i
SPORULASI DAN VIABILITAS KONIDIA CENDAWAN
ENTOMOPATOGEN
Beauveria bassiana
(Balsamo) Vuillemin DI
BERBAGAI MEDIA TUMBUH
SUSILAWATI
DEPARTEMEN PROTEKSI TANAMAN FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR BOGOR
iii
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Sporulasi dan Viabilitas Konidia Cendawan Entomopatogen Beauveria bassiana (Balsamo) Vuillemin di Berbagai Media Tumbuh adalah benar karya saya dengan arahan dari dosen pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
v
ABSTRAK
SUSILAWATI. Sporulasi dan Viabilitas Konidia Cendawan Entomopatogen Beauveria bassiana (Balsamo) Vuillemin di Berbagai Media Tumbuh. Dibimbing oleh TEGUH SANTOSO.
Cendawan Beauveria bassiana merupakan entomopatogen yang secara luas digunakan dalam pengendalian beberapa jenis hama tanaman. Cendawan ini diketahui menginfeksi walang sangit Leptocorisa oratorius dan penggerek batang jagung Ostrinia furnacalis. Penelitian ini bertujuan mempelajari sporulasi dan perkecambahan konidia B. bassiana di media alternatif dan inang alami. Isolat B. bassiana yang berasal dari walang sangit dibiakan di media beras, beras + jagung, PDA (Potato Dextrose Agar), larva O. furnacalis dan imago L. oratorius. Kerapatan konidia dihitung pada 21 dan 33 HSI (hari setelah inokulasi). Daya kecambah dari biakan yang sama diamati dalam PDA cair dan PDA tipis. Hasil penelitian menunjukkan bahwa kerapatan konidia cendawan berumur 21 HSI pada media beras + jagung dan media beras berturut-turut mencapai 1.82x1010 dan 1.65x1010 per 50 g media. Sedangkan pada umur 33 HSI berturut-turut menghasilkan kerapatan 9.96x109 dan 5.80x1010 per 50 g media. Pembentukan konidia meningkat sampai 33 HSI. Peningkatan jumlah konidia juga terjadi pada cendawan yang ditumbuhkan di inang alami O. furnacalis dan L. oratorius: berturut-turut dari 5.38x108 dan 1.68x108 mencapai 9.73x108 dan 4.64x108 konidia/bangkai. Kerapatan konidia di media PDA umur 33 HSI (2.02x108/cawan) tidak berbeda nyata dengan umur 21 HSI (2.93x108/cawan). Daya kecambah konidia umur 33 HSI yang diinkubasi pada media PDA cair selama 24 JSIn (jam setelah inkubasi) lebih tinggi dari konidia umur 21 HSI, kecuali pada media beras + jagung. Inkubasi selama 24 JSIn pada PDA tipis menunjukkan persentase daya kecambah yang tinggi, mencapai 97% (O. furnacalis); PDA tipis menghasilkan daya kecambah yang lebih tinggi dibandingkan PDA cair. Pada kedua umur biakan baik 21 maupun 33 HSI, daya kecambah konidia dari inang alami (O. furnacalis dan L. oratorius) secara signifikan lebih tinggi dari media alternatif (beras dan beras + jagung).
vii
ABSTRACT
SUSILAWATI. Conidial Sporulation and Viability of Entomopathogenic Fungi Beauveria bassiana (Balsamo) Vuillemin on Different Media. Supervised by TEGUH SANTOSO.
Entomopathogenic fungus B. bassiana is widely used in biological control of various plant pests. Naturally this fungus known to infect rice bug Leptocorisa oratorius and Asiatic corn borer Ostrinia furnacalis. This research aims to study the sporulation and germination of B. bassiana conidia on alternative media and on insect natural host. B. bassiana originated from rice bug was cultured on rice, rice + corn, PDA (Potato Dextrose Agar), O. furnacalis larvae and L. oratorius adult. Density of conidia was calculated at 21 and 33 days after culture. The conidial viability of same culture was also observed in liquid PDA and in thin PDA. The result showed that the density of conidia at age 21 days on rice + corn and rice media reached 1.82x1010 and 1.65x1010 per 50 g media. At age 33 days, the obtained density were 9.96x109 and 5.80x1010 per 50 g on rice + corn and rice media, respectively. We noted the increase of conidiogenesis until 33 days. The increase was also noted when fungi were grown on natural host O. furnacalis and L. oratorius: from 5.38x108 and 1.68x108 reached 9.73x108 and 4.64x108 conidia/cadaver respectively. However, on PDA the conidial density at 33 days (2.02x108/dish) did not differ with that at 21 days (2.93x108/dish). When incubated during 24 hours in liquid PDA, generally the germination of conidia aged 33 days were higher than that of 21 days, excepted on rice + corn treatment. Incubation during 24 hours in thin PDA showed the high percentage of conidial germination, reached 97% (O. furnacalis); thin PDA yielded higher germination than liquid PDA. Both at age 21 and 33 days, conidial germination from natural host (O. furnacalis and L. oratorius) was significantly higher than from alternative media (rice and rice + corn).
ix
©Hak Cipta milik IPB, tahun 2015 Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
xi
SPORULASI DAN VIABILITAS KONIDIA CENDAWAN
ENTOMOPATOGEN
Beauveria bassiana
(Balsamo) Vuillemin DI
BERBAGAI MEDIA TUMBUH
SUSILAWATI
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian
pada
Departemen Proteksi Tanaman
DEPARTEMEN PROTEKSI TANAMAN FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR BOGOR
xv
PRAKATA
Alhamdulillahirobbilalamin, puji dan syukur penulis panjatkan ke hadirat Alloh SWT yang telah memberikan nikmat dan rahmatnya sehingga penulis dapat menyelesaikan tugas akhir yang berjudul “Sporulasi dan Viabilitas Konidia Cendawan Entomopatogen Beauveria bassiana (Balsamo) Vuillemin di Berbagai Media Tumbuh”, sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian pada Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor.
Terima kasih penulis ucapkan kepada Dr. Ir. Teguh Santoso, DEA. selaku dosen pembimbing skripsi yang banyak memberi motivasi, bimbingan, saran, materi, waktu, dan hal lainnya. Dr. Ir. Abdul Muin Adnan, MS. selaku dosen penguji tamu yang telah memberikan kritik dan saran untuk penyempurnaan penulisan skripsi. Seluruh Staff Departemen Proteksi Tanaman IPB baik Dosen Pengajar, Laboran, Petugas Teknis, dan yang lainnya. Keluarga tercinta Ibu, Ayah, Kakak-Kakak, beserta keluarga yang lainnya untuk kasih sayang, doa, serta dukungan yang selalu diberikan. Teman-teman laboratorium Patologi Serangga (Ushbatul Umami, kak Tia, kak Daniar, ibu Diana, ibu Tuti, kak Agung, kak Farriza, ibu Silvi, Syifa, Suci, Arif dan yang lainnya) atas bantuan dan motivasi yang telah diberikan selama penulis melakukan penelitian. Teman-teman seperjuangan angkatan 47 di Departemen Proteksi Tanaman, serta pihak lain yang turut membantu dalam pelaksanaan tugas akhir ini. Semoga skripsi ini bermanfaat.
Bogor, Januari 2015
xvii
DAFTAR ISI
DAFTAR TABEL x
DAFTAR GAMBAR x
DAFTAR LAMPIRAN x
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Manfaat Penelitian 2
METODE 3
Tempat dan Waktu Penelitian 3
Bahan dan Alat 3
Prosedur 4
Perbanyakan Cendawan B. bassiana pada Media Tumbuh 4
Kerapatan Konidia B. bassiana 4
Daya Kecambah Konidia B. bassiana 5
Rancangan Percobaan 5
HASIL DAN PEMBAHASAN 6
Morfologi Cendawan Entomopatogen Beauveria bassiana 6 Pertumbuhan Cendawan Entomopatogen B. bassiana di Media Tumbuh 7 Pengaruh Umur Biakan dan Jenis Media Tumbuh terhadap Kerapatan Konidia
Cendawan B. bassiana 8
Pengaruh Jenis Media Tumbuh, Waktu Inkubasi dan Umur Biakan terhadap Daya Kecambah Konidia B. bassiana dalam Media PDA cair 10 Pengaruh Jenis Media Tumbuh, Waktu Inkubasi dan Umur Biakan terhadap Daya Kecambah Konidia B. bassiana dalam Media PDA tipis 12
SIMPULAN DAN SARAN 15
Simpulan 15
Saran 15
DAFTAR PUSTAKA 16
LAMPIRAN 18
xix
DAFTAR TABEL
1 Jumlah konidia B.bassiana umur biakan 21 dan
33 HSI (hari setelah inokulasi) 8
2 Pengaruh umur biakan terhadap jumlah konidia B. bassiana 9 3 Pengaruh jenis media tumbuh terhadap jumlah total konidia B. bassiana 9 4 Daya kecambah konidia B. bassiana umur 21 dan 33 HSI
pada media inkubasi PDA cair 10
5 Pengaruh jenis media dan umur biakan terhadap daya
kecambah konidia B. bassiana dalam media PDA cair selama 16 JSIn 11 6 Pengaruh jenis media dan umur biakan terhadap daya
kecambah konidia B. bassiana dalam media PDA cair selama 24 JSIn 12 7 Daya kecambah konidia B. bassiana umur 21 HSI dan 33 HSI
dengan media inkubasi PDA tipis 13
8 Pengaruh jenis media dan umur biakan terhadap daya
kecambah konidia B. bassiana dalam media PDA tipis selama 16 JSIn 13 9 Pengaruh jenis media dan umur biakan terhadap daya
kecambah konidia B. bassiana dalam media PDA tipis selama 24 JSIn 14
DAFTAR GAMBAR
1 Tempat pemeliharaan larva O. furnacalis dan imago L. oratorius 4
2 Cendawan B. bassiana 6
3 Koloni cendawan B. bassiana di media 7
DAFTAR LAMPIRAN
1 Hasil RAL terhadap daya kecambah konidia B. bassiana
dalam media PDA cair 19
2 Hasil RAL terhadap daya kecambah konidia B. bassiana
1
PENDAHULUAN
Latar Belakang
Usaha petani dalam pengendalian hama telah dilakukan dengan berbagai cara, namun sampai saat ini yang paling populer di kalangan petani yaitu dengan menggunakan pestisida kimia sintetik. Faktor pendorong petani dalam menggunakan pestisida sintetik yaitu hasil aplikasi pestisida dapat langsung terlihat dan praktis dalam penggunaannya. Dampak negatif dari penggunaan pestisida sintetik secara tidak bijaksana selain pencemaran terhadap lingkungan juga dapat menyebabkan resistensi hama, resurjensi, munculnya hama sekunder, membunuh serangga bukan sasaran, serta residu pada hasil produksi yang dapat membahayakan manusia (Gullan dan Cranston 2010).
Pengendalian hama terpadu (PHT) merupakan pengendalian yang memadukan beberapa cara yaitu secara kultur teknis, pengendalian fisik mekanik dan penggunaan agens hayati (Norris et al. 2003), namun cara ini jarang dilakukan oleh petani. Komponen PHT yang telah banyak dikembangkan dan banyak diteliti yaitu pengendalian dengan menggunakan musuh alami. Pengendalian hama dengan menggunakan musuh alami dapat menggunakan beberapa agens seperti predator, parasitoid dan patogen serangga yang meliputi bakteri, virus, protozoa, nematoda dan cendawan (Tanada dan Kaya 1993). Banyak cendawan entomopatogen yang telah diteliti virulensinya dan diuji dalam pengendalian hama baik skala laboratorium maupun lapangan, salah satunya yaitu cendawan Beauveria bassiana (Balsamo) Vuillemin. Cendawan B. bassiana termasuk dalam filum Ascomycota, subfilum Pezizomycotina, kelas Sordariomycetes, ordo Hypocreales (Vega et al. 2012). Cendawan ini bersifat entomopatogen yang memiliki inang terbanyak di antara cendawan entomopatogen lain. Inang dari cendawan ini paling banyak dari ordo Lepidoptera, Coleoptera dan Hemiptera, namun juga ditemukan menyerang ordo Diptera dan Hymenoptera (Tanada dan Kaya 1993).
Cendawan B. bassiana telah banyak diekplorasi kemampuannya dalam mengendalikan hama. Beberapa peneliti telah menguji keefektifan cendawan ini terhadap beberapa jenis hama di antaranya penggerek batang jagung Ostrinia furnacalis (Lepidoptera: Crambidae) (Agustin 2014), hama boleng Cylas formicarius (Coleoptera: Brentidae) (Ahdiaty 2013), Thrips Frankliniella occidentalis (Thysanoptera: Thripidae) (Gao et al. 2012), kutudaun kedelai Aphis glycine (Hemiptera: Aphididae) (Afifah 2011), kumbang bubuk Sitophilus zeamais (Coleoptera: Curculionidae) (Surtikanti et al. 2011), ulat grayak Spodoptera litura (Lepidoptera: Noctuidae) (Surtikanti dan Yasin 2009), hama kubis Crocidolomia pavonana (Lepidoptera: Pyralidae) (Trizelia 2005) dan kumbang kolorado pada kentang Leptinotarsa decemlineata (Coleoptera: Chrysomelidae) (Klinger 2003).
Media tumbuh akan mempengaruhi virulensi suatu biakan cendawan. Dalam hal ini, cendawan dalam keadaan menginfeksi serangga inang akan tetap bertahan dengan membentuk struktur yang resisten bila kondisi lingkungan yang tidak mendukung (Tanada dan Kaya 1993).
Virulensi suatu cendawan dapat diketahui dengan melihat tingkat pembentukan konidia serta kemampuan konidianya dalam berkecambah. Semakin tinggi daya kecambah konidia suatu cendawan maka semakin tinggi virulensi cendawan tersebut. Menurut Altre et al. (1999) virulensi cendawan entomopatogen berkaitan dengan ukuran konidia, kecepatan perkecambahan konidia, dan produksi enzim yang berfungsi sebagai pendegradasi kutikula inang.
Tujuan Penelitian
Penelitian ini bertujuan menguji produksi dan daya kecambah spora B. bassiana pada inang alami larva O. furnacalis dan imago L. oratorius serta pada media tumbuh PDA, beras dan beras + jagung.
Manfaat Penelitian
3
METODE
Tempat dan Waktu Penelitian
Penelitian dilaksanakan di Laboratorium Patologi Serangga Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor dari bulan Februari 2014 sampai bulan Juli 2014.
Bahan dan Alat
Isolat Cendawan Beauveria bassiana
Isolat cendawan B. bassiana didapatkan dari walang sangit Leptocorisa oratorius (Hemiptera: Alydidae) yang terinfeksi cendawan ini di pertanaman padi Desa Cikarawang, Kecamatan Dramaga, Kabupaten Bogor. Cendawan yang tumbuh diisolasi dan dimurnikan pada media PDA (Potato Dextrose Agar) yang mengandung chloramphenicol dalam cawan petri berdiameter 9 cm. Koloni cendawan yang tumbuh diisolasi kembali dan dibiakan di media PDA baru, kemudian diidentifikasi menggunakan buku Illustrated Genera of Imperfect Fungi (Barnet dan Hunter 1998). Cendawan B. bassiana yang telah murni kemudian ditumbuhkan pada suhu ruang selama ± 21 HSI (hari setelah inokulasi), guna keperluan penelitian lebih lanjut.
Pemeliharaan Ostrinia furnacalis dan Leptocorisa oratorius
Telur O. furnacalis didapatkan dari kebun jagung Leuwi Kopo, Dramaga Bogor yang kemudian dipelihara di laboratorium. Telur yang didapat disimpan dalam cawan petri yang dialasi tisu lembab dan dibiarkan sampai menetas. Larva kemudian disimpan dalam cawan petri berdiameter 15 cm yang dialasi tisu dan diberi pakan buah jagung muda. Setelah larva mencapai instar IV kemudian dipindah ke wadah plastik berukuran panjang 20 cm x lebar 15 cm x tinggi 6 cm yang telah diberi lubang untuk sirkulasi udara dan dialasi tisu (Gambar 1A). Pupa dimasukan ke dalam wadah plastik dengan diameter 9 cm dan tinggi 10.5 cm yang diberi penutup dan dibiarkan sampai menjadi imago. Imago O. furnacalis dipindah ke dalam kurungan berukuran panjang 38.5 cm x lebar 31 cm x tinggi 30 cm yang pada bagian atasnya ditutup dengan kertas roti (baking paper) sebagai tempat bertelurnya imago. Kurungan kemudian ditutup dengan busa lembab begitu pula pada bagian dasar kurungan diberi alas busa lembab (Gambar 1B). Imago diberi pakan madu dengan konsentrasi 10% yang diresapkan ke busa di dalam cawan petri. Pakan madu diganti setiap dua hari sekali. Kertas roti yang diletaki telur O. furnacalis kemudian digunting dan disimpan di dalam cawan petri berdiameter 9 cm yang dialasi tisu lembab. Telur tersebut disimpan beberapa hari sampai menetas menjadi larva.
Gambar 1 Tempat pemeliharaan larva O. furnacalis (A), imago O. furnacalis (B), L. oratorius (C)
Prosedur
Perbanyakan Cendawan B. bassiana pada Media Tumbuh
Konidia B. bassiana hasil pemurnian pada media PDA diinokulasikan pada lima media baru yaitu 50 g beras, 50 g beras + jagung, PDA, larva O. furnacalis instar III dan imago L. oratorius. Suspensi konidia dibuat dengan cara mencampurkan biakan B. bassiana dengan aquades steril. Inokulasi suspensi konidia ke media beras, beras + jagung, larva O. furnacalis instar IIIdan imago L. oratorius dilakukan dengan cara disemprotkan pada media tersebut di dalam laminar flow. Sedangkan inokulasi ke media PDA baru dilakukan dengan memindahkan inokulum menggunakan jarum inokulasi. Media beras, beras + jagung dan PDA telah disterilkan lebih dulu dengan autoklaf selama 35 menit pada suhu 121oC. Media beras atau beras + jagung ditempatkan dalam kantong plastik HDPE (High Density Polyethylene) yang tahan panas. Larva O. furnacalis instar III dan imago L. oratorius yang telah diinokulasi B. bassiana masing-masing disimpan di dalam wadah steril kemudian dipelihara sampai ada larva O. furnacalis dan imago L. oratorius yang mati. Serangga yang mati kemudian dipindahkan ke cawan petri steril berdiameter 6 cm yang dialasi tisu lembab.
Kerapatan Konidia B. bassiana
Kelima media tumbuh yang telah diinokulasi cendawan B. bassiana dan diinkubasi selama 21 HSI dan 33 HSI diselimuti miselium berwarna putih. Media beras dan beras + jagung ditumbuk menggunakan mortar dan ditambahkan 0.05% larutan Tween 20 (v/v) sebanyak 100 ml agar konidia menyebar (Goettel dan Inglis 1997), kemudian disaring menggunakan kain berbahan nilon. Selanjutnya, suspensi dikocok menggunakan vortex selama 30 detik.
Cendawan B. bassiana yang menyelimuti permukaan media PDA, larva O. furnacalis maupun imago L. oratorius diambil menggunakan kuas steril. Cendawan kemudian dibuat suspensi dalam 0.05% larutan Tween 20 (v/v) sebanyak 10 ml
C
5
kemudian disaring menggunakan kain berbahan nilon. Selanjutnya, suspensi konidia dikocok menggunakan vortex selama 30 detik. Kerapatan konidia dari media tumbuh dan inang alami dihitung menggunakan haemocytometer Neubauer improved sebanyak dua kali ulangan dan dilakukan dengan menggunakan mikroskop compound merk Olympus model BX51 dengan perbesaran 400 kali.
Daya Kecambah Konidia B. bassiana
Pengamatan daya kecambah dilakukan setelah suspensi konidia B. bassiana diinokulasikan pada media PDA cair dalam tabung reaksi dan media PDA tipis dalam cawan berdiameter 9 cm. Media PDA cair dibuat dengan cara melarutkan 1 gram PDA yang telah diformulasikan (Difco) ke dalam 100 ml aquades. Media PDA tipis dibuat dengan cara melarutkan 39 gram PDA yang sama dalam 1 liter aquades ditambah 1 gram chloramphenicol. Suspensi konidia dalam dua media tersebut kemudian diinkubasi pada suhu kamar. Pengamatan daya kecambah konidia dilakukan pada 16 dan 24 JSIn (jam setelah inkubasi). Perhitungan konidia dalam PDA cair menggunakan haemocytometer sedangkan pengamatan daya kecambah pada PDA tipis dilakukan langsung di bawah mikroskop compound setelah dilakukan pewarnaan dengan lactophenol cotton blue. Konidia dianggap berkecambah apabila tabung kecambah melebihi dua kali diameter konidia (Goettel dan Inglis 1997). Pengamatan daya kecambah diulang sebanyak tiga kali. Persentase daya kecambah dihitung menggunakan rumus Goettel dan Inglis (1997):
Daya kecambah =jumlah konidia yang berkecambahjumlah konidia yang diamati × %
Rancangan Percobaan
HASIL DAN PEMBAHASAN
Morfologi Cendawan Entomopatogen Beauveria bassiana
Isolat Cendawan B. bassiana diperoleh dari walang sangit Leptocorisa oratorius (Hemiptera: Alydidae) yang terinfeksi cendawan ini dari pertanaman padi Desa Cikarawang, Kecamatan Dramaga, Kabupaten Bogor. Gambar 2A merupakan ciri makroskopis dari cendawan B. bassiana yang menginfeksi serangga inang L. oratorius. Koloni B. bassiana berwarna putih kekuningan menyelimuti tubuh serangga sehingga cendawan ini disebut dengan penyakit white muscardine (Tanada dan Kaya 1993). Cendawan B. bassiana dalam keadaan menginfeksi inang akan mengeluarkan metabolit sekunder yaitu berupa antibiotik yang mencegah pembusukan oleh bakteri sehingga serangga inang menjadi mengeras dan kaku atau disebut dengan mumifikasi (McCoy et al. 1988). Cendawan B. bassiana terdapat di seluruh dunia, memiliki inang yang luas dan juga terdapat dalam tanah sebagai saprofit (Tanada dan Kaya 1993), sehingga cendawan ini mudah didapatkan untuk dijadikan isolat untuk perbanyakan. Trizelia (2005) mendapatkan berbagai isolat cendawan B. bassiana dari berbagai cadaver serangga inang yang kadang-kadang tidak bisa dibedakan secara morfologi.
Gambar 2 Cendawan B. bassiana. (A) koloni cendawan B. bassiana yang menginfeksi imago L. oratorius, terlihat pertumbuhan intensif miselia menyelimuti tubuh serangga, (B) konidiofor B. bassiana khas berbentuk zig-zag, perbesaran 400x.
Hasil pengamatan mikroskopis menunjukkan konidiofor tumbuh secara bergerombol bercabang-cabang dan berbentuk zig-zag, konidia muncul pada konidiofor berbentuk bulat dan tidak berwarna (Gambar 2B). Hal ini sesuai dengan deskripsi Boucias dan Penland(1998) bahwa B. bassiana memiliki konidia tidak berwarna (hyaline) dan memiliki bentuk yang bulat sampai lonjong, hifa bersekat-sekat, dan percabangan konidiofor berbentuk zig-zag, sedangkan miselium di bawahnya menggelembung. Konidiofor dapat terbentuk secara sendiri-sendiri atau dapat berkelompok dalam bentuk kelompok beraturan atau gelungan.
7
Pertumbuhan Cendawan Entomopatogen B. bassiana di Media Tumbuh
Cendawan B. bassiana pada media PDA, media beras dan beras + jagung mulai terlihat tumbuh pada 2-3 HSI (hari setelah inokulasi), sedangkan pada inang alami cendawan mulai tumbuh keluar dari dalam tubuh serangga pada 5-6 hari setelah serangga mati. Cendawan B. bassiana menyebabkan bercak cokelat pada tubuh larva O. furnacalis sebelum menyebabkan kematian. Bercak ini merupakan nekrotik yang disebabkan oleh enzim yang diproduksi oleh B. bassiana saat perkecambahan diantaranya enzim protease, kitinase dan lipase yang dapat mendegradasi integumen inang (Boucias dan Penland 1998). Cendawan B. bassiana pertama kali keluar dari serangga L. oratorius melalui bagian tubuh yang lunak seperti antar ruas-ruas tungkai dan antena, sedangkan pada larva O. furnacalis langsung keluar dari tubuh larva.
Koloni cendawan B. bassiana umur 21 dan 33 HSI terlihat jelas menyelimuti semua media tumbuh (Gambar 3). Cendawan tumbuh dan berkembang menyelimuti penuh media beras, beras + jagung dan inang alami, kecuali pada media PDA cendawan tidak tumbuh memenuhi cawan petri (Gambar 3A). Hal ini disebabkan oleh sifat dari isolat B. bassiana yang ditumbuhkan. Berdasarkan kriteria yang dikemukakan oleh Rayner dan Boddy (1988) karakteristik tekstur dari pertumbuhan cendawan isolat ini mempunyai tipe Velvety dengan ciri hifa pendek, lurus dan tebal. Hasil penelitian Trizelia (2005) juga menunjukkan bahwa beberapa isolat B. bassiana dari berbagai sumber mempunyai karakteristik tekstur pertumbuhan yang berbeda pada media SDAY (Sabouraud dextrose agar dengan yeats extract) dalam cawan petri.
Gambar 3 Koloni cendawan B. bassiana di media PDA (A), imago L. oratorius (B), larva O. furnacalis (C), media beras (D) dan media beras + jagung (E)
Pengaruh Umur Biakan dan Jenis Media Tumbuh terhadap Kerapatan Konidia Cendawan B. bassiana
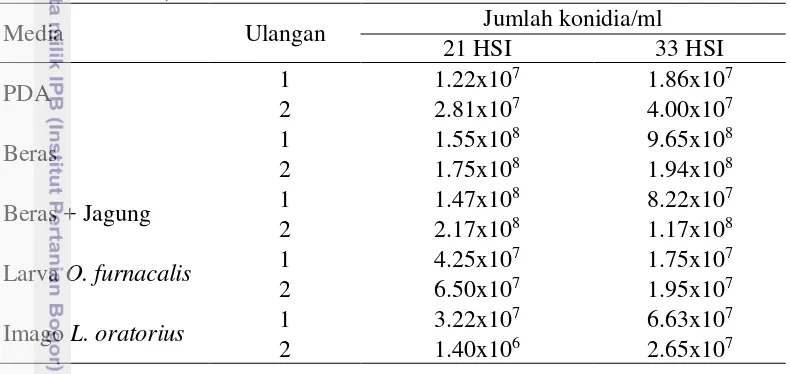
Cendawan B. bassiana dari setiap media biakan menghasilkan kerapatan konidia yang berbeda-beda. Secara umum kerapatan konidia cendawan yang berasal dari media beras dan beras + jagung memiliki kerapatan konidia 108 konidia/ml lebih tinggi dibandingkan dengan PDA dan inang alami yang mencapai 107 konidia/ml (Tabel 1). Kelima media mengandung jenis nutrisi yang diperlukan oleh B. bassiana untuk pertumbuhannya terutama sumber karbon dan nitrogen, akan tetapi dimungkinkan terdapat perbedaan dalam jumlah nutrisi sehingga menghasilkan kerapatan konidia yang berbeda.
Tabel 1 Jumlah konidia B.bassiana umur biakan 21 dan 33 HSI (hari setelah inokulasi)
Media Ulangan Jumlah konidia/ml
21 HSI 33 HSI
Larva O. furnacalis 1 4.25x10
7 1.75x107
2 6.50x107 1.95x107
Imago L. oratorius 1 3.22x10
7 6.63x107
2 1.40x106 2.65x107
Faktor yang mempengaruhi produksi konidia B. bassiana dalam biakan massal yaitu jenis dan jumlah nutrisi yang terkandung pada media tumbuh yang digunakan terutama perbandingan kandungan antara karbon dan nitrogen. Media yang mengandung 3% sukrosa sebagai sumber karbon dan 1% asam casamino sebagai sumber nitrogen dengan rasio perbandingan 22.4 menyebabkan produksi konidia B. bassiana optimal mencapai 107 konidia/ml (Pham et al. 2009). Luas permukaan media tumbuh juga mempengaruhi jumlah konidia yang dihasilkan. Semakin luas permukaan media maka akan semakin banyak konidia yang dihasilkan. Media yang cenderung menggumpal akan memiliki luas permukaan yang sempit, sehingga produksi konidia juga sedikit. Media yang ideal adalah media yang tidak hanya mempunyai partikel dengan permukaan luas, tetapi juga yang dapat mempertahankan keutuhan partikel selama proses produksi (Maheva et al. 1984; Bradley et al. 1992).
9
permukaan PDA, atau dengan satu larva O. furnacalis maupun dengan satu imago L. oratorius.
Tabel 2 Pengaruh umur biakan terhadap jumlah total konidia B. bassiana
1Analisis umur biakan terhadap jumlah konidia B. bassiana menggunakan uji t dengan P=0.05
HSI= hari setelah inokulasi
Umur biakan tidak berpengaruh signifikan terhadap kerapatan konidia yang dihasilkan. Semua media menunjukkan nilai |t hitung| kurang dari t tabel yaitu 4.30 yang berarti rata-rata kerapatan konidia pada semua media pada umur 21 HSI tidak berbeda nyata dengan rata-rata kerapatan konidia pada umur 33 HSI pada P=0.05. Secara umum jumlah konidia menunjukkan kenaikan seiring bertambahnya umur biakan. Kenaikan jumlah konidia terjadi pada media beras, larva O. furnacalis dan imago L. oratorius. Penurunan jumlah konidia terjadi pada media beras + jagung. Sedangkan pada media PDA jumlah konidia tidak mengalami perubahan dari 21 HSI sampai 33 HSI. Penurunan jumlah konidia pada media beras + jagung diduga sama dengan media jagung yaitu penurunan ini disebabkan oleh nutrisi yang terkandung dalam media menurun seiring lamanya umur penyimpanan sehingga cendawan tidak dapat meneruskan pertumbuhannya. Hasil penelitian Purnama et al. (2003) menunjukkan bahwa kerapatan konidia pada media jagung menurun dari 2.30 x 109 konidia/mm3 pada umur 21 HSI menjadi 8.17 x 108 konidia/mm3 pada umur 28 HSI. Sedangkan pada media PDA diduga karena pertumbuhan optimal B. bassiana pada PDA hanya sampai 21 HSI.
Tabel 3 Pengaruh jenis media tumbuh terhadap jumlah total konidia B. bassiana Media
Beras + Jagung 1.82x1010 9.96x109
Larva O. furnacalis 5.38x108
-1.94 9.73x10 8
-0.63 4.30 Imago L. oratorius 1.68x108 4.64x108
1Analisis jenis media tumbuh terhadap jumlah konidia B. bassiana menggunakan uji t dengan
P=0.05
Selain pengaruh umur biakan terhadap kerapatan konidia B. bassiana, dapat dilihat juga pengaruh jenis media terhadap kerapatan konidia yang dihasilkan. Berdasarkan hasil uji (Tabel 3), media beras dan beras + jagung baik pada umur 21 HSI maupun umur 33 HSI nilai |t hitung| kurang dari t tabel yang berarti rata-rata kerapatan konidia pada media beras tidak berbeda nyata dengan rata-rata kerapatan konidia pada media beras + jagung pada P=0.05. Hal yang sama juga dihasilkan
Media Jumlah total konidia t hitung1 t tabel
21 HSI 33 HSI
Beras 1.65x1010 5.80x1010 -1.07 4.30
Beras + Jagung 1.82x1010 9.96x109 2.11 4.30
PDA 2.02x108 2.93x108 -0.69 4.30
nilai |t hitung| dari inang alami larva O. furnacalis dan imago L. oratorius baik pada umur 21 HSI maupun 33 HSI kurang dari t tabel yang berarti rata-rata kerapatan konidia dari kedua inang alami tersebut tidak berbeda nyata.
Dosis aplikasi B. bassiana di lapangan bergantung pada jenis hama sasaran yang akan dikendalikan dan infektivitas isolat cendawan yang digunakan. Isolat cendawan B. bassiana dari kelima media dapat digunakan sebagai biopestisida untuk pengendalian hama di lapangan. Namun terlebih dahulu perlu diuji tingkat infektivitasnya terhadap hama sasaran. Pengendalian larva penggerek batang jagung di lapangan dengan alat semprot dapat menggunakan B. bassiana kerapatan konidia 107 konidia/g dengan dosis 2 kg/100 liter air untuk aplikasi 0.5 ha (Greenmax 2013). Berdasarkan rujukan tersebut pengendalian hama penggerek batang jagung di lapangan membutuhkan B. bassiana asal PDA sebanyak 200 cawan petri/ha, atau 80 larva O. furnacalis/ha, atau 80 imago L. oratorius/ha, atau 4 kantong/ha media beras dengan berat 50 g/kantong, atau 4 kantong/ha media beras + jagung dengan berat 50 g/kantong.
Pengaruh Jenis Media Tumbuh, Waktu Inkubasi dan Umur Biakan terhadap Daya Kecambah Konidia B. bassiana dalam Media PDA cair
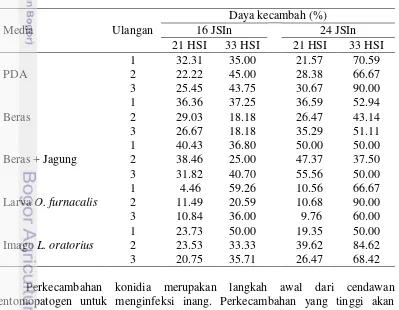
Hasil penelitian menunjukkan bahwa daya kecambah dari lima media mencapai 4%-59% pada 16 JSIn (jam setelah inkubasi) dan 9%-90% pada 24 JSIn (Tabel 4).
Tabel 4 Daya kecambah konidia B. bassiana umur 21 dan 33 HSI pada media inkubasi PDA cair
11
kecambah konidia B. bassiana sebesar 92.4% dapat menyebabkan mortalitas tertinggi pada Sitophilus zeamais.
Kemampuan daya kecambah (viabilitas) konidia cendawan B. bassiana dipengaruhi oleh beberapa faktor yaitu jenis dan asal isolat cendawan, umur cendawan, media pertumbuhan, media tempat perkecambahan, suhu dan lama inkubasi. Menurut Tanada dan Kaya (1993) viabilitas konidia dapat menurun apabila selama subkultur terjadi penurunan sumber karbon seperti glukosa, glukosamin, nitrogen dan pati. Tabel 5 menunjukkan setiap media tumbuh menghasilkan daya kecambah yang berbeda-beda. Daya kecambah konidia cendawan umur 21 HSI yang diinkubasi 16 JSIn paling tinggi yaitu sebesar 36.9% dari media beras + jagung. Sedangkan daya kecambah konidia cendawan umur 33 HSI paling tinggi yaitu 41.3% berasal dari media PDA. Berdasarkan uji Duncan pada α=0.05 daya kecambah konidia 16 JSIn dari cendawan umur 21 HSI menunjukkan adanya perbedaan yang nyata antar media. Daya kecambah asal media beras + jagung tidak berbeda nyata dengan media beras, tetapi berbeda nyata dengan media lainya. Sedangkan daya kecambah konidia cendawan umur 33 HSI tidak berbeda nyata di setiap media.
Tabel 5 Pengaruh jenis media dan umur biakan terhadap daya kecambah konidia B. bassiana dalam media PDA cair selama 16 JSIn
Media Rata-rata daya kecambah (%) ± SD 1
1Angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji Duncan
α= 0.05
2 Analisis umur biakan terhadap daya kecambah konidia B. bassiana menggunakan uji t dengan P=
0.05
Selain jenis media tumbuh, umur biakan juga mempengaruhi daya kecambah. Umur biakan dan kondisi lingkungan mempengaruhi kualitas fisiologi inokulum cendawan dan sangat menentukan potensi cendawan tersebut untuk dijadikan biopestisida (Hallsworth dan Magan 1996). Ahdiaty (2013) menunjukkan bahwa daya kecambah konidia B. bassiana yang diinkubasi dalam air selama 10 JSIn meningkat seiring bertambahnya umur cendawan: daya kecambah umur 13 HSI mencapai 25.7%, 36 HSI mencapai 33.5% dan 48 HSI mencapai 48.6%.
Dengan waktu inkubasi 24 JSIn secara berturut-turut daya kecambah cendawan umur 21 dan 33 HSI paling tinggi yaitu sebesar 50.9% (media beras + jagung) dan 75.8% (media PDA) (Tabel 6). Menurut Kassa (2003) cendawan yang telah memenuhi syarat untuk dikembangkan sebagai bioinsektisida yaitu konidia yang memiliki daya kecambah di atas 80%. Liu et al. (2003) menyarankan bahwa daya kecambah konidia cendawan yang akan digunakan sebagai agens hayati harus diatas 90%. Namun, secara umum daya kecambah yang diinkubasi dalam PDA cair paling tinggi hanya mencapai 75.8%.
Tabel 6 Pengaruh jenis media dan umur biakan terhadap daya kecambah konidia B. bassiana dalam media PDA cair selama 24 JSIn
Media Rata-rata daya kecambah (%) ± SD 1
1Angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji Duncan
α= 0.05
2 Analisis umur biakan terhadap daya kecambah konidia B. bassiana menggunakan uji t dengan P=
0.05
Daya kecambah konidia dari semua media kecuali beras + jagung mengalami peningkatan seiring dengan bertambah tua umur biakan. Peningkatan ini nyata pada P=0.05 dimana nilai |t hitung| lebih besar dari t tabel, ini berarti rata-rata daya kecambah konidia cendawan umur 21 HSI dari keempat media ini berbeda nyata dengan rata-rata daya kecambah konidia cendawan umur 33 HSI. Daya kecambah konidia dari media beras + jagung mengalami penurunan tetapi penurunan tersebut tidak nyata perbedaannya antara umur 21 HSI dengan 33 HSI. Berdasarkan uji Duncan daya kecambah pada media beras + jagung 21 HSI berbeda nyata dengan daya kecambah konidia dari media lainnya. Daya kecambah konidia cendawan umur 33 HSI asal PDA dan inang alami merupakan daya kecambah yang termasuk tinggi di antara semua media. Daya kecambah konidia asal PDA tidak berbeda nyata baik dengan larva O. funacalis maupun L. oratorius.
Waktu inkubasi mempengaruhi daya kecambah konidia B. bassiana. Dalam penelitian ini ditunjukkan bahwa semakin lama waktu inkubasi maka semakin tinggi daya kecambah konidia. Semakin lama konidia berada dalam media inkubasi maka semakin banyak nutrisi yang diserap untuk perkecambahan konidia sehingga perkecambahan terus berlangsung.
Pengaruh Jenis Media Tumbuh, Waktu Inkubasi dan Umur Biakan terhadap Daya Kecambah Konidia B. bassiana dalam Media PDA Tipis
13
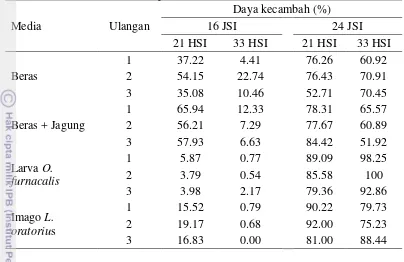
Tabel 7 Daya kecambah konidia B. bassiana umur 21 HSI dan 33 HSI dengan media inkubasi PDA tipis
Tabel 8 menunjukkan bahwa konidia B. bassiana dari media beras + jagung umur 21 HSI yang diinkubasi 16 JSIn mempunyai daya kecambah paling tinggi sebesar 60%. Konidia asal beras, larva O. furnacalis dan imago L. oratorius mempunyai daya kecambah yang lebih rendah.
Tabel 8 Pengaruh jenis media dan umur biakan terhadap daya kecambah konidia B. bassiana dalam media PDA tipis selama 16 JSIn
Media Rata-rata daya kecambah (%) ± SD 1
1Angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji Duncan
α= 0.05
2 Analisis umur biakan terhadap daya kecambah konidia B. bassiana menggunakan uji t dengan P=
0.05
Inkubasi selama 24 JSIn (Tabel 9) memberikan pola perkecambahan yang berbeda. Pada semua media, konidia menunjukkan persentase perkecambahan yang tinggi.
Tabel 9 Pengaruh jenis media dan umur biakan terhadap daya kecambah konidia B. bassiana dalam media PDA tipis selama 24 JSIn
Media Rata-rata daya kecambah (%) ± SD 1
t hitung2 t tabel
21 HSI 33 HSI
Beras 68.5±13.6b 67.4±5.6c 0.12 2.78
Beras + Jagung 80.1±3.7ab 59.5±6.9c 4.55 2.78 Larva O. furnacalis 84.7±4.9a 97.0±3.7a -3.47 2.78 Imago L. oratorius 87.7±5.9a 81.1±6.7b 1.28 2.78
1Angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji Duncan
α= 0.05
2 Analisis umur biakan terhadap daya kecambah konidia B. bassiana menggunakan uji t dengan P=
0.05
15
SIMPULAN DAN SARAN
Simpulan
Kelima jenis media tumbuh berpengaruh terhadap kerapatan dan daya kecambah konidia cendawan B. bassiana. Media beras dan beras jagung menghasilkan konidia total mencapai 1010, PDA dan inang alami menghasilkan 108. Umur biakan tidak berpengaruh terhadap kerapatan konidia, namun berpengaruh terhadap daya kecambah konidia pada beberapa media. Daya kecambah meningkat seiring meningkatnya waktu inkubasi, daya kecambah konidia 24 JSIn lebih tinggi dari 16 JSIn. Daya kecambah konidia cendawan yang ditumbuhkan pada inang alami lebih tinggi dibandingkan dengan daya kecambah konidia cendawan yang ditumbuhkan pada media alternatif.
Saran
DAFTAR PUSTAKA
Afifah L. 2011. Pertumbuhan cendawan entomopatogen Lecanicillium lecanii pada berbagai media serta infektivitasnya terhadap kutudaun kedelai Aphis glycines Matsumura (Hemiptera: Aphididae) [skripsi]. Bogor (ID): Fakultas Pertanian, Institut Pertanian Bogor.
Agustin D. 2014. Keefektifan cendawan entomopatogen Beauveria bassiana (Balsamo) Vuillemin dan Lecanicillium lecanii (Zimm.) Zare & Gams terhadap penggerek batang jagung asia Ostrinia furnacalis Guenee (Lepidoptera: Crambidae) [tesis]. Bogor (ID): Institut Pertanian Bogor. Ahdiaty I. 2013. Pengaruh umur cendawan Beauveria bassiana (Balsamo)
Vuillemin terhadap infektifitasnya pada Cylas formicarius Fabricius (Coleoptera: Brentidae) [skripsi]. Bogor (ID): Fakultas Pertanian, Institut Pertanian Bogor.
Altre JA, Vandenberg JD, Cantone FA (1999). Pathogenicity of Paecilomyces fumosoroseus isolates to Diamondback Moth, Plutella xylostella: correlation with spore size, germination speed, and attachment to cuticle. J Invertebr Pathol. 73(3): 332-338.
Barnet HL, Hunter BB. 1998. Illustrated Genera of Imperfect Fungi. 4th ed. Minnesota (US): APS press.
Boucias DG, Pendland JC. 1998. Principles of Insect Pathology. Massachusetts (US): Kluwer Academic Publishers.
Bradley CA, Black WE, Kearns R, Wood P. 1992. Role of production technology in mycoinsecticide development. Di dalam: Leatham GF, editor. Frontiers in Industrial Mycology. London (GB): Chapman and Hall. hlm 160-173. Gao Y, Reitz SR, Wang J, Xu X, Lei Z. 2012. Potential of a strain of the
entomopathogenic fungus Beauveria bassiana (Hypocreales: Cordycipitaceae) as a biological control agent against western flower thrips, Frankliniella occidentalis (Thysanoptera: Thripidae). Biocontrol Science and Technology. 22(4): 491-495.
Goettel MS, Inglis GD. 1997. Fungi: Hyphomycetes. Di dalam: Lacey LA, editor. Manual of Technique in Insect Pathology. San Diego (US): Academic Press. hlm 213-249.
Greenmax. 2013. Talc based Beauveria bassiana [internet]. Coimbatore (IN): Greenmax; [diunduh 2014 Desember 14]. Tersedia pada: http//www.greenmaxagrotech.com/beauveria-bassiana.html.
Gullan PJ, Cranston PD. 2010. The Insect. 4th ed. West Sussex (GB): Wiley-Blackwell.
Hallsworth JE, Magan N. 1996. Culture age, temperature, and pH affect the polyol and trehalose contents of fungal propagules. Applied and environmental microbiology. 62(7): 2435-2442.
17
Klinger E. 2003. Susceptibility of adult colorado potato beetle (Leptinotarsa decemlineata) to the fungal entomopathogen Beauveria bassiana [tesis]. Maine (US): University of Maine.
Liu H, Skinner M, Brownbridge M, Parker BL. 2003. Characterization of Beauveria bassiana and Metarhizium anisopliae isolates for management of tarnished plant bug Lygus lineolaris (Hemiptera: Miridae). J Invertebr Pathol. 82(3):139-147.
Maheva E, Djelveh G, Larroche C, Gros JB. 1984. Sporulation of Penicillium roqueforti in solid substrate fermentation. Biotechnology Letters. 6: 97-102. McCoy CW, Samson RA, Boucias DG. 1988. Entomogenous fungi. Di dalam: Ignofo CM, editor. CRC Handbook of Natural Pesticides. Microbial insecticides, Part A Entogenous Protozoa and Fungi. Florida (US): CRC Press. hlm 151-236.
Norris RF, Caswell-Chen EP, Kogan M. 2003. Concepts in Integrated Pest Management. Upper Saddle River (NJ): Prentice Hall.
Pham TA, Kim JJ, Kim SG, Kim K. 2009. Production of blastospore of entomopathogeneic Beauveria bassiana in a submerged batch culture. Mycobiology. 37(3): 218-224.
Purnama PC, Nastiti SJ, Situmorang J. 2003. Uji patogenitas jamur Beauveria bassiana (Bals.) Vuill. isolat Magelang terhadap Aphis craccivora Koch. Biosmart. 5(2): 81-88.
Rayner ADM, Boddy L. 1988. Fungal Decomposition of Wood. New York (US): John Wiley & Sons.
Surtikanti, Yasin M, Tandiabang J. 2011. Pengendalian hama kumbang bubuk menggunakan cendawan Beauveria bassiana Vuill. berupa tepung. Prosiding Seminar Nasional Serealia 2011; Maros. Maros (ID): Balai Penelitian Tanaman Serealia.
Surtikanti, Yasin M. 2009. Keefektifan entomopatogenik Beauveria bassiana Vuill. dari berbagai media tumbuh terhadap Spodoptera litura F. (Lepidoptera : Noctuidae) di laboratorium; 2009; Maros. Maros (ID): Balai Penelitian Tanaman Serealia.
Tanada Y, Kaya HK. 1993. Insect Pathology. San Diego (US): Academic Press, INC.
Trizelia. 2005. Cendawan entomopatogen Beauveria bassiana (Bals.) Vuill. (Deuteromycotina: Hypomycetes): keragaman genetik, karakterisasi fisiologi dan virulensinya terhadap Crocidolomia pavonana (F.) (Lepidoptera: Pyralidae) [disertasi] Bogor (ID): Institut Pertanian Bogor.
19
19
Lampiran 1 Hasil RAL terhadap daya kecambah konidia B. bassiana dalam media PDA cair
HSI= hari setelah inokulasi JSIn= jam setelah inkubasi
Lampiran 2 Hasil RAL terhadap daya kecambah konidia B. bassiana dalam media PDA tipis
HSI= hari setelah inokulasi JSIn= jam setelah inkubasi
Umur biakan (HSI)
Waktu inkubasi
(JSIn)
DB JK KTG F
hitung P
21 16 4 1320.937267 330.234317 18.29 0.0001
24 4 2539.711427 634.927857 18.02 0.0001
33 16 4 546.3254000 136.5813500 1.01 0.4483
24 4 2263.771027 565.942757 3.61 0.0453
Umur biakan (HSI)
Waktu inkubasi
(JSIn)
DB JK KTG F
hitung P 21 16 3 5573.468292 1857.822764 52.76 <.0001
24 3 643.6678917 214.5559639 3.31 0.0781
33 16 3 311.3806250 103.7935417 4.24 0.0454
RIWAYAT HIDUP
Penulis dilahirkan di Sukabumi pada tanggal 17 Januari 1991, sebagai putri dari Bapak Karmun dan Ibu Tuti. Penulis adalah putri terakhir dari tiga bersaudara. Penulis lulus dari SMA Negeri I Cibadak Kabupaten Sukabumi pada tahun 2009 dan pada tahun 2010 penulis lulus seleksi masuk Institut Pertanian Bogor (IPB) melalui jalur Ujian Talenta Mandiri dan diterima di Departemen Proteksi Tanaman, Fakultas Pertanian.