antara lain tidak dapat membedakan individu homozygot dan heterozygot karena bersifat dominan dan kesulitan dalam skoring data. Kelemahan lain dari tehnik RAPD adalah tingkat reprodusibilitasnya sangat rendah sehingga sering memberikan hasil yang tidak konsisten (Dowling et al.
1996; Gupta et al. 2002).
Sejumlah pendekatan berbeda dapat dilakukan untuk meningkatkan keakuratan hasil analisis tehnik RAPD, di antaranya membuat ulangan dalam elektroforesis, membuang semua pita yang tidak dapat diulang dan menggunakan semua pita DNA dengan tingkat kesalahan tertentu (Hallden et al. 1996).
Selain pendekatan di atas untuk mengatasi kelemahan tehnik RAPD dilakukan tehnik marka inter simple seguence repeats (ISSR) menggunakan
simple seguence repeats sebagai primer dengan atau tanpa penambahan sekuen
anchor pada ujung 3‟ atau 5‟. Sekuen anchor berfungsi untuk menyesuaikan annealing primer ke situs target DNA cetakan yang diapit oleh sekuen anchor yang komplementer. Amplifikasi terjadi jika dua mikrosatelit sekuen berulang yang sama, dalam orientasi terbalik, berlokasi cukup dekat satu sama lain sehingga memungkinkan sekuen diantaranya untuk teramplifikasi (Pharmawati, 2009).
Marka ISSR dihasilkan oleh amplifikasi DNA dengan PCR yang menggunakan primer tunggal. Primer disusun dari sekuen mikrosatelit yang biasanya dijaga pada ujung 3‟ atau 5‟ oleh dua atau empat nukleotida. Marka ISSR merupakan metode yang cepat, sederhana dan mempunyai reproduksibilitas tinggi dengan penggunaan primer yang panjang dan kekuatan yang tinggi dicapai dengan suhu annealing. ISSR dapat digunakan untuk tiap spesies karena tidak memerlukan informasi sekuen genom yang diuji (Gurcan et al. 2009).
Marka RAPD dengan alel yang berbeda pada lokus yang sama ditandai dengan ada atau tidaknya pita hasil amplifikasi. Individu tanaman dibedakan berdasarkan ada atau tidak ada pita hasil amplifikasi. Jika muncul pita maka genotipe individu adalah homozigot dominan atau heterozigot. Jika pita tidak muncul untuk lokus yang sama maka individu tanaman adalah homozigot resesif. Marka RAPD merupakan marka dominan sehingga individu homozigot dan heterozigot tidak dapat dibedakan, tetapi dapat membedakan individu homozigot dominan atau heterozigot dengan individu homozigot resesif dalam populasi yang masih bersegregasi. Individu homozigot dan heterozigot akan menghasilkan pita amplifikasi yang sama (Liu 1998).
Bulk segregant analisis (BSA) yang dikembangkan Michelmore et al.
lokus yang dituju secara cepat dan tepat. Metode bulk segregant analisis
menghasilkan marka yang terpaut erat karakter yang diinginkan karena marka yang dihasilkan telah mengalami skrinning sebanyak tiga kali yaitu pada kedua tetua, kelompok toleran dan peka serta masing-masing individu toleran dan peka. Marka metode bulk segregant analisis dapat dipergunakan sebagai marker
assisted selection untuk generasi lanjut.
Pengembangan marka molekuler sebagai alat bantu seleksi dalam pemuliaan tanaman selain metode BSA, juga melalui analisis molekuler dengan pemetaan marka molekuler yang terpaut dengan QTL (quantitative trait loci) yang mengendalikan karakter yang akan diperbaiki. Penggunaan marka molekuler yang terpaut dengan QTL yang mengendalikan karakter yang akan diperbaiki dikenal dengan marker assisted selection (MAS).
Memanfaatkan marka molekuler sebagai MAS dalam program seleksi terhadap karakter yang diinginkan maka marka yang berasosiasi dengan QTL yang mengendalikan karakter yang diinginkan harus diidentifikasi terlebih dahulu melalui analisis dan pemetaan QTL (Azrai et al. 2002; Ruswandi et al. 2002).
Quantitative trait loci (QTL) adalah lokus yang mengendalikan karakter
kuantitatif. Pemetaan QTL merupakan kombinasi antara analisis pautan kualitatif dengan analisis genetika kuantitatif. Pemetaan QTL meliputi konstruksi pemetaan genom dan penelusuran hubungan antara karakter kualitatif dengan marka polimorfik. Hasil pemetaan QTL dapat menyediakan informasi tentang jumlah dan aksi gen yang mengontrol suatu karakter serta lokasinya pada kromosom. Bila memanfaatkan marka molekuler yang terpaut dengan QTL sebagai MAS maka seleksi dapat dilakukan pada generasi awal dan langsung pada individu tanaman sehingga hasil yang diperoleh lebih akurat dan dalam waktu yang lebih cepat (Terry et al 2000; Korzun 2004).
STUDI PEWARISAN SIFAT TOLERANSI KEDELAI
TERHADAP TANAH MASAM
Abstrak
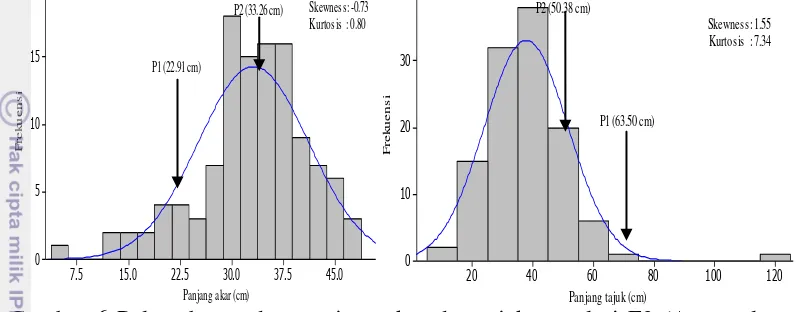
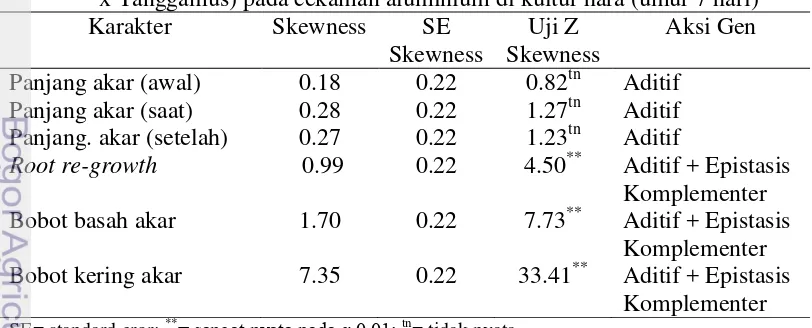
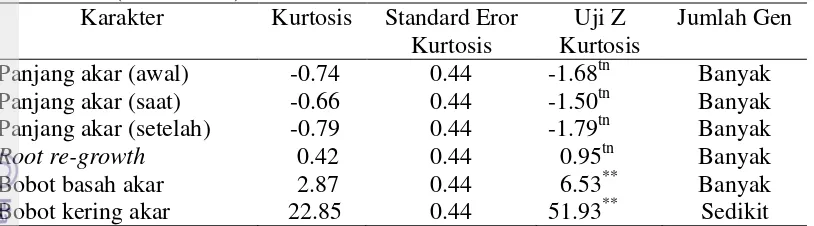
Penelitian ini bertujuan untuk mendapatkan informasi tentang aksi gen dan pewarisan sifat toleransi kedelai terhadap cekaman aluminium di tanah masam dan keracunan Al pada stadia bibit di kultur hara, serta mempelajari aksi gen kemampuan akar kedelai dalam memperbaiki kerusakan akar setelah tercekam Al di larutan hara. Penelitian dilaksanakan di Jasinga, Kabupaten Bogor bulan Mei sampai Agustus 2012 dan di Rumah Kaca University Farm IPB Cikabayan Bogor bulan Maret 2013. Materi yang digunakan adalah Argomulyo (tetua peka), Tanggamus (tetua toleran) dan F2. Hasil penelitian pewarisan sifat toleransi tanah masam menunjukkan bahwa populasi F2 kedelai hasil persilangan Argomulyo dan Tanggamus di tanah masam memiliki nilai tengah lebih tinggi dari nilai tengah kedua tetua untuk beberapa karakter agronomi yang diamati dan terdapat segregan transgesif. Karakter agronomi yang diamati dikendalikan oleh gen aditif dan epistasis komplementer, kecuali karakter tinggi tanaman yang hanya dikendalikan oleh banyak gen dan aksi gen aditif. Nilai heritabilitas untuk karakter agronomi yang diamati tergolong rendah sampai sedang. Percobaan pewarisan sifat toleransi terhadap aluminium di kultur hara, karakter panjang akar dikendalikan oleh banyak gen dengan aksi gen aditif dan epistasis duplikat, sedangkan karakter lainnya dikendalikan oleh aksi gen aditif dan epistasis komplementer. Nilai heritabilitas karakter pertumbuhan yang diamati tergolong sedang sampai tinggi. Sifat root re-growth kedelai pada cekaman aluminium dikendalikan oleh banyak gen dengan aksi gen aditif dan epistasis komplementer tetapi bobot kering akar dikendalikan oleh sedikit gen dengan aksi gen aditif dan epistasis komplementer.
Kata kunci: aksi gen, aditif, epistasis, heritabilitas, nilai tengah, kultur hara, root
regrowth
Abstract
inheritance study of soybean seedlings tolerance to aluminum in nutrient culture showed that the root length character is controlled by many genes with additive gene action and duplicate epistasis, while other character are controlled by additive genes action and complementary epistasis. The heritability estimates for root growth characters were classified as moderate to high. The character of soybean root re-growth under aluminum stress is controlled by many genes with additive gene action and complementary epistasis but root dry weight is controlled by a few genes with additive gene action and complementary epistasis.
Keywords: additive, epistasis, gene action, heritability, mean value, nutrient
culture, root re-growth
Pendahuluan
Umumnya karakter agronomi pada tanaman dikendalikan secara kompleks. Oleh karena itu perlu dilakukan penelitian mengenai kendali genetik terhadap suatu karakter. Sejauh ini belum banyak informasi tentang dasar genetik dan pola pewarisan sifat toleransi kedelai terhadap kandungan aluminium tinggi. Beberapa hasil penelitian tentang pola pewarisan sifat ketahanan tanaman pada kondisi cekaman aluminium di tanah masam telah dilaporkan oleh Wulandari (2003) pada tanaman padi. Hasil penelitiannya menunjukkan bahwa sifat toleransi terhadap Al pada karakter panjang akar relatif dikendalikan aksi gen dominan resesif. Hasil penelitian Hariyanto (2009) pada tanaman padi memperlihatkan bahwa karakter tinggi tanaman, panjang akar dan komponen produksi merupakan karakter poligenik, sedangkan Sa‟diyah et al. (2013) melakukan penelitian pada kondisi optimum terhadap tanaman kedelai, telah melaporkan bahwa tinggi tanaman, bobot 100 biji dan bobot biji per tanaman adalah karakter kuantitatif.
Tehnik hibridisasi atau persilangan buatan dapat menimbulkan keragaman baru melalui rekombinasi yang terbentuk dari alel-alel yang berasal dari tetua-tetua persilangan (Baenziger 2006). Menurut Mahendra (2010) populasi F2 merupakan populasi yang bersegregasi. Tingkat segregasi dan rekombinan yang luas pada generasi F2 ini tergambar melalui sebaran frekuensi genotipenya. Sebaran frekuensi tersebut dapat digunakan sebagai penduga pola pewarisan sifat dan jumlah gen yang terlibat dalam pengendalian suatu sifat dari karakter agronomi tanaman (Christiana 1996).
Pertanaman di lapangan sangat dipengaruhi banyak faktor lingkungan selain cekaman Al, kekurangan unsur hara P dan Mg, kekeringan, penyakit dan hama, sehingga mengakibatkan nilai heritabilitas arti luas tergolong rendah sampai sedang pada percobaan lapangan. Untuk mengatasi permasalahan tersebut maka diperlukan suatu metode yang efisien, cepat dan kondisi lingkungan lebih terkendali dan seragam, terutama pada pengamatan di fase awal pertumbuhan tanaman atau fase kecambah. Metode yang biasa digunakan adalah metode kultur hara (Zhang et al. 2004).
setelah tanaman mendapat perlakuan cekaman Al. Salah satu karakter yang sudah digunakan untuk mengamati sifat toleransi Al pada populasi segregasi adalah karakter root re-growth. Karakter root re-growth ini telah digunakan pada tanaman triticale (Zhang et al. 1999), padi (Roslim et al. 2010) dan rye (Secale
cereale L.) (Miftahudin et al. 2004).
Pembentukan populasi bersegregasi untuk studi pola pewarisan sifat toleransi kedelai terhadap kandungan aluminium tinggi dengan menyilangkan antara varietas kedelai peka tanah masam