FILOGEOGRAFD INTWASPES1
ES GELATIK
( P a d d a oryzivora
(L.))
(PASSERIFORYES: PLOCEIDAE)
01
PEILAU BALI
1'Oleh
S A N G P U T U KAhER S U R A T A
PROGRAM PASCASARJANA INSTITLIT PERTANlAtd BOGOR
FILOGEOGRAFI
INTRASPESIES
GELATIK
(Pa&
olyzzvora
(L.))(PASSERIFORMES:
PLOCEIDAE)
DI PULAU
BALI
SANG PUTU KALER SURATA, Filogeografi intraspesies gelatik (Pacida olyzivora
(L.)) (~asseriformes: Ploceidae) di Pulau Bali (dibimbing oleh Prof Dr NAWANGSARI
SUGIRI (Ketua), Prof Dr R E M
WIDJAJAKUSUMA,
Dr AHMAD ANSORIMATTJIK, Dr DEDY DURYADI dan Dr ASEP S. ADHMERANA sebagai anggota)
Pada masa yang akan datang dikhawatirkan gelatik (Padda oryzivora (L.)) di Bali
mengalami ancaman kepunahan. Oleh karena itu, upaya-upaya untuk mencegah gelatik
dari ancaman kepunahan hams diprioritaslcan. Akan tetapi sebelum upaya tersebut
dilakukan, maka unit-unit konservasi perlu diidentifikasi terlebih dahulu.
Penelitian ini dilakukan dengan maksud mempelajari pola-pola filogeografi gelatik
di Bali. Sebanyak tiga penanda (morfometri, gambaran darah dan genetik) digunakan
untuk mengidentifikasi unit konservasi gelatik. Selain itu, penelitian ini juga mencakup
profil vegetasi dan avifauna dari kawasan tempat mencari makan gelatik di Bali.
Tujuan dari penelitian ini adalah ( i ) inventarisasi profil vegetasi, avifauna dan
kelimpahan gelatik pada beberapa kawasan tempat mencari makan burung tersebut; (ii)
membandingkan pola filogeografi gelatik berdasarkan karakter morfometri, gambaran
darah dan genetik; (iir) memperoleh penanda yang tepat dalam penetapan unit
konservasi gelatik.
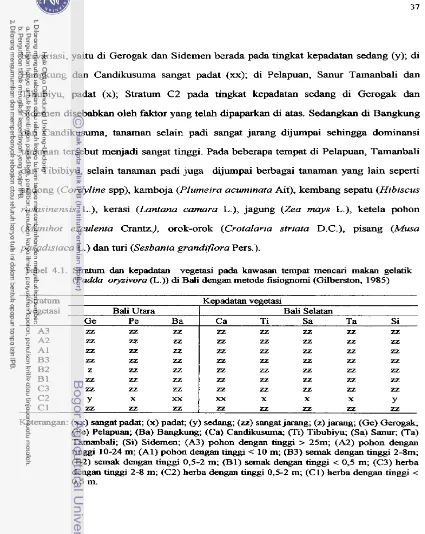
Stratum dan kelimpahan vegetasi diamati dengan metode fisiognomi. Keragaman
avifauna diamati dengan metode garis transek dengan teknik pengamatan garis tunggal.
Tanita, (ii) panjang total dan panjang ekor dengan memakai penggaris; serta ( i i i )
panjang pamh, lebar paruh, kedalaman bukaan paruh, panjang sayap dan panjang tarsus
dengan menggunakan kaliper. Analisis gambaran darah mencakup penentuan (i)
hematoluit dengan teknik mikrohematokrit; (ii) jumlah eritrosit dan leukosit dengan
hemocyiorneter; dan (iii) penghitungan diferensial Ieukosit dengan metode hapusan
darah. Sedangkan pengamatan genetik dilakukan dengan analisis DNA total berdasarkan
teknik RAPD (Random Amplified Polymorphic DNAs).
Jurnlah sampel yang digunakan untuk analisis morfometri, gambaran darah dan
genetik adalah 15-20% dari kelimpahan gelatik yang terdapat pada setiap kawasan
tempat mencari makan burung tersebut. Sebagai outgroup lgunakan gelatik yang
berasal dari Jawa Barat (lima ekor) dan Jawa Timur (enam ekor). Berdasarkan adanya
barrier fisik yang memisahkan Jawa Barat, Jawa Timur, Bali Utara dan Bali Selatan,
populasi gelatik diasumsikan terpisah menjadi empat subpopulasi, yaitu subpopulasi
gelatik Jawa Barat, Jawa Timur, Bali Utara dan Bali Selatan.
Dari hasil pengarnatan teridentifikasi sebanyak delapan kawasan tempat mencari
makan gelatik di Bali, yaitu Gerogak, Pelapuan, Bangkung, Candikusuma, Tibubiyu,
Sanur, Tarnanbali dan Sidemen. Semua kawasan tersebut didominansi oleh vegetasi
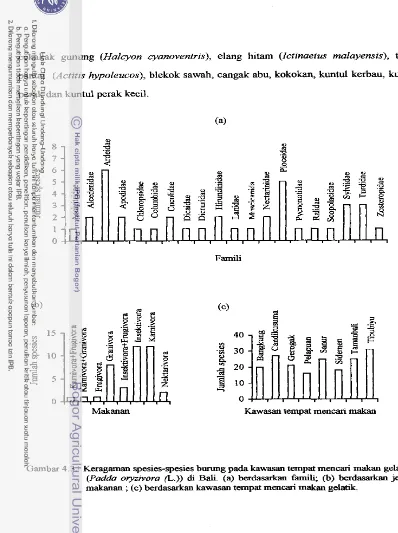
herba (55-84% dari luas total) dengan tinggi 0,5-2 m. Jurnlah spesies burung yang
ditemukan sebanyak 39 spesies, yang tergolong dalam 7 ordo dan 20 famili. Kelimpahan
avifauna pada kawasan tempat mencari makan gelatik didominansi oleh kelompok
burung granivora (86,52%). Sedangkan kelimpahan gelatik hanya 5,3396, jauh lebih
kecil dibanding bond01 dada sisik (Lonchura puncfulata) (34,52%). Hasil analisis
kawasan tempat mencari makannya sendiri. Diduga gelatik mengalami tekanan populasi
akibat kalah bersaing dalam memperebutkan sumber makanan, terutarna dengan bond01
dada sisik.
Gelatik asal Bali memiliki rataan bobot badan 24,88+.1,78 g, panjang total
146f2,63 mm, panjang paruh 15,59+0,45 mm, lebar paruh 8,91+0,37 mm, kedalaman
bukaan pamh 12,93M,29 mm, panjang sayap 66,87*1,87 mm, panjang tarsus
18,5 1+0,55 mm dan panjang ekor 48,9232,03 mm. Hasil analisis komponen utama dan
analisis kelompok menunjukkan tidak terdapat hubungan antara keragaman morfometri
gelatik dan distribusi geografi burung tersebut.
Jumlah eritrosit gelatik asal Bali 3,13+0,66 juta/mm3, hematohit: 62,33+ 12,07 %/
mrn3 dan leukosit 22,40kO,77 ribu/mm3. Sedangkan jumlah basofil 0,06*,24%,
eosinofil 0,91+1,33%, heterofil 38,18+15,48%, limfosit 57,21+16,05% dan monosit
3,09f3,35% dari total leukosit. Hasil analisis komponen utama dan analisis kelompok
menunjukkan tidak terdapat hubungan antara keragarnan gambaran darah gelatik dan
distribusi geografi burung tersebut.
Profil DNA RAPD gelatik menunjukkan adanya struktur populasi yang berbeda
antara gelatik asal Jawa Barat, Jawa Timur, Bali Utara dan Bali Selatan. Hal itu
didukung pula dari hasil analisis komponen utama dan analisis kelompok. Kedua hasil
analisis itu menunjukkan terdapat hubungan antara keragaman genetik gelatik dan
distribusi geografi burung tersebut. Oleh karena itu pengelolaan gelatik yang berasal dari
Bali Utara clan Bali Selatan dalam dua unit konservasi yang saling terpisah perlu
INTRASPECIFIC PHYLOGEOGRAPHY OF
JAVA
SPARROW (Padda O ~ ~ Z ~ V O ~ Q (L.)) (PASSER1IFORIMES: PLOCEIDAE) ON BALK ISLANDSANG PUTU KALER SURATA, Intraspecific phylogeography of Java Sparrow (Padda
oryzzvora (L.)) (Passeriformes: Ploceidae) on Bali Island (Under the supervision of
NAWANGSARI SUGIIU, REVIANY WIDJAJAKUSUMA, AEPvlAD ANSORI
MATTJIK, DEDY DURYADI
AND
ASEP S. ADHIKERANA)The population of Java Sparrow (Padda oryzivora (L.)) on Bali Island is
threatened by the risk of extinction in the near future. The conservation of this
population might provide a solution for its fizture survival. However, before conservation
will be applied, the potential units of conservation must be identified earlier. Hence, the
main purpose of this study is to clarify phylogeographical patterns of Java Sparrow
based on morphometrical, hematological and genetical markers. The study includes also
avifauna diversity and vegetation profile of Java Sparrow feeding areas in Bali.
The objectives of this study are ( 1 ) to investigate habitat profile, avifauna diversity
and the abundance of Java Sparrows in their feeding areas; (2) to compare
morphometrical, hematological and genetical variations, (3) to find markers for
identifying conservation units of Java Sparrows.
Physiognomical survey method was used for recording data on vegetation
stratification and abundance. Avifauna diversity was observed using the line kransect.
Eight morphological measurements were taken consistently on body weight, total length,
bill length, bill width, bill depth, wing Ien& tzrsal length and tail length.
Standard haemotological assay procedures were utilized in determining packed
count for each sample. The genetical character was observed by DNA RAPD (random
amplified polymorphic DNA's) analyses.
About 20% of Java Sparrows from each feeding area were collected for
morphometrical, hematological and genetical analyses. Five birds from West Java and
six birds fiom East Java were used as outgroups. Based on physical barriers among West
Java, East Java, North Bali, and South Bali, the population of Java Sparrow is assumed
to be separated into four subpopulations.
The results indicated that there are eight feeding areas of Java Sparrows on Bali
Island. All of these are dominated by herbs (0,5-2 m), covering 55 to 84% of the total
area. Thirty-nine species of birds, including 7 orders and 20 families were observed. The
abundance of avifauna are dominated by granivorous birds (86,52%). However the
abundance of Java Sparrow is only 5,33%, or about one-sixth from the total of Scaly-
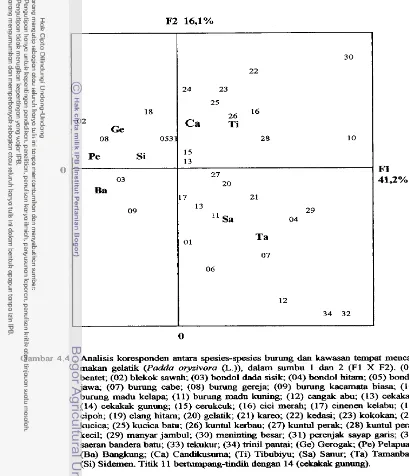
breasted Munia (Lonchura puncrulata). The result of correspondence analysis did not
show any relationship between Java Sparrows and their feeding areas.
The average body weight of Java Sparrows is about 24.88+1.78 g, total length
1 4 6 s . 6 3 mm, bill length 15.59M.45 mm, bill width 8.91&0.37 mm, bill depth
12.93H.29 mm, wing length 66.87&1.87 mm, tarsal length 18.51M.55 rnm and tail
length 48.92f2.03 mm. There is no correlation between geographical distribution and
morphometrical variations, both from the results of principal component analysis and
cluster analysis.
The average leukocyte of Java Sparrow is about 22.40M.77 thousand/mm3,
erythrocyte 3.13-10.66 million/mm3, packed cell volume 62.32f12.07%, basophil
0.06B,24%, heterophil 38.18&15.48%, lymphocyte 57.21f 16.01°h, monocyte
cluster analysis, no correlations between geographical distribution and haematological
variations were observed.
The estimate of genetic diversity (N) among Java Sparrow subpopulations ranged
from 0.174 to 0.294. The RAPD allele frequencies data (Fst>0.25) revealed high
differentiation among subpopulations. These indicated, that Java Sparrow belong to
large ancestral population. A remarkable degree of concordance was found between the
geographic distribution and the DNA polymorphism. The population of Java Sparrow
was separated into four clearly distinct lineage's, namely North Bali, South Bali, East
Java and West Java subpopulations. The potential conservation units could be deduced
from these results, and a management policy could consequently be inferred. This study
clearly indicates the relevance of the genetical approach for the identification of
FILOGEOGRAFI INTRASPESIES GELATm (Padda oryzivora (L.)) (PASSERIFORMES: PLOCEWAE) DI PULAU BALI
Oleh
SANG PUTU KALER SURATA BIO 965081
Disertasi
Sebagai Salah Satu Syarat untuk Memperoleh Gelar Doktor
pada Program Pascasarjana Institut Pertanian Bogor
PROGRAM PASCASARJANA INSTITUT PERTANLAN BOGOR
Judul Disertasi : FlLOGEOGRAFI TNTRASPESIES GELATIK (Padda oryzrvora (L.)) (PASSERIFORMES: PLOCEIDAE) D'.
PULAU BALI
Nama Mahasiswa : SANG PUTU KALER SURATA
Nomor Pokok 965081
Program Studi BIOLOGI
Menyetujui 1. Komisi Pembirnbing
,
<
(Prof Dr Nawangsari Sugiri) Ketua
(Prof Dr ~ e v i a h ~ Widjajakusuma) (Dr Ir Ahmad Ansori Mattjik)
(Dr IT Dedy Duryadi) Anggota
(Dr Asep S. Adhikerana) Anggota - -
2. Ketua Program Studi Biologi gram Pascasaqana
A
(Dr Ir Dede Setiadi)Sang Putu Kaler Surata dilahirkan di Bangli, Bali pada tanggal 24 Pebruari 1959
sebagai anak pertama dari lima bersaudara dari Ibu Sang Ayu Kompyang Tunjung dan
Ayah Sang Made Ratep. Penulis menyelesaikan pendidikan SD tahun 1972 di SDN 2
Bangli, S mtahun 1975 di SMPN Bangli dan Sh4TA tahun 1979 di SM!3 Saraswati
Denpasar, Bali.
Pada tahun 1985, penulis memperoleh gelar sarjana pendidikan biologi pada
Fakultas Keguruan dan Ilmu Pendidikan, Universitas Mahasaraswati Denpasar.
Sedangkan gelar magister sains dalam bidang ilmu pengelolaan sumberdaya alarn dan
lingkungan pada Program Pascasarjana Institut Pertanian Bogor, berhasil diraih pada
tahun 1993. Mulai tahun 1996 penuiis mendapat beasiswa dari Departemen Pendidikan
dan Kebudayaan melalui program beasiswa pendidikan program pascasaqana (BPPS)
untuk mengikuti pendidikan program doktor pada Program Studi Biologi, Program
Pascasarjana Institut Pertanian Bogor.
Sejak tahun 1979 sampai tahun 1983, penulis bekerja sebagai asisten apoteker di
Apotik Kosala Fanna Denpasar. Mulai tahun 1983 sampai 1990, penulis bertugas
sebagai guru honor di SLUA (SMA) Saraswati 2 dan 4 Denpasar, serta di SMF
Saraswati Denpasar. Sedangkan mulai tahun 1989 sampai sekarang penulis bekerja
sebagai staf pengajar pada Fakultas Pendidikan Matematika clan Ilmu Pengetahuan
Alam, Institut Keguruan dan Ilmu Pendidikan Saraswati Tabanan, Bali.
Pada tanggal 15 Januari 1988, penulis menikah dengan Ni Wayan Kari, dan
dikaruniai dua orang anak. Anak yang sulung bernama Sang Putu Arik Priastawan (12
tahun). Sedangkan anak yang bungsu bemama Sang Ayu Made Ika Utari Dewi (9
UCAPAN TERIlMA KASIR
Apabila laporan penelitian ini dapat terselesaikan itu hanyalah berkat rakhmat Ida
Shang Hyang Widhi Wasa atau Tuhan Yang Maha Esa. Untuk itu puji syukur penulis
panjatkan kehadapanNYA. Penulis sangat menyadari bahwa segala usaha dan upaya
yang telah dilakukan adalah berkat bantuan langsung maupun tidak langsung dari
berbagai p i M . Semoga Tuhan Yang Maha Esa memberi imbalan yang sepadan atas
berbagai jasa yang telah diberikan.
Melalui kesempatan ini penulis menyampaikan penghargaan dan terima kasih
kepada yang terhonnat Ibu Prof Dr Nawangsari Sugiri atas kesediaan beliau menjadi
ketua komisi pembimbing. Penghargaan clan ucapan terima kasih yang tulus j,uga
disampaikan kepada Ibu Prof Dr Reviany Widajakusurna, Bapak Dr Ahmad Ansori
Mattjik, Bapak Dr Dedy Duryadi dan Bapak Dr Asep S. Adhikerana atas kesediaan
beliau menjadi anggota komisi pembimbing. Bimbingan dan saran yang diberikan
sangat membantu penulis dalam merurnuskan dan rnenajamkan pemikiran. Tidak kecil
peranan Bapak Dr Dedy Duryadi yang ikut membantu dalam pengadaan material dan
fasilitas penelitian sehingga mernperlancar penelitian clan penulisan disertasi ini.
Ucapan terima kasih disampaikan kepada Bapak Prof Dr S. Somadikarta dan Ibu
Dr Ir Ani Mardiastuti Pakpahan, MSc. atas kesediaan beliau berdua untuk m e n j d
penguji di luar komisi pembimbing.
Penulis juga mengucapkan terima kasih kepada Bapak Koordinator Kopertis
Wilayah VIII, Bapak Ketua Yayasan Perguruan Rakyat Saraswati Tabanan, dan Bapak
Rektor Institut Keguruan dan Ilmu Pendidikan Saraswati Tabanan, atas ijin dan
kesempatan yang diberikan kepada penulis untuk mengikuti pendi&kan program doktor
(S3) di TPB. Ucapan senada disampaikan pula kepada Ibu Direktur Program
xii
IPB dan Pengelola BPPS Direbur Jenderal Perguruan Tinggi Departemen Pendidikan
dan Kebudayaan Republik Indonesia, atas kesempatan dan dukungan dana yang telah
diberikan sehingga proses penyelesaian studi penulis dapat be jalan dengan lancar.
Melalui kesempatan ini, secara khusus penulis sampaikan terima kasih yang tulus
kepada Ibu Dr Sukarti Moeljoprawiro, M.App.Sc (Fakultas Biologi UGM) atas
ketulusan beliau memberikan bantuan enzim Tag polimerase. Kepada Bapak Ir Bagus
Ketut Lodji, MS, Bapak Ir Ahmad Faradjallah MS, Ibu
Ir
Ani Aryani (Lab ZoologiFMIPA Biotrop), Bapak Ir Wahyu Wododo dan Bapak Drs Daryono (Puslitbang
Zoologi LIPI), penulis ucapkan terima kasih yang tulus atas ban- yang diberikan
selama penelitian.
Kepada Dr Ir Made Yasa, MS (alrnarhum); Dr I Wayan Supartha MS; Drs Ketut
Junitha, MS, Drh I Ketut Suatha, Msi; Ir I Gede Wijana, MS; Ir I Gusti Komang Dana
Arsana, Ir Ida Bagus Suryawan, MSi; Ir Komang Puma, MSi; Ir Nengah Suarya, MSi; Ir
Nyoman Wijaya, MS; Ir Komang Sukarsa; Drh Nyoman Suarta, MS1; Drs I Made Sara
Wijana, MSi; Ir I Gde Suranjaya, MSi; Ir Anak Agung Sugiarta, Msi; Ir I Wayan Alit
Artha Wiguna, Msi dan beserta seluruh anggota Himpunan Mahasiswa Pascasarjana
(Punhawacana) Bali-IPB, penulis mengucapkan terima kasih yang setulus-tulusnya atas
dorongan dan dukungannya selama penulis berada di Bogor.
Akhimya, ucapan terima kasih disarnpaikan kepada seluruh keluarga, ayah, iby
adik, ipar, istri dan anak-anak tercinta yang telah memberi ban- talc ternilai baik
moral maupun material selama penulis mengikuti studi di IPB, Bogor. Semoga Tuhan
Yang Maha Esa memberikan rakhmatNYA kepada semuanya
Bogor, Awal Mei 2000
DAFTAR iSI
... DAFTAR TABEL
DAFTAR G A M B M ...
BAB 1 . PENDAHULUAN ...
Latar Belakang ... . .
... Tujuan Penelltian
. .
Manfaat Penelltian ...
... .
BAB I1 TINJAUAN PUSTAKA
... Filogeografi Intraspesies
... Nama dan Sistimatika
... Distribusi dan Habitat
Kawasan Tempat Mencari Makan Gelatik ...
... Isolasi Populasi
... Keragaman Geografi Gelatik
Morfologi Gelatik ...
Darah Burung ...
... Darah sebagai Surnber DNA
Garnbaran Darah ...
Analisis Genetik ...
Amplifikasi DNA dengan Primer Acak ...
BAB I11 . BAHAN DAN METODE ...
Ruang Lingkup ...
Tempat dan Waktu ...
... Kelimpahan dan Sarnpel
... Aspek Habitat dan Avifauna
... Aspek Morfologi
... Aspek Gambaran Darah
... Aspek Genetik
Isolasi dan Purifikasi DNA ...
... Ampflifikasi RAPD
Seleksi Primer dan Pemberian Skor Pita RAPD ... . .
Anal~sis Data ...
BAE3 IV . HASIL DAN PEMBAHASAN ...
Kawasan Tempai Mencari Makan Gelatik ...
Kawasan Tempat Bersarang Gelatik ...
Avifauna ...
Kekayaan Spesies Burung ...
Kelimpahan Spesies Burung ... Asosiasi antara Avifauna dan Kawasan Mencari Tempat Makan
... Gelatik
Kelimpahan Gelatik ...
Morfologi Gelatik ... . . .
Jenls-jenls Bulu Gelatik ...
Bulu Sayap Primer dan Sekunder ... Morfometri ...
Pola Sebaran Gelatik ... Gambaran Darah ...
. .
... Diferensial Leukosit
...
Gelatik Asal Daerah Berbeda... Pola Sebaran Gelatik
... Genetik
Profil DNA Gelatik dengan RAPD ... Lokus Polimorf d m Heterozigositas
...
... Diferensiasi Genetik dan Aliran Gen
...
Pola Sebaran Gelatik... Keragaman Genetik dan Populasi Nenek Moyang Gelatik
...
Penanda RAPD dan Penanda Genetik yang LainFilogeografi dan Konservasi ...
...
Lahan Persawahan. Goa Gelatik dan Hutan Prapat BenuaBAB V . KESIMPULAN DAN SARAN ...
Saran ...
..
...DAFTAR PUSTAKA ...
DAFTAR TABEL Nomor 2.1 3.1 Teks
...
Garnbaran darah normal beberapa spesies burungBeberapa kondisi fisik dan jurniah transek yang dibuat pada kawasan tempat mencari makan gelatik (Padda oryzivora
...
( L . ) ) di Bali
Kelimpahan dan sampel gelatik (Padda oryzivora (L.) pada delapan kawasan tempat mencari makan b m g tersebut ... Sekuen 12 primer RAPD yang digunakan untuk arnplifikasi
DNA gelatik (Padda orytivora ( L . ) ) ... Stratum dan kepadatan vegetasi pada kawasan tempat mencari makan gelatik (Padda oryzivora ( L . ) ) di Baii ... Keragaman kelompok b m g - b u r u n g granivora pada delapan kawasan tempat mencari makan gelatik (Pad& oryzivora
(L.)) di Bali ... Rataan (+ s.b.) delapan karakter morfometri gelatik (Padda oryzivora ( L . ) ) asal Jawa Barat, Jawa Timilr, Bali Utara dan Bali Selatan. ... Hasil pengukuran morfometri geIatik (Padda oryzivora ( L . ) )
asal Jawa d m Bali oleh beberapa orang peneliti lain
...
Rataan garnbaran darah(+
s.b.) gelatik (Padda olyzivora (L.)) asaI Jawa Barat, Jawa Timur, Bali Utara dan Bali Selatan...
Estimasi heterozigositas untuk empat subpopulasi gelatikNomor
DAFTAR GAMBAR
Teks Halaman
...
Hipotesis Avise tentang asal-usul DNA 6
Sepasang gelatik (Padda oryzivora (L.)) &lam penangkaran ... 9
...
Distribusi gelatik (Padda oryzivora ( L . ) ) 13
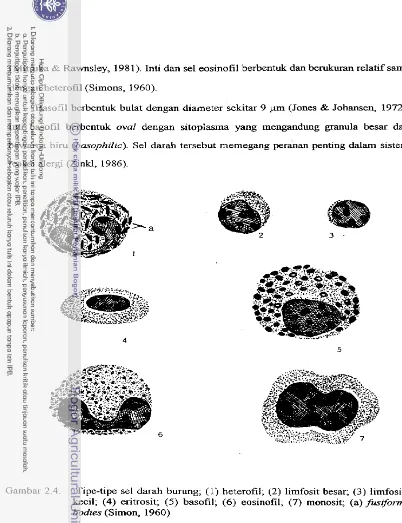
Tipe-tipe sel darah burung
...
18Beberapa kawasan tempat mencari makan gelatik (Padda
nryzivora (L.)) di Pulau Bali ... 26
Beberapat kawasan tempat mencari makan gelatik (Padda
oryzivora ( L . ) ) di Bali ... 3 8
Burung gereja (Passer rnontanus), gelatik (Padda oryzivora ( L . ) )
...
dan kawasan t e m p t mencari makan gelatik di Tamanbali.. 41
Keragarnan spesies burung pada kawasan tempat mencari makan ...
gelatik (Pa& oryzivora ( L . ) ) di Bali 43
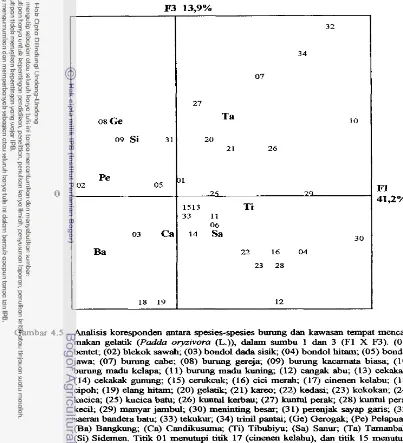
Analisis koresponden antara spesies-kawasan t e m p t mencari ...
makan gelatik (Padda oryzivora (L.)) dalam sumbu F1 d m F2. 5 1
Analisis koresponden antara spesies-kawasan tempat mencari
makan gelatik (Pa& oryzivora ( L . ) ) dalarn sumbu
F1
dan F3....
52Analisis koresponden antara spesies-kawasan tempat mencari makan gelatik (Padda oryzivora (L.)) &$am sumbu F2 dan
F3 ... 5 3
Populasi gelatik (Padda olyzivora (L.)) dewasa dan rnuda pada
delapan kawasan tempat mencari makan burung tersebut di Bali. 55
Morfologi geIatik (Padda oryzivora (L.)) ... 58
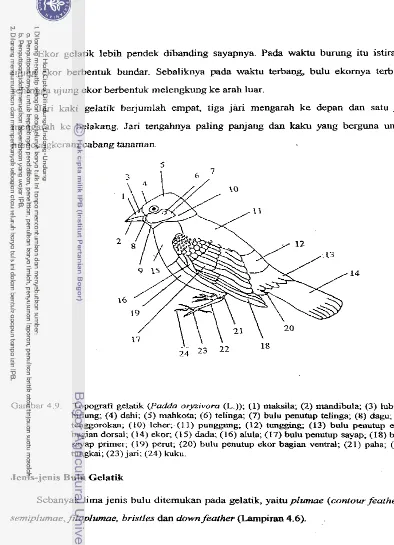
Topografi geiatik ((Padda oryzivora (L.))
...
5 9...
Grafik analisis komponen utama morfornetri gelatik (Padda oryzivora (L.)) alarn sumbu F 1 dan F2.. . .
.
. ..
.). ..
. ..
..
. .. . .
.. . .
. .. . . .
Grafik analisis komponen utama morfometn' gelatik (Padda oryzivora (L.)) dafam sumbu F1 dan F3
.
... .. .
.
..
. .. .
..
..
..
.. . .
...
. .
. . . .Grafik analisis komponen utama morfornetri gelatik (Padda oryzivora (L.))dalam sumbu F2 dan F3 ...
... .
... [image:188.529.42.415.36.581.2]Dendrogram morfometri gelatik (Padda oryzivora (L.)) berdasarkan metode W P G M
... ... . ...
....
. ... .
Grafik analisis kornponen utama gambaran darah gelatik (Padda oryzivora (L.)) dalam surnbu F1 dan F2
...
...
.. ...Gratik analisis komponen utama gambaran darah gelatik ( M a oryzivora (L.)) dalam sumbu F 1 dan F3
...
... . ....Grafik analisis komponen utama gambaran darah gelatik (Padda oryzivora (L.)) dalam sumbu P2 dan F3
... .
...Dendrogram gambaran darah gelatik (Padda oryzivora (L.)) berdasarkan metode U P G M
...
... .
. ... . .Pita hasil amplifikasi DNA total dengan primer
RP
01 (5'- AGTCCTCCCC-3') dalam agarosa 1,2%...
Pita hasil amplifikasi DNA total dengan primer RP 10 (5'- CCGGACACGA-3') dalam akrilamid 5% . . .
.
. . . ..
..
. . ..
.
. ..
.
.
. . . ..Keragaman lokus polimorf DNA antara subpopulasi-subpopulasi gelatik dalam limaprimer
RAPD
Diferensiasi genetik, aliran gen clan hubungan antara diferensiasi genetik dan aliran gen .. ...
Grafik hasil analisis komponen utarna DNA gelatik (Padda oryzivora).
. .
.. . . .
.. .
.. . .
. . .
.. . .
.. . . .
.
.. . .
.
. . .
BABI PENDAHULUAN
Latar Belakang
Pada saat ini, predikat gelatik (Padda olyzzvora (L.)) telah berubah dari "burung
hama" menjadi "burung hias". Burung tersebut tidak lagi dimusuhi sebagai binatang
yang meruskan terutama bagi para petani, tetapi sebaliknya semalcin disayangi sebagai
satwa peliharaan. Gelatik digemari banyak orang karena keelokan bentuk tubuh,
keindahan wama bulu, kemerduan suara dan kelincahan gerak-geriknya.
Adanya permintaan pasar yang t e n s bertambah menyebabkan harga gelatik
semakin meningkat. Sebagai contoh, jika pada tahun 1996 harga seekor gelatik di Pasar
Burung Satria Denpasar dan Pasar Burung Rarnayana Bogor berkisar antara Rp 5000,OO
sampai Rp 7000,OO maka dua tahun kemudian harganya meningkat 4-5 kali lipat
(Surata, 1996; 1998). Bahkan untuk burung yang telah pintar bersiul harga untuk setiap
ekornya bisa mencapai Rp 400 000,OO sampai Rp 500 000,OO (Karjono, 1998).
Peningkatan permintaan pasar terhadap gelatik bukan saja berasal dari &lam
negeri tetapi juga dari luar negeri. Hal tersebut menjadikan gelatik sebagai satu
komoditas ekspor Indonesia (Widodo et al., 1998). Sampai 30 tahun lalu, ekspor gelatik
asal Indonesia mencapai lebih dari 100 000 ekor (King, 1974). Akan tetapi sejak 10
tahun terakhir jumlah ekspor burung tersebut diperkirakan menurun bersamaan dengan
menurunnya populasi burung di alam.
Sebagian besar kebutuhan gelatik untuk memenuhi permintaan pasar masih
mengandalkan pada penangkapan dari alam. Perburuan yang terus berlangsung secara
besar-besaran terutama di Pulau Jawa dan Pulau Bali telah mengakibatkan populasi
gelatik pada daerah-daerah yang sebelumnya dilaporkan banyak ditemukan burung
tersebut. Pada masa lalu, gelatik dalam kelompok-kelornpok besar yang terdiri atas
ratusan individu didapati hampir pada semua tempat di Pulau Bali (Stresemann, 1913).
Sedangkan di Pulau Jawa gelatik sangat umum dijumpai di Jawa Tengah dan Jawa
Timur (Kuroda, 1930). Penurunan populasi gelatik sejak 20 tahun terakhir ini
mengakibatkan burung tersebut semakin jarang dijumpai di PuIau Jawa maupun Pulau
Bali (Ash, 1982; Holmes & Nash, 1991; Collar et a;., 1994). Penelitian yang dilakukan
oleh van Helvoort (198 1) terhadap burung-burung yang hidup pada ekosistem pedesaan
di Jawa Barat, tidak pemah sekalipun dijumpai gelatik pada kawasan penelitiannya.
Populasi gelatik juga tidak ditemukan dalam survei keragaman spesies burung pada
desa-desa seniman di Bali (Surata, 1993). Akan tetapi, melalui kegiatan pengamatan
burung (birdwatchzng), pada beberapa tempat di Bali masih dijumpai adanya populasi
gelatik dengan masing-masing jumIah individu kurang dari 10 ekor (Surata, 1998).
Adanya penurunan populasi yang sangat drastis mendorong International Union of
Concervation Nature and Natural Resources (IUCN/ menggolongkan gelatik dalam
katagori "rentan" atau vulnerable (Collar et al., 1994). Katagori itu rnencakup kriteria
tentang kemungkinan terjadi penurunan populasi lebih dari 50 persen dalam waktu
20 tahun (sekitar lima generasi), atau peluang punahnya 10 persen dalam
kurun
waktu100 tahun (IUCN Spesies Survival Commission, 1994).
Apabila fenornena di atas tidak segera diperhatikan, dikhawatirkan Indonesia akan
kehilangan sumber devisa dari ekspor gelatik karena kalah bersaing dengan ekspor
serupa asal negara lain. Salah satu negara yang berpotensi menyaingi ekspor gelatik
Indonesia adalah Amerika Serikat. Hal itu disebabkan populasi gelatik di Kepulauan
Hawai meningkat pesat dalam 30 tahun terakhir (Islam, 1997). Ironisnya lagi, gelatik
Jepang (Widodo et al., 1998). Burung tersebut dikembangbiakan dalam penangkaan
dengan menggunakan gelatik yang berasal dari Indonesia sebagai induknya
(Nurmaliatasari, pers. comm.).
Upaya konservasi baik secara in situ (dalam habitat alami) maupun e x situ (di luar
habitat alami) harus segera dilakukan untuk menghindarkan gelatik dari ancarnan
kepunahan. Akan tetapi, kendala utama &lam konservasi satwa liar (temasuk gelatik)
adalah keterbatasan informasi tentang sistematika dan evolusi biologi yang sesuai
dengan prioritas konservasi (Ryder & Chemnick, 1993). Penelitian yang sudah
dilakukan terhadap gelatik baru mencakup desknpsi rnorfologi (Stresemann, 1913;
Kuroda, 1933; Dupond, 1942 clan Hoogenverf, 1966); fisiologi (Nurani, 199 1; Saito et
al., 1992); prilaku (Goodwin, 1963; Baptista & Atwood, 1980) dan formula pakan
dalam penangkaran (Handini, 199 1 ; Waluyo, 199 1). Analisis genetik pada tingkat
interspesies baru dilakukan untuk memperoleh data tentang kromosom (Christidis,
1986), protein enzim (Christidis, 1987) dan albumin (Baverstock et al., 1991). Pada
tingkat intraspesies hanya tercatat penelitian tentang protein enzim (Tempuhireng,
1999). Sedangkan analisis DNA gelatik belum pernah dilaporkan, padahal penelitian
tersebut dapat memberikan kontribusi penting terhadap sistimatika dan evolusi biologi.
Untuk mengatasi kendala itu, maka dilakukan penelitian tentang filogeografi
intraspesies, yaitu proses-proses atau prinsip-prinsip yang mengarahkan penyebaran
geografi dari gelatik (Avise, 1989). Hal itu berarti fiiogeografi sebagai kajian tentang
variasi geografi, mencakup berbagai karakter seperti karakter morfologi, fisiologi,
etologi dan genetik (Lougheed & Handford, 1993). Akan tetapi ruang lingkup kajian ini
terbatas pada aspek profil vegetasi dan avifauna dari kawasaa tempat mencari makan
gelatik. Penelitian dilakukan dengan rnaksud mengungkap tingkat divergensi karakter
morfologi, garnbaran darah dan DNA untuk dijadikan bahan pertimbangan &lam
strategi konservasi.
Pulau BaIi dipilih sebagai lokasi penelitian lapang karena ( i ) tersedia informasi
yang memadai mengenai sebaran populasi gelatik; ( i i ) ada kecenderungan te rjadi isolasi
populasi gelatik, ( i i i ) terdapat rintangan (barrier) berupa pegunungan dengan ketinggian
rata-rata di atas 1000 m yang memisahkan Bali Utara dan B d i Selatan.
Tujuan Penetitian
Tujuan penelitian adalah ( i ) inventarisasi profil vegetasi, keragaman avifauna dan
kelimpahan gelatik pada kawasan tempat mencari makan burung tersebut; ( i r )
membandingkan pola filogeografi gelatik berdasarkan karakter morfometri, gambaran
darah dan genetik; ( i i i ) memperoleh penanda filogeografi yang tepat dalam penetapan
unit konservasi gelatik.
Man faat Penelitian
Penelitian ini bermanfaat dalam perancangan konservasi. Dalarn waktu dekat ini,
diperkirakan banyak populasi gelatik di alam terisolasi sehingga terancam mengalami
kepunahan lokal (local extinction). Pemulihan populasi dengan reintroduksi burung
yang berasal dari populasi lain rnerupakan satu solusi untuk rnencegah agar tidak te rjadi
kepunahan lokal pada populasi-populasi geIatik. Akan tetapi sebelum reintroduksi
dilakukan, ada atau tidaknya unit-unit konservasi yang berbeda hams diidentifikasi
terlebih dahulu. Pendekatan filogeografi dapat memberikan rekomendasi untuk tujuan
BABII
TINJAUAN PUSTAKA
Filogeografi Intraspesies
Sebagai prinsip atau proses yang mengarahkan distribusi geografi spesies tertentu,
filogeografi menekankan pada kajian variasi
gee-
yang mencakup berbagai tipekarakter seperti morfologi, fisiologi, etologi dan genetik (Lougheed & Handford, 1993).
Kajian tersebut memegang peranan penting dalam memahami proses-proses evolusi,
karena variasi-variasi yang bersifat menurun adalah materi dasar yang terlibat dalam
proses seleksi alam dan adaptasi. Penyebaran populasi suatu spesies dimulai dari variasi-
variasi geografi yang semakin lama semakin berkembang. Filogeografi pada tingkat
intraspesies dapat mengungkap keragaman di dalam spesies, mdai dari tingkat populasi
subpopulasi, subspesies sampai spesies.
Selain dapat menjelaskan pola-pola biogeografi (Mustrangi & Patton, 1997),
filogeografi juga memberikan kontribusi pada pemahaman respon populasi terhadap
perubahan lingkungan yang sangat cepat (Avise, 1997). Pertama, populasi konspesies
yang berbeda (khususnya yang terpisah dalam kurun waktu panjang) kemungkinan
memberikan respon yang tidak sama terhadap perubahan lingkungan. Adaptasi terhadap
lingkungan menyebabkan gen-gen yang terakumulasi berbeda antara populasi satu
dengan populasi lainnya. Kedua, perkembangan pesat &lam metode analisis genetik
memungkinkan dilakukannya identifikasi respon populasi terhadap modifikasi
lingkungan pada tingkat intraspesies. Ketiga, analisis filogeografi dapat mernbuktikan
hubungan dekat antara demografi populasi dan genetik. Hal tersebut &an berirnplikasi
pada konsewasi khususnya dalam periode perubahan lingkungan yang sangat cepat.
sebagai akibat sejarah kejadian demografi yang aneh. Ekspansi, fkagrnentasi dan
distribusi geografi populasi merupakan proses evolusi yang terjadi jauh sebelum
manusia mempengaruhi planet bumi. Kehadiran manusia menimbulkan bmyak
perubahan lingkungan sehingga banyak spesies perlu beradaptasi. Berdasarkan data hasil
analisis genetik maka kemampuan adaptasi suatu spesies dapat diestimasikan.
Pada Gambar 2.1 teriihat hubungan asal-usul genetik antara dua populasi
konspesies yang semula berasal dari satu daerah geografi tetapi kemudian terpisah
dalam dua daerah geografi (daerah I dan daerah 11).
Garnbar 2.1. Hipotesis Avise tentang asai-usul DNA (Avise, 1997). (A) populasi nenek moyang; (B) populasi yang merupakan keturunan dari populasi A
pada daerah I; (C) populasi yang merupakan keturunan dari popuiasi A
Barrier rnenyebabkan tidak adanya aliran gen antara populasi B dan C. Filogeni
antara keturunan populasi B dan C semakin berbeda akibat kurun waktu isolasi yang
semakin lama. Perbedaan tersebut dapat ditunjukkan dengan jarak genetik (genefic
distance), yaitu ukuran perbedaan genetik dari dua takson (Hillis et al., 1996).
Pada masa lalu, perspektif filogeografi intraspesies jarang diaplikasikan karena
kekurangan pendekatan genetik molekul untuk mendapatkan hubungan historis. Selain
itu, juga masih terdapat persepsi luas bahwa pada populasi-populasi yang masih dapat
melakukan silang antar (interbreeding), filogeni tidak memiliki pengertian yang nyata
(Avise, 1989). Hal tersebut disebabkan analisis filogeografi dilakukan berdasarkan
karakter morfologi saja sehingga hanya sedikit dapat menunjukkan keragaman
intraspesies.
Sejak dua dekade terakhir, penelitian-penelitian keragaman pada burung
berdasarkan DNA mitokondrion atau mirochondriaZ DNA (mrDNA) berkembang pesat
pada tingkat populasi. Analisis mrDNA pada tingkat intraspesies marnpu menunjukkan
keragaman yang tinggi, yang tidak terdeteksi dengan metode lain (Ball & Avise, 1992).
Dari berbagai ordo burung, filogeni Anseriformes paling banyak diteliti. Hal itu
disebabkan hubungan kekerabatan antara takson-takson dalam ordo tersebut belum
jelas, walaupun berbagai pendekatan sudah dilakukan (Zimmer et al., 1994). Spesies
burung lain yang juga telah dianalisis DNA mitokondrionnya adalah Ficedula hypoleuca
(Tegelstrom et al., 1990), Pomatostomur temporalis (Edwards, 1993),
Zonotrichia capensis (Lougheed & Hanford, 1993), Microstus agrestis (Jaarola &
Tegelstrom, 1996), Lanius lundovicianus (Mundy et al., 1997), Fringilla spp. dan
Carduelis chloris (Marshall & Baker, 1997) serta tujuh spesies dari genus Alectoris
Banyak analisis filogeografi pada burung menggunakan pendekatan distribusi
geografi dan filogeni DNA untuk menunjukkan hubungan antara aspek genetik dan
morfologi (Mourn et al., 1991; Taberlet & Bouvet, 1994). Analisis tersebut rnisalnya
pa& Melospiza melodia (Arctander et al., 1994), Anthus novaeseelandiae
(Foggo et al.. 1997), Larus glaucescens dan L. occidentalis (Bell, 1996). Berdasarkan
komparasi aspek morfologi dengan DNA lalu ditentukan ada atau tidaknya konsistensi
antara karakter morfologi dan genetik. Hal ini disebabkan status suatu populasi atau
spesies tidak dapat ditentukan hanya dengan menggunakan pendekatan genetik saja.
Analisis karakter-karakter yang lain seperti morfologi serta perilaku juga perlu
dilakukan dalam penentuan status organisme (Ball & Avise, 1992).
Analisis filogeografi dalarn penelitian ini terutama bertujuan untuk mengetahui
variasi morfologi c l a n genetik pada gelatik. Diharapkan pula &pat diungkap aspek-aspek
lain yang diduga ikut berpengaruh dalam pembentukan variasi geografi intraspesies
seperti kawasan ternpat mencari makan, keragaman avifauna dan garnbaran darah.
Diasumsikan bahwa variasi-variasi geografi merupakan keberhasilan populasi
suatu spesies dalarn rnenghambat aliran gen yang cenderung menghalangi penyebaran
populasi. Perbedaan fenotipe timbul karena daya adaptasi gen terhadap keadaan
lingkungan sekitarnya.
Ada empat kemungkinan pola filogeografi pada gelatik, (i) keragaman genetik
tinggi dan struktur geografi kuat, keduanya saling berhubungan; ( i i ) keragaman genetik
tinggi dan stmktur geografi kuat tetapi tidak saling berhubungan; (iii) keragaman
genetik tinggi dan struktur geografi lemah atau sebaliknya; ( i v ) tidak ada keragaman
Nama dan S i m a t i k a
Gelatik mempunyai
banyak
nama
lokal,
misalnya
di
Bali
disebut Jelatik, Jawa:
glatik,
Malaysia
dan
Sunda:
glatik;
Perancis:
Pa&
C a m ;
Jerman:
Reisfink
Reintogel
(Kuroda,
1932;
Mason
&
Jarvis, 1989).
Sedangkan
Mam
Bahasa
Inggris
I
gelatik disebut
J a w Sparrow, Java
Finch,
Java Ricebird, Gray Java Ricebird
dan
Pnddy
ficebird
(Berger,
1975;
Baptista
&
Atwood,
1980).
Nama ilmiah gelatik adalah Padda o y i v o r a (L.)
wonderman,
1885). Selain itu
gelatik memiliki
nama ilmiah sinonim,
yaitu
Loxia javensis
Sparm.,
Padda Verecwzda
Reich,
dan
Oryzivora leucotis Jerd. (Chasen,
1935).
Sesuai dengau prinsip the rule of
Gelatik termasuk ordo Passeriformes, famili Ploceidae, subordo Oscine dan genus
Padda (Mayr & Amadon, 1951). Semua burung yang termasuk Passeriformes tidak
dapat berdiri tegak karena otot fleksor dan jari-jari kaki beradaptasi untuk berpegangan
pada ranting dan cabang turnbuhan (Campbell & Lack, 1985). Ciri-ciri Oscine (burung
penyanyi): semua ujung dari otot siring melekat pada separuh cincin bronkus (Mayr &
Amadon, 195 1). Ciri-ciri Ploceidae: tubuh berukuran 10-15 cm, ekor pendek dan paruh
tebal yang berbentuk kerucut untuk memecah biji-bijian (MacKinnon, 1991). Ciri-ciri
Padda: paruh tebal dan besar, sayap lebih panjang dari ekor dan kaki benvarna pucat.
Bentuk morfologi burung jantan dan betina sarna (monomorf) sehingga sukar dibedakan
berdasarkan bentuk clan wama bulu (Kuroda, 1933).
Distribusi dan Habitat
Secara alami (endemik) gelatik terdapat di Pulau Jawa dan Pulau Bali. Akan
tetapi sekarang burung tersebut telah tersebar luas ke berbagai belahan dunia seperti
Kepulauan Hawai (Berger, 1975), Pvlalaysia dan Filipina (Dickinson et al., 1991),
Bangkok (Lekagul & Round, 1991), serta Sulawesi, Kalimantan dan Sumatera
(MacKinnon et al., 1999).
Pada masa lalu, Pulau Bawean, Madura dan Sumatera sempat pula diduga sebagai
habitat alami gelatik (Kuroda, 1933). Akan tetapi sekarang dipastikan ketiga pulau
bukan merupakan habitat alami gelatik. Pa& waktu penjajahan Belanda, kemungkinan
gelatik dibawa ke pulau-pulau tersebut sebagai satwa peliharaan. Beberapa ekor burung
Habitat utama gelatik adalah daerah persawahan, semak belukar dan kawasan
terbuka lainnya sampai ketinggian 1500 m dpl (Holmes & Nash, 1991). Di Kepulauan
Hawai, gelatik umumnya lebih menyukai semak-belukar, padang rumput, taman-taman
dan kawasan pemukiman penduduk yang terletak di daerah dataran rendah (Islam,
1997). Di Pulau Jawa, kawasan pertanian, padang nunput, sekitar kolam ikan dan hutan
bakau merupakan habitat utama gelatik (Stresemann, 1930). Di Pulau Bali gelatik
dijumpai pada semak-belukar, hutan bakau, hutan pantai, daerah pertanian dan kawasan
pemukiman penduduk (Ash, 1982; van Helvoort, 1981).
Kawasan tempat mencari makan
Gelatik termasuk kelompok burung granivora karena makanan utamanya adalah
biji tanaman dari famili Graminae terutama padi (Oryza sativa L.). Makanan lainnya
adalah biji sorgum (Andropogon sorghum Brot.), biji bambu (Bambusa spp.), biji kerasi
(Lantana carnara L.), biji glagah (Saccharurn sponfaneum L.) dan biji bayam
(Amaranthus spp.) (Sody, 1989; Surata, 1998).
Pada waktu jumlah populasi masih besar, gelatik sering dijumpai berkelompok
sampai mencapai ratusan ekor menyerbu tanaman padi secara bersama-sama sehingga
dianggap sebagai pesaing oleh manusia (Johnston & Klitz, 1977). Jika daerah
persawahan di Pulau Jawa terendam banjir, burung tersebut membentuk kelompok-
kelompok kecil atau malah hanya sepasang-sepasang mencari makanan berupa blji-
bijian, buah-buahan dan insekta (van Balen, 1997). Di Pulau Bali gelatik juga dijumpai
mencari makan di gudang beras, penyosohan gabah, kawasan perhotelan dan di sekitar
bondol jawa (L. leucogastroides), bondol dada hitam (L. malacca), burung gereja
(Passer rnontanus) dan tekukur (Streptopelia chinensis) (Surata, 1998).
lsotasi Populasi
Pada masa yang lalu, gelatik tersebar di Pulau Bali mulai dari Buleleng, Negara,
Singaraja, Gitgit, Candikusuma, Bali Barat, Tanah Lot, Banyuwedang, Petitenget, Uiu
Watu, Pesanggaran, Wangaya Gede, Padangbai, Nusa Dua, Amed dan Prapat Benua
(Ash et al., 1987). Menurut Ash (1982), di Bali gelatik belum pernah dijurnpai di atas
ketinggian 900 m. Padahal dari ujung barat sampai timur Pulau Bali terdapat
pegunungan yang memanjang dengan ketinggian 500 sampai 3000 m dpl (Bappeda Tk.1
Bali, 1996). Pegunungan tersebut kemungkinan menjadi rintangan yang memisahkan
antara subpopulasi gelatik yang terdapat di Bali Utara dengan subpopulasi gelatik yang
terdapat di Bali Selatan.
Subpopulasi yang saling terpisah diperkirakan juga terbentuk akibat fragrnentasi
habitat, yaitu tercerai-berainya habitat karena alih fungsi lahan, terutarna dari daerah
persawahan menjadi kawasan pemukiman (Ferron, 1990; Soeriatmadja, 1992). Kawasan
perkotaan umumnya mengalami fragrnentasi habitat lebih banyak dibanding kawasan
pedesaan karena alih fungsi lahan pada kawasan perkotaan jauh lebih pesat dibanding
kawasan pedesaan. Alih fungsi lahan telah mengakibatkan berkurangnya kawasan
tempat mencari makan dan bersarang bagi gelatik (Fuller er al., 1995). Intensifikasi
pertanian yang diikuti dengan pembersihan semak-belukar dan penanaman padi dengan
usia yang semakin pendek, selain mengakibatkan kehilangan tempat bersarang dan