ANALISIS PENGELOMPOKAN DAN HUBUNGAN
KEKERABATAN SPESIES ANGGREK PHALAENOPSIS
BERDASARKAN KUNCI DETERMINASI DAN MARKA RAPD
OLEH:
KRISTINA DFVIATMINI
PROGRAM PASCA SARJANA INSTITUT PERTANIAN B O W R
ABSTRAK
KRISTINA DWIATMINI. Analisis Pengelompokan dan Hubungan Kekerabatan Spesies Anggrek Phalaenopsis Berdasarkan Kunci Determinasi dan Marka RAPD. Dibimbing oleh NURHAYATI A MATTJIK, HAJRIAL ASWIDINNOOR, dan NURITA L TORUAN-MATIUS.
Bunga anggrek Bulan (salah satu spesiedjenis dari marga Phalaenopsrs)
adalah puspa pesona bangsa Indonesia. Di Indonesia terdapat kira-kira 26 spesies
Phalaenopsls yang endemik dari 70 spesies yang dilaporkan. Tanaman anggrek merupakan jenis tanaman yang mempunyai keragaman fenotipik yang sangat besar. Anggrek juga tanaman yang dapat cross fertrl secara intergenerik, sehingga konsep s p i e s secara biologi pada anggrek menjadi rancu. Sampai saat ini sering terjadi pergeseran pengelompokan dan perubahan nama pada suatu jenis anggrek. Karena itu perlu dilakukan penelitian hubungan kekerabatan spesies-spesies Phalaenopsrs dengan menggunakan teknik molekuler untuk melengkapi sistem klasifikasi yang hanya mengandalkan karakter morfologi tanaman.
Penelitian dllakukan di Laboratorium Biologi molekuler dan Imunologi, Unit Penelitian Bioteknologi Perkebunul, Bogor dari bulan September 2001 sarnpai April 2002. Penelitian ini bertujuan untuk mengetahui pengelompokan spesies-spesies anggrek Phalaenopsrs berdasarkan penanda morfologi yang dijadikan basis penyusunan kunci determinasi, mengetahui hubungan kekerabatan berdasarkan analisis pola pita DNA dan mengetahui seberapa jauh keselarasan,korelasi keduanya Enam belas primer digunakan untuk mengamplifikasi DNA berdasu Polymerase Chain Reaction
(PCR)
dan menghasilkan 300 hgrnen DNA yang polimorfik. Dendrogram tanaman anggrek Phalaenopsisdari
data RAPD menggunakan koefisien kemiripan DICE, dari data morfologi menggunakan rumus jarak taksonomi (koefisienDIST) dan korelasi antara matriks kerniripan dan matriks jarak menggunakan fasilitas
MXCOMP pada program NTSYS versi 2.02. Analisis komponen utama (KU) dilakukan untuk mengetahui pita-pita yang berperan dan posisi relatif pengelompokan secara terpisah 19 spesies yang diuji.
Hubungan kekerabatan berdasarkan koefisien kemiripan Dice adalah antara 0.24 - 0.66 (jarak genetik antara 0.34
-
0.76). Sedangkan jarak taksonomi berdasarkan koefisien Dist adalah 1.42 - 0.08. Nilai korelasi ( r ) antara matriks kemiripan dan matriks jarak adalah kecil yaitu -0.38197, dengan nilai koefisien determinasi R' = 0.1459. Nilai koefisien determinasi yang sangat kecil menunjukkanenam genotipe anggrek yang mempunyai posisi relatif dekat yaitu P. pantherina, P.
SllJRAT
PERNYATAAN
Saya menyatakan dengan sebenar-benarnya bahwa segala pernyatan dalam tesis saya
yang berjudul :
"ANALISIS PENGELOMPOKAN DAN HUBUNGAN KEKERABATAN SPESIES ANGGREK PHALAENOPSIS BERDASARKAN KUNCI DETERMINASI DAN MARKA RAPD"
merupakan gagasan atau has11 peilelitian tesis saya sendiri, dengan pembimbingan
Komisi Pembimbing kecuali yang dengan jelas ditunjukkan rujukannya. Tesis ini
belum pernah diajukan untuk memperoleh gelar pada program sejenis di perguruan tinggi lain. Semua data dan informasi yang digunakan telah dinyatakan secara jelas
dan dapat diperiksa kebenarannya.
Bogor, September 2002
Krist ' a Dwiatmini
ANALISIS PENGELOMPOKAN DAN HUBUNGAN
KEKERABATAN SPESIES ANGGREK PHALAENOPSIS
BERDASARKAN KUNCI DETERMINASI DAN MARKA RAPD
KRISTINA DWIATMINI
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Agronomi
PROGRAM PASCA SARJANA INSTITUT PERTANIAN BOGOR
Judul Tesis : Analisis Pengelompokan dan Hubungan Kekerabatan Spesies Anggrek Phalaenopsis Berdasarkan Kunci Deterrninasi dan Marka RAPD
Nama Mahasiswa : Kristina Dwiatrnini Nomor Pokok : 99635
Program Studi : Agronomi
Menyetuj ui 1. Komisi Pembimbing
Dr. Ir. Nurhayati A. Mattjik. MS Ketua
Dr. Hairial Aswidinnoor, MSc Dr. ~ d t a L. Toruan Matius, MS APU
Anggota Anggota
2.Ketua Program Studi Agrono rogram Pascasa jana
2%-
Dr. Ir. Hajrial Aswidinnoor, MS frida Manuwoto, MSc
RJWAYAT HIDUP
Penulis dilahirkan pada tanggal 25 Desember 1966 di Sukoharjo, Jawa Tengah,
sebagai anak kedua dari delapan bersaudara, puteri dari pasangan R. Sastromadyono dan Ibu Suminah. Pada tahun 1996 menikah dengan Eko Artanto dan dikaruniai dua
orang putera yaitu Yonatan Kristanto Nugroho (4.5 tahun) dan Yohanes Kristanto
Raharjo (1 tahun).
Pada tahun 1979 penulis menyelesaikan sekolah dasar di SD Kristen Sukoharjo
dan berturut-turut pada tahun 1982 tamat dari SMP N 2 Sukoharjo dan tahun 1985
tamat dari SMA N Sukoharjo. Selanjutnya melalui program PMDK penulis diterima sebagai mahasiswa di Institut Pertanian Bogor, Fakultas Pertanian dengan program
studi Hama dan Penyakit Tumbuhan. Fenulis memperoleh gelar sajana pertanian
pada tahun 1990.
Setelah bekerja di swasta selama 3 tahun, pada tahun 1993 penulis diterima
sebagai staf peneliti di Sub Balai Penelitian Hortikultura Cipanas yang selanjutnya
menjadi Balai Penelitian Tanaman Hias Segunung. Penulis mengkuti program
Magister Sains pada Program Studi Agronomi/Pemuliaan Tanaman sejak tahun 1999.
PRAKATA
Puji syukur penulis panjatkan kehadirat Tuhan yang Maha Kasih atas berkat dan karunia-Nya yang selalu dilimpahkan sehingga penulis &pat menyelesaikan &is ini.
Terima kasih yang tulus penulis sampaikan kepada Ibu Dr. Nurhayati A Mattjik, MS sebagai ketua komisi pembimbing dan Dr. Ir. Hajrial Aswidinnoor, MSc serta Dr. Nurita L Toruan-Matius, MS APU sebagai anggota komisi pembimbing, atas segala bimbingan dan bantuannya dari awal penelitian hingga selesainya penulisan tesis. Penyelesaian penelitian ini tidak lepas dari bantuan dana proyek PAATP, Badan Litbang Pertanian.
Kepada mereka yang telah banyak berperan, penulis juga ingin berterima kasih kepada Dr. M. Kosim Kardin, MSc atas saran, petunjuk dan sebagian material tanaman yang digunakan dalam penelitian, juga kepada Bapak Ir. Rizal Jafaarer atas material tanaman dan bantuan identifikasinya, Bapak Tolhas Hutabarat, Dipl. Kim., mbak Nanik dan mbak Umi atas bantuan dan kerjasama di laboratorium. Kepada rekan-rekan mahasiswa program studi agronomi angkatan 99 talc lupa diucapkan terima kasih atas dorongan dan semua bantuan yang diberikan selama masa perkuliahan, penelitian dan penulisan tesis.
Kesuksesan dan keberhasilan studi ini tidak terlepas dari pengertian, bantuan serta doa seluruh keluarga. Atas semua ini, dengan sepenuh hati penulis mengucapkan terima kasih kepada Bapak dan Ibu mertua (almarhurnah), ayah dan ibu dan terutarna kepada suarni dan anak-anak yang telah turut berkorban dalam banyak hal. Semoga Tuhan melimpahkan anugerah-Nya kepada kita semua.
Akhirnya penulis berharap semoga tesis ini dapat memberikan informasi yang bermanfaat.
Bogor, September 2002
DAFTAR IS1
Halaman DAFTAR TABEL DAFTAR GAMBAR DAFTAR LAMPIRAN FENDAHULUAN Latar Belakang Tujuan Penelitian Hipotesis TINJAUAN PUSTAKAGenus Phalaenopsis dan Sejarah Klasifikasi Penanda Morfologi clan Penanda Molekuler RAPD Kaitan antara Penanda Morfologi dan Molekuler BAHAN
DAN
METODEBahan Tanaman
Bahan Kimia dan Peralatan Metode Penelitian
Pelaksanaan Percobaan Analisis Data
Tahapan Percobaan Untuk Analisis RAPD Analisis Polimorfisme dengan RAPD
Pengelompokan Berdasarkan Kunci Determinasi
Pengelompokan Gabungan Data RAPD dan Data Biner Fenotipik Korelasi antara Matriks Tingkat Kemiripan Genetik dan
Matriks Jarak Taksonomi
xii
PEMBAHASAN
Hubungan Kekerabatan Berdasarkan Analisis Marka RAPD Pengelompokan Berdasarkan Kunci Determinasi
Pengelompokan Berdasarkan Gabungan Data RAPD dan Fenotipik Keselarasan Pengelompokan Berdasar
RAPD
dan Karakter Fenotipik KESIMPULAN DAN SARANDAFTAR TABEL
1 Kriteria goodness offir
2 Kuantitas DNA hasil ekstraksi pada 2 1 spesies anggrek
Phalaenopsis dibandingkan dengan besamya pita DNA lamda 3 Jenis primer, susunan basa nukleotida, persentase kandungan
GIC dan jumlah pita yang diseleksi
4 Jenis primer, susunan basa nukleotida, kandungan G/C, dan jumlah pita hasil amplifikasi
5 Matriks perkiraan kesamaan koefisien DICE pada 19 spesies
Phalaenopsis
6 Katagorial nilai karakter pembeda fenotipe pada 1 Y spesies dalam genus Phalaenopsis
7 Matriks perkiraan jarak genetik koefisien
DIST
pada 18 spesiesPhalaenopsis
8 Matriks perkiraan kemiripan genetik koefisien DICE pada 18 spesies Phalaenopsis dengan data gabungan
DAFTAR GAMBAR
Bagan alir analisis data
Kuantitas DNA total 20 spesies Phalaenopsis dari bahan daun Kualitas DNA total 20 spesies Phalaenopsis didigesti EcoHl
Tujuh belas primer hasil seleksi menggunakan DNA cetakan
P. violaceae 'Borneo'
Profil pita DNA 19 spesies Phalaenopsis hasil amplfikasi ~nenggunakan primer OPA 1 1
Dendogram 19 spesies Phalaenopsis menggunakan 16 primer, dengan h g s i DICE dan metode UPGMA pada Program NTSYS 2.02.
Pemetaan 19 genotipe Phalaenopsis berdasar 2 dan 3
KU
Pengelompokan spesies Phalaenopsis berdasar kunci determinasi cara non katagorial
Dendogram 18 spesies anggrek dalam genus Phalaenopsis
berbasis karakter fenotipik dengan prosedur STAND dan h g s i SlMMINT diolah dengan progran NTSYS versi 2.02. Dendrogram 18 spesies anggrek Phalaenopsis h a i l analisis klaster berdasarkan data biner gabungan RAPD dm fenotipik dangan metode UPGMA pada program NTSYS 2.02.
Diagram hubungan antara matiks taksonomi berdasarkan fenotipe dengan matiks kemiripan genetik berdasarkan RAPD dari 1 8 genotipe anggrek Phalaenopsis.
DAFTAR
LAMPIRAN
Halaman Daftar Sgesies Anggrek Phalaenopsis yang digunakan untuk
penelitian. 69
Komposisi pereaksi dan buffer untuk analisis DNA tanaman
anggrek. 70
Data biner 19 spesies Phalaenopsis menggunakan 16 primer
acak dekamer. 7 1
Nilai kategorial fenotipik 18 spesies anggrek dalarn genus
Plzalaenopsis. 77
Transformasi nilai katagorial karakter fenotipik menjadi data
biner. 78
Nilai komponen utarna dari pita-pita hasil arnplifikasi 8 1 Teladan analisis pengelompokan berdasarkan data fenotip
menggunakan rumus Average Taxonomic Distance. 86 Teladan analisis pengelompokan berdasarkan pola pita DNA
menggunakan koefisien DICE (rumus Nei dan Li, 1978) 88 Teladan analisis korelasi antara matriks koefisien kemiripan
dengan matriks rata-rata jarak taksonomi dengan korelasi rank
sperman 9 1
Profil pita DNA 19 spesies Phalaenopsis menggucakan primer; OPA 1, OPA 2, OPA 3, OPA 4, OPA 7, OPA 1 1,
OPA 13 dan OPA 16 92
Profil pita DNA 19 spesies Phalaenopsis menggunakan primer; OPB 5, OPB 7, OPB 10, OPB 17, OPC 6, OPC 9,
12 Variasi tonjolan bibir (kalus) beberapa Phalaenopsis
13 Variasi bentuk bibir beberapa Phalaenopsis
14 Variasi bentuk bibir dan detail bunga beberapa Phalaenopsis
PENDAHULUAN
Latar Belakang
Indonesia menggunakan anggrek bulan (Phalaenopsis amabilis (L.) Blurne) sebagai bunga Nasional dengan sebutan puspa pesona. Salah satu pertimbangan anggrek bulan dipilih sebagai bunga Nasional disebabkan Indonesia merupakan wilayah penyebaran marga Phalaenopsis yang terkaya di dunia. Dari 70 spesies Phalaenopsis, 26 spesies di antaranya terdapat di Indonesia (Anonim 1997). Sejarah mencatat bahwa selain Phalaenopsis amabilis, pada tahun 1 865 Phalaenopsis sumatrana mendapat " First Class Certificate" dari Royal Horticultural Society
London (Rivai 1995).
Keindahan bunga anggrek bulan (marga Phulaenopsis) sangat digemari oleh pasar luar negeri. Sebagai contoh, pada tahun 1993 Phalaenopsis telah mencapai peringkat ke-14 dengan penjualan sebesar US $ 12,8 juta di pasar lelang Belanda (Abidin 1997). Anggrek Phalaenopsis yang tersebar di pasaran tersebut berupa spesies maupun hibrida. Statistika hibrida anggrek menunjukkan jurnlah silangan anggrek marga Phalaenopsis (13.064 silangan) menempati peringkat kedua setelah marga Paphiopedilum (Irawati 1997).
Sej arah klasifikasi dari genus Phalaenopsis menunj ukkan beberapa kal i perubahan nama suatu spesies. Sebagai contoh spesies yang pertama kali ditemukan di Ambon pada tahun 1750, yaitu Phalaenopsls amabills (L.) Blume. Rumphius memberi nama Angraecum album-majus yang diterbitkan dalam "Herbarium Arnboinense 6:99". Dalan waktu yang hampir bersamaan pada tahun 1752 Peter Osbeck membawa spesimen jenis ini dari Jawa Barat dan diidentifikasi oleh Linnaeus sebagai Epidendrum amabile yang diterbitkan dalam "Spesies Plantar~rn~~, tanpa mengetahui publikasi yang terdahulu oleh Rumphius. Tahun 1814 Roxburgh juga mengidentifikasi jenis ini sebagai Cymbidium amabile, dan pada tahun 1825 Blurne memasukkan tumbuhan ini ke &lam marga Phalaenopsis dan diberi nama
Phalaenopsis arnabilis (L.) Blume (Sweet 1980; Puspitaningtyas dan Mursidawati
1999). Perubahan nama atau pemindahan pengelompokan suatu spesies sering te rjadi. Sebagai contoh Phalaenopsis serpentilingua (masuk genus Phalaenopsis) namanya diganti menjadi Paraphafaenopsis serpentilingua, terrnasuk genus Paraphafaenopsis
(Hawkes 1970). Pemindahan dalarn genus tersebut didasarkan bentuk daunnya yang berbeda (Hawkes 1970) dan secara genetik Phalaenopsis tidak kompatibel dengan
Paraphalaenopsis (Yam 1 994).
Secara taksonomi fakta suatu spesies ditunjukkan oleh satu set karakter yang diberikan oleh sifat-sifat morfc!oginya. Sifat-sifat tersebut merupakan produk dari
enzim yang spesifik akan hilang dan sifat khusus tersebut tidak terekspresi. Menurut . Griesbach (1981) ilmu genetika &pat membantu klarifikasi permasalahan yang timbul dalam taksonomi. Sebagai contoh permasalahan pada Phalaenopsis digambarkan sebagai berikut. Pada tahun 1859 di Palembang, Sumatera; Gersen menemukan satu
Phalaenopsis baru, yang oleh Teijsmann dan Binnendijk pada tahun 1862 diidentifikasi sebagai P. zebrina var. Garsenii. Bentuk lain dari Phalaenopsis ini ditemukan oleh Lobb dan diklasifikasikan oleh Teijsmann dan Bennedijk sebagai P. zebrina var. lilacina. Beberapa dekade kemudian P. zebrina disetujui sinonim dengan
P. sumatrana, selanjutnya Reichenbach menyatakan kedua tanarnan tersebut sebagai varietas dari P. sumatrana. Kemudian pada tahun 1882 ditemukan bentuk lain dan diidentifikasi oleh Reichenbach sebagai P. violacea var. schroederana. Pada saat ini disetujui bahwa ketiga takson tersebut merclpakan intermediate d a i P. violacea clan
P.
sumatrana. Namun Phalaenopsis x Gersenii yang diyakini sebagai hibrid alam antara
P. violacea dan P. smatrana ternyata menunjukkan beberapa inkonsistensi, tergantung ekotipe dari tetuanya dan terjadinya introgresi. Pada P. sumatrana var. gersenii sangat sedikit terjadi rekombinasi genetik selama meiosis, kemudian
segregasi te rjadi pada generasi yang sudah sangat lanjut.
1883, Rolfe pada tahun 1886,
dan
Pfitzer pada tahun 1889 dikutip oleh Arends( 1970).
Anggrek merupakan jenis tanaman yang sangat dipengaruhi lingkungan, sehingga keragaman karakter fenotipik yang nampak sering rnembingungkan dalam pengelompokan hubungan kekerabatannya. Berdasarkan kunci determinasi terkini yang disusun oleh Sweet (1980) dicoba disusun pengelompokan antar spesies dalarn genus Phalaenopsis. Selanjutnya dalam penelitian ini akan dilakukan studi hubungan kekerabatan beberapa spesies anggrek Phalaenopsis menggunakan marka Randomly Amplrfied Polymorphic DNA (RAPD). Diharapkan dapat diketahui sejauh mana keselarasan antara penanda morfologi berdasarkan kunci determinasi berbasis fenotipik dengan penanda RAPD berbasis DNA, sehingga aplikasi RAPD dapat digunakan untuk studi taksonomi
dan
genetik genus Phalaenopsis tersebut.Tujuan Penelitian
Penelitian ini bertujuan untuk: (I) mengelompokan spesies anggrek
Phalaenopsis berdasarkan karakter fenotipik yang dijadikan basis penyusunan kunci determinasi, (2) mempelajari hubungan kekerabatan spesies anggrek Phalaenopsis
berdasar analisis berbasis DNA dengan t e h k RAPD,
,dan
(3) menetapkan hubunganJkorelasi keduanya.DNA (RAPD) dilakukan analisis komponen utama untuk mengetahui pita-pita berperan dalam pengelompokan dan posisi relatif antar genotipe.
Hipotesis
TINJAUAN PUSTAKA
Genus Phalaenopsis dan Sejarah Klasifikasi
Hamparan daratan negara Indonesia hanyalah 1,3 % dari luas permukaan bumi, namun diperkirakan tidak kurang dari 30 % dari sekitar 17.000 jenis anggrek menghuni bumi nusantara. Jadi di Indonesia terjadi konsentrasi jumlah jenis tanaman anggrek; serta beberapa kelompok tetumbuhan khas lainnya yang sangat besar, dan sering disebut sebagai pusat keanekaragaman hayati dunia (Rivai 1995). Irawati (1999) melaporkan di Jawa terhpat 700 jenis anggrek, sedang di Surnatera dan Kalimantan tidak kurang dari 2000 jenis. Irian Jaya dilaporkan menyimpan berbagai jenis anggrek unik dan khas seperti anggrek macan (Grammatoplyllum scriptum) dan jenis-jenis lainya (Anonim 1992), selain jenis anggrek yang terdapat di kepulauan Indonesia lainnya yang belum teridentifikasi. Identifikasi jenis-jenis anggrek yang ada di Indonesia harus dilakukan secepatnya sebelurn berlomba dengan kerusakan habitatnya. Oleh karena itu melalui program Flora Malesiana dilakukan revisi jenis- jenis tumbuhan di dunia yang diperkirakan baru akan selesai pada tahun 2 135.
Phalaenopsis merupakan anggrek mono@al yang dicirikan sebagai tanaman epifit atau litofit (menempel atau menempel pada batu). Akarnya agak pipih, berdaging dan mengandung klorofil. Berbatang pendek yang seluruhnya terbungkus oleh pangkal pelepah daun. Daunnya berwarna hijau atau hijau muda mengkilat, berbentuk lonjong yang biasanya makin melebar pada ujungnya, tanpa tangkai daun. Susunan bunga bermacam-macam bentuknya ada yang tunggal, tandan atau malai. Marga Phalaenopsis dapat berbunga serentak atau bergantian. Jumlah bunganya dapat sedikit (satu kuntum) hingga banyak (30 kuntum). Ke!opak mahkotanya tidak berlekatan. Seringkali ukuran kelopak dan mahkota hampir sama atau mahkotanya sedikit lebih besar dan lebar. Ukuran bunganya bervariasi dari yang kecil (2-3 cm) hingga besar (9-10 cm). Marga Phalaenopsis umumnya memiliki warna yang mencolok dengan variasi warna putih, merah jambu, ungu, kuning yang dihiasi dengan pola garis-garis, bintik-bintik atau totol-tot01 berwarna merah hati, coklat, merah jambu yang menimbulkan kesan warna kontras. Perhiasan bunga (kelopak dan mahkotanya) sering kali mempunyai p l a clan warna yang sama (Puspitaningtyas dan Mursidawati 1999).
kembali oleh Sweet (1980) melalui beberapa kali tahapan dengan data terbaru (material segar) dan spesimen yang berasal dari berbagai herbarium di seluruh dunia.
Genus Phalaenopsis merupakan satu dari 90 genus dalam sub tribe Sarcanthinae, dengan jumlah kromosom 2n = 38 (de Vogel 1990). Pengelompokan sub tribe Sarcanthinae ke dalam genus-genus didasarkan pada bentuk bagian-bagian atau struktur bunganya yang khas. Genus Arachnis karena bentuk bunganya seperti kalajengking, Renanthera (ren = ginjal dan anthera) karena bentuk anternya seperti ginjal. Genus Phalaenopsis (phalaina = ngengat dan opsis = penampilan) karena bentuknya seperti ngengat, Rhynchostilis (rhynchos = paruh dan stylis = column) karena bentuk column berparuh (Sidran 1985 dikutip Kartikaningrum 2002). Namun Sweet (1980) menyatakan, jika satu kriteria tertentu digunakan
untuk
mengevaluasi pengelompolcan, maka beberapa genus yang berkerabat dekat dapat menjadi terpisah. Persamaan dari genus-genus yang tergolong dalam sub tribe Sarcanthinae adalah pertumbuhannya monopodial, pada umumnya polinarium terdapat stipe dan viscidium,bunga muncul di bagian lateral dari batang tanaman (de Vogel 1990).
Penanda Morfologi dan Penanda Molekuler RAPD
atau karakter molekuler yang dapat didcur dengan pewarisan sederhana d m mengikuti pewarisan Mendel. Penan& molekuler berperan penting dalam program pemuliaan tanaman sebagai sumber genetik dalam bentuk sidik jari Vinger printing) dan sebagi alat seleksi berdasar penanda yang terpaut dengan karakter fenotipik yang dituju.
Beberapa teknik yang digunakan sebagai penanda antara lain RFLP
(Restrrctron Fragment Length Polymorphism), PCR (Polymerase Chain Reaction), SSR (Srmple Sequence Repeat), AFLP (Ampllfed Fragment Length Polymorphism)
dan W D (Randomly Ampllfed Polymorphic DNA). Penanda molekuler melalui teknik RAPD dihasilkan melalui arnplifikasi DNA yang berdasar pada PCR. Hasil reaksi PCR berupa potongan-potongan DNA yang dengan mudah dapat dipisahkan melalui
teknik
elektroforesis yang hasilnya berupa pita-pita DNA dalam berbagaiukuran
(William et al. 1990). Potongm-potongan DNA hasil amplifikasi, masing- masing dapat diperlakukan sebagi karakter untuk keperluan analisis.Teknik
RAPD
telah banyak diaplikasikan sebagai infonnasi genetik, antara lain analisis kekerabatan pada Chicory (Koch dan Jung 1997), keragaman genetik pada anggrek Cymbidium (Obara-Okeyo dan Kako 1998), keragaman genetik kapas (Tatineni et al. 1994), atlaiisis sidik jari dalam pemuliaan anggrek Phalaenopsis (Chenlemon (Deng et al. 1995), dan analisis kekerabatan anggrek sub tribe Sarcanthinae (Kartikaningrum 2002). Selain itu teknik RAPD juga banyak digunakan untuk tujuan tertentu antara lain sebagai proteksi hak paten, pemetaan genetik, diversitas genetik, variabilitas genetik, karakterisasi galur tangkar dalam (inbreed line), taksonomi dan evolusi, variasi populasi genetik (Rajapakse dan Ballard 1997).
Kaitan antara Penanda Morfologi dan Molekuler
Penanda morfolop baik karakter vegetatif maupun generatif ada yang mudah dibedakan, namun sering pula sulit karena sangat dlpengaruhi lingkungan terutama untuk karakter kuantitatif. Pada anggrek tanah Spathoglotis aurea dan S. plicata yang tersebar di Jawa, kedua spesies tersebut sangat mudah dibedakan berdasarkan karakter warna pelepah, warna helaian daun, jarak antar tulang daun, bentuk bulk warna bunga, bentuk bibir, perbuluan
dm
bentuk kalus (Chilanawati 1994). Namun untuk jenis anggrek lainnya yang mempunyai keragaman sangat besar, sangat sukar untuk membedakan dan mengelompokan fenotipe-fenotipe tersebut secara akurat. Oleh karena itu saat ini mulai banyak penelitian yang membandingkan antara kaitan karakter morfologs dengan penanda molekuler (isozim, DNA).Penanda DNA secara genetik terpaut dengan suatu karakter yang diinginkan, yang digunakan untuk kloning gen, diagnosa medis, dan karakter introgresi pacia
disebut penanda RAPD yang berlaku sebagai penanda dominan, yakni dalam populasi yang bersegregasi tidak dapat membedakan .individu homosigot dan heterosigot, karena memberikan hasil pita DNA yang sama.
Dalam pemuliaan, penanda molekuler berperan sebagai penentu identitas atau sidik jari dan sebagai penanda dalam seleksi terpaut dengan sifat fenotipik tertentu. Chen et al. (1995) dalam penelitiannya menunjukkan bahwa primer OPQ103so pb merupakan penanda warm bunga merah berdasarkan analisis Fl hasil persilangan D. pulcherima dan Phalaenopsis equestris. Penanda dalam seleksi terhadap karakter
terpaut dilakukan dengan cara skoring tidak langsung terhadap ada tidaknya fenotip tanaman yang diinginkan atau komponen fenotip yang didasarkan pada pola pita dari
BAHAN DAN METODE
Penelitian ini dilakukan di laboratorium Biologi Molekuler dan Imunologi, Unit Bioteknologi Perkebunan (UPBP), Bogor dari bulan September 2001 sampai bulan April 2002.
Bahan Tanaman
Bahan tanaman yang digunakan dalam penelitian ini adalah 19 spesies anggrek
Phalaenopsis yaitu Phalaenopsis violacea 'Borneo', P. sumatrana, P. kunstleri, P. parrtherina, P. cornu-cervi,
P.
micholitzii,P.
gigantea,P.
lueddemanniana 'pulchra',P. amboinensis 'Ambon', P. parishii, P. celebensis, P. amabilis, P. javanica,
P.
speciosa Tetraspis',
P.
venosa, P. viridis. P. schillerana, P. manii dan P. equestris(Lampiran 1 ).
Bahan Kimia dan Peralatan
Bahan kimia yang digunakan adalah cetyl trimethyl ammonium bromide
(CTAB) 10 %, Tris-HC1 1 M pH 8, NaCl 5 M, larutan etilendiamin tetra asetat (EDTA) 0.5 M pH 8, bufer ekstraksi Orozco-Castillo (Lampiran 2), klorofonn :
bromida, 16 primer, larutan bufer primer, enzim taq DNA polimerase (taq GeneAmp dari Applied Biosystem), larutan stok dNTPs, larutan ion M$+, dan penanda DNA yang dipotong enzim HindZZZ.
Alat yang digunakan adalah PCR Thermolyne-Arnplitron 11, alat elektroforesis, kamera polaroid 665, scanner, transimulafor T 2201, serztrifuRe high sonic Sorvall RC-55 Du-pont, sentrifuge high sonic
MR
1812, inkubator, neraca analitik (4 desimal) Sartorius, oven, microvawe National, Penangas air, pipet mikro Eppendorf berbagai ukuran, pipet mohr, pipet tetes, erlenrneyer, gelas piala, tabung Eppendorf berbagai ukuran, tips Eppendorfberbagai ukuran, mortar,fieezer dan pipet tetes.Metode Penelitian
Isolasi DNA tanaman anggrek genus Phalanopsis dilakukan menurut metode Orozco-Castillo et al. (1994) yang dimodifikasi khususnya penambahan antioksidan polivinilplipirolidon (PVPP) pada waktu penggerusan dan merkaptoetanol yang ditambahkan dalam bufer ekstraksi (Toruan-Matius et al. 1997). Pereaksi yang digunakan disajikan dalam Lampiran 2.
Pelaksanaan Perco baan
Orozco-Castillo et al. (1994) yang sebelumnya telah diinkubasikan pada suhu 65°C selama 30 menit dan diberi 25 p1 merkaptoetanol. Campuran dikocok selama 5 menit dan diinkubasikan pada suhu 65°C selama 30 menit. Kemudian larutan dibiarkan pada suhu kamar selama 5 menit. Untuk mendegradasi protein, ditambahkan 1 ml larutan kloroform : isoamilalkohol (24 : 1) dan dikocok dengan vorteks selama 5 menit, diikuti dengan sentrihgasi dengan kecepatan 11000 rpm selama 10 menit. Cairan bagian atas dipindahkan dengan menggunakan pipet ke dalam tabung Eppendorf yang lain. Pemberian larutan kloroform : isoamilalkohol (24 : 1) dapat diulang kembali dilanjutkan dengan proses sentrifugasi untuk mendapatkan DNA yang murni.
Tahap untuk mendapatkan DNA yang mumi adalah dengan menambahkan 1 ml isopropanol dingin ke dalarn tabung Eppendorfyang berisi larutan DNA, kemudian dikocok perlahan sampai terlihat benang-benang halus, lalu disimpan dalam lemari es
pada suhu 4OC selama 30 menit. Selanjutnya disentrifugasi selama 10 menit dengan
kecepatan 1 1000 rpm untuk mengendapkan DNA. Cairan dibuang kemudian peletnya dicuci dengan alkohol 70% dan dikeringkan 5 menit dengan cara membalikkan tabung. Pelet DNA dilarutkan dengan 500 p1 larutan TE, dikocok perlahan hingga larut dan disimpan difieezer pada suhu -20°C.
Penetapan kuantitas (konsentrasi) dan kualitas DNA dilakukan dengan cara elektroforesis msnurut Sambrook et al. (1989). Besarnya kuantitas DNA dibandingkan dengan besarnya pita DNA lambda yang telah diketahui konsentrasinya yaitu 250, 500, dan 1000 nglpl. Uji kualitas DNA dilakukan dengan menggunakan sebanyak 4
(Lampiran 2), 4 unit 0.3 p1 EcoRI
dan
18.2 p1 aquades steril, kemudian campurn dikocok perlahan-iahan dan diinkubasikan pada suhu 37°C selama 120 menit dalam penangas air. Setelah masa inkubasi berakhir, ke dalam campuran ditambahkan 5 p1 loading buffer. Kuantitas dan kualitas DNA dapat ditentukan dengan cara elektroforesis menggunakan gel agarose 1 % (blv).Pembuatan gel agarose dilakukan dengan melarutkan 0.3 g bubuk agarose dalam 30 ml larutan Tris Asetat (TAE) lX, dan dipanaskan dalarn microwave selama
2 menit dan ke dalamnya ditambahkan pewarna DNA yaitu 1.5 p1 etidium bromida. Campuran tersebut dikocok perlahan dan dituangkan ke dalam cetakan elektroforesis yang telah diberi sisir. Gel didiamkan selama 60 menit supaya padat, kemudian cetakan diletakkan ke dalam bak elektroforesis dan direndam dengan larutan
TAE
lX, kemudian ke &lam l a . tersebut ditambahkan 12 p1 etidium bromida.Elektroforesis dijalankan pada tegangan 50 volt selama 60 menit. Untuk menetapkan konsentrasi clan kualitas DNA, hasil elektroforesis diamati di bawah
UV
transluminator. Konsentrasi DNA ditetapkan berdasarkan ketebalan pita DNA contoh dibandingkan dengan pita marker DNA lamda yang telah diketahui konsentrasinya. Sedangkan kua!itas DNA dapat diketahui dari mampu terpotongnya DNA yang ditunjukkan dengan garis putih tebal (smear) pa& sumur elektroforesis.
Seleksi primer
RAPD
untuk amplifikasi DNAUntuk memperoleh tingkat polimorfisme yang tinggi maka dilakukan tabapan
OperonTechnology (Almaeda, USA) menggunakan contoh DNA spesies Phalaenopsis violacea 'Borneo'. Dari 17 primer tersebut terpilih 9 primer, sedangkan 4 primer mengadopsi dari hasil seleksi yang telah dilakukan pada anggrek sub tribe Sarcanthinae (Kartikaningrum 2002), 1 primer mengadopsi hasil penelitian pada anggrek Vanda (Kardin et al. 1999), 2 primer mengadopsl hasil penelitian
Trichoderma. Berdasarkan informasi dan tahapan seleksi terpilih 16 primer untuk amplifi kasi selanjutnya.
Amplifikasi DNA
Amplifikasi DNA dilakukan menurut metode Williams et al. (1990). Reaksi amplifikasi dilakukan dalam 25pl campuran larutan (Lampiran 2) yang terdiri atas 10
X bufer reaksi (50
m M
KCl; 10m M
Tris-HC1 pH 9.0; 0.1 % Triton X-loo), 0.2 rnMdNTP, 2.5
m M
MgC12, 1 unit Taq DNA polimerase (GenAmp, Applied Biosystem), H20, 50 ng DNA cetakan dan 0.3 p M primer yang dipipet ke dalam tabungEppendorf Untuk menghlndari penguapan yang dapat menyebabkan perubahan konsentrasi selama proses PCR dalam alat Themal Cyler, ke dalam tabung Eppendorf
selama 2 menit, kemudian reaksi diakhiri dengan post extention pada suhu 72°C selama 4 menit.
Produk amplifikasi dipisahkan menggunakan elektroforosis pada 1,4% agarose dengan buffer TAE. Gel diwarnai dengan etidium bromida menurut Sambrook et al.
(1989). Hasil elektroforesis divisualisasikan di atas
UV
transimulator dan didokumentasikan dengan film Polaroid 665 atau dengan menggunakan scanner.Analisis Data
Data molekuler
Hasil analisis RAPD diperoleh &lam bentuk pita-pita hasil pemotretan gel dengan ukuran tertentu dari masing-masing genotipe anggrek. Analisis data berdasarkan ada (1) tidaknya (0) pita yang sarna yang dimiliki secara bersama pada
masing-masing genotipe tanaman yang dibandingkan. Untuk menentukan tingkat kesarnaan pasangan genotipe yang terdapat pada lajur yang berbeda ditentukan berdasarkan rumus Nei dan Li (1979) yaitu :
keterangan : S = koefisien persamaan
a dan b = dua individu yang dibandingkan
flab = jumlah pita DNA yang sarna posisinya baik pada
individu a maupun b
% = jumlah pita DNA pada individu a
Pengelompokan data matriks dan pembuatan dendogram dilakukan dengan metode UPGMA, fungsi SIMQUAL program NTSYS versi 2.02 (Rohlf 1993). Data matriks dihitung melalui koefisien Drce (S), yang pada prinsipnya sama dengan rumus Nei dan Li tersebut di atas. Untuk menghitung koefisien jarak, data h m dikonversikan dengan rurnus d = 1 - S, di mana d adalah jarak dan. S adalah nilai matriks persamaan (koefisien persamaan). Tingkat kepercayaan dari dendrogram yang terbentuk dinilai dengan anslisis nilai kofenetik (Rohlf 1993).
Matriks kemiripan genetik dari data RAPD dianalisis lanjut berdasarkan
Principal Component (komponen utama) yang diturunkan dari matriks varian- kovarian. Skor komponen utama untuk setiap pengamatan dihitung melalui persamaan Yij = ali xij + azi xzj + ...
.
+ +i xpj, dimana Yij adalah skor komponen ke-iuntuk pengarnatan ke-j, aij menunjukkan besarnya kontribusi peubah ke-i terhadap
komponen utarna ke-j dan xij adalah peubah ke-i dengan komponen utama ke-j. Analisis komponen utama dilakukan untuk menentukan pita yang berperan dalam pengelompokkan dan posisi relatif genotipe-genotipe tersebut. Dari pita-pita yang berperan dapat dicari pita yang spesifik untuk genotipe tertentu yang memiliki korelasi yang besar antara setiap pita (peubah ke-i) dengan komponen utama ke-j dengan rumus menurut Dillon dan Goldstein (1984) sebagai berikut:
keterangan : ri : korelasi pita ke-i pada komponen utama ke-j aq : unsur ke-i dari vektor ciri ke-j
h . : akarciri ke-j
-{
Data fenotipik
Berdasarkan kunci determinasi terkini yang disusun oleh Sweet (1980), dibuat dendrogram yamg menunjukkan pengelompokan spesies-spesies yang terrnasuk dalam genus Plzalaenopsis. Pembuatan dendrogram dilakukan dengan dua cara. Cara pertama yaitu langsung menggunakan kunci tersebut untuk mengelompokan spesies-spesies yang diuji menurut seksi dan sub seksinya. Sedangkan cara kedua yaitu dengan mencari karakter fenotipik pernbedanya dan memberikan nilai katagorial / skor karakter tersebut. Penentuan katagori nilai berdasarkan pada beberapa acuan antara lain Bechtel et al (1981), Cameron dan Chase (1999), dan Tatineni et a1 (1996) yang dimodifikasi. Kemudian data katagorial nilai karakter fenotipik tersebut, ditranformasi dengan prosedur standardisasi STAND pada program NTSYS (Numerical Taxonomy and Multivariate Analysis System) versi 2.02, yang pada prinsispnya adalah nilai observasi setiap karakter dikurangi nilai rata-rata karakter tersebut dibagi standar deviasi (Beer et al. 1993; Autrique et al. 1996; Tatineni et al. 1996; Rohlf 1993). Analisis data yang sudah ditransforrnasi, menggunakan fhngsi SIMMZNT pada program NTSYS berdasar koefisien DIST I rata-rata jarak taksonomi :
keterangan : i dan j : dua genotipe yang dibandingkan k : karakter morfologi
X : nilai pengamatan n : jumlah genotipe
E : jarak taksonomi
didasarkan suatu asumsi bahwa karakter tesebut dikendalikan ale1 ganda (multiple alel). Artinya jika gen A bermutasi menjadi a1 atau a2 atau a3 dan seterusnya dengan urutan dominansi tertentu, maka fenotipik dari sel somatik yang terdiri dua seri alel tersebut ditentukan oleh gen yang doininan. Berarti biia suatu karakter morfoiogi telah berperan, maka peranan karakter lain tidak terlihat. Dalam penelitian ini deskripsi karakter pembeda merupakan sub karakter, sehingga banyaknya sub karakter tersebut dianggap sebagai gen yang bermutasi (anggota alel). Sub karakter yang tampak pada genotipe tersebut ditandai dengan nilai 1 dan yang tidak nampak nilainya 0. Selanjutnya data tersebut digabung dengan data RAPD. Analisis gerombol dilakukan seperti pada data genotipik yaitu dengan mctode U P G M , fungsi SIMQUAL dengan koefisien
DICE
pada program NTSYS (Rohlf 1993). Tingkat kepercayaan dari dendrograrn dinilai berdasarkan analisis nilai kofenetik (Rohlf 1 993).Keselarasan antara penanda RAPD dengan fenotipik
Keselarasan antara penanda molekuler dan penanda fenotipik dapat ditinjau berdasarkan matriks rata-rata jarak taksonomi dan matriks kemiripan genetik. Kedua matriks tersebut dibandingkan dengan uji korelasi product-moment dari Pearson fungsi MXCOMP pada program NTSYS (Rohlf 1993). Korelasi antara pasangan dua matriks diuji dengan statistik Z Mantel (Mantel 1967 dikutip Beer et at. 1993) yang dihitung sebagai :
keterangan : Xjk : elemen baris matriks ke j dan kolom ke k dari
,
,
X
Yjk : elemen baris matriks ke
j
dan kolom ke k dari Yjk k < jPada fbngsi MXCOMP, distribusi Z untuk nilai X dan Y tertentu diturunkan dengan menghitung Z
untuk
setiap permutasi matrib yang mungkin sehingga persenti1 Zobservasi dapat diestimasi. Nilai Z kemudian ditrafisformasi melalui normalisasi statistik Mantel yang ekuivalen dengan koeftsien korelasi product- moment Pearson untuk X dan Y (Smouse et al. 1986 dikutip Beer et al. 1993). Dengan rumus :
Z =
X,
Xr
XJ' yjk, keterangan XJ' : hasil transformasi XjkyJk : hasil transformasi Yjk
Jika baik
X
maupun Y berdistribusi normal, dan koefisien korelasi populasi sama dengan 0 maka nilai t = r(n-2)lRI
(l-?)lR berdistribusi sebagai t-Student(Snedecor dan Cochran 1989 dikutip Beer et al. 1993).Hubungan penduga antara
peubah
X dengan Y dinotasikan dengan persamaan regresi linier sederhana (MattJikdan Sumertajaya 2000):
dimana
Hipotesis : Ho : nilai ik tidak berkorelasi dengan nilai
qk
H1 : nilai dk berkorelasi dengan nilaiqk
Dasar pengambilan keputusan :1. Jika probabilitas > 0.05, maka
JJO
diterima Jika probabilitas I 0.05, rnaka & ditolak 2. Jika t hitung < t (n-2; d2), maka HO diterimaKeselarasan pengelompokan ditentukan dari lcriteria goodness of$t Berdasarkan nilai
korelasi menmt Rohlf (1993) yang disajikan pada Tabel 1.
Tabel 1. Kriteria goodness offit berdasarkan nilai korelasi
Level Kriteria g d i ~ e s s offit -- -
0.9 5 r Sangat baik
0.8 5 r < 0.9 Eaik
0.7 5 r < 0.8 Lemah
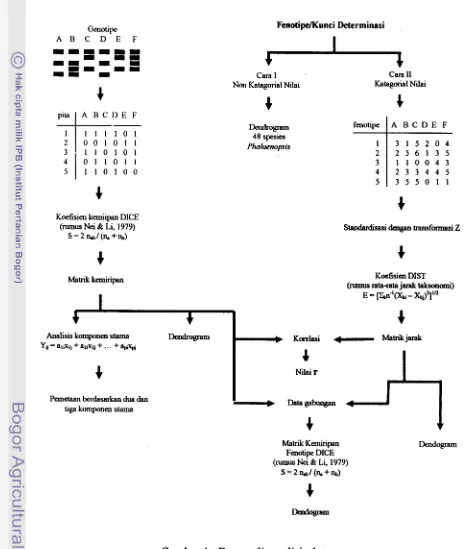
Analisis data tersebut dapat digambarkan seperti tercanturn pada bagan alir berikut (Gambar 1).
Genotipe
A B C D E F
Feaotipe/Kunci Determinasi
[image:161.598.63.539.137.686.2]Cara I Cara I1
Gambar 1. Bagan alir analisis data
Non Katagorial Nilai Katagorial Nilai
+
+
C
D e n d r v
48 s p i e s
0 0 1 0 1 1 Phalaenopsis
1 1 0 1 0 1 O I l O l l
1 1 0 1 0 0 2 3 3 4 4 5
3 5 5 0 1 1
+
Koefisien kemiipan DICE
+
(rumus Nei & Li, 1979) Standardisasi dengan transformasi Z
S=2na/(n.+nb)
+
+
Uatrikkemiripan Koefisien DIST
(rumus rata-tata jarak taksonomi)
I
E = [ml&-
x ~ ) ~ ] ' ~S
.)Analisis kompwen utama D = h P m
Y8=alixlj+az,x2j+... +a,,&,
+
Pemetaan bedasarkan dua dan
+
Korelasi 4- Matrikjarak4
Nilai r
tiga komponen utama III,btagsbungan
+
Matrik Kemiripan Dendogram
Fenotipe DICE (mus N k & Li, 1979)
S=Znal(n.+nb)
HASIL
Tahapan Percobaan untuk Analisis RAPD
Analisis RAPD terdiri atas beberapa tahapan percobaan yaitu : isoiasi DNA genom dilanjutkan dengan pemurnian dan penetapan kuantitas dan kualitas DNA, seleksi primer dekamer acak dan analisis polimorfisme.
isolasi, pemurnian, perretapan kuantitas dan kualitas DNA
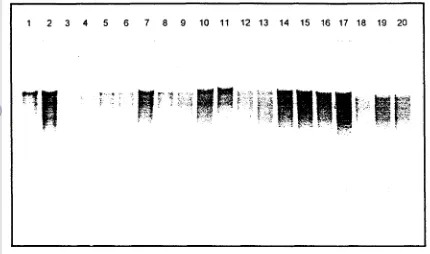
Konsentrasi DNA yang diperoleh yaitu antara antara 75 ngp1 sarnpai 700 ng/pl (Gambar 2, Tabel 2).
Gambar 2. Kuantitas DNA total 20 spesies Phalaenopsis dengan bahan daun (1) DNA h 250 ng/pl, (2) DNA h 500 ndpl, (3) DNA h 1000 ng/pl, (4) P. violacea 'Borneo' (5) P. sumatrana, (6) P. kuntsleri, (7) P. pantherzna, (8) P. cornu-cervi, (9) P. floresensis, (10) P. micholitzii, (1 1) P. gigantea, (12) P. lueddemanniana 'Pulchra', (13) P. arnboinensis 'Ambon', (14) P. parishii, (15) P. celebensis, (16) P. amabilis, (17) P. javanica, (18) P.
[image:162.598.74.542.345.737.2]Tabel 2. Kuantitas
DNA
hasil ekstrabi pada 2 1 spesies anggrek PhaIaenopsisdibandingkan dengan besarnya pita
DNA
lamdaSpesies Tanaman Anggrek Phalaenopsis Konsentrasi DNA (nglpl)
1 . P. violacea 'Borneo'
2. P. sumatrana
3. P. krrnstleri
4. P. pantherina 5 . P. cornri-cervi
6 . P. jloresetds*'
7 . P. micholitzii
8. P. giganlea
9. P.l~~eddemmlnia~a 'Pulchra'
10. P. amboinensis' Ambon'
11. P. p i s h i i
12. P. celebetts.;~
1 3 . P. amabilis
14. P. javanica
15. P. speciosa 'Tetraspis'
1 6. P. zebrinae8'
1 7 . P. venom
18. P. viridis
19. P. schillerana
20. P. manii
2 1. P. equestris
Keterangan : *) spesies tidak disertakan dalam penelitian selanjutnya
Hasil penetapan kualitas
DNA
total hasil ekstraksi menunjukkan bahwa seluruhDNA
contoh yang dianalisis terpotong dengan baik yang ditunjukkan denganDNA
yang smear (Garnbar 3). Uji restriksi merupakan salah satu uji kualitasDNA
Gambar 3. Kualitas DNA total 20 spesies Phalaenopsis dengan digesti EcoRl lajur (I) P. violacea 'Borneo', (2) P. sumatrana, (3) P. kunstleri, (4)
P.
pantherina, (5) P. cornu-cervi, (6) P. jloresensis, (7) P. micholitzii, (8) P. gigantea, (9) P. lueddemanniana 'Pulchra7, (10) P. amboinensis
'Ambon', (1 1) P. parishii, (12) P. celebensis, (13) P. amabilis, (14) P.
javanica, (15) P. speciosa 'Tetraspis', (16) P. zebrinae, (17) P. venosa, (1 8) P. viridis, (19) P. schillerana, (20) P. manii.
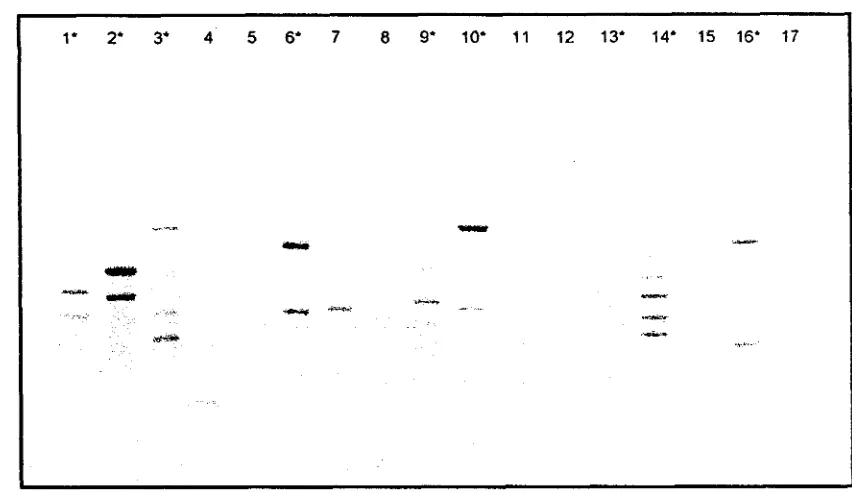
Seleksi primer acak dekamer
[image:164.601.87.517.87.341.2]'Borneo'(Gambar 4), dipilih 9 primer yang menunjukkan polirnorfisme yang tinggi, jurnlah pita yang banyak dan meaghasilkan pita-pita yang tajam, yaitu OPA- 1,OPA-3,
OPA-7, OPA-11, OPA-16, OPB-5, OPB-7, OPB- 10, dan OPB- 17. Dua primer lainnya yaitu OPC-6 clan OPC-9 merigadopsi dan penelitian pa& Triclmderma.
Gambar 4. Tujuh belas primer hasil seleksi menggunakan DNA cetakan P. violuceue
'Borneo';
*
: primer terpilih, lajur (1) OPA 1, (2) OPA 3, (3) OPA 7, (4) OPA 8, (5) OPA 9, (6) OPA 1 1, (7) OPA 12, (8) OPA 14, (9) OPA 16, (1 0) OPB 7, (1 1) OPB 2, (12) OPB 3, (13) OPB 5, (14) OPB 10, (15) OPB 16, (16) OPB 17, (17) OPB 19.Seluruh primer yang diseleksi dapat mengamplifikasi DNA genom tanaman P.
[image:165.603.90.516.196.443.2]tunggal adalah kedua utas DNA mempunyai sekuen komplemen dengan primer yang digunakan. Ketajaman pita dapat ditentukan oleh banyaknya fiagmen DNA yang teramplifikasi, makin banyak maka resolusi pita DNA yang dihasilkan makin jelas dan sebaliknya makin sedikit menghasilkan pita yang kabur. Menurut Weising et at.
[image:166.598.76.518.363.683.2](1995) pada genom tanaman lebih kurang 90 % dari DNA genom merupakan urutan yang berulang. Selain itu adanya kompetisi situs penempelan primer pada DNA genom menyebabkan suatu fragmen a k a . diamplifikasi dalam jurnlah banyak dan fragmen lain dalam jumlah yang sedikit.
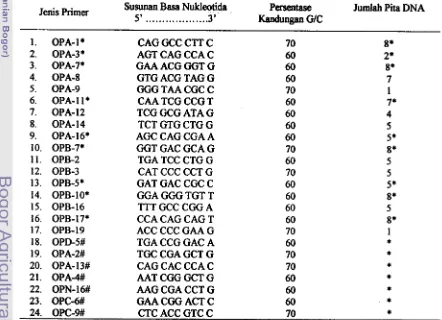
Tabel 3. Jenis primer, susunan basa nukleotida, persentase kandungan GIC dan jumlah pita dari primer yang diseleksi
Jenis Primer Susunan Basa Nukle~tida Pexsentase Jumlah Pita DNA 5'
. .
.
.
. . .
. . .
.3' Kanduagan WCOPA- 1
*
CAG GCC CTT C 70 OPA-3*
AGT CAG CCA C 60 OPA-7* GAA ACG GGT G 60OPA-8 GTG ACG TAG G 60
OPA-9 GGGTAACGCC 70 OPA- 1 1
*
CAA TCG CCG T 60OPA-12 TCG GCG ATA G 60
OPA- 1 4 TCT GTG CTG G 60 OPA- 1 6* AGC CAG CGA A 60
OPB-7* GGT GAC GCA G 70 OPB-2 TGA TCC CTG G 60
OPB-3 CAT CCC CCT G 70 OPB-5
*
GAT GAC CGC C 60OPB- lo* GGA GGG TGT T 60 OPB- 16 TTT GCC CGG A 60
OPB-17* CCA CAG CAG T 60 OPB- 19 ACC CCC GAA G 70 OPD-5# TGA CCG GAC A 60 OPA-2# TGC CGA GCT G 70 OPA- 13# CAG CAC CCA C 70 OPA-4# AAT CGG GCT G 60
OPN- 1 6# AAG CGA CCT G 60
OPG6# GAA CGG ACT C 60 OX-9# CTC ACC GTC C 70
Analisis Polimorfime dengan RAPD
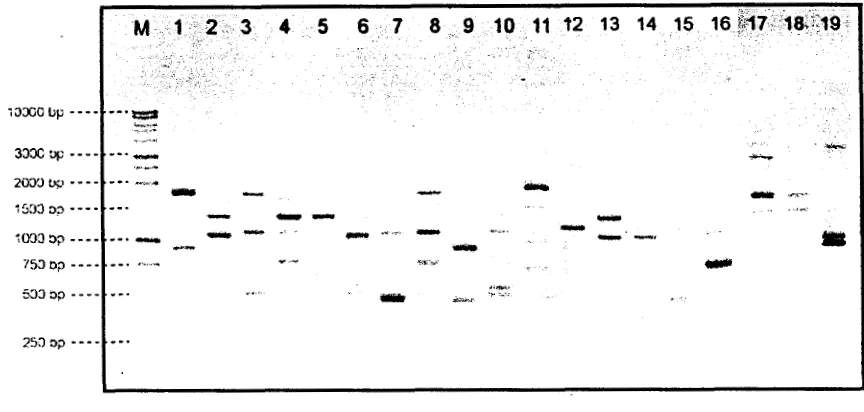
Enarn belas primer yang digunakan pada amplifikasi 19 genotipe Phalaenopsis
menghasilkan 13
-
24 pita (Tabel 4), dengan total pita yang dihasilkan 300 pita dengan ukuran fragrnen berkisar antara 250 pb-
3000 pb. Seluruh pita yang dihasiikan menunjukkan polimorfisme. Salah satu contoh profil pita hasil amplifikasi dengan primer OPA 11 disajikan pada Gambar 5.Tabel 4. Jenis primer, susunan basa nukleotida, persentase kandungan G/C dan jumlah pita hasil amplifikasi 19 spesies Phalaenopsis
Jenis Primer Susunan Basa Nukleotida Persentasc Jurnlah Pita Hasii 5' . . .
.. .
..
. ..
. . . .3' Kandungan G/C AmplifikasiOPA-1 OPA-2 OPA-3 OPA-4 OPA-7 OPA-11 OPA- 13 OPA-16 OPB-5 OPB-7 OPB- 10 OPB- 17 OPC-6 OPC-9 OPD-5 OPN-16
CAG GCC CTT C TGC CGA GCT G
AGT CAG CCA C AAT CGG GCT G GAA ACG GGT G CAA TCG CCG T CAG CAC CCA C AGC CAG CGA A
TGC GCC CTT C GGT GAC GCA G GGA GGG TGT T CCA CAG CAG T GAA CGG ACT C CTCACCGTCC TGA GCG GAC A AAG CGA CCT G
Gambar 5. Profil pita DNA 19 spesies Phalaenopsis hasil amplfikasi menggunakan primer OPA 11; lajur (M) 1 kb DNA Ladder, (1) P. violacea 'Borneo', (2)
P. sumatrana, (3) P. kunstleri, (4) P. parzterina, (5) P. cornucervi, (6) P. micholitzii, (7) P. gigantea, (8) P. lueddemanniana 'Pulchra', (9) P.
amboinensis 'Ambon', (10) P. parishii, (1 1) P. celebensis, (12) P. amabilis, (13) P. jmanica, (14) P. speciosa 'Tetraspis', (1 5) P. venosa, (16) P. viridis, (17) P. schillerana, (18)
P.
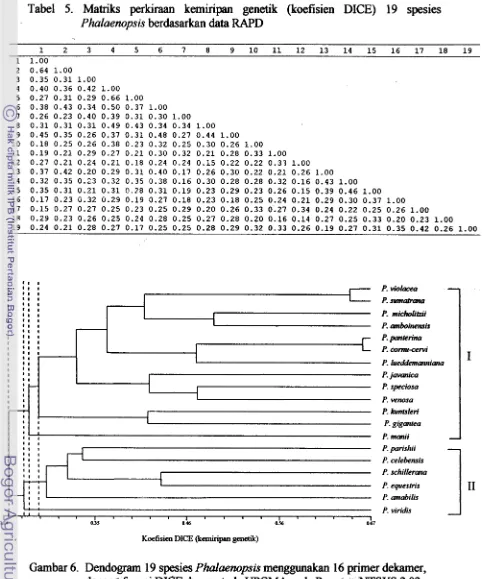
manii, (19) P. equestris.Hasil analisis kemiripan genetik menunjukkan bahwa 19 spesies tersebut mengelompok dengan tingkat kemiripan antara 24 sampai dengan 66 % (Gambar 6). Dendrogram tersebut telah diuji nilai goodness of fit (kebaikan suai) dengan uji kofenetik dan menunjukkan nilai korelasi kofenetik (Cophenetic value) sebesar 80 %
Tabel 5. Matriks perkiraan kemiripan genetik (koefisien DICE) 19 spesies
Phalaenopsis berdasarkan data RAPD
Koefisien DICE &emkipan genetik)
P. violocea
P. nmralraw P. micholihii
P. amboinemis P. panterina
P. comu-cewi
P. lueddemmmiana I
P. jovrmica
P. speciosa
P. m o s a P. h t r b r i
P. gigantea
P. manii
P. parishii
P. celebemis
P. schillerana
P. equeslris
P. amabilis
P. viridis
[image:169.598.71.553.84.663.2]Analisis pengelompokan juga menunjukkan beberapa spesies dalarn satu seksi mengelompok membentuk satu garpu dengan koefisien kemiripan berturut-turut 66 %, 65 %, dan 48 % yaitu P. pantherina dan P. cornu-cervi (seksi Polychillos), P. violacea
'Borneo' dan
P.
sumatrana (seksi Zebrinae), dan P. micholitzii dan P. amboinensis'Ambn7 (seksi ambinensis). Sedangkan pada tingkat kemiripan sekitar 47 % P.
kunstleri dan P. gigantea membentuk satu garpu, padahal dua spesies tersebut berasal dari dua seksi yang berbeda.
Pada tingkat kemiripan 30 % spesies-spesies dari seksi Parishianae, Stauroglotis dan Phalaenopsis membentuk satu garpu yaitu F. parishii, P. celebensis, P. schillerana, P. equestris dan P. amabilis. Dm spesies yang termasuk seksi Fuscatae yaitu P. kunstleri
dan
P. viridis terpisah dari dua kelompok utama tersebut.Sedangkan
spesies-spesies lainnya membentuk kelompok dengan tingkat kemiripan yang kecil. Tingkat kemiripan yang kecil atau polimorfisme yang tinggi tersebut menghasilkan pengelompokan yang tidak sepenuhnya selaras dengan berdasarkan fenotipenya.
(ii)
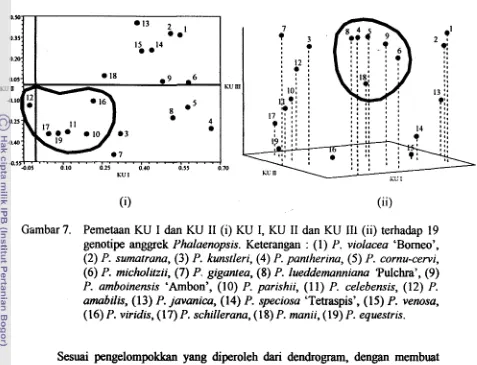
Gambar 7. Pemetaan KU I dan KU I1 (i) KU I, KU I1 dan KU I11 (ii) terhadap 19 genotipe anggrek Phalaenopsis. Keterangan : (1) P. violacea 'Borneo',
( 2 ) P. sumatrana, ( 3 ) P. kunstleri, ( 4 ) P. pantherina, ( 5 )
P.
cornu-cervi, ( 6 ) P. micholitzii, (7) P. gigantea, ( 8 ) P. lueddemanniana 'Pulchra', (9) P. amboinensis 'Arnbon', (10) P. parishii, (1 1) P. celebensis, (12) P. amabilis, (13) P. javanica, (14) P. speciosa 'Tetraspis', (15) P. venosa,(16) P. viridis, (17) P. schillerana, (18) P. manii, (19) P. equestris.
Sesuai pengelompokkan yang diperoleh dari dendrogram, dengan membuat pemetaan antara KU I dan KU I1 diperoleh tiga kelompok yang berada pada tiga kuadran yang berbeda. Kelompok pertarna terdiri atas satu spesies yaitu P. amabilis,
[image:171.601.78.555.75.440.2](seksi Amboinensis), dan P. lueddemanniana 'pulchra' (seksi Zebrinae). Enam spesies tersebut mempunyai pita OPM7@ yang berkorelasi dengan KU I1 sebesar 0.82. Analisis komponen utama yang diterapkan dalam penelitian ini menghasilkan 11 komponen utarna yang mempunyai eigenvalue (akar ciri) lebih dari satu (Dillon dan Goldstein 1984; Shanna 1996) dan mempunyai persentase keragaman kumulatif yang menerangkan keragaman total
data
minimum 75 %. Total 11 KU tersebut menerangkan keragaman data asal sebesar 76, 83 %. Nilai masing-masing KU, akar ciri, proporsi keragamandata
asal dan proporsi kumulatif disajikan pada Lampiran 6. Besarnya keragaman yang dapat diterangkan oleh komponen utarna tersebut sesuai dengan kecilnya tingkat kemiripan antar genotipe yang diarnati yaitu 24 % (Gambar5). Artinya 76.83 % kemgaman dari karakter pita RAPD terhadap 19 genotipe anggrek Phalaenopsis tersebut dapat diterangkan oleh 1 1 KU.
Berdasarkan nilai komponen utarna dari 300 pita hasil amplifikasi 16 primer &pat diidentiwi 231 pita
RAPD
yang berperan dalam mengelompokan secara terpisah 19 genotipe anggrek Phalaenopsis (Lampiran 6). Berclasarkan perhitungan nilai korelasi antara pita dengan komponen utama, diperoleh nilai r yang besar (>0.80) yaitu OPA3880 berkorelasi dengan komponen utama ke-1 (KU-1) sebesar 0.974.
dilakukan persilangan terkontrol untuk mengetahui korelasi suatu pita den* karakter tertentu.
Pengelompokan Berdasarkan Kunci Determinasi Pengelompokan cara-1 Non katagorial nilai
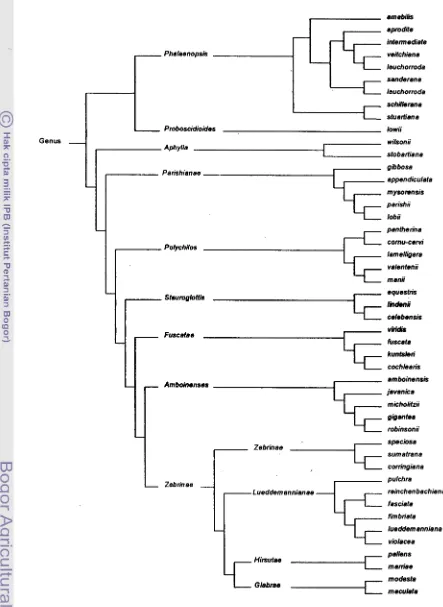
Herman Sweet (1980) menggarap reklasifikasi genus Phalaenopsis dimulai sejak tahun 1968, dengan menggunakan ratusan spesimen dari herbarium di seluruh dunia dan material hiduplsegar. Terdapat 48 spesies Phalaenopsis yang diklasifikasi ulang dan dalam penelitian ini digunakan sebagai data skunder untuk dikelompokan dengan menggunakan cara ke-1 (Gambar 8). Pengelompokan tanpa pemberian katagorial nilai menghasilkan dendrogram yang dibuat secara manual tanpa informasi perkiraan koefisien jarak genotipenya Pengelompokan tersebut merupakan visualisasi kunci detenninasi
dari
Sweet, berturut-turut berdasarkan deskripsi perbedaan karakterpada 9 seksi, 4 sub seksi Zebrinae clan spies-spesies anggotanya.
Pengelompokan spesies-spies pada genus Phalaenopsis didasarkan terutama pada karakter fenotipik bunga. Genus Phalaenopsis dibagi menjadi sembilan seksi yaitu seksi Phalaenopsis, Proboscidioides, Aphyllae, Parishianae, Polychilos, Stauroglottis, Fuscatae, Amboinenses, dm seksi Zebrinae yang dibagi lagi menjadi empat sub seksi yaitu sub seksi Zebrinae, Lueddemannianae, Hirsutae dan Glabrae. Kunci determinasi menuju seksi ditekankan pada karakter fenotipik pembeda yang menonjol yang merupakan ciri
mum
pada seksi tersebut. Seksi Phalaenopsis dicirikanujung column, dan bentuk bagian tengah bibir terbelah dua atau terbelah menjadi dua tendril. Seksi Proboscidioides terutama dicirikan adanya perpanjangan pada ujung tugu yang menyerupai belalai gajah (proboscis). Seksi Aphyllae ditandai dengan tidak berkembangnya pertumbuhan daun (rudimenter). Seksi Parishianae mempunyai ciri adanya pangkal column pada dasar petal dan bagian tengah bibir tidak menyatu. Seksi Polychilos dicirikan dengan bentuk bibir tengah yang menyerupai jangkar. Seksi Stauroglot!is ditandai dengan adanya tonjolan I kalus berbentuk perisai pada persimpangan bibir bagian tengah dan bibir lateral, dan tidak adanya bagian yang menonjol dan menebal di antara bibir lateral. Seksi Fuscatae dicirikan oleh bagian tengah bibir cekung dengan lunas (keel) yang berdaging tebal pada tengahnya Seksi Amboinenses dicirikan mempunyai bentuk bunga bulat, petal bunga elip meluas den- panjang kurang dari duakali lebarnya. Sedangkan seksi Zebrinae mempunyai bentuk bunga menyerupai bintang, petal bunga sempit berbentuk bulat telur terbalik
Genus
eprudite intennediate
-
Phalaenopsis veitchianaleuchormda sanderana
leuchorroda <