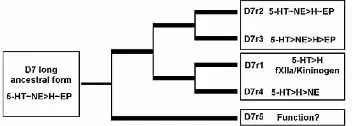
Isolasi Gen Pengkode Arthropod Odorant Binding Protein D7 dari Kelenjar Saliva Vektor Malaria Anopheles sundaicus
Bebas
6
0
0
Teks penuh
Gambar


Dokumen terkait
