MODIFIKASI KANDUNGAN LIGNIN KAYU SENGON
(
Paraserianthes falcataria
(L.) Nielsen) MELALUI REKAYASA
GEN 4-COUMARATE COA LIGASE (4CL)
N. SRI HARTATI
DEPARTEMEN BIOLOGI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI
DAN SUMBER INFORMASI
Saya menyatakan dengan sebenar-benarnya bahwa segala pernyataan dalam disertasi saya yang berjudul Modifikasi Kandungan Lignin Kayu Sengon
(Paraserianthes falcataria (L.) Nielsen) Melalui Rekayasa Gen 4-Coumarate
CoA Ligase (4CL) adalah karya saya sendiri bersama pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal dan dikutip dari karya yang diterbitkan maupun tidak diterbitkan oleh penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini
© Hak cipta milik IPB, tahun 2011 Hak cipta dilindungi undang-undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
2. Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis dalam bentuk apapun
ABSTRACT
N. SRI HARTATI. Modification of Lignin Content of Wood of Sengon (Paraserianthes falcataria (L.) Nielsen) by Engineered gene of 4-Coumarate CoA Ligase (4CL). Under direction of Suharsono, Kurnia Sofyan, and Enny Sudarmonowati
pCAMBIA 2301. Recombinant expression vector of antisense 4CL has been successfully introduced into sengon via Agrobacterium tumefaciens. Gene integration test by PCR method showed that 19 out of 112 transformed sengon seedlings which resistant to kanamycine were transgenic seedlings. Lignin content determination of transgenic seedlings stem including histochemical assay, Klason lignin content and FTIR analysis to determine the S/G ratio was indicated that two transgenic sengon seedlings namely 4CLAS-4 and 4CLAS-1 possessed lower lignin content (15:53%) and higher S/G ratio than the control. These plants also showed the best growth characteristic and normal morphological appearance.
RINGKASAN
N.SRI HARTATI. Modifikasi Kandungan Lignin Kayu Sengon (Paraserianthes falcataria (l.) Nielsen) Melalui Rekayasa gen 4-Coumarate CoA Ligase (4CL). Dibimbing oleh Suharsono, Kurnia Sofyan, Enny Sudarmonowati
Pemisahan lignin dari selulosa membutuhkan input bahan kimia dan energi yang tinggi yang berdampak pada tingginya biaya produksi dan resiko ekologi yang membahayakan. Keberadaan lignin pada sel tanaman merupakan faktor pembatas efisiensi pengolahan material lignoselulosa menjadi produk-produk industri berbahan dasar kayu termasuk pulp baik secara kimia maupun biologis. Kadar dan komposisi lignin yang terkandung dalam tanaman akan mempengaruhi efisiensi proses pulping. Oleh karena itu bahan baku pulp dengan kadar lignin rendah atau yang memiliki komposisi lignin dengan reaktivitas tinggi lebih mudah dipisahkan dari selulosa, sehingga akan sangat menguntungkan bagi industri pulp karena akan menghemat energi dan biaya.
Sengon (P. falcataria) memiliki keunggulan untuk digunakan sebagai bahan baku pulp karena dimensi seratnya yang baik, sifat fisik dan mekanik lembaran kertas dari kayu sengon kualitasnya tinggi, disamping itu sengon dapat tumbuh cepat. Penelitian ini bertujuan untuk (1) mengisolasi cDNA penyandi 4-coumarate CoA ligase (4CL) dari tanaman sengon, (2) mengkonstruksi vektor biner gen 4CL dengan orientasi antisense (terbalik) untuk tranformasi pada sengon, (3) merakit tanaman sengon transgenik yang mempunyai karakteristik rendah lignin ataupun rasio siringil yang lebih tinggi dengan mengintroduksikan konstruk vektor biner gen (4CL) dengan orientasi antisense. Kegiatan penelitian yang telah dilakukan untuk modifikasi kadar ataupun komposisi lignin sengon melalui teknologi DNA rekombinan (rekayasa genetik) terdiri dari beberapa percobaan yaitu (1) analisis kuantitatif dan uji histokimia lignin, (2) induksi embriogenesis dan induksi tunas dari nodal kotiledon, (3) isolasi dan pengklonan cDNA fragmen gen penyandi 4-coumarate: Coenzyme A ligase dari sengon, dan (4) transformasi genetik sengon dengan fragmen gen 4CL antisense.
sekuen nukleotida produk RT-PCR memiliki homologi yang tinggi dengan sekuen 4CL terkonservasi dari Rubbus ideaus, Oryza sativa, Populus tomentosa, Populus balsamifera, Betulla platyphilla, Nicotiana tabacum, and Arabidopsis thaliana dengan kisaran identity 78-90%. Fragmen ini telah berhasil disisipkan diantara promotor 35S dan terminator NOS dengan arah terbalik (antisen) di dalam vektor biner pCAMBIA 2301. Vektor ekspresi rekombinan berupa antisense 4CL telah berhasil diintroduksikan ke dalam sengon melalui Agrobacterium tumefaciens. Hasil uji integrasi gen terhadap 112 bibit sengon yang tahan pada media seleksi yang mengandung antibiotik kanamisin, menunjukkan bahwa 19 diantaranya merupakan tanaman transgenik. Berdasarkan hasil pengujian kadar dan komposisi lignin yang meliputi uji histokimia, kadar lignin Klason dan analisis spektroskopi FTIR (Fourier Transformed Infra Red) untuk mengetahui S/G rasio, diperoleh 2 tanaman transgenik yaitu 4CLAS-4 yang memiliki keunggulan rendah kadar lignin (15.53%) dibanding kadar lignin sengon secara umum yaitu sekitar 26% dan 4CLAS-1 yang memiliki rasio S/G tinggi dibanding kontrol. Kedua tanaman tersebut menunjukkan pertumbuhan yang paling baik dan memiliki sifat morfologi yang normal.
MODIFIKASI KANDUNGAN LIGNIN KAYU SENGON
(
Paraserianthes falcataria
(L.) Nielsen) MELALUI REKAYASA
GEN 4-COUMARATE COA LIGASE (4CL)
N. SRI HARTATI
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Departemen Biologi
DEPARTEMEN BIOLOGI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
Judul :Modifikasi Kandungan Lignin Kayu Sengon (Paraserianthes falcataria (L.) Nielsen) Melalui Rekayasa Gen 4- coumarate CoA Ligase (4CL) Nama Mahasiswa : N. Sri Hartati
NRP : G361050111
Program Studi : Biologi
Disetujui,
Komisi Pembimbing
Dr. Ir. Suharsono, DEA Ketua
Prof. Dr. Ir. Enny Sudarmonowati Prof. Dr. Ir. Kurnia Sofyan Anggota Anggota
Diketahui,
Ketua Program Studi Biologi Dekan Sekolah Pascasarjana
Dr. Ir. Dedy Duryadi Solihin, DEA Dr. Ir. Dahrul Syah, M.Sc.Agr.
Penguji pada ujian tertutup : 1. Dr. Ir. Miftahuddin, M.Si 2. Dr. Ir. Sri Wilarso Budi R
PRAKATA
Puji syukur penulis panjatkan ke hadirat Allah SWT karena hanya atas perkenanNya penulis dapat menyelesaikan penelitian dan penulisan disertasi dengan judul “Modifikasi Kandungan Lignin Kayu Sengon (Paraserianthes falcataria (l.) Nielsen) Melalui Rekayasa gen 4-Coumarate CoA Ligase (4CL)”. Penelitian ini dilaksanakan dari bulan April 2006 hingga September 2010. Sumber dana penelitian adalah dari kegiatan Riset Unggulan Terpadu XII dan dana penelitian Laboratorium Biologi Molekuler 3- Puslit Bioteknologi LIPI.
Pada kesempatan ini, penulis mengucapkan terimakasih yang sebesar-besarnya kepada Dr. Ir. Suharsono, DEA selaku ketua komisi pembimbing, Prof. Dr. Ir. Enny Sudarmonowati dan Prof. Dr. Ir. Kurnia Sofyan selaku anggota komisi pembimbing atas gagasan, dukungan dan motivasi yang sangat berguna dalam pelaksanaan penelitian dan penyusunan disertasi. Ucapan terimakasih disampaikan pula kepada Kepala Pusat Penelitian Bioteknologi LIPI yang telah memberikan ijin kepada penulis untuk melanjutkan studi pada program doktor, Lembaga Ilmu Pengetahuan Indonesia atas dana beasiswa yang telah diberikan kepada penulis untuk mengikuti program doktor, Sekolah Pasca Sarjana IPB, Rektor IPB, Dekan SPs IPB, Dekan Fakultas MIPA, Ketua Program Studi Biologi SPs IPB, Ketua Departemen Biologi IPB yang telah memberikan kesempatan untuk menimba ilmu di Institut Pertanian Bogor serta keluarga atas segala dukungan dan doa yang selalu diberikan. Ucapan terima kasih juga disampaikan kepada semua pihak yang tidak dapat disebutkan satu persatu yang telah membantu penulis dalam melakukan penelitian dan penulisan disertasi ini.
Semoga karya ilmiah dalam bentuk disertasi ini dapat memberikan manfaat dan berguna bagi pengembangan ilmu pengetahuan dan teknologi.
Bogor, Februari 2011
RIWAYAT HIDUP
Penulis dilahirkan di Tasikmalaya pada tanggal 26 Desember 1969 sebagai anak sulung dari ayah Dr. Maryoto Hadi Purnomo (Alm) dan ibu Ati Susilawati. Pendidikan sarjana di tempuh di Program Studi Kimia, Institut Teknologi Sepuluh November Surabaya, lulus pada tahun 1992. Pada tahun 1999, penulis diterima di Program Studi Bioteknologi pada Program Pascasarjana IPB dan menyelesaikannya pada tahun 2002. Kesempatan untuk melanjutkan ke Program doctor pada Program Studi Biologi, Program Pascasarjana IPB diperoleh pada tahun 2005. Beasiswa pendidikan pascasarjana diperoleh dari Lembaga Ilmu Pengetahuan Indonesia (LIPI).
Penulis bekerja sebagai Peneliti di Pusat Penelitian Bioteknologi-LIPI sejak tahun 1993 pada Bidang Biologi Molekuler. Bidang penelitian yang didalami oleh penulis adalah analisis keragaman genetik tanaman, isolasi dan pengklonan gen, serta transformasi genetik tanaman dengan gen yang berhubungan dengan biosintesis dinding sel.
DAFTAR ISI
Halaman
DAFTAR TABEL vii
DAFTAR GAMBAR viii
PENDAHULUAN UMUM
Latar belakang……… 1
Tujuan penelitian……… 4
Manfaat penelitian……….. 4
Hipotesis penelitian……… 4
Ruang lingkup penelitian………... 5
TINJAUAN PUSTAKA……….. 6
ANALISIS KUANTITATIF DAN UJI HISTOKIMIA LIGNIN SENGON (Paraserianthes falcataria)……….. Abstract……….. 19
Abstrak………... 19
Pendahuluan………... 20
Bahan dan metode………... 22
Hasil dan pembahasan……… 24
Kesimpulan dan saran………. 30
PRODUKSI TUNAS MELALUI INDUKSI EMBRIOGENESIS DAN MULTIPLIKASI TUNAS NODAL KOTILEDON SENGON Abstract……….. 31
Abstrak………... 31
Pendahuluan………... 32
Bahan dan metode………... ... 33
Hasil dan pembahasan……… 34
Kesimpulan dan saran………. 38
PENGKLONAN DAN KONSTRUKSI VEKTOR EKSPRESI ANTISENSE DARI FRAGMEN GEN PENYANDI 4-COUMARATE: COENZYME A LIGASE DARI SENGON Abstract……….. 39
Abstrak………... 39
Pendahuluan………... 40
Bahan dan metode………... 42
Hasil dan pembahasan……… 45
Kesimpulan dan saran………. 52
TRANSFORMASI GENETIK SENGON DENGAN FRAGMEN GEN 4-COUMARATE COA LIGASE (4CL) ANTISENSE PADA SENGON MELALUI Agrobacterium tumefaciens Abstract……….. 53
Abstrak………... 53
Pendahuluan………... 54
Bahan dan metode………... 56
Hasil dan pembahasan……… 59
PEMBAHASAN UMUM……….. KESIMPULAN DAN SARAN……….
DAFTAR TABEL
Halaman Aktivitas katalitik beberapa enzim yang terlibat dalam biosintesis
lignin………... 15
Kadar lignin kayu sengon yang dikoleksi dari daerah Cibinong, Cikampek, Tasikmalaya dan Yogyakarta………. 25 Persentase pembentukan somatik embrio yang diinduksi dari aksis embrio sengon pada 4 jenis media……….. 35 Jumlah tunas yang terbentuk dari nodal kotiledon pada beberapa jenis dan komposisi media... 37 Efisiensi hasil transformasi konstruk antisense 4CL pada sengon
berdasarkan PCR………... 62
Kadar lignin sengon hasil transformasi konstruk antisense 4CL
dan kontrol………... 67
Rasio siringil dan guaiasil lignin sengon hasil transformasi
konstruk antisense 4CL dan kontrol……… 71 Karakteristik gugus fungsi lignin Klason tanaman sengon transgenik antisense 4CL dan kontrol berdasarkan serapan infra
merah……… 72
DAFTAR GAMBAR
Halaman
Diagram alur penelitian……… 5
Pohon sengon……… 6
Struktur kimia penyusun lignin……… 11
Diagram struktur lignin kayu lunak……….. 13
Jalur biosintesis prekursor monolignol lignin………... 14
Isolasi dan uji histokimia lignin bibit sengon berbagai umur……... 27
Intensitas histokimia lignin dengan phloroglucinol dan kadar lignin kecambah sengon pada berbagai umur………... 28
Profil histokimia lignin kayu sengon bagian bawah, tengah dan atas pada pohon sengon umur 1 tahun………... 29
Profil histokimia lignin kayu sengon bagian bawah, tengah dan atas pada pohon sengon umur 2 tahun………... 29
Profil histokimia lignin kayu sengon dewasa yang dikoleksi dari Cibinong………... 30
Kalus embriogenik sengon yang diinduksi dari aksis embrio pada media MS + 0.05 mg/l IAA + 2 mg/l TDZ dan MS + 0.15 mg/l IAA dan 2 mg/l TDZ pada umur 1 bulan………. 35
Struktur kalus embriogenik dan embriosomatik yang diinduksi dari aksis embrio pada media MS + 0.1 mg/l TDZ + 0. 25 mg/l IAA pada umur umur 2, 4, 5 dan 7 minggu... 36
Induksi dan regenerasi tunas dari nodal kotiledon……… 37
Bibit sengon umur 4 bulan yang berasal dari kultur in vitro nodal kotiledon dari kecambah umur 10 hari pada media MS yang mengandung 1 mg/l TDZ……….. 38
Peta situs restriksi PGEM®-T Easy dan pCAMBIA 2301……….. 44
Fragmen produk RT-PCR berukuran 342 dan 500 bp yang diamplifikasi menggunakan degenerate primer kondisi annealing 54.4oC selama 45 detik dan ekstensi 60 detik dan fragmen hasil pemisahan………. 45
Koloni hasil transformasi E. coli DH5α dengan vektor ekspresi fragmen 4CL antisense……… 46
Urutan nukleotida dan deduksi asam amino dari fragmen gen 4CL sengon………... 47
Hasil analisis BLAST P sekuen asam amino yang dideduksi dari produk RT-PCR sengon……… 48
Dendogram 4CL berdasarkan urutan nukleotida beberapa tanaman 49 49 Analisis situs restriksi dengan program NEB cutter……… Konstruksi vektor ekspresi fragmen gen 4CL antisense…………... 50
Koloni A. tumefaciens dan E. coli. A. Koloni A. tumefaciens pCAMBIA 2301, E. coli DH5α transforman hasil transformasi konstruk antisense fragmen gen 4Cl, A. tumefaciens LBA 4404, A. tumefaciens LBA4404 hasil transformasi konstruk antisense…….. 51
Hasil PCR tanaman sengon hasil transformasi fragmen gen 4CL antisen dengan primer NPTII……….. 62 Tanaman yang tahan pada media seleksi yang mengandung kanamisin 300 mg/l………... 63 Tanaman transgenik sengon umur 4 bulan yang mengandung fragmen gen 4CL antisense yang digunakan untuk analisis
komposisi lignin……… 64
Morfologi daun sengon transgenik yang mengandung fragmen gen 4CL antisense dan kontrol……… 64 Batang sengon transgenik antisense 4CL dan kontrol……….. 65 Tinggi dan diameter tanaman sengon hasil gransformasi dengan fragmen gen 4CL antisen dan kontrol pada umur 4 bulan…………
66 Uji histokimia lignin bagian tengah batang bibit sengon transgenik
dan kontrol……… 68
DAFTAR LAMPIRAN
Halaman Perhitungan rasio siringil dan guaiasil lignin Klason tanaman
sengon transgenik kontrol berdasarkan metoda garis dasar…….. 88
PENDAHULUAN UMUM
Latar Belakang
Kayu merupakan material lignoselulosa yang memiliki nilai ekonomi tinggi dengan pemanfaatan yang luas mencakup kayu konstruksi, kayu lapis, pulp dan kertas hingga perkembangan terkini yang telah memulai memanfaatkan kayu sebagai bahan baku bioetanol. Khusus untuk industri pulp dan kertas, hingga kini kayu masih diandalkan sebagai bahan baku utama. Industri pulp terus berkembang dan produksinya cenderung meningkat dari tahun ke tahun seiring dengan bertambahnya konsumsi kertas dunia. Secara umum, produksi kertas dan pulp dunia pada tahun 2004 adalah sekitar 360 juta ton dan diperkirakan akan meningkat hingga 494 juta ton pada 2020 (Teraäs 2007).
Potensi Indonesia masih sangat mendukung perkembangan sektor industri pulp dunia dengan mengedepankan visi kelestarian lingkungan. Berbagai upaya telah dilakukan untuk meningkatkan produksi pulp ramah lingkungan antara lain modifikasi proses bleaching tanpa senyawa organik terklorinasi (AOX). Hal tersebut berkaitan dengan salah satu aspek kebijaksanaan pengelolaan lingkungan hidup yang berkenaan dengan baku mutu lingkungan yaitu pengurangan produksi limbah, dalam rangka peningkatan efisiensi produksi dengan maksud untuk mengurangai produksi limbah yang berupa limbah bahan berbahaya dan beracun (B3), cair, padat, dan gas. Pemilihan bahan baku, pengembangan teknologi, pemanfaatan ulang dan lain-lain dapat dilakukan dalam rangka peningkatan efisiensi produksi (KLH 1994).
Warna gelap pada pulp umumnya disebabkan oleh lignin, salah satu komponen utama penyusun kayu yang tergolong ke dalam senyawa fenolik yang sangat mudah teroksidasi. Pada sisi lain, limbah organik terklorinasi yang dihasilkan dari proses bleaching sulit untuk didegradasi atau didaur ulang sehingga berpotensi sebagai bahan pencemar yang membahayakan lingkungan. Lebih dari sepuluh tahun terakhir ini, berbagai cara minimisasi limbah dalam industri pulp telah ditempuh yaitu melalui pengolahan pulp dengan memanfaatkan mikroba (biopulping) maupun modifikasi proses bleaching. Modifikasi proses
bleaching yaitu Totally Chlorin Free (TCF) bleaching yang menggunakan bahan kimia hidrogen peroksida dan ozon (Johnston et al. 1996) telah berhasil menurunkan toksisitas efluen menjadi lebih rendah dibanding proses yang umum dipakai sebelumnya yaitu Elemental Chlorin Free (ECF). Namun demikian, modifikasi proses bleaching pulp bersifat menurunkan saja dan tidak dapat menghilangkan limbah berbahaya sama sekali. Keberadaan lignin merupakan faktor pembatas efisiensi pengolahan material lignoselulosa menjadi produk-produk industri terkait termasuk pulp baik secara kimia maupun biologis (biopulping) menggunakan jamur pelapuk putih. Asosiasi lignin dengan selulosa membentuk suatu matriks hidrofobik yang membatasi efektifitas kerja enzim-enzim hidrolitik yang berperan dalam proses biopulping.
Pengurangan limbah dapat pula dilakukan melalui subtitusi atau pemilihan bahan baku yang mendukung efisiensi proses bleaching. Selain itu struktur lignin mempengaruhi proses kimia dan fisika pulping kayu dan penggunaan serat yang dihasilkannya. Dengan demikian penggunaan bahan baku kayu dengan kadar lignin rendah atau yang memiliki komposisi lignin dengan reaktivitas tinggi yaitu yang rasio siringil ligninnya lebih tinggi sehingga lebih mudah dipisahkan dari selulosa, akan sangat menguntungkan bagi industri pengolahan pulp karena akan menghemat energi dan biaya disamping dapat menekan limbah berbahaya.
genetik) guna mengontrol biosintesis komponen dinding sel tanaman termasuk lignin. Studi molekuler mengenai enzim-emzim yang terkait dengan biosintesis lignin telah dimulai sejak tahun 1990-an dari tanaman model Arabidopsis thaliana
dan tembakau hingga tanaman berkayu seperti poplar dan pinus. Tanaman berkayu trasgenik rendah lignin pertama adalah antisense 4-Coumarate Coenzim A ligase (4CL) poplar yang dilaporkan oleh kelompok peneliti dari North Carolina University. Berbagai macam gen terkait biontesis lignin dari berbagai tanaman saat ini telah terdokumentasi pada situs gene bank. Selain itu studi mengenai fungsi gen terkait biosintesis lignin dengan memanfaatkan teknik transgenesis baik yang bersifat up regulasi maupun down regulasi telah banyak dilaporkan.
Sengon (P. falcataria) merupakan salah satu komoditas yang diprioritaskan pada Hutan Tanaman Industri (HTI) karena sifatnya yang tumbuh cepat. Kayu sengon memiliki keunggulan untuk digunakan sebagai bahan baku pulp karena dimensi seratnya memenuhi syarat untuk digunakan sebagai bahan baku pulp serta sifat fisik dan mekanik lembaran kertas dari kayu sengon sangat baik yaitu memiliki nilai tensile strength (daya regang) dan bursting strength (kekuatan robek) yang tinggi.
transgenik kadar lignin termodifikasi dapat menjadi salah satu pilihan bahan baku industri pulp dan kertas yang lebih efisen dan ramah lingkungan.
Tujuan Penelitian
Penelitian ini bertujuan untuk:
1. Mengetahui kisaran kadar lignin kayu sengon dari berbagai lokasi tumbuh. 2. Memperoleh metoda dan bahan yang tepat untuk proses transformasi genetik. 3. Mengisolasi cDNA penyandi 4-coumarate CoA ligase (4CL) dari tanaman
sengon.
4. Mengkonstruksi vektor biner rekombinan yang mengandung fragmen gen 4CL dengan orientasi antisense (terbalik) untuk tranformasi pada sengon.
5. Merakit tanaman sengon transgenik yang mempunyai karakteristik rendah lignin ataupun rasio siringil yang lebih tinggi dengan mengintroduksikan fragmen gen 4CL dengan orientasi antisense.
Manfaat Penelitian
1. Gen 4-coumarate CoA ligase 4CL) yang diisolasi dari sengon yang merupakan tanaman asli Indonesia, HKI nya menjadi milik sendiri dan dapat melengkapi deretan gen 4CL yang sudah diperoleh dari tanaman lain dan akan memberikan kontribusi untuk perkembangan mutakhir dalam perbaikan tanaman hutan tropis.
2. Vektor ekspresi gen 4CL antisense yang dapat berpengaruh terhadap modifikasi komposisi lignin pada sengon dapat diterapkan pada tanaman berkayu (tanaman hutan) bahan baku pulp lainnya.
Hipotesis Penelitian
Ruang lingkup penelitian
Kegiatan penelitian yang telah dilakukan untuk modifikasi kadar ataupun komposisi lignin sengon melalui teknologi DNA rekombinan (rekayasa genetik) terdiri dari beberapa percobaan (Gambar 1). Penelitian ini terdiri dari: (1) analisis kuantitatif dan uji histokimia lignin, (2) induksi embriogenesis dan induksi tunas dari nodal kotiledon, (3) isolasi dan pengklonan cDNA fragmen gen penyandi 4-coumarate: Coenzyme A ligase dari sengon, dan (4) transformasi genetik sengon dengan fragmen gen 4CL antisense.
Analisis kuantitatif dan uji histokimia bibit dan sengon dewasa bertujuan untuk mengetahui kadar lignin sengon dari berbagai lokasi tumbuh, distribusinya pada perbedaan ketinggian pohon dan untuk menganalisis pembentukan lignin atau senyawa serupa lignin pada berbagai tahapan umur pohon. Percobaan teknik
in vitro yaitu induksi embriogenesis dan tunas bertujuan untuk mendapatkan metoda dan bahan yang tepat untuk proses transformasi genetik sehingga menghasilkan tanaman transgenik. Dalam rangka memodifikasi kadar lignin kayu sengon melalui teknologi DNA rekombinan harus dilakukan isolasi salah satu gen yang berkaitan dengan biosintesis lignin yaitu 4-coumarate CoA ligase (4CL). Untuk menghambat ekspresi gen 4CL yang terdapat di dalam sengon, fragmen gen 4CL dengan arah ekspresi terbalik diintroduksikan ke dalam tanaman sengon dibawah kendali promotor kuat 35S.
Tanaman sengon
Tanaman
transgenik
Pengklonan dan konstruksi vektor
ekspresi antisense fragmen gen penyandi 4-Coumarate: Coenzyme A ligase
Induksi embriogenesis
dan tunas
Analisis kuantitatif dan uji histokimia
lignin
TINJAUAN PUSTAKA
Klasifkasi dan botani sengon
Sengon merupakan salah satu jenis tanaman yang tumbuh cepat di daerah tropis dan ditemukan pertama kali oleh Teysman di Pulau Banda pada tahun 1871 (Santosa 1992). Jenis-jenis P. falcataria terdapat di berbagai macam habitat dari permukaan laut hingga ketinggian 1600 m atau lebih, di hutan primer terutama di hutan basah sekunder di dataran rendah, juga di hutan pegunungan, hutan lumut, dan dataran berumput atau di sepanjang tepi jalan dekat laut. Tumbuh pada tanah berpasir dan pada tanah laterit dengan drainase cukup baik.
Di Indonesia, P. falcataria mempunyai nama daerah bermacam-macam. Di Jawa misalnya dikenal dengan nama jeunjing, sengon laut, albizia, sengon landak, sengon lendi, sengon sarang dan kalbi. Di Sulawesi dikenal dengan nama tedehu pute, sedangkan di Maluku dikenal dengan nama rawe, selawoku, selawoku merah, sika, sika bot, tawasela atau sikas dan di Irian Jaya dikenal dengan nama bae, bai, wahogon, wai atau wikie. Di Madura dikenal dengan nama jing laut sedangkan di Malaysia dan Brunei Darussalam dikenal dengan nama puah dan batai atau kayu macis (Samingan 1982).
Menurut Samingan (1982) sistematika taksonomi Paraserianthesfalcataria
adalah sebagai berikut :
Divisi : Spermatophita Sub-divisi : Angiospermae Klas : Dikotyledone Ordo : Rosales Famili : Leguminoseae Sub-famili : Mimosaceae Genus : Paraserianthes
Spesies : Paraserianthes falcataria
Sengon merupakan salah satu jenis tanaman yang tumbuh cepat di daerah tropis dan ditemukan pertama kali oleh Teysman di Pulau Banda pada tahun 1871 (Santosa, 1992). Pada umur satu tahun dapat mencapai tinggi 7 m dan pada umur 12 tahun dapat mencapai tinggi 39 m dengan diameter 63,5 cm. Diameter pohon yang sudah tua dapat mencapai 100 cm dan kadang- kadang lebih. Batang umumnya tidak berbanir, lurus dan silindris. Kulit licin berwarna abu-abu atau kehijau- hijauan. Tajuk berbentuk perisai, jarang dan selalu hijau (Hidayat et al. 2002).
Kayu sengon termasuk kelas kuat dan kelas awet IV – V (Abdurachman & Hadjib 2009). Sengon merupakan salah satu jenis pohon yang dikembangkan dalam program Hutan Tanaman Industri. Tanaman ini mempunyai sifat-sifat unggul yaitu dapat tumbuh cepat pada tanah miskin hara dan drainase yang kurang baik, batang lurus, dan multi guna sebagai kayu pertukangan maupun bahan baku industri pulp. Sifatnya yang tumbuh cepat sangat sesuai digunakan dalam reboisasi dan penghijauan lahan-lahan kritis sebagai penyubur tanah.
lapis, venir, bahan bangunan, perabot rumah tangga dan kayu bakar dan daunnya untuk makanan ternak dan pupuk hijau (Anggraeni 2008).
P. falcataria termasuk jenis kayu yang diprioritaskan untuk hutan tanaman karena kayunya sesuai untuk bahan baku pulp, kertas dan kayu pertukangan selain karena pertumbuhannya yang cepat. Kayu sengon memiliki massa jenis sekitar 0.4 g/cm3 (Ishiguri et al. 2007; Yahya 2010). Dibandingkan jenis-jenis lain dengan massa jenis yang sama, keawetan kayu ini lebih tinggi sehingga sengon banyak dipergunakan sebagai bahan bangunan (Prajadinata & Masano 1989).
Kayu sengon memiliki panjang dan diameter serat 1.373 dan 0.0160 mm. Berdasarkan dimensi serat dan juga massa jenisnya, kayu sengon sesuai untuk digunakan sebagai bahan baku pulp sebagaimana yang dikemukakan oleh Haroen (2006), bahwa umumnya massa jenis kayu yang digunakan untuk pulp massa jenisnya lebih kecil dari 0.7, panjang serat lebih dari 0.9 mm, kadar lignin kurang dari 33% dan ekstraktif lebih kecildari 5%. Demikian pula jika ditinjau dari sifat fisik dan mekanik kertas yang dibuat dari kayu sengon, kualitasnya termasuk kategori kualitas 1 dengan keunggulan sifat fisik dan mekanik diantaranya kekutan sobek (burst factor), tensile strength, daya regang (stretch) dan panjang putus (breaking length) dengan nilai berturut-turut 88.78, 6.44, 3.74 dan 10.23 (Yahya 2010).
Permasalahan pada industri pulp dan kertas
Sejalan dengan peningkatan kebutuhan pulp dan kertas dunia, industri pulp dan kertas dituntut untuk meningkatkan efisiensi produksi dengan tetap menjaga kelestarian sumber bahan baku. Selain itu tuntutan mutu lingkungan dengan menekan pencemaran industri, mendorong penelitian yang sangat intensif di bidang teknologi proses maupun rekayasa jenis tanaman bahan baku pulp. Modifikasi bahan baku pulp dengan meminimalkan faktor-faktor pembatas pada proses pembuatan pulp akan meningkatkan efisiensi proses.
maupun campuran, waktu pengolahan yang relatif pendek dan menghasilkan pulp dengan kualitas tinggi, namun demikian pulp yang dihasilkan berwarna gelap sehingga membutuhkan banyak bahan pemutih dan menghasilkan limbah berupa bahan organik terklorinasi (Siagian 2003).
Pulp coklat (unbleached pulp) yang merupakan hasil pencucian kemudian disaring, diputihkan, atau dikelantang pada unit pemutih (bleaching) yang umumnya dilakukan dalam tiga hingga enam tahap. Pada proses pemutihan menggunakan zat-zat kimia dari golongan klorin terutama Cl2, akan bereaksi dengan lignin menghasilkan limbah berupa senyawa organoklorin yang umumnya beracun. Bleaching pulp ditinjau dari segi proses industri sangat penting karena derajat putih pulp tidak hanya merupakan standard kualitas yang digunakan oleh industri tetapi juga sebagai salah satu kriteria pemilihan oleh konsumen.
Warna gelap pada pulp umumnya disebabkan oleh lignin, salah satu komponen utama penyusun kayu yang tergolong ke dalam senyawa fenolik yang sangat mudah teroksidasi. Pada sisi lain, limbah organik terklorinasi yang dihasilkan dari proses bleaching sulit untuk didegradasi atau didaur ulang sehingga berpotensi sebagai bahan pencemar yang membahayakan lingkungan. Lebih dari sepuluh tahun terakhir ini, telah ditempuh beberapa cara untuk untuk menekan limbah berbahaya dalam industri pulp yaitu melalui pengolahan pulp dengan memanfaatkan mikroba (biopulping) maupun modifikasi proses
bleaching.
Pengolahan pulp secara biologi merupakan proses yang memanfaatkan mikroba yaitu jamur pelapuk putih (white rot fungi) untuk melemahkan struktur kayu melalui cara degradasi lignin sehingga akan mengurangi pemakaian bahan kimia dan energi didalam pemisahan serat (Siagian et al. 2003). Modifikasi proses
Program minimisasi limbah dalam industri kertas yang efektif akan mengurangi biaya produksi dan beban pengelolaan limbah berbahaya sehingga akan meningkatkan efisiensi dan kualitas produk. Teknik minimisasi limbah yang dapat dilakukan selain yang terkait langsung dengan proses produksi seperti perencanaan produksi dan tahapannya, penyesuaian peralatan/proses atau modifikasi, pemisahan limbah dan daur ulang limbah, penggantian (substitusi) bahan baku juga merupakan salah satu alternatif minimisasi limbah (Setyorini 2002). Substitusi bahan baku yang dapat diterapkan untuk efisiensi produksi pulp adalah penggunaan bahan baku dengan komposisi lignin yang memudahkan proses pulping sehingga dapat menekan biaya produksi.
Struktur lignin
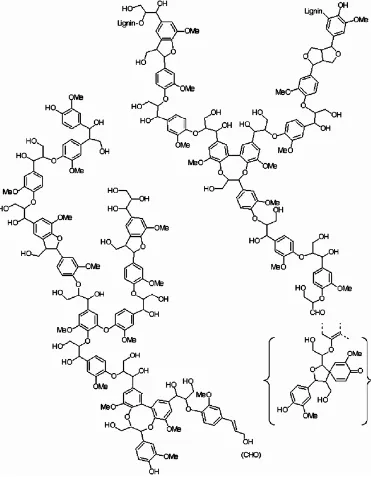
Lignin merupakan komponen penyusun dinding sel tumbuhan (17-33%) dengan komposisi bahan penyusun yang berbeda-beda bergantung jenisnya. Lignin adalah polimer dari unit fenilpropana: unit guaiasil (G) dari prekusor trans-coniferyl-alcohol, siringil (S) unit dari trans-sihapyl-alcohol, dan p-hydroxyphenyl (H) unit dari prekursor trans-p-coumaryl alcohol. Lignin terutama terakumulasi pada lignin berfungsi sebagai bahan pengikat komponen penyusun lainnya, sehingga suatu pohon bisa berdiri tegak. Berbeda denga gugus Gugus fenilpropana kebanyakan adalah ikatan eter (lebih dari 2/3) dan sisanya dengan ikatan karbon-karbon.
Senyawa ini dikelompokkan ke dalam 3 grup berdasarkan unit monomer penyusunnya (Gambar 3) yaitu gymnosperm lignin (disusun oleh monomer coniferil alkohol), angiosperm lignin (disusun oleh monomer coniferil alkohol dan sinapil alkohol) serta grass lignin (terdiri dari campuran monomer coniferil alkohol, sinapil alkohol dan ρ-coumaril alkohol) (Higuchi 1980).
sedang (18-33%) dan lignin rendah (18%) (Pari 1996). Kadar lignin kayu sengon umumnya adalah 26.8%, dan pada penelitian Pari et al. (1997) kadar lignin bervariasi tergantung umur yaitu pada sengon yang berumur 5, 10 dan 15 tahun kadarnya berturut-turut adalah 29.10%, 29.79% dan 30.19%.
Gambar 3. Struktur kimia penyusun lignin (A) p-koumaril alkohol, (B) koniferil alkohol, (C) sinafil alkohol (Fengel dan Wegener 1995).
Lignin dalam kayu terutama terdapat dalam lamela tengah dan dinding sel sekunder (Fengel & Wegener 1995). Lignin bersama-sama dengan selulosa merupakan suatu komponen penting pada tumbuhan berpembuluh dan dapat ditemukan dalam jumlah yang besar pada dinding sel sekunder, serat dan pembuluh angkut xilem. Fungsi lignin dalam tumbuhan selain sebagai penunjang mekanik juga sangat penting dalam membantu pertahanan tumbuhan terhadap patogen.
Komposisi lignin di alam sangat bervariasi tergantung pada spesies tanaman, yang dapat dikelompokkan berdasarkan kayu daun jarum, kayu daun lebar, dan rumput-rumputan. Kayu lunak terutama tersusun atas unit guaiasil, sedangkan kayu keras juga tersusun atas unit siringil. Kayu lunak ditemukan lebih resisten untuk didelignifikasi dengan ekstraksi basa daripada kayu keras. Hal ini diduga karena guaiasil lignin membatasi pemekaran (swelling) serat dan dengan demikian menghalangi serangan enzim terhadap siringil lignin. Beberapa studi yang terbaru mengenai lignin ditemukan bahwa terdapat struktur lignin yang
bermacam-macam seperti terdiri dari daerah amorphous dan bentuk-bentuk terstruktur seperti partikel tabung dan globul. Ada indikasi pula bahwa struktur kimia dan dimensi tiga lignin sangat dipengaruhi oleh matrik polisakarida. Simulasi dinamik menunjukkan bahwa gugus hidroksil dan metoksil di dalam prekusor lignin dan oligomer mungkin berinteraksi dengan mikrofibril selulosa sejalan dengan fakta bahwa lignin memiliki karakteristik hidrofobik. Sebagai contoh ditemukan bahwa tipe ikatan utama lignin di dalam kayu spruce adalah ikatan eter-aril eter. Gugus fungsional yang mempengaruhi reaktifitas lignin meliputi gugus phenolic hydroxyl bebas, methoxyl, benzylic hydroxyl, benzyl alcohol, noncyclic benzyl ether dan carbonyl. Guaiasil lignin mengandung lebih banyak gugus phenolic hydroxyl dibanding siringil.
Lignin pada kayu daun lebar disusun terutama oleh unit guaiasil dan siringil. Kadar lignin dan rasio siringil/guaiasil (S/G) berbeda-beda tergantung jenis dan lapisan selnya. Perbedaan rasio S/G ini mempengaruhi sifat kimia dan kecepatan degradasinya pada proses pulping sehingga sangat penting untuk mengkarakterisasi lokalisasi unit guaiasil dan siringil pada dinding sel kayu keras (Watanabe 2004). Berdasarkan komposisi unit strukturalnya, lignin diklasifikasikan kedalam beberapa tipe. Lignin pada kayu daun jarum atau disebut lignin guaiasil atau G lignin sebagian besar disusun oleh unit guaiasil (sekitar 90%) dan p-kumaril alkohol (sekitar 10%). Lignin pada kayu daun lebar atau disebut lignin guaiasil siringil atau G-S lignin disusun oleh unit guaiasil dan siringil dengan perbandingan tertentu, tergantung dari jenis kayu, umur kayu, tempat tumbuh dan iklim (Davin & Lewis 2005).
Kompleksitas struktur lignin hingga kini masih belum jelas. Namun demikian struktur dominan lignin telah diteliti seiring dengan semakin berkembangnya metoda identifikasi degradasi produk dan model sintesis. Beberapa studi berhasil menjelaskan repesentasi struktur lignin. Diketahui bahwa ikatan antar unit fenilpropana dan berbagai gugus fungsi menyebabkan lignin memiliki stuktur yang unik dan kompleks (Gambar 4). Ikatan yang dominan pada
lignin mengandung gugus methoxyl, phenolic hydroxyl serta sedikit gugus aldehid terminal. Hanya sebagian kecil saja proporsi gugus phenolic hydroxyl yang bebas karena sebagian besar terikat dengan fenilpropana lainnya. Gugus-gugus karbonil dan hidroksil alkohol terikat dalam struktur lignin ketika proses dehidrogenasi enzimatik.
Biosintesis lignin dan enzim-enzim terkait
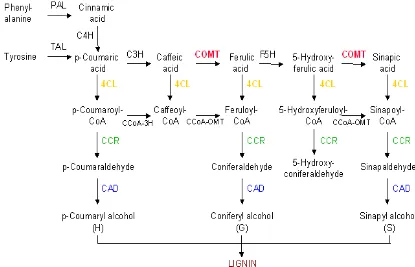
Lignin terbentuk dari polimerisasi dehidrogenasi monolignol p-koumaril alkohol, koniferil alkohol, and sinapil alkohol. Monolignol tersebut disintesis melalui jalur biosintesis fenilpropanoid yang diinisiasi dari deaminasi fenil alanin oleh enzim fenilalanin ammonia liase (Zong & Morrison 2000a; Harakava 2005).
Pada beberapa tanaman telah diketahui enzim-enzim yang berkaitan dengan biosintesis lignin diantaranya phenylalanine ammonia–lyase (PAL) (Kao
[image:32.595.86.501.378.645.2]et al. 2002), o-methyltransferase (CCoAoMT) (Ibrahim et al. 1998; He et al. 1998); 4-coumarate CoA ligase (4CL) (Allina et al 1998; Ehlting et al. 1999; Chukovic et al. 2000; Ehlting et al. 2001: Rogers et al. 2005), cinnamoyl-CoA reductase (CCR) dan cinnamyl alcohol dehydrogenase (CAD) (Ralph et al. 1998). Enzim-enzim tersebut terlibat di dalam jalur biosintesis lignin yang dimulai dari konversi prekursor fenil alanin hingga pembentukan monolignol (Gambar 5).
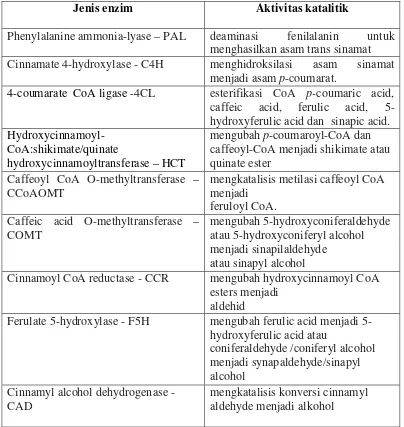
Gen-gen penyandi enzim yang berkaitan dengan biosintesis lignin telah diisolasi dan dikarakterisasi. Saat ini sudah banyak dilaporkan data sekuen DNA yang berupa fragmen cDNA ataupun sekuen gen utuhnya. Sekuen yang terdaftar pada data gene bank jumlahnya sangat banyak hingga mencapai ratusan jenis sekuen dari berbagai macam tanaman juga organisme lainnya. Berdasarkan studi EST (Expressed Sequence Tag), diketahui bahwa masing-masing enzim memiliki aktivitas terhadap substrat yang spesifik (Tabel 1).
Tabel 1. Aktivitas katalitik beberapa enzim yang terlibat dalam biosintesis lignin (Harakava 2005).
Jenis enzim Aktivitas katalitik
Phenylalanine ammonia-lyase – PAL deaminasi fenilalanin untuk menghasilkan asam trans sinamat
Cinnamate 4-hydroxylase - C4H menghidroksilasi asam sinamat
menjadi asam p-coumarat.
4-coumarate CoA ligase -4CL esterifikasi CoA p-coumaric acid, caffeic acid, ferulic acid, 5-hydroxyferulic acid dan sinapic acid.
Hydroxycinnamoyl-CoA:shikimate/quinate
hydroxycinnamoyltransferase – HCT
mengubah p-coumaroyl-CoA dan caffeoyl-CoA menjadi shikimate atau quinate ester
Caffeoyl CoA O-methyltransferase – CCoAOMT
mengkatalisis metilasi caffeoyl CoA menjadi
feruloyl CoA. Caffeic acid O-methyltransferase –
COMT
mengubah 5-hydroxyconiferaldehyde atau 5-hydroxyconiferyl alcohol menjadi sinapilaldehyde
atau sinapyl alcohol
Cinnamoyl CoA reductase - CCR mengubah hydroxycinnamoyl CoA esters menjadi
aldehid
Ferulate 5-hydroxylase - F5H mengubah ferulic acid menjadi 5-hydroxyferulic acid atau
coniferaldehyde /coniferyl alcohol menjadi synapaldehyde/sinapyl alcohol
Cinnamyl alcohol dehydrogenase - CAD
Modifikasi transgenik komposisi lignin kayu dan prospek proses pulping
yang lebih efisien
Pada proses pembuatan pulp lignin harus dihilangkan untuk memperoleh serat selulosa. Pemisahan lignin dari selulosa memerlukan bahan kimia dan energi dalam jumlah yang besar sehingga biaya yang diperlukan juga tinggi disamping menimbulkan resiko pencemaran lingkungan. Banyak upaya yang telah dilakukan untuk mengembangkan varitas bibit pohon dengan kadar lignin rendah atau termodifikasi untuk mendukung efisiensi produksi pulp. Modifikasi transgenik dengan cara mengatur ekspresi gen-gen terkait dengan metabolism fenilpropanoid sangat memungkinkan untuk dikembangkan guna mendapatkan kayu dengan kadar lignin rendah atau yang lebih mudah didelignifikasi.
Perbaikan genetik tanaman berkayu seperti tanaman kehutanan dan buah-buahan dibatasi oleh berbagai faktor antara lain ukuran pohon, siklus hidup yang panjang dan kurangnya informasi mengenai basis genetik. Teknologi DNA dapat menjadi solusi untuk mengatasi keterbatasan tersebut. Penelitian mengenai transgenik tanaman tinggi ditemukan lebih dari 100 laporan yang meliputi ketahanan terhadap herbisida dan serangga (Strauss & Bradshaw 2001) dan modifikasi lignin tanaman berkayu melalui represi ekspresi enzim yang berkaitan dengan biosintesis lignin (Hauffe 1993; Kajita 1997; Lee 1997; Zhong et al.
2000b). Pendekatan untuk menekan biosintesis lignin adalah dengan memanfaatkan fenomena PTGS (Post Trancriptional Gene Silencing) yang dapat dilakukan diantaranya dengan konstruk antisense.
Pada tanaman model Arabidopsis transformasi konstruk antisense dapat menurunkan aktivitas enzim 4CL hingga tersisa 8% dan menyebabkan perubahan rasio G/S lignin (Lee et al. 1997). Bahkan pada tanaman kehutanan transgenik subtropis yang cepat tumbuh yaitu aspen (Populus tremuloides) mengandung gen penyandi 4CL yang dikonstruksi secara antisense sehingga terjadi down regulated
ekspresi 4CL menyebabkan turunnya kadar lignin hingga 45% dan kadar selulosa meningkat hingga 15%, selain itu terbukti pula memacu pertumbuhan daun, akar dan batang (Sederoff 1999, Harding et al. 1999).
mudah dipisahkan dari selulosa yang akan sangat menguntungkan bagi industri pengolahan pulp karena akan menghemat energi dan biaya disamping dapat menekan limbah berbahaya. Siringil lignin yang tersusun atas monomer sinapil alkohol lebih peka terhadap degradasi kimia maupun enzimatik dibanding guaiasil lignin, karena unit posisi C5 aromatik pada guaiasil bebas berikatan antar karbon. Ferulate 5-hydroxylase (F5H) terlibat dalam jalur sintesis 5-hidroksi koniferaldehid berupa prekursor sinapilalkohol sebagai penyusun siringil lignin.
Over ekspresi ferulate 5-hydroxylase Arabidopsis pada poplar, menghasilkan kayu poplar transgenik dengan kadar siringil lignin tinggi yang dapat meningkatkan efisiensi proses pulping yaitu dalam hal peleraian selulosa dari lignin (U.S. Departement of Energi 2006). Over ekspresi F5H sweetgum dibawah kontrol promotor xylem-specific Pt4CLIP pada aspen transgenik berhasil meningkatkan rasio siringil/guaiasil (S/G) hingga 2.5 kali (Baucher et al. 2003). Demikian pula over ekspresi ferulate 5-hydroxylase Arabidopsis pada poplar, menghasilkan kayu poplar transgenik dengan kadar siringil lignin tinggi yang dapat meningkatkan efisiensi pulping yaitu dalam hal peleraian selulosa dari lignin (Boudet et al. 2003).
Perbaikan sifat tanaman dengan cara konvensional yaitu melalui hibridisasi seksual (persilangan) akan membutuhkan waktu lama terutama karena panjangnya siklus hidup tanaman kehutanan. Dengan demikian usaha perbaikan sifat tanaman melalui teknologi DNA atau rekayasa genetika untuk memodifikasi kadar lignin kayu sengon akan lebih menguntungkan karena modifikasi terjadi dalam waktu relatif singkat.
Pendekatan untuk menekan biosintesis lignin adalah dengan memanfaatkan fenomena PTGS (Post Transkriptional Gene Silencing) yang dapat dilakukan diantaranya dengan konstruk antisense dan RNAi (RNA interference) yang selanjutnya ditransformasikan pad tanaman. Efektivitas PTGS yang dihasilkan melalui teknik antisense umumnya sekitar 50%. Knock out gen dengan teknik RNAi dapat lebih tinggi dibanding antisense, misalnya efek silencing beberapa gen (GUS, PVY dan FAD2∆12-desaturase) pada tanaman tembakau, arabidopsis,
Pada Pinus radiata konstruk RNAi 4CL dapat menurunkan kadar lignin total hingga 36% - 50% dan rasio S/G meningkat, tetapi pada beberapa individu bentuk fenotifiknya menjadi kerdil (Wagner et al. 2009) . Mengingat peranannya yang penting dalam struktur dinding sel dan ketahanan terhadap hama dan penyakit, penurunan kadar lignin kayu bisa dilakukan hingga batas tertentu yang tidak mengganggu fenotif dan kekuatan pohon.
ANALISIS KUANTITATIF DAN UJI HISTOKIMIA LIGNIN
SENGON (
Paraserianthes falcataria
)
Abstract
The presence of lignin in plant cells is a factor limiting the efficiency of processing lignocellulosic materials of wood-based industries including pulp and paper industry either chemically or biologically (biopulping) process using white rot fungi. Lignin content determination of sengon collected from different areas in Indonesia indicated that it ranged low to moderate category (16.58 – 35.59%). Lignin histochemical assay of transverse section of stems using phloroglucinol-HCl staining showed that the initiation of lignin deposition was noted in 2 weeks old seedling. Lignin quantitative and qualitative assessment through histochemical assay showed that lignin content was varied in trees depending on height.
Keywords: Paraserianthes falcataria, lignin, histochemical, phloroglucinol-HCl, pulp
Abstrak
Keberadaan lignin pada sel tanaman merupakan faktor pembatas efisiensi pengolahan material lignoselulosa menjadi produk-produk industri berbahan dasar kayu termasuk pulp baik secara kimia maupun biologis (biopulping) menggunakan jamur pelapuk putih. Pengujian kadar lignin yang dikoleksi dari beberapa daerah menunjukkan bahwa kadar lignin sengon termasuk kategori rendah hingga sedang (16.58% - 35.59%). Uji histokimia lignin pada potongan transversal batang dengan pewarnaan phloroglucinol-HCl menunjukkan bahwa deposisi lignin tampak jelas dimulai pada umur 2 minggu. Pengujian kadar lignin secara kuantitatif menggunakan metoda Klason dan kualitatif melalui uji histokimia jaringan, menunjukkan bahwa deposisi lignin berbeda pada ketinggian pohon yang berbeda.
Pendahuluan
Industri pulp terus berkembang dan produksinya cenderung meningkat dari tahun ke tahun seiring dengan bertambahnya konsumsi kertas dunia. Pertumbuhannya dalam dekade yang akan datang diperkirakan antara 2% hingga 3.5% per tahun, sehingga membutuhkan kenaikan kayu log yang dihasilkan dari lahan hutan seluas 1 sampai 2 juta hektar setiap tahun. Kondisi ini menuntut tersedianya bahan baku yang cukup dan kontinyu khususnya kayu dari berbagai jenis tanaman kehutanan sebagai bahan baku yang paling banyak digunakan.
Kayu sebagai bahan dasar dalam industri kertas mengandung beberapa komponen antara lain selulosa, hemiselulosa, lignin dan zat ekstraktif. Di dalam proses industri pulp secara kimia seperti proses sulfat/Kraft, memerlukan proses
bleaching karena pulp yang dihasilkan berwarna gelap yang disebabkan oleh oksidasi senyawa lignin. Kandungan lignin pada sel tanaman (monomer guaiasil dan siringil) berpengaruh terhadap pelepasan dan hidrolisis polisakarida. Pada
biopulping, asosiasi lignin dengan selulosa membentuk suatu matriks hidrofobik yang membatasi kerja enzim-enzim hidrolitik.
Lignin adalah polimer penyusun biomassa tanaman yang kelimpahannya menduduki urutan kedua setelah selulosa. Pada tanaman tingkat tinggi senyawa ini memiliki peran penting pada kekutan pohon, transport air dan ketahanan terhadap penyakit (Lappierre et al. 1999). Pemisahan lignin dari selulosa merupakan proses dasar pada Kraft pulping yaitu proses yang banyak digunakan pada pembuatan pulp. Pada berbagai spesies kayu kadar lignin bervariasi antar 15 - 36% (Doorsselaere et al. 1995).
tahan robeknya (tensile strength dan bursting strength) yang tinggi yang mungkin disebabkan karena kayu sengon seratnya panjang.
Dalam proses pulping secara kimia, delignifikasi merupakan proses terpenting karena bertujuan untuk mendegradasi dan melarutkan lignin sebanyak mungkin dan menghindari kerusakan pada serat selulosa seminimal mungkin. Perbedaan laju delignifikasi tidak hanya dipengaruhi oleh kadar lignin tetapi juga oleh reaktivitas komponen penyusun lignin. Walaupun sifat-sifat fisik kayu sengon sangat baik sebagai bahan baku industri pulp, kayu sengon bukan merupakan bahan baku pulp yang digunakan secara luas karena berat jenisnya yang relatif rendah dibanding jenis kayu pulp lainnya. Untuk meningkatkan penggunaan kayu sengon sebagai bahan baku pulp karena sifatnya yang tumbuh sangat cepat dan sifatnya fisik kayunya yang menguntungkan, diperlukan upaya pemuliaan pohon untuk memodifikasi kadar maupun komposisi sub unit lignin yang merupakan faktor pembatas perolehan rendemen selulosa yang tinggi. Berkaitan dengan hal ini diperlukan studi pendahuluan mengenai analisis kadar lignin kayu dan waktu mulainya pembentukan lignin pada sengon. Lignin terdapat pada semua sel jaringan tumbuhan, akan tetapi banyak ditemukan pada jaringan sklerenkim (Soukupova et al. 2000). Keberadaan lignin tersebut secara kualitatif dalam suatu jaringan tumbuhan dapat diketahui dengan metode histokimia menggunakan pewarnaan phloroglucinol-HCl 1%. Section (irisan penampang melintang) suatu akar atau batang tanaman yang dicelupkan ke dalam larutan phloroglucinol-HCl 1% akan memberikan warna merah pada ligninnya (Valette
et al. 1998).
Penelitian mengenai perbandingan keberadaan lignin pada dinding sel kayu telah dilakukan pada beberapa spesies tanaman diantaranya Arabidopsis thaliana (Zhong et al. 2000b), spruce (Soukovova et al. 2000) dan Eucalyptus
Bahan dan metode
Analisis kadar lignin kayu sengon dilakukan terhadap kayu dari tanaman sengon yang dikoleksi dari beberapa lokasi yaitu koleksi kebun plasma nutfah Puslit Bioteknologi LIPI di Cibinong, Kebun Percobaan Puslitbang Hutan-Kementrian Kehutanan di Cikampek, kebun koleksi sengon Dinas Kehutanan dan Perkebunan Tasikmalaya serta koleksi sengon Pusat Penelitian Hutan Tanaman-Kementrian Kehutanan di Yogyakarta. Umur tanaman yang dianalisis hampir sama yaitu sekitar 10 tahunan. Selain itu pohon yang dipilih untuk dianalisis kadar ligninnya merupakan pohon yang memiliki respon pertumbuhan paling baik pada masing-masing lokasi tumbuhnya.
Sengon yang dikoleksi dari kebun plasma nutfah Puslit Bioteknologi-LIPI (Cibinong) merupakan pohon plus, yang diuji dari 105 pohon yang diseleksi berdasarkan berbagai parameter yaitu diameter, tinggi, Diameter breast height
(DBH), bentuk batang, bentuk percabangan, sudut percabangan, pembuahan, ketahanan hama dan penyakit serta cacat lain (Hartati et al. 2007). Pada setiap lokasi penelitian masing-masing dianalisis sebanyak 5 pohon. Tinggi dan diameter pohon yang diuji adalah sekitar 30-37 m dan 40-70 cm. Kayu di koleksi dengan cara mengambil kayu beserta kulit dengan ukuran 20 x 20 cm dan ketebalan ±3 cm, pada dua titik yaitu pada ketinggian pohon 2 m dan 4 m dari pangkal batang. Selain uji kuantitatif kadar lignin dilakukan pula uji histokimia lignin terhadap irisan melintang hipokotil bagian atas, tengah dan bawah bibit sengon yang berasal dari Kebun Botani Serpong pada umur 3 hari, 1 minggu dan 2 minggu, 1 bulan dan 2 bulan untuk mengetahui distribusi lignin pada bagian dan umur bibit yang berbeda.
Uji kadar lignin
dalam gelas piala yang berisi air dan dilakukan pengadukan pada 200 rpm selama 2.5 jam. Selama pengadukan suhu air dipertahankan pada 20±1oC. Kemudian sampel dipindahkan kedalam Erlenmeyer 500 ml dan ditambah dengan akuades sebanyak 171 ml dan ditutup dengan aluminium foil. Sampel diautoklaf pada 121oC selama 15 menit. Selanjutnya disaring dalam keadaan panas menggunakan gelas filtrate. Sebelum digunakan gelas filtrate dioven pada 105oC selama 24 jam, didinginkan dalam desikator selama 2 jam dan ditimbang. Gelas filtrate dicuci berturut-turut dengan 20 ml air dan acetone dan selanjutnya di oven pada 105oC selama 24 jam. Setelah dioven, sample didinginkan dalam desikator.
Kadar lignin ditetapkan berdasarkan perhitungan sebagai berikut: Kadar lignin = C – A X 100
(100%-Ka) X B
A: Berat gelas awal (sebelum penyaringan), B: Berat sample bebas ekstraktif, C: Berat gelas filtrate setelah digunakan untuk menyaring
Ka: Kadar air sampel
Uji histokimia lignin
Hasil dan pembahasan
Analisis kadar lignin
Secara umum kadar sengon dari seluruh lokasi yang diuji pada ketinggian pohon 2 m adalah (17.93 -34.11%) dengan rata-rata 27.29% dan pada ketinggian 4 m adalah (16.58 – 35.59%) dengan rata-rata 27.73%. Sengon yang memiliki kadar lignin terendah adalah pohon PII yang merupakan koleksi dari Cibinong dengan kadar lignin rata-rata pada ketinggian 2 m dan 4 m adalah 17.25%. Kadar lignin tersebut termasuk kategori kayu rendah lignin yaitu kurang dari 18% (Pari 1996). Selain memiliki kadar lignin yang rendah pohon ini juga kadar selulosanya cukup tinggi yaitu 51.54% (Hartati et al. 2009).
Berdasarkan hasil analisis kadar lignin pohon PII tersebut berpotensi untuk dibudidayakan dan diperbanyak sebagai bahan baku pulp yang efisien ataupun sebagai material pemuliaan tanaman untuk menurunkan lignin hingga kadar yang lebih rendah lagi tetapi respon pertumbuhan dan daya hasilnya tetap menguntungkan. Perbanyakan koleksi pohon unggul rendah lignin tersebut dapat dilakukan melalui stek ataupun propagasi in vitro untuk menjamin kesamaan identitas genetiknya.
Tabel 2. Kadar lignin kayu sengon yang dikoleksi dari daerah Cibinong, Cikampek, Tasikmalaya dan Yogyakarta.
No. Asal koleksi Kode Pohon
Kadar lignin (%) pada ketinggian: 2 m 4 m
1 Cibinong P I
P II P III P IV P V 26.57 17.93 25.63 22.53 21.80 25.26 16.58 16.96 24.13 25.34 Rata-rata 22.89 21.65
2 Cikampek 94
56 57 39 75 32.88 19.68 34.11 31.79 29.25 33.10 33.62 28.25 31.93 34.44 Rata-rata 29.54 32.27 3 Tasikmalaya TSM 1
TSM 2 TSM 3 TSM 4 TSM 5 28.95 27.79 31.95 31.69 24.14 35.59 30.38 29.61 26.91 32.78 Rata-rata 28.9 31.05 4 Yogyakarta CND1
CND3 CND4 CND5 CND6 29.47 29.04 23.79 26.77 30.01 21.30 32.09 22.75 24.11 29.47 Rata-rata 27.82 25.94 Rata-rata total 27.29 27.73
Uji histokimia lignin bibit sengon umur 3 hari, 1 minggu, 2 minggu, 1 bulan
dan 2 bulan
Hal ini sesuai dengan uji histokimia pada Picea abis dengan pewarnaan phloroglucinol-HCl yang menunjukkan bahwa kandungan senyawa lignin banyak ditemui didaerah sklerenkim, hipodermis dan sel atau jaringan vaskular (Soukupova, 2000). Berdasarkan penelitian Watanabe et al. (2004) tentang studi histokimia distribusi lignin pada dinding sel dari dua spesies Eucalyptus yaitu E. calmadulins dan E. globulus dengan metode Klason menunjukkan bahwa dinding sel E. calmadulins mengandung gugus guaiasil dan siringil dan pada vessel walls
umumnya mengandung gugus guaiasil. Sedangkan E. globulus dinding selnya hanya mengandung gugus siringil sedangkan pada vessel walls mengandung gugusguasil dan siringil.
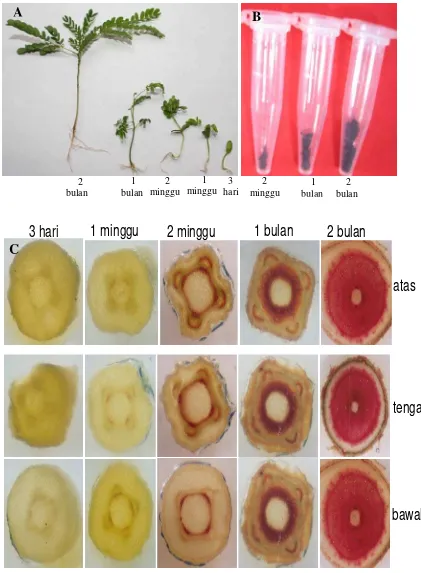
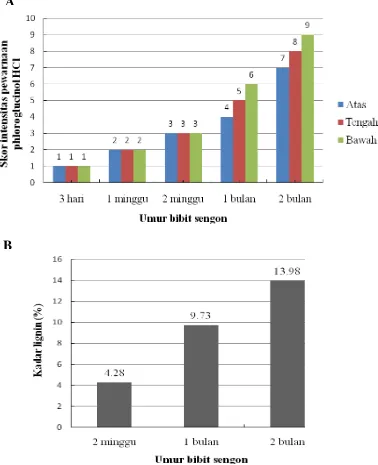
Pewarnaan lignin mulai tampak pada bibit umur 2 minggu yang semakin tinggi intensitasnya pada umur 1 bulan dan 2 bulan (Gambar 6). Intensitas pewarnaan lignin pada setiap bagian yang diuji (atas, tengah, bawah) pada bibit umur 1 dan 2 bulan berbeda dimana pewarnaan lignin dengan intensitas tertinggi tampak pada bagian bawah (Gambar 7). Pada umur bibit 2 minggu, walaupun sudah terdekteksi adanya pembentukan lignin, tetapi intensitas warna pada ketiga bagian yang diuji skornya sama (Gambar 7). Seperti halnya hasil uji histokimia yang menunjukkan meningkatnya deposisi lignin seiring bertambahnya umur tanaman, kadar lignin Klason juga meningkat pada bibit umur 2 minggu, 1 dan 2 bulan yaitu masing-masing 4.28 %, 9.73% dan 13.98% (Gambar 7). Perbedaan distribusi dan deposisi lignin pada bagian tanaman dan umur berbeda juga terjadi pada bagian atas, tengah dan bawah dari ruas ke- 10 tanaman jagung yang diamati pada 3, 5, 7,9, 11, 13, dan 15 hari (Morrison et al. 1994). Hasil penelitian tersebut menunjukkan bahwa pada tanaman jagung, terjadi peningkatan lignin hingga 50% pada pengamatan 15 hari dibanding 3 hari dengan kadar tertinggi diperoleh pada bagian bawah ruas tanaman. Pada penelitian mengenai perkembangan xilem dan dinding sel kecambah kedelai umur 5 hari menunjukkan adanya deposisi lignin yang diamati dengan epi-fluorescen microscopy
Gambar 6. Isolasi dan uji histokimia lignin bibit sengon berbagai umur. A. Bibit sengon yang digunakan untuk uji histokimia dan uji kadar lignin, B. isolat lignin untuk pengujian kadar lignin, C. Profil histokimia bibit sengon.
3 hari
1 minggu
atas
tengah
bawah
1 bulan
2 bulan
2 minggu
2 bulan 1
bulan 2
minggu
A
B
C
1 bulan 2
bulan
1 minggu
3 hari 2
Gambar 7. Intensitas pewarnaan histokimia lignin dengan phloroglucinol-HCl (A) dan kadar lignin kecambah sengon pada berbagai umur (B).
Pada jaringan kayu sengon umur 1 dan 10 tahun, pewarnaan phloroglucinol menghasilkan warna merah yang sangat jelas. Percobaan uji histokimia pada berbagai umur bibit dan pohon sengon akan berguna untuk menguji kadar lignin secara kualitatif kadar lignin sengon hasil transformasi dengan konstruk antisense gen penyandi enzim yang berkaitan dengan biosintesis lignin seperti halnya yang dilakukan untuk menguji kadar lignin tembakau dan poplar transgenik rendah lignin (Sewalt et al. 1997 dan Zhong et al. 2000a ).
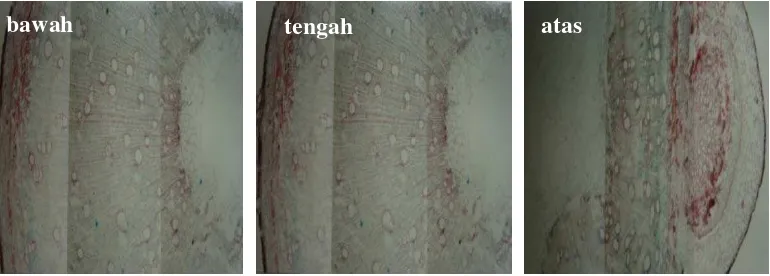
A
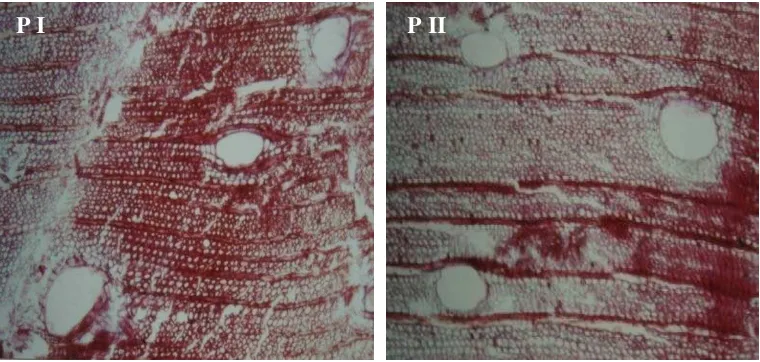
Perbedaan intensitas warna lignin pada sengon umur 1 tahun pada ketiga bagian yang diuji tidak tampak jelas (Gambar 8.), tetapi pada bibit umur 2 tahun bagian pangkal memiliki intensitas warna yang lebih tinggi (Gambar 9). Adanya variasi intensitas warna pada tiga bagian kayu yang diuji yaitu atas, tengah dan bawah, mengindikasikan adanya perbedaan kadar lignin yang berbeda tergantung ketinggian atau jarak pengambilan sampel dari pangkal batang atau bibit. Uji kualitatif deposisi lignin secara histokimia dengan phloroglucinol-HCl sesuai pula dengan data kuantitatif sebagaimana pada hasil uji histokimia sengon PII dibanding dengan PI, dimana PII yang memiliki intensitas pewarnaan lignin lebih lemah dari PI kadar ligninnya pun lebih rendah (Gambar 10). Distribusi lignin pada PI tampak lebih jelas dibandingkan PII.
[image:47.595.121.506.333.471.2]
Gambar 8. Profil histokimia lignin kayu sengon bagian atas, tengah dan bawah pada pohon sengon umur 1 tahun.
Gambar 9. Profil histokimia lignin kayu sengon bagian atas (a), tengah (b) dan bawah (c) pada pohon sengon umur 2 tahun.
atas tengah
bawah
[image:47.595.114.511.546.683.2]
Gambar 10. Profil histokimia lignin kayu sengon dewasa yang dikoleksi dari Cibinong.
Kesimpulan dan saran
Analisis kayu sengon yang dikoleksi dari daerah Cibinong, Cikampek, Tasikmalaya dan Yogyakarta menunjukkan bahwa kadar lignin sengon secara umum termasuk kategori kadar lignin sedang dengan kisaran (16.58 -35.59%). Sengon dengan kadar lignin rendah teridentifikasi pada koleksi sengon asal Cibinong yang merupakan kandidat pohon plus. Berdasarkan uji histokimia tampak bahwa pembentukan lignin dimulai pada bibit sengon umur 2 minggu. Baik melalui pengujian kadar lignin secara kuantitatif maupun uji histokimia menunjukkan bahwa deposisi lignin pada setiap bagian batang berbeda kadarnya berbeda pula. Sengon dengan kadar lignin rendah sangat berpotensi untuk diperbanyak dan dibudidayakan untuk mendukung ketersediaan bahan baku industri pulp dan kertas yang efisien dan ramah lingkungan. Data kualitatif melalui uji histokimia lignin yang menunjukkan waktu dimulainya deposisi lignin merupakan informasi yang sangat berguna untuk penelitian lebih lanjut terkait dengan upaya modifikasi kadar dan komposisi sub unit penyusun lignin yang menguntungkan untuk industri pulp dan kertas. Teknologi DNA rekombinan dengan mengatur tingkat ekspresi gen-gen yang berkaitan biosintesis lignin melalui Post transcription Gene Silencing (PTGS) dapat menjadi alternatif untuk upaya penurunan kadar lignin.
PRODUKSI TUNAS MELALUI INDUKSI EMBRIOGENESIS
DAN MULTIPLIKASI TUNAS NODAL KOTILEDON
SENGON
Abstract
Sengon micropropagation has been carried out using two methods i.e. embryogenesis induction of embryo axis and shoots induction of cotyledon node on MS medium containing various types and concentrations of plant growth regulators. Embryogenic callus could be induced using all media tested, but only on MS medium containing 0.1 mg/l TDZ and 0.25 mg/l IAA, the callus could develop into somatic embryos-like structures. The highest number and the longest shoots was obtained from cotyledon node-derived 10 days seedling on MS medium containing 1 mg/l TDZ. Shoots induction of cotyledon node was selected as the most appropriate system for genetic transformation to improve the sengon wood quality for pulp and paper industry.
Key words: embryogenesis, shoot induction, TDZ, IAA, P. falcataria
Abstrak
Mikropropagasi sengon telah dilakukan melalui dua jenis metoda yaitu induksi embryogenesis dari aksis dan induksi tunas dari nodal kotiledon pada media MS yang mengandung beberapa jenis dan konsentrasi ZPT. Kalus embriogenik dapat diinduksi pada semua media yang dicoba, namun hanya pada media MS yang mengandung 0.1 mg/l TDZ dan 0.25 mg/l IAA dapat berkembang menjadi struktur menyerupai embrio somatik. Tunas majemuk terbanyak dan rata-rata tunas terpanjang dapat diinduksi dari nodal kotiledon kecambah umur 10 pada media MS yang mengandung 1 mg/l TDZ. Dengan demikian induksi tunas menggunakan nodal kotiledon dapat dipilih sebagai sistem regenerasi pada transformasi genetik sengon untuk meningkatkan kualitas bibit sengon yang sesuai dengan kebutuhan indusri pulp dan kertas.
Pendahuluan
Propagasi tanaman melalui teknik in vitro dapat dilakukan melalui induksi somatik embrio maupun induksi tunas dari berbagai eksplan. Selain untuk perbanyakan masal bibit tanaman, teknik kultur jaringan digunakan pula untuk penyedian bahan tanaman untuk transformasi genetik. Terdapat beberapa zat pengatur tumbuh (ZPT) yang dapat digunakan untuk induksi tunas dari golongan sitokinin seperti BAP dan TDZ. Thidiazuron merupakan zat pengatur tumbuh yang sesuai untuk digunakan pada tanaman berkayu seperti Albizia sinensis
(Sinha et al. 2000). Selain itu dapat pula digunakan kombinasi ZPT. Jenis eksplan yang digunakan untuk induksi tunas diantaranya kotiledon (Ardiana 2010), nodal kotiledon (Rauf et al.2004; Aasim et al. 2010; Singh & Tiwari 2010), daun (Ozyigit 2009) dan ruas tunas batang (Semivay et al. 2005).
Regenerasi melalui embriogenesis somatik memiliki banyak keuntungan, antara lain waktu perbanyakan lebih cepat, pencapaian hasil dalam mendukung program perbaikan tanaman lebih cepat dan jumlah bibit yang dihasilkan tidak terbatas jumlahnya (Mariska 1996). Di samping itu, dengan strukturnya yang bipolar dan kondisi fisiologis yang menyerupai embrio, embrio somatik lebih menguntungkan daripada pembentukan tunas adventif yang unipolar (Sukmadjaja 2005).
Morfogenesis tunas dan embriogenesis somatik merupakan dua proses regenerasi yang berbeda dan keduanya sangat bergantung kepada sumber eksplan dan jenis media kultur yang digunakan. Morfogenesis tunas (organogenesis) merupakan proses pembentukan dan perkembangan tunas dari jaringan meristem tunas. Tunas selanjutnya dapat diakarkan untuk mendapatkan tanaman utuh, sedangkan embriogenesis somatik merupakan proses regenerasi tanaman melalui pembentukan struktur menyerupai embrio (embrioid) dari sel-sel somatik yang telah memiliki calon akar dan tunas (serupa embrio zigotik). Tanaman utuh diperoleh dari hasil perkecambahan embrio somatik (Pardal et al. 2002).
pemeliharaan, pendewasaan, perkecambahan, dan aklimatisasi. Embrio somatik biasanya berasal dari sel tunggal yang kompeten dan berkembang membentuk fase globuler, hati, torpedo, dan akhirnya menjadi embrio somatik dewasa yang siap dikecambahkan membentuk planlet/tanaman utuh (Finer & Mc Mullen 1991, Finer et al. 1996).
Keberhasilan regenerasi melalui embriogenesis somatik dipengaruhi oleh berbagai faktor, antara lain formulasi media (sumber nitrogen dan gula serta zat pengatur tumbuh) yang berbeda pada setiap tahap perkembangan embrio somatik serta jenis eksplan yang digunakan (Purnamaningsih 2002). Perbanyakan tanaman melalui embriogenesis somatik memerlukan beberapa tahapan dengan formulasi media yang berbeda, bergantung pada tahapan perkembangan embrio somatik (Sukmadjaja 2005). Penggunaan eksplan yang bersifat meristematik umumnya memberikan keberhasilan pembentukan embrio somatik yang lebih tinggi. Eksplan yang digunakan dapat berupa aksis embrio zigotik muda dan dewasa, kotiledon, mata tunas, epikotil maupun hipokotil (Purnamaningsih 2002).
Penelitian ini bertujuan untuk memperoleh metoda yang tepat untuk penyedian bahan untuk trasnformasi genetik sengon guna modikasi komposisi lignin.
Bahan dan Metode
Bahan tanaman
Bahan tanaman yang digunakan untuk induksi embriognesis adalah aksis embrio yang diisolasi dari biji sengon yang berasal dari Kebun Botani Serpong dan terlebih dahulu dikecambahkan selama 1 hari, sedangkan tunas majemuk diinduksi dari nodal kotiledon kecambah sengon umur 7 dan 10 hari yang telah dihilangkan tunas terminalnya. Jumlah eksplan untuk setiap perlakukan adalah 30.
Sterilisasi biji
Dithane 4%, dibiarkan selama 30 menit sambil dikocok. Biji dibilas dengan akuades sampai bersih lalu ditambahkan larutan Masalgine 2% dan dibiarkan selama 10 menit sambil dikocok. Larutan Masalgine dibuang dan biji dibilas dengan aquades sampai bersih. Kemudian ditambahkan larutan etanol 70% sambil dikocok selama 5 menit. Setelah etanol dibuang, biji dibilas dengan akuades steril sampai bersih. Lalu ditambahkan larutan HgCl2 0.5 % selama 3 menit selanjutnya
biji sengon dibilas dengan akuades steril 3-5 kali hingga bersih.
Induksi embriogenesis
Aksis embrio yang diisolasi dari biji sengon yang sudah disterilkan dan dikultur selama 1 hari, dikultur secara langsung pada media MS yang mengandung dua jenis zat pengatur tumbuh yaitu Thidiazuron (TDZ) dan Indole Acetic Acid (IAA) dengan konsentrasi bervariasi, yang dipadatkan dengan 0.3% gelrite. Kultur diinkubasi pada ruangan yang bersuhu 25-26°C.
Induksi tunas majemuk dari buku kotiledon
Buku kotiledon dari kecambah sengon berumur 7 dan 10 hari dikultur pada media MS yang mengandung TDZ, BAP dan NAA dengan beberapa variasi konsentrasi. Kultur diinkubasi pada suhu 25-26°C.
Hasil dan Pembahasan
Induksi embriogenesis dari aksis embrio
0. 25 mg/l IAA kalus embriogenik dapat berkembang lebih lanjut menjadi embrio somatik pada umur 5 minggu (Gambar 11). Namun demikian embrio somatik ini tidak dapat berkembang menjadi planlet utuh.
Penelitian Tampubolon (2007) mengenai induksi kalus yang responsif embriogenik dari eksplan kotiledon dari kecambah biji sengon berumur 10 hari telah berhasil dengan menggunakan media MS yang ditambah dengan 1 mg/l TDZ dan 0.25 mg/l IAA. Tetapi kalus yang dapat berkembang lebih lanjut ke tahapan embriogenesis berikutnya frekuensinya masih rendah. Sehingga masih diperlukan penelitian untuk optimasi media induksi embriogenesis somatik pada sengon dengan mengkombinasikan TDZ dengan ZPT yang lain seperti IAA dengan jenis eksplan yang berbeda.
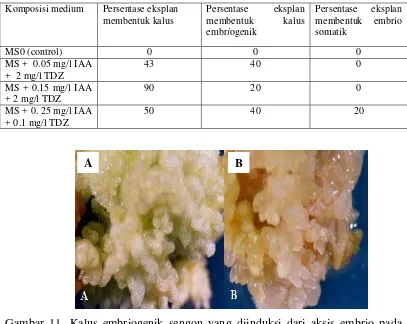
Tabel 3. Persentase pembentukan somatik embrio yang diinduksi dari aksis embrio sengon pada 4 jenis media.
Komposisi medium Persentase eksplan membentuk kalus
Persentase eksplan membentuk kalus embriogenik
Persentase eksplan membentuk embrio somatik
MS0 (control) 0 0 0
MS + 0.05 mg/l IAA + 2 mg/l TDZ
43 40 0
MS + 0.15 mg/l IAA + 2 mg/l TDZ
90 20 0
MS + 0. 25 mg/l IAA + 0.1 mg/l TDZ
[image:53.595.108.515.372.696.2]50 40 20
Gambar 11. Kalus embriogenik sengon yang diinduksi dari aksis embrio pada media MS + 0.05 mg/l IAA + 2 mg/l TDZ (A) dan MS + 0.15 mg/l IAA dan 2 mg/l TDZ (B) pada umur 1 bulan.
[image:54.595.182.417.102.357.2]
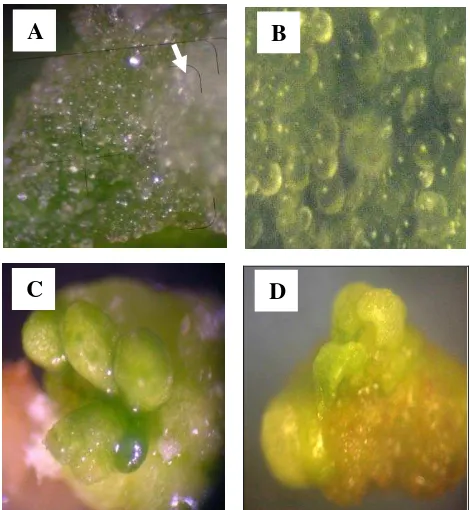
Gambar 12. Struktur kalus embriogenik dan embriosomatik yang diinduksi dari aksis embrio pada media MS + 0.1 mg/l TDZ + 0. 25 mg/l IAA pada umur umur 2 minggu (A), 4 minggu (B) , 5 minggu (C) dan 7 minggu (D).
Induksi tunas dari buku kotiledon
Ekplan nodal kotiledon baik dari kecambah umur 7 hari maupun 10 hari dapat diinduksi pembentukan tunas majemuknya pada semua media yang mengandung ZPT yang dicoba (Tabel 4) tetapi persentase pembentukan tunas dan jumlah tunasnya berbeda. Jenis media terbaik yang dapat menginduksi tunas terbanyak (rata-rata 5.87 tunas per eksplan) adalah media MS yang mengandung 1 mg/l TDZ dengan eksplan buku kotiledon dari kecambah umur 10 hari, selain itu kisaran panjang tunas yang terbentuk pun untuk jenis eksplan ini lebih besar dibanding media lain yang mengandung BAP atau BAP dan NAA (Tabel 4). Hasil penelitian serupa mengenai pengaruh BAP dan TDZ terhadap respon pembentukan tunas dari nodal kotiledon juga terjadi pada induksi tunas
Nyctanthes arbor-tritis L, dimana pada konsentrasi yang sama TDZ memberikan respon jumlah tunas yangh lebih tinggi dibanding BAP (Siddique et al. 2006).
Pembentukan tunas majemuk terjadi sejak minggu pertama yang dilanjutkan dengan penggandaan dan pendewasaan tunas yang berlangsung
A B
hingga minggu ke 7. Tunas selanjutnya dapat membentuk akar pada media 1/2MS pada minggu 8 dan dapat diaklimatisasi pada minggu ke 10 (Gambar 13). Planlet dapat berkembang lebih lanjut menjadi bibit sengon yang dipelihara pada media tanah (Gambar 14).
Tabel 4. Jumlah tunas yang terbentuk dari nodal kotiledon pada beberapa jenis dan komposisi media
Medium
Umur eksplan (hari)
7 10
Persentase eksplan membentuk tunas majemuk Jumlah rata-rata tunas /eksplan Kisaran panjang tunas (cm) Persentase eksplan membentuk tunas majemuk Jumlah rata-rata tunas /eksplan Kisaran panjang tunas (cm)
MS (Kontrol) 0 0 0 0 0 0
1 TDZ 100 2.3 0.2-1.5 100 5.87 0.4-2
1.5 TDZ 100 2.5 0.2-1.5 100 4.7 0.4-2
1.75 TDZ 100 3.6 0.2-1.5 100 5.4 0.4-2
1 BAP 73.3 0.9 0.2-0.4 100 1.8 0.2-0.4
1B + 0.1 N 16.7 0.5 0.2-0.5 100 3 0.2-0.7
3B + 0.1 N 100 2.4 0.2-0.5 72.7 1.64 0.2-0.6
3B + 0.5 N 0 0 0 100 3.16 0.2-0.5
Gambar 13. Induksi dan regenerasi tunas majemuk dari nodal kotiledon. A. tahap awal induksi tunas, B. inisiasi pembentukan tunas majemuk, C. multiplikasi tunas majemuk, D. pemisahan dan pendewasaan tunas , E. induksi perakaran, F. bibit