Serangan Patogen Dan Gangguan Terhadap Proses
Fisiologis Pohon
Oleh
Y U N A S F I
NIP. 132288490
DEPARTEMEN KEHUTANAN
FAKULTAS PERTANIAN
UNIVERSITAS SUMATERA UTARA
KATA PENGANTAR
Puji dan syukur penulis panjatkan ke hadirat Allah SWT yang telah memberikan rahmat dan berkahnya sehingga penulis dapat menyelesaikan tulisan ini.
T e r j a d i n y a p e n y a k i t p a d a p o h o n h u t a n d i s e b a b k a n o l e h i n t e r a k s i a n t a r a p a t o g e n , i n a n g ( p o h o n ) d a n l i n g k u n g a n . P e n y a k i t p a d a p o h o n h u t a n a k a n me m b e r i k a n d a m p a k y a n g me r u g i k a n b a g i p o h o n , k a r e n a t e r g a n g g u n y a p e r t u m b u h a n d a n p e r k e m b a n g a n p o h o n . P e r k e mb a n g a n p o h o n y a n g t i d a k n o r ma l a k i b a t s e r a n g a n p e n y a k i t , me n y e b a b k a n m e n u r u n n y a k u a l i t a s d a n k u a n t i t a s p r o d u k s i k a y u y a n g d i h a s i l k a n . P a t o g e n s e b a g a i s a t u k o mp o n e n p e n y e b a b p e n y a k i t p a d a p o h o n , d a p a t b e r k o n t r i b u s i t e r h a d a p g a n g g u a n p r o s e s f i s i o l o g i s p a d a p o h o n .
D a l a m t u l i s a n i n i d i u r a i k a n p e n g a r u h s e r a n g a n p a t o g e n t e r h a d a p b e r b a g a i p r o s e s - p r o s e s f i s i o l g i s y a n g t e r j a d i p a d a p o h o n . A k i b a t s e r a n g a n p a t o g e n b a n y a k p r o s e s f i s i o l g i s y a n g t e r g a n g g u p a d a p o h o n d i a n t a r a n y a p r o s e s f o t o s i n t e s i s , r e s p i r a s i d a n l a i n -l a i n . D i h a r a p a k n s e mo g a t u -l i s a n i n i b e r ma n f a a t , d a -l a m r a n g k a p e n g e n d a l i a n p e n y a k i t h u t a n .
P e n u l i s ,
DAFTAR ISI
1.5. Pertumbuhan dan Reproduksi 3
1.6. Pemencaran 3
II. TERGANGGUNYA PROSES FOTOSINTESIS DAN RESPIRASI PADA
POHON YANG TERSERANG PATOGEN 5
2.1. Terganggunya proses fotosintesis pada pohon yang terserang
patogen 5
2.2. Ter ganggunya pr ose s re spira si pa da pohon yang terserang
patogen 6
III HUBUGAN BIOSINTESIS ASAM NUKLEAT, ASAM AMINO DAN PROTEIN ; PERUBAHAN-PERUBAHAN FISIOLOGIS YANG TERJADI PADA POHON YANG SAKIT APABILA METABOLISME N DAN P
NYA TERGANGGU 10
3.1. Perubahan-perubahan fisiologis pada pohon yang sakit apabila metabolisme N dan P nya terganggu 15 IV MEKANISME PENGGUNAAN ENZIM PEKTINASE DAN SELULASE
OLEH PATOGEN DALAM MERUSAK DINDING SEL 16 4.1. Mekanisme penggunaan enzim pektinase 16 4.2. Mekanisme penggunaan enzim selulase 17 4.3. PERAN ENZIM PROKSIDASE DAN POLIFENOLOKSIDASE
PADA INANG SEBAGAI PERTAHANAN TERHADAP
PATOGEN 22
DAFTAR GAMBAR
No. Teks Halaman
1. Komponen penyusun asam nukleat 11 2. Sebagian rantai asam nukleat yang menunjukan ikatan antara
nukleotida-nukleotida penyusunnya.
12 3. Ikatan hidrogen yang berperan dalam pembentukan struktur spiral
ganda molekul DNA, antara pasangan Thimin-Adenin dan Sitosin--Guanin.
13 4. Skema yang mewakili struktur dan komposisi dinding sel 16 5. Perombakan rantai pektin oleh tiga jenis pektinase menjadi
molekul-molekul yang dimodifikasi dan lebih sederhana
17 6. Mekanisme Hidrolisis Selulosa secara Enzimatis (Norkrans, 1967) 29 7. Skema tahap-tahap hidrolisis selulosa secara enzimatis (Sasaki, 1982). 21
8. Bentuk-bentuk akohol lignin 23
I. PENDAHULUAN
Pada suatu penyakit infektif, terjadi keadaan yang sedikit banyaknya dapat
dibedakan dalam suatu suksesi dan membawa pada perkembangan dan
melestarikan keadaan penyakit dan patogen. Kejadian-kejadian utama yang
terjadi dalam satu siklus penyakit adalah inokulasi, penetrasi, pembentukan
infeksi, penyerangan (invasi), pertumbuhan dan reproduksi patogen,
pemencaran patogen.
1.1. Inokulasi
Inokulasi adalah terjadinya kontak antara patogen tumbuhan.
Patogen--patogen yang sampai dan menyebabkan terjadinya kontak dengan
tumbuhan disebut inokulum. Inokulum adalah bagian patogen yang dapat
memulai infeksi. Pada fungi inokulum dapat berupa spora, sklerotium atau
bagian-bagian miselium. Pada bakteri dan virus selalu berupa keseluruhan
individu bakteri dan virus.
1.2. Penetrasi
Patogen mempenetrasi permukaan tumbuhan secara langsung melalui
lubanglubang alami, atau melalui luka. Fungi ada yang dapat melakukan
penetrasi dengan satu cara dan ada yang dua cara. Bakteri umumnya
masuk melalui luka, jarang melalui lubang alami dan tidak pernah secara
langsung. Adapun virus dapat masuk melalui luka yang dibuat vektornya
dan juga melalui luka-luka mekanik yang disebabkan oleh alat-alat
pertanian.
Proses penetrasi ini dapat dilakukan dengan beberapa cara yaitu :
mempenetrasi tumbuhan inangnya dengan menggunakan
apresorium.
b. Penetrasi melalui luka, seperti semua bakteri, sebagian besar
fungi dan beberapa virus.
c. Penetrasi melalui lubang-lubang alami, banyak fungi dan bakteri
masuk ke dalam tumbuhan melalui stomata, hidatoda, nektartoda
dan lentisel.
1.3. Infeksi
Infeksi adalah proses saat patogen melakukan kontak dengan sel atau
jaringan tumbuhan yang rentan dan mendapatkan makanan dan tumbuhan
tersebut. Infeksi yang berhasil akan mengakibatkan timbulnya bagian yang
berubah warna, berubah bentuk, atau nekrosis pada tumbuhan inang yang disebut
gejala, dan ada yang tidak menghasilkan gejala yang disebut laten dan gejala ini
akan kelihatan pada waktu berikutnya di saat kondisi lingkungan lebih
menguntungkan.
Interval waktu antara inokulasi dengan munculnya gejala penyakit
disebut periode inkubasi. Lama periode inkubasi berbagai penyakit
bervariasi, khususnya dengan kombinasi inang-patogen, dengan tingkat
perkembangan inang, dan dengan suhu lingkungan tumbuhan yang terinfeksi.
1.4. Penyerangan (Invasi)
Patogen dengan jenis yang berbeda akan menyerang inang dengan
cara yang berbeda pula. Sebagian besar fungi masuk kedalam bagian
organ-organ tumbuhan (daun, batang, akar) yang diinfeksinya, baik
dengan jalan tumbuh menembus selmiselium, atau dengan jalan tumbuh di
antara sel-sel miselium interseluler.
Bakteri menyerang jaringan tumbuhan secara interseluler, namun demikian
Virus menyerang jaringan tumbuhan dengan berpindah dari sel ke
sel secara intraselular. Banyak infeksi yang disebabkan oleh fungi,
bakteri, virus, nematoda dan tumbuhan tingkat tinggi parasit bersifat lokal yaitu
patogen melibatkan satu sel, beberapa sel atau sebagian kecil sel tumbuhan.
Selain itu ada infeksi alami bersifat sistemik yaitu patogen dan titik awal
infeksi menyebar dan menyerang sebagian besar atau seluruh sel atau jaringan
yang rentan dan seluruh bagian tumbuhan.
1.5. Pertumbuhan dan Reproduksi
Sebagian besar patogen baik yang menghasilkan bercak kecil,
bagian yang terinfeksi lebih meluas atau nekrosis umum yang terdapat
pada tumbuhan, pertumbuhan dan perkembangannya terus berlanjut
terus-menerus di dalam inang yang terinfeksi tanpa henti, sehingga individu patogen
terus menyebar. Sedangkan patogen lain - bakteri, virus - ukurannya tidak banyak
meningkat, karena bentuk dan ukurannya relatif tidak berubah selama masa
hidupnya.
Fungi berkembangbiak dengan spora, yang mungkin terjadi secara
aseksual ataupun secara seksual. Sebagian besar fungi patogenik tumbuhan
menghasilkan miselium hanya dalam tumbuhan yang terinfeksi. Hanya sedikit
fungi yang menghasilkan miselium pada permukaan tubuh inangnya. Bakteri
berkembangbiak dengan cepat di dalam jaringan yang terinfeksi. Di
bawah kondisi makanan dan lingkungan yang optimal bakteri membelah
diri setiap 20 sampai 30 merit.
Virus berkembangbiak di dalam sel-sel inang yang hidup - partikel virus
baru pertama-tama dapat dideteksi beberapa jam setelah infeksi. Setelah
itu partikel-partikel virus akan terakumulasi di dalam sel-sel hidup yang
terinfeksi hingga dapat mencapai 100.000 sampai 10.000.000 partikel virus dalam
1.6. Pemencaran
Hampir semua penyebaran patogen yang bertanggung jawab terhadap ledakan penyakit tumbuhan, dilakukan secara pasif oleh agensia-agensia seperti udara, air, serangga dan hewan-hewan tertentu serta manusia.
a. Pemencaran oleh udara
Aliran udara melepaskan spora dan biji dan sporofor, atau sewaktu
spora dan biji tersebut dikeluarkan secara paksa atau jatuh pada saat
matang, dan tergantung pada turbulensi dan kecepatan aliran udara.
b. Pemencaran oleh air
Air penting bagi penyebaran patogen dalam tiga hal; (1) Bakteri
nematoda dan spora, sklerotium dan bagian miselium fungi yang terdapat
dalam tanah disebarkan oleh air hujan atau air irigasi, (2) Bakteri dan
spora banyak jenis fungi terlarut ke dalam larutan yang dapat melengket
dan penyebarannya tergantung kepada air hujan dan air irigasi, (3)
Butir-butir hujan yang jatuh atau air irigasi yang disemprot dari atas akan
mengambil spora fungi dan bakteri yang terdapat di udara.
c. Pemencaran oleh serangga
Serangga, khususnya aphid dan wereng, merupakan vektor virus
yang paling penting, disamping itu wereng juga vektor utama mikoplasma
II. TERGANGGUNYA PROSES FOTOSINTESIS DAN
RESPIRASI PADA POHON YANG TERSERANG
PATOGEN
2.1. Terganggunya proses fotosintesis pada pohon yang terserang patogen
Fotosistesis merupakan proses satu-satunya sumber semua energi yang
dapat dimanfaatkan oleh sel tumbuhan dan hewan. Proses fotosintesis adalah
proses sintesa senyawa organik dari senyawa-senyawa anorganik biasanya
berupa CO2 dan H20 dengan bantuan energi matahari dari sel-sel yang
mengandung klorofil, dapat digambarkan sebagai berikut :
cahaya
6 CO2 + 6 H2O C6 H12 06 + 6 O2
klorofil
Karena pentingnya peranan fotosintesis dalam kehidupan
tumbuhan, maka apabila ada serangan dari patogen pada tumbuhan
mengakibatkan terganggunya proses fotosintesis dan tanaman akan menjadi
sakit.
Gangguan patogen terhadap prses fotosintesis jelas terlihat dari
klorosis yang terjadi pada banyak tumbuhan yang terinfeksi, dari luka
nekrotik atau nekrotik meluas yang dihasilkan patogen pada bagian
tumbuhan hijau dan menurunnya pertumbuhan dan jumlah buah yang
dihasilkan pada tumbuhan yang terinfeksi.
Perubahan utama akibat terserang patogen pada fotosintesis tumbuhan
adalah terjadi perubahan dan fungsi dan kloroplas yang tidak normal, dimana
terjadinya degenerasi yang dapat menghambat perkembangan pada jaringan
terjadinya penurunan dan aktivitas ribosom. Adapun penyebab dan
ketidak-normalan kloroplas ini diperkirakan karena adanya toksin yang dikeluarkan
oleh patogen, selain itu toksin ini jugs dapat menghambat proses
fotofosforilasi dan menekan sintesis klorofil.
Pada beberapa penyakit yang disebabkan oleh fungi dan bakteri
fotosintesis menurun karena toksin yang dihasilkannya, seperti
tentoksin dan tabtoksin yang dapat menghambat enzim yang terlibat
baik secara langsung maupun secara tidak langsung dalam proses
fotosintesis.
Infeksi patogen yang dapat menyebabkan terjadinya perubahan
pada lubang stomata yang dapat mempengaruhi laju asimilasi, karena
terhambatnya laju aliran CO2. Adanya perubahan dalam fiksasi CO2
akan menyebabkan terjadinya perubahan dalam aktivitas enzim-enzim
yang berperanan dalam proses fotosintesis dan menyebabkan terjadinya
perubahan dalam metabolisme akumulasi asam amino dan asam
organik dalam pelepasan gula dan gula posfat.
Pada bercak daun, hawar, dan berbagai jenis penyakit lain yang
menyebabkan kerusakan jaringan daun atau defoliasi (pengguguran
daun) maka proses fotosintesis akan menurun, karena permukaan yang
berfotosintesis pada tumbuhan menjadi berkurang.
2.2. Terganggunya proses respirasi pada pohon yang terserang patogen
Respirasi (pernafasan) adalah suatu proses dimana sel melalui
oksidasi (pembakaran) yang dikendalikan secara enzimatik terhadap
karbohidrat dan asam lemak berenergi tinggi, membebaskan energi dalam
bentuk yang dapat digunakan untuk berlangsungnya berbagai proses
langkah yaitu :
1. Perombakan glukosa menjadi pirufat melalui reaksi glikolisis
atau melalui reaksi yang lebih pendek, yaitu rangkaian reaksi
pentosa (pentosa pathway).
2. Perombakan pirufat yang menghasilkan CO2 dan air melalui reaksi yang
dikenal dengan siklus Kreb, yaitu diiringi dengan oksidasi
akhir (terminal oxidation).
Di bawah keadaan normal (aerobik), yaitu apabila terdapat oksigen,
kedua langkah tersebut dapat berlangsung, dan satu molekul glukosa
menghasilkan hasil akhir enam molekul CO2 dan enam molekul air dapat
digambarkan sebagai berikut :
C6 H12 06 + 6 02 6 CO2 + 6 H2 0
Langkah pertama respirasi memberikan dua molekul ATP per
mol glukosa, dan langkah kedua memberikan sisanya. Akan tetapi,
di bawah kondisi anaerob (tanpa oksigen) pirufat tidak dapat
dioksidasi, tetapi mengalami fermentasi dan menghasilkan asam
laktat atau alkohol. Penggandaan oksidasi glukosa dengan
penambahan fosfat ke ADP menghasilkan ATP disebut posforilasi
oksidatif (oxidative phosphorylation). Aktivitas sel yang
membutuhkan energi yang tersimpan pada ATP, secara simultan
merombak ATP menjadi ADP dan posfat arorganik. Selanjutnya
terdapatnya ADP dan posfat di dalam sel, akan merangsang laju
respirasi. Jumlah ADP dan posfat di dalam sel ditentukan oleh laju
penggunaan energi, selanjutnya laju tersebut menentukan laju
respirasi dalam jaringan tumbuhan.
tumbuhan untuk semua jenis kerja seluler, seperti penumpukan dan mobilisasi senyawasenyawa, sintesis protein, mengaktifkan enzim pertumbuhan dan pembelahan sel, reaksi-reaksi pertahanan dan proses-proses inang lainnya.
Kekompleskan respirasi, jumlah enzim yang terlibat dalam respirasi, kejadiannya dalam setiap sel, dan pengaruhnya yang luas terhadap fungsi dan keberadaan sel maka respirasi merupakan salah satu fungsi pertama yang dipengaruhi di saat tumbuhan diinfeksi patogen. Pada saat tumbuhan diinfeksi patogen, umumnya laju respirasi meningkat, berarti jaringan terserang patogen menggunakan cadangan karbohidratnya lebih cepat dibanding dengan jaringan yang sehat.
Infeksi tumbuhan oleh fungi, bakteri dan virus umumnya
menunjukan gejala yang sama yaitu meningkatnya laju respirasi, yang
dapat dilihat sebagai fenomena yang umum dalam fisiologi penyakit
tanaman. Tapi kondisi ini tidak spesifik disebabkan oleh petogen saja,
tapi juga disebabkan oleh beberapa bahan kimia dan gangguan
mekanik.
Respirasi meningkat lebih cepat pada infeksi yang terjadi varitas
tahan, yang dibutuhkan dan digunakan untuk produksi atau mobilitas
mekanisme pertahanan sel yang lebih cepat. Hal ini terjadi karena
meningkatnya konsumsi 02 yang ditunjukan oleh adanya gejala
nekrotik baik itu disebabkan oleh virus, fungsi atau bakteri.
Selanjutnya pada fase-fase perkembangan penyakit pada tanaman
resisten peningkatan respirasi akan lebih cepat dan akan berkurang
secara perlahan-lahan.
Pada penghilangan effek Pasteur, jaringan tanaman yang diinfeksi
oleh jamur dan bakteri, menunjukan penurunan efisiensi respirasi, hal
ini terjadi karena meningkatnya penyerapan O2. Di sini dapat
dijelaskan bahwa peningkatan respirasi akan mempengaruhi proses
Peningkatan respirasi kadang-kadang juga diikuti oleh
peningkatan fermentasi secara luar biasa dibanding dengan yang
terjadi pada tumbuhan sehat, mungkin sebagai akibat
meningkatnya kebutuhan energi pada tumbuhan yang sakit karena
pada keadaan respirasi aerob yang normal tidak dapat menyediakan
energi yang mencukupi.
Ada beberapa sebab peningkatan respirasi pada tumbuhan yang
terserang patogen antara lain :
1. Tidak berpasangannya fosforilasi oksidatif
Pada kondisi ini, energi (ATP) yang tidak dapat digunakan,
dihasilkan melalui respirasi yang normal meskipun menggunakan
ATP yang ada dan akumulasi ADP, yang merangsang respirasi.
2. Sebagai akibat peningkatan metabolisme
Pada banyak penyakit tumbuhan, pertama-tama tumbuhan
di-rangsang, aliran protoplasmik meningkat, dan bahan-bahan
disintesis, ditranslokasikan, dan diakumulasikan pada bagian yang
sakit. Energi yang dibutuhkan untuk melaksanakan
aktivitas-aktivitas tersebut berasal dan ATP yang dihasilkan melalui
respirasi. Jika lebih banyak ATP yang digunakan maka lebih
banyak pula ADP yang dihasilkan dan selanjutnya merangsang
respirasi.
Pada proses respirasi juga dapat mengakibatkan terjadinya
perubahan katabolisme glukosa. Perubahan pada lintasan
Embden-Meyerhof-Parnas terjadi perubahan lintasan pentosa posfat pada
jaringan yang diparasit oleh fungi, virus dan bakteri. Perubahan ini
adalah tidak spesifik tergantung pada tipe luka, umur, stress yang
III. HUBUGAN BIOSINTESIS ASAM NUKLEAT, ASAM
AMINO DAN PROTEIN ; PERUBAHAN-PERUBAHAN
FISIOLOGIS YANG TERJADI PADA POHON YANG
SAKIT APABILA METABOLISME N DAN P NYA
TERGANGGU
Asam nukleat terdiri atas dua jenis, yakni asam ribonukleat
(RNA) dan asam deoksiribonukleat (DAN). Masing-masing tersusun
dan molekul yang disebut nukleotida. Nukleotida terbentuk dan asam
fosfat, gula pentosa, dan senyawa basa purin (adenin dan guanin) atau basa
pirimidin (timin dan sitosin).
Nukleotida RNA mengandung gula ribosa, sedangkan
nukleotida DNA mengandung gula deoksiribosa yang memiliki kurang
satu atom oksigen dibanding ribosa. RNA dijumpai pada seluruh bagian sel,
dimana RNA berperan dalam sintesis protein, sedangkan DNA hanya dijumpai
pada inti sel. DNA merupakan bagian utama kromosom.
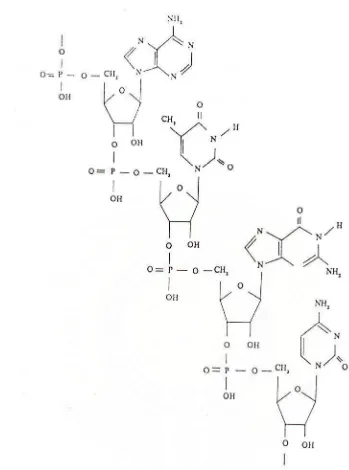
Pemecahan asam nukleat menjadi komponen penyusunnya dapat dilihat
pada Gambar 1, sedangkan bentuk ikatan dalam penggabungan nukleotida dapat
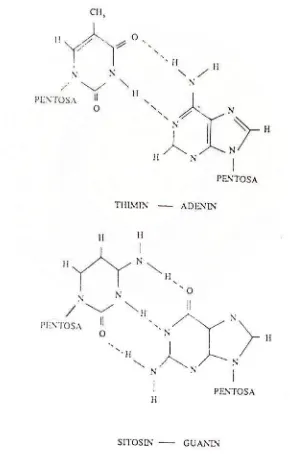
dilihat pada Gambar 2. DNA umumnya berbentuk spiral ganda (Double
helix). Bentuk spiral ganda ini terjadi karena adanya ikatan hidrogen antara atom
H dan 0 atau N dan basa purin dan pirimidin. Pasangan antara basa purin dan
pirimidin yang saling tank menarik oleh ikatan hidrogen dalam struktur
spiral ganda adalah sangat spesifik, dimana adenin selalu berpasangan dengan
timin, sedangkan guanin selalu berpasangan dengan sitosin dapat dilihat pada
Gambar 1. Komponen penyusun asam nukleat
Penyusun utama protein adalah carbon, hidrogen, oksigen, nitrogen dan
sulfur. Adapun monomer dari protein adalah asam amino, yang mana suatu
molekul asam amino terdiri atas suatu gugusan karboksil (-COOH) dan
gugusan amino (-NH2 ).
Sintesis protein merupakan proses perangkaian asam-asam amino
sehingga membentuk suatu rantai yang panjang. Rantai asam amino ini disebut
polipeptida. Molekul protein dapat terdiri atas satu atau lebih rantai polipeptida
Gambar 2. Sebagian rantai asam nukleat yang menunjukan ikatan antara nukleotida-nukleotida penyusunnya.
Pada DNA, selalu terdapat molekul adenin (kelompok nukleotida
purin) sebanyak molekul timin (kelompok pirimidin) dan molekul guanin
tersebut bersambungan membentuk rantai panjang dan setiap molekul
DNA terdiri atas dua rantai yang memilin bersama-sama bagaikan dua
pegangan tangga yang berbentuk spiral, sehingga membentuk heliks
rangkap. Struktur nukleotida sedemikian rupa sehingga adenin pada rantai
yang satu selalu berpasangan dengan timin pada rantai kedua yang
berhadapan dan guanin selalu berpasangan dengan sitosin.
spiral ganda molekul DNA, antara pasangan Thimin-Adenin dan Sitosin-Guanin.
Urutan nukleotida pada DNA menentukan urutan komplementer
nukleotida pada molekul RNA yang dapat dilihat pada proses transkripsi.
Hanya satu dari kedua unting DNA pada gen yang membawa kendali
tersandi untuk mensintesis protein kadang disebut unting bermakna. Pada
transkripsi, kedua rantai DNA terberai dan terpisah : unting bermakna
tersebut berfungsi sebagai tempat untuk sintesis RNA.
Pada sintesis protein, urutan nukleotida yang terdiri dari empat
macam nukleotida di m RNA melaksanakan translasi untuk urutan asam
amino protein. Pada tahap pertama, asam amino berinteraksi dengan ATP
dalam reaksi yang dikatalisatori enzim yang melepaskan pirafosfat dari
ATP dan menggabungkan asam amino tersebut dengan AMP. Pada tahap kedua,
asam amino dipindahkan dari AMP ke molekul RNA pemindah (t RNA) yang
tepat, membentuk komplek aminoasil-t RNA, yang terlepas dari
pembentukannya. Sebagian besar energi awal ATP dipindahkan ke komplek
aminoasil-t RNA jadi ini merupakan pengaktifan asam amino.
Bagian dari molekul t RNA yang terdiri dari tiga nukleotida
membentuk antikodon. Urutan ini terletak pada molekul t RNA, sehingga
terbentuk ikatan komplementer dengan urutan tiga nukleotida yang disebut
kodon pada unting m RNA. Pengaturan posisi tersebut membuat molekul t
RNA, yang masing-masing membawa asam amino khasnya, dapat sebaris
dengan unting m RNA, sehingga asam amino akan berada pada urutan yang
tepat.
Nitrogen merupakan penyusun asam amino, amida, purin dan protein serta
nukleoprotein. Enzim mengandung molekul protein yang berantai panjang
dan kompleks serta kelompok reaktif yang bukan protein yang umumnya suatu
memiliki N dalam cincinnya seperti pada triptopan. Nitrogen juga merupakan
bahan penyusun dari sekumpulan senyawa yang disebut alkaloid dimana
senyawa ini diperkirakan sebagai senyawa cadangan N.
Fosfor merupakan komponen struktural dan sejumlah senyawa
penting, molekul pentransfer energi ADP dan ATP, NAD, NADPH, dan senyawa
sistem informasi genetik DNA dan RNA. Fosfor juga merupakan bahan
penyusun fosfolipid seperti lesitin dan kolin, yang memegang peranan penting
dalam integritas membran.
3.1. Perubahan-perubahan fisiologis pada pohon yang sakit apabila metabolisme N dan P nya terganggu
Perubahan-perubahan fisiologis pada pohon apabila metabolisme N
terganggu akan menyebabkan terjadinya defisiensi N yang dapat membatasi
pembesaran sel dan pembelahan sel, dimana pertumbuhan kerdil, dan kuning,
terutama di bagianbagian tanaman yang lebih tua. Gangguan pertumbuhan ini
dapat menyebabkan penimbunan gula dan menyebabkan jaringan basal
berubah warna menjadi ungu karena pembentukan antosianin. Akibat adanya
redistribusi apabila pengambilan N terbatas adalah terjadinya kebakaran
(menguning dan menua) daun-daun sebelah bawah.
Perubahan-perubahan fisiologis pada pohon apabila metabolisme P
terganggu dalam hal ini juga menyebabkan terjadinya defisiensi P yang
akan berpengaruh kepada pertumbuhan tanaman seperti daun berwarna
hijau gelap atau hijau kebiru-biruan. Tanaman kerdil, jumlah dan panjang
akan berkurang, terjadi penimbunan gula yang ditunjukan dalam bentuk
pigmentasi antosianin pada bagian dasar batang dan urat daun. Bila asam
nukleat tidak terbentuk dengan baik akan menghambat pembelahan sel
karena DNA-nya terganggu, menghambat pembentukan jaringan baru,
menghambat pembentukan organ dan secara keseluruhan dapat
IV. MEKANISME PENGGUNAAN ENZIM PEKTINASE DAN
SELULASE OLEH PATOGEN DALAM MERUSAK
DINDING SEL
4.1. Mekanisme penggunaan enzim pektinase
Zat pektik merupakan komponen penyusun utama lamela tengah,
yaitu semen interseluler yang menempatkan sel-sel pada jaringan
tumbuhan (Gambar 4)
Gambar 4. Skema yang mewakili struktur dan komposisi dinding sel
Zat pektik adalah polisakarida yang terdiri dari molekul
galakturonan yang diselingi dengan sejumlah kecil molekul
rhamnosa dan rantai galakturonan samping sedikit dan beberapa gula
lainnya. Beberapa enzim telah diketahui sebagai pektinase atau enzim
pektolitik yang menguraikan zat pektik. Zat pektik adalah polisakarida yang
terdiri dari molekul galakturonan yang diselingi dengan sejumlah
kecil molekul rhamnosa dan rantai galakturonan samping sedikit
dan beberapa gula lainnya. Beberapa enzim telah diketahui sebagai
pektinase atau enzim pektolitik yang menguraikan zat pektik.
rantai pektin dan tidak mempunyai pengaruh terhadap panjang rantai
pektin, tetapi e nzi m ter se but me rubah daya lar ut pekt in dan
me mpe ngar uhi l aju penyerangan yang disebabkan oleh
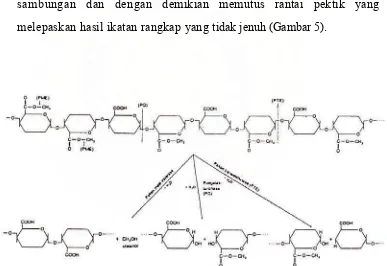
chain-splitting pectinase (enzim yang dapat memotong rantai pektik).
Chain-splitting pectinase memotong rantai pektin dan melepaskan
bagian rantai yang lebih pendek yang mengandung satu atau beberapa
molekul galakturonan. Beberapa chain-splitting pectinase
(poligalakturonase) memotong rantai pektik dengan menambah satu
molekul air dan memutuskan (menghidrolisis) sambungan antara dua
molekul galakturonan enzim yang lain (pektin liase atau transeliminase)
memotong rantai tersebut dengan membuang satu molekul air dari
sambungan dan dengan demikian memutus rantai pektik yang
melepaskan hasil ikatan rangkap yang tidak jenuh (Gambar 5).
Gambar 5. Perombakan rantai pektin oleh tiga jenis pektinase menjadi molekul-molekul yang dimodifikasi dan lebih sederhana
Masing-masing enzim tersebut terdapat dalam bentuk yang
dan melepaskan rantai yang lebih pendek atau hanya dapat
memutuskan hubungan ujung rantai (ekso-pektinase) dan
melepaskan unit galakturonan tunggal.
Patogen menghasilkan enzim pektolitik dalam jumlah yang
paling rendah pada tempat yang terdapat pektin, dan dan pektin tersebut
dilepaskan sejumlah kecil monomer atau oligomer galakturonan.
Molekul-molekul tersebut apabila diserap oleh patogen, berperan sebagai
pengimbas (inducer) untuk meningkatkan sintesis dan melepaskan
enzim-enzim pektolitik (autocatalytic induction), yang selanjutnya
meningkatkan monomer galakturonan. Monomer galakturonan
dengan mudah diasimilasi oleh patogen, tetapi pada konsentrasi yang
lebih tinggi molekul galakturonan menekan enzim yang sama (catabolite
respession).
Enzim-enzim yang merombak pektin terlibat dalam menyebabkan banyak penyakit, terutama penyakit-penyakit penyebab jaringan busuk lunak. Perombakan pektin menghasilkan kumpulan cairan zat pektik yang menyatukan sel-sel tumbuhan bersamaan dengan melemahnya dinding sel, mendorong maserasi jaringan (jaringan tumbuhan menjadi lunak dan longgar serta terpisahnya sel-sel, dan akhirnya mati). Melunaknya dinding sel dan terjadinya maserasi jaringan tidak diragukan lagi memberi peluang terjadinya penyerangan secara inter atau intra seluler oleh patogen. Enzim pektik juga menyediakan makanan bagi patogen pada jaringan yang
terinfeksi.
4.2. Mekanisme penggunaan enzim selulase
Selulosa terdapat pada semua tumbuhan tingkat tinggi sebagai
zat rangka (skeletal) dinding sel dalam bentuk mikrofibril.
p-D-glukopiranosa yang terikat satu sama lain dengan ikatan-ikatan
glikosida (1 —> 4). Dalam bantuk kayu dan kapas selulosa mengandung
nilai makanan sebanyak sukrosa, namun selulosa tidak dapat dicerna
oleh manusia karena cairan tubuh hanya dapat menghidrolisis
ikatan tipe a. bukan Namun binatang-binatang tertentu (pemamah
biak) mampu memanfaatkan selulosa sebagai makanannya karena
di dalam saluran pencernaannya terdapat koloni mikroorganisme
yang menghasilkan enzim yang dikenal dengan nama selulase.
Perombakan selulosa secara enzimatik menghasilkan hasil akhir
berupa molekul-molekul glokosa. Glukosa tersebut dihasilkan
dengan serangkaian reaksi enzimatik yang dilakukan oleh sejumlah
selulase dan enzim-enzim lain.
Selulase dihasilkan secara ekstraseluler oleh berbagai cendawan dan
bakteri. Kemampuan masing-masing mikroba untuk menghasilkan selulase
tidak sama. Ada yang mampu menghasilkan enzim dalam jumlah besar
dengan semua enzim penyusun yang lengkap, sebaliknya ada mikroba yang
hanya mampu menghasilkan enzim dalam jumlah sedikit dan tidak lengkap
enzim penyusunnya (Enari, 1983 dalam Ahmad, 1992).
Selulase merupakan enzim yang komplek yang terdiri dan
beberapa enzim yang bekerja secara bertahap atau bersama-sama
menguraikan selulosa menjadi glukosa (Norkrans, 1967 dalam
Ahmad, 1992). Enari (1983 diacu oleh Ahmad 1992) terdapat empat
kelompok enzim utama sebagai komponen penyusun selulase
berdasarkan spesifikasi substrat masing-masing enzim yaitu :
a. Endo- -1.4-glukanase (3-1-4-glukan-4-glukanohidrolase,
EC.3.2.1.4), menghidrolisis ikatan glikosida -1.4, secara acak.
Enzim ini tidak menyerang selabiosa tetapi menghidrolisis
dan selulosa yang telah disubstitusi seperti CMC dan HES
(Hydroxy Ethyl Cellulose).
b. -1.4-D-glukan selobichidrolase (EC.3.2.1.91), menyerang ujung
rantai selulose non pereduksi dan menghasilkan selobiosa. Enzim
ini dapat menyerang selodekstrin tetapi tidak menyerang
selulosa yang telah disubstitusi serta tidak dapat menghidrolisis
selubiosa.
c. -1.4-D-glukan glukohidrolase (EC.3.2.1.74), menyerang ujung
rantai selulosa non pereduksi dan menghasilkan glukosa. Enzim
ini menyerang selulosa yang telah dilonggarkan dengan asam fosfat,
selo-oligosakarida dan CMC.
d. -1.4-D-glukosidase ( -1.4-D-glukosida glukohidrolase, EC.3.2.1.21),
menghidrolisis selobiosa dan Selo-oligosakarida rantai pendek serta
menghasilkan glukosa. Enzim ini tidak menyerang selulosa
atau selodekstrin.
Selulase dalam melakukan hidrolisis berlangsung dalam dua tahap
proses, yaitu aktifasi sebagai tahap awal yang diikuti dengan reaksi hidrolisis
(Norkans, 1967). Adapun mekanismenya dapat dilihat pada Gambar 6.
C1 Cx -glukosidase s e l u l o s a s e l u l o s a s e l o b i o s a g l u k o s a
r e a k t i f
Gambar 6. Mekanisme Hidrolisis Selulosa secara Enzimatis (Norkrans, 1967)
Secara rinci konsep mekanisme hidrolisis selulosa secara enzimatis
dapat dilihat pada Gambar 7.
Endoglukonase (EG=Cx ) menyerang daerah "amorfous" serat selulosa,
kedua enzim tersebut bekerja bahu membahu membebaskan selobiosa dari
serat selulosa. Baik selobiohidrolase maupun endoglukanase tidak mampu
memecah selobiosa, sehingga diperlukan enzim lain yaitu -glukosidase yang
menguraikan selobiosa menjadi glukosa.
Gambar 7. Skema tahap-tahap hidrolisis selulosa secara enzimatis (Sasaki, 1982).
Enzim selulotik yang disekresikan oleh patogen memainkan peranan
penting dalam melunakan dan menguraikan bahan penyusun dinding sel.
Enzim-enzim tersebut memudahkan penetrasi dan penyebaran patogen di dalam
inang dan menyebabkan pecah (kolapse) dan terurainya struktur seluler,
mungkin berpartisipasi secara tidak langsung dalam perkembangan penyakit
dengan melepaskan, dari rantai selulosa, gula yang dapat larut, yang berperan
sebagai makanan patogen.
4.3. PERAN ENZIM PROKSIDASE DAN POLIFENOLOKSIDASE PADA INANG SEBAGAI PERTAHANAN TERHADAP PATOGEN
Aktivitas enzim phenol-oxidazing (enzim pengoksidasi-fenol)
(polifenoloksidase) lebih tinggi pada jaringan varitas tahan yang terinfeksi
dibanding dengan jaringan varietas rentan yang terfinfeksi. Peranan
aktivitas enzim polifenoloksidase dalam ketahanan penyakit mungkin
berasal dan sifatnya yang dapat mengoksidasi senyawa fenol menjadi
kinon, yang sering lebih beracun bagi mikroorganisme dibanding
dengan fenolnya sendiri. Maka patut diasumsikan bahwa peningkatan
aktivitas polifenol oksidase akan menghasilkan produk toksin yang lebih
tinggi dari basil oksidasi dan oleh karena itu menghasilkan tingkat
ketahanan lebih tinggi terhadap infeksi.
Enzim peroksidase, tidak hanya mengoksidasi fenolik tetapi juga
meningkatkan laju polimerisasi senyawa-senyawa fenolik menjadi
senyawa-senyawa seperti lignin, yang terdeposit dalam dinding sel dan
papila yang selanjutnya menggangu pertumbuhan dan perkembangan
patogen . Secara umum aktivitas peroksidase pada ekstrak daun yang
diinfeksi adanya peningkatan aktivitas dan konsentrasi dan enzim pada
jaringan yang tidak terinfeksi. Peningkatan ini dipengaruhi oleh
Disini diperkirakan adanya isozim barn sebagai akibat infeksi virus
pada sitoplasma. Peroksidase berperanan dalam degradasi H202
yang berhubungan dengan bertambahnya respirasi pada produksi
etilen.
Peroksidase keberadaannya berkaitan dengan lignin, pada umumnya
lignin engandung tiga alkohol aromatik yaitu alkohol koniferil, yang
banyak terdapat pada ka y u l u n a k , a l k o h o l s i n a p i l , d a n a l k o h o l p
-k u ma r i n , d a p a t d i l i h a t p a d a Gambar 8. Beberapa -kemung-kinan cara
alkohol tersebut berhubungan dalam lignin disajikan pada Gambar 9.
Semua alkohol aromatik dalam lignin berasal dari lintasan asam
sikimat. Fenilalanin diubah menjadi asam aromatik seperti kumarat dan
ferulat, kemudian diubah menjadi ester CoA. Ester ini direduksi
menjadi alkohol aromatik oleh NADPH, dan alkohol tersebut
kemudian dipolimerisasi menjadi lignin oleh mekanisme radikal
bebas. Enzim mengandung besi yang dinamakan peroksidase
mengkatalis dua reaksi terpisah yang kemudian menyebabkan
polimerisasi.
Peroksidase terdapat dalam bentuk isozim, sedikit diantaranya
terdapat di dinding sel. Isozim mula-mula berfungsi dengan membentuk
H202 dari NADH dan 02. Kemudian, isozim mengambil atom hidrogen
masing-masing satu dari kedua alkohol aromatik, dan menggabungkan
kedua atom hidrogen itu dengan satu H202 untuk melepaskan dua molekul
H20 sebagai produk samping.
PUSTAKA ACUAN
Achmad. 1992. Cendawan Selulolitik. Kursus Singkat Pemanfaatan Limbah Lignoselulolitik untuk Media Semai Tanaman Kehutanan. Pusat Antar Universitas Institut Pertanian Bogor. Bogor.
Agrios. 1996. Ilmu Penyakit Tumbuhan (Terjemahan Munzir Busnia). Gadjah Mada University Press. Yogyakarta.
Bidwek. R.G.S. 1979. Plant Physiology. Macmillan Publishing Co. Inc. New York. Coller Macmillan Publisher London. London.
Dwijoseputro, D. 1980. Pengantar Fisiologi Tumbuhan. PT. Gramedia Jakarta.
Friend. J, D.R. Threlfall. 1976. Bichemical Aspects of Plant-Parasite Relationships. Department of Plant Biology. University of Hull, England. Academic Press London. New York. San Francisco.
Gardner, F.P., R.B. Pearce, R.L. Mitchell. 1991. Fisiologi Tanaman Budidaya (Terjemahan Herawati Susilo). Universitas Indonesia. Jakarta.
Goodman, R.N., Z. Kiraly, K.R. Wood. 1989. The Biochemistry and Phyisiology of Plant Disease. University of Missouri Press. Columbia.
Lakitan, B. 1993 Dasar-dasar Fisiologi Tumbuhan. PT. Raja Grafindo Persada Jakarta. Jakarta.