54
Ann. N.Y. Acad. Sci. 1066: 54–66 (2005). © 2005 New York Academy of Sciences. doi: 10.1196/annals.1363.005
Stability
FLORIN DESPA,a DENNIS P. ORGILL,b AND RAPHAEL C. LEEa
aDepartment of Surgery, The University of Chicago, Chicago, Illinois 60637, USA
bDepartment of Surgery, Brigham and Woman’s Hospital, Harvard Medical School,
Boston, Massachusetts 02115, USA
ABSTRACT: The volume fraction occupied by the dry matter of the cell can be as large as 40%, of which more than half (~60%) are proteins. Thus, cellular proteins and protein assemblies occupy a large volume that can have a pro-found effect on their own native-state stabilities and on their unfolding/refold-ing rates. In addition, macromolecular crowdunfolding/refold-ing can change the properties of a significant fraction of the water in the cell. We review features of the molec-ular crowding effect which are relevant for describing the microscopic mecha-nism of thermal injuries.
KEYWORDS: crowding effects; protein denaturation; thermal injury
INTRODUCTION
To a large extent cells are made of proteins, which constitute more than half (~60%) of the dry weight of the cell.1 Proteins determine the structure of the cell and, more importantly, they represent the physical apparatus which performs designed functions in the cell. Specific proteins, such as actin and myosin, are organized in large macromolecular arrays (e.g., cytoskeleton fibers) and play the essential role in shaping the cell. Besides proteins, the interior of cells contains several other kinds of macromolecules like lipids, sugars, and nucleic acids. Because no single macromo-lecular species may be present at high concentration, but all species taken together oc-cupy a significant fraction of the volume of the medium, such media are referred to as “crowded.” The volume fraction occupied by the dry matter of the cell (FIG. 1) can be as large as ϕ = 0.4. The large volume occupied by these crowding agents can have profound effect on the native state stability and unfolding/refolding rates of cellular proteins. Molecular crowding is considered as a source of nonspecific interactions between cellular proteins. Steric repulsion is the most common of all interactions between macromolecules and is always present in crowded environments, inde-pendent of the magnitude of the general electrostatic and hydrophobic interactions.
Because molecules are mutually impenetrable, the presence of a significant vol-ume fraction of macromolecules in the medium is a source of constraints on the
Address for correspondence: Dr. Raphael Lee, Department of Surgery, MC 6035, University of Chicago, Chicago 60637, IL. Voice: 773-702-6302; fax: 773-702-1634.
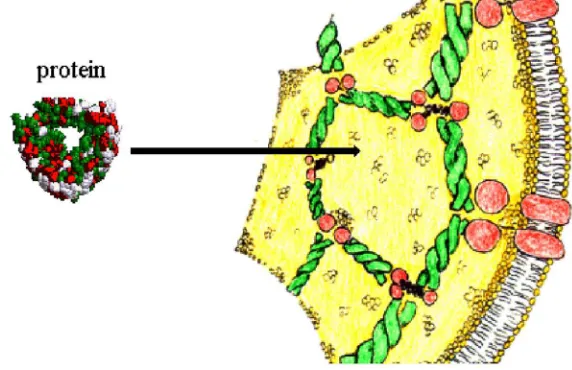
placement of an additional macromolecule. These constraints depend upon the rela-tive sizes, shapes, and concentrations of all macromolecules in that environment. Volume may be excluded also by the surfaces of “immobile” structures, that is, mem-branes and large macromolecular assemblies (FIG. 2). Excluded volume effect, as de-scribed by Minton and others,2–6 can predict many of the aspects of molecular crowding in vivo, but other physical factors need to be considered. For instance, a
FIGURE 1. Cell compartments are crowded. Actin filaments, ribosomes, membrane structures and other macromolecular assemblies occupy a volume fraction which can be as large as ϕ = 0.4.
FIGURE 2. The volume of certain cellular compartments, though comparable with
significant outcome of molecular crowding is exerted via diffusion effects on the process of aggregation of unfolded proteins.7 In addition, interfacial water mole-cules within a few hydration layers are also a sensor of the cell crowding.8,9 The physical properties of confined water differ considerably from those corresponding to bulk water and affect protein–protein interactions.8
In this chapter we will review features of the molecular crowding effect that are relevant for describing the microscopic mechanism of thermal injuries. For details regarding various mathematical formulations of the crowding effects, the reader is directed to the original papers2–7 in the reference list.
NONSPECIFIC INTERACTION AND ENTROPIC EFFECTS ON PROTEIN STABILITY
Proteins are made from an assortment of 20 very different amino acids, each with a distinct chemical personality. This leads to specific interactions among the amino acids (FIG. 3), which are important for the primary and secondary structure, as well
as nonspecific interactions. A nonspecific interaction does not depend strongly upon details of the primary, secondary, or tertiary structure(s) of the interacting molecules, but rather upon global properties of the molecules, such as polarity and macromo-lecular shape, or/and properties of the surrounding environment. Hydrophobic inter-actions between molecules are promoted by structuring effects of water. Molecular crowding is a source of nonspecific interactions.
Because of the complexity of these interactions between the protein of interest and crowding agents (all the other cellular components), it is difficult to predict the net energetic effect of macromolecular crowding on the protein dynamics.6 Small molecules (water, amino acids, etc.) alter protein dynamics by short-range site–site excluded interactions typically over distances of a few angstroms. In contrast, the range of macromolecular excluded volume interactions can be on the order of tens of angstroms, which is given by the actual size of globular proteins. By modeling the crowding particles as hard spheres, Cheung, Klimov, and Thirumalai6 predicted the changes in the folding of two-state folders (proteins that can be characterized by two states, folded or unfolded, and have no other intermediate states) by using entropic arguments. They assume that proteins would prefer to be localized in a region that is free of the macromolecular objects. The probability to find such a region decreases at a high fractional volume occupancy f > 0. Therefore, at high values of f, there is
an increased probability that a protein is in its compact form (folded in its native state). If the protein is compact at large f values, then the entropy change
∆S = S (f > 0) − S (0) < 0
because conformations involving unfolded states are suppressed. Thus, the stability of the native state of a protein is predicted to increase as fincreases.
MOLECULAR CROWDING INCREASES THERMAL STABILITY OF CELLULAR PROTEINS
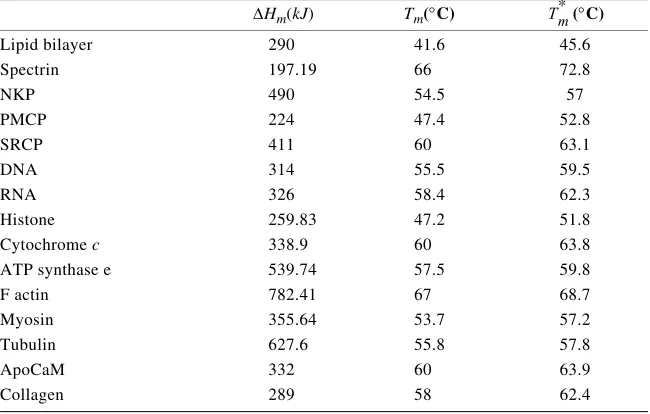
Proteins and protein assemblies optimally perform their function when they are in specific three-dimensional conformations. High temperatures alter these confor-mations and often lead to irreversible processes (denaturation) which affect the cell viability and trigger cell death. Because the functional structure of each protein and organelle is unique, so is its vulnerability to denaturation at high temperatures. Char-acteristic vulnerability to thermal denaturation of each cellular component can effi-ciently be characterized by two main thermodynamic parameters, the melting temperature (Tm) and denaturation enthalpy (∆Hm).10Tm represents the temperature at which half of the proteins are denaturated and is the enthalpy of unfolding at this temperature. Tm and ∆Hm are obtained routinely by calorimetric measurements of proteins in dilute solutions.11 In the crowded environment of a living cell, the work required for a protein to unfold is much greater than that required for unfolding in a dilute solution. Crowding increases usually the value of the melting temperature of a protein.12,13 Evidence for the increase of Tm due to crowding can be obtained sim-ply by calorimetric measurements of proteins incubated with surfactants. For exam-ple, the melting temperature of actin increases by approximately 5°C in the presence
the melting temperature of the protein in solution to account for volume exclusion effects,12
where R is the gas constant, R⳩ 8.315 JK−1 mol−1. K represents the equilibrium
re-action constant defined as the ratio between the unfolding (ku) and folding (kf) rates of the protein. The above equation states that any isothermal variation of K changes
the temperature at which half of the proteins are denaturated. TABLE 1 displays the values of the thermodynamic parameters Tm and DHm, as well as the expected values , =Tm + ∆Tm, for the melting temperatures of these biomolecules in situ. We
can see that crowding effects can substantially increase the thermal stability of the cellular components. However, the amount of unfolded protein increases dramatical-ly at supraphysiological temperatures. We have shown that,7 at temperatures above 60°C,tissue proteins are most likely denatured, with probabilities approaching unity. We can infer from TABLE 1 that the lipid bilayer and membrane-bound ATPases are the proteins most predisposed to thermal denaturation. Therefore, the alteration of the plasma membrane is likely to be the most significant cause of the tissue ne-crosis. This hypothesis correlates with the observation that edema is considered to be the first evidence of thermal injury in tissue. This edema is likely due to early dis-turbances in the cell membrane or cell membrane ion pumps (NKP). In many cases it appears that these cells can recover from this injury (i.e., most first-degree burns heal). Temperatures above the first-degree burn threshold lead to irreversible
dam-. (1)
TABLE 1. The values of the thermodynamic parameters Tmand ∆Hm as determined in calorimetric experiments and the expected values corrected for crowding effects
ATP synthase e 539.74 57.5 59.8
age to the cell membrane or other macromolecules and yield a critical injury. F actin
seems to be a very stable protein at elevated temperatures, as one can deduce from TABLE 1. This protein has a low probability of unfolding in the temperature range corresponding to a second degree burn and is damaged extensively only at higher temperatures, that is, in a third-degree burn.7 Cells contain also other very thermally stable proteins, as for example, heat-shock proteins (Hsps). Hsps are assumed to act
as molecular chaperones to assist in refolding denatured proteins. Hsp25 and Hsp27
have a midpoint transition temperature of Tm = 69.9°C,15 which is higher than that
corresponding to F actin, for example. Crowding effects inherently enhance the
sta-bility of these proteins, too. One can predict that Hsps can exist in functional form
of 80% even above 75°C. However, above 45°C, the cell membrane breakdown is so extensive that it is improbable that Hsps occur in high enough concentration to be an
effective protector against cell disruption.7
MOLECULAR CROWDING PLACES GEOMETRICAL RESTRICTIONS ON THE UNFOLDING OF PROTEINS
Let rg be a measure of the compactness of the protein structure, i.e., the radius of
gyration of a protein (FIG. 4). This rg expands during unfolding excluding volume to other surrounding proteins. If the increase ∆r of the radius of gyration is in the range
of the protein interspace, defined as the mean distance between proteins in solution [ , where n is the protein concentration], the subsequent confinement
would provide stability for adjacent proteins which are in compact, native states. The self-stabilization effect develops progressively during protein unfolding. The results are also relevant for describing the stability of proteins in tightly packed fibers and membranes.16
To understand this effect in a more quantitative way, we can write the apparent equilibrium constant K of the protein in a crowded environment as7
where K0 stands for the equilibrium constant of proteins in the ideal case of a dilute
solution, rN is the radial size of the molecule in the native form and f represents the
fractional volume occupancy of the protein,
is the total number of proteins in the particular environment. It is not difficult to ob-serve that any conformational change that increases the volume of a protein
changes the protein interspace d. Consequently, d is related to the evolution of the
density distribution of the population in the unfolded state of the protein PU,
FIGURE 4. The compactness of the protein structure in the folded state is different
from that in the unfolded state. rg expands during unfolding, excluding volume to other sur-rounding proteins.
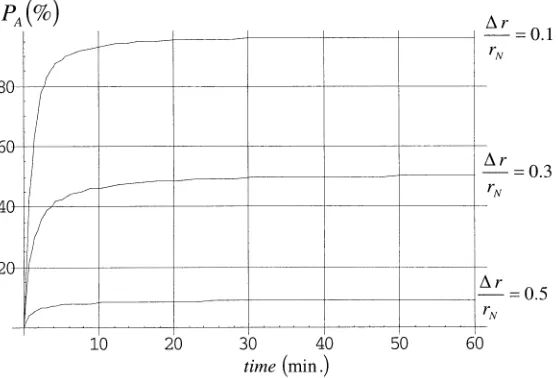
FIGURE 5. Denaturation of adenylate kinase in time for a temperature history
PU=1−PN, where PN is the density distribution of the population in the native state. The above observation leads to the conclusion that the protein unfolding pro-cess is progressively inhibited by the conformational changes of proteins that in-crease their coresponding volumes and shrink the characteristic interspace (d)
between proteins.7 In FIGURE 5 one can observe that, for example, the probability of distribution of adenylate kinase in the denaturated state (PA) is much lower in a
so-lution in which proteins have a finite volume occupancy (i.e., f= 20%) than in an
ideal case of a dilute solution (f → 0). The self-stabilization occurs because of steric
effects (∆r increases) induced by the unfolding of a fraction of proteins in solution.
As the volume available per unfolded protein is larger than that corresponding to a native protein, this will impose geometrical constraints (volume exclusion) on the proteins in native states, as described above.
A more general approach will describe the unfolding of a protein species i in a
crowd formed by M various other protein species. This shows that the volume
exclu-sion effect leads to a decrease of the equilibrium constant Ki of the protein species i in a mixture of different protein kinds J.7
UNFOLDING OF THE MOST THERMOLABILE PROTEINS IN A CELL INCREASES THE STABILITY OF THE
OTHER CELLULAR PROTEINS
The vulnerability to denaturation at high temperatures of various cellular proteins is different.10 Thus, by increasing the temperature over the physiological level, pro-teins with low midpoint transitions will unfold first. The excluded volume theory tells us that the unfolding of these proteins can provide extra stability for the other proteins in the cell, having presumably higher melting temperatures. Obviously, the stabilization of proteins with the highest melting point transitions (e.g., proteins making up the cytoskeleton, such as actin and myosin) is a result of the excluded
vol-ume yielded by the unfolding of all the other protein species. The stability of the most thermolabile proteins in the cell are also affected by molecular crowding, that is, these proteins have a higher stability in a cell than in a dilute solution. However, it is unlikely that their dynamics can be influenced at any extent by the unfolding of the proteins with high melting temperatures.7
DIFFUSION OF THE PROTEINS MODIFIES THE NET EFFECTS OF MOLECULAR CROWDING
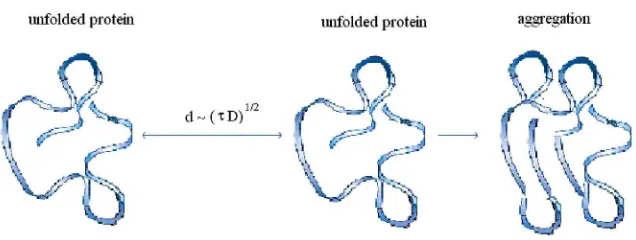
in-creases with the increase of the population in the unfolded state. However, the rate of aggregation is limited by the diffusion of the unfolded proteins (FIG. 6). Inherent-ly, crowding in homogeneous solutions of unfolded proteins leads to a rapid irrevers-ible aggregation of those proteins.
The rate of aggregation can be approximated by the inverse of the diffusion time τi, ka,i = 1/τi. τi for an unfolded molecule of a protein species i relates to its corre-sponding diffusion coefficient Diby τi=d′2/4Di. Here, Di is an effective diffusion
coefficient which includes a correction due to the restriction on the movement of the molecules in a crowded environment.7
As discussed above, the aggregation applies only to unfolded proteins. Under such circumstances, the translational diffusion length d depends on density
distribu-tion of the populadistribu-tion in the unfolded state of the protein (PU), and on the fractional volume occupancy of the protein (f).7 In this way, the rate of protein aggregation is
directly related to the extent of crowding.
QUANTITATIVE ESTIMATION OF CROWDING EFFECTS ON THERMAL DENATURATION OF PROTEINS
As shown above, crowding can substantially affect the transition of a protein be-tween its native (N) and unfolded (U) states via volume exclusion effects. Also,
crowding influences considerably the aggregation (A) of unfolded proteins. To
ex-amine the details, one can study the protein transition
as described by Despa, Orgill and Lee.7 ku,i and kf,i are the unfolding/folding rate co-efficients and ka,i denotes the rate constant for the irreversible aggregation. ku,i is a
function of the melting temperature Tm,i and the enthalpy of denaturation ∆Hun,i
. (3)
FIGURE 6. If the distance between two unfolded proteins is comparable with the diffusion length d, then the probability of irreversible aggregation of unfolded proteins increase.
N
kf i, ku i,
U
ka i,
A is a constant which determines the time scale of the unfolding process. This
de-pends, among others, on the coupling of the protein with the solvent.17–20 The back-ward rate is simply kf,i = ku,i/Κi. Tm,i and ∆Hun,i are derived by calorimetric
measurements of dilute protein solutions. Corrections for crowding effects are incor-porated in Ki (see Eq. 2) via volume exclusion and in ka,i by rescaling the
translation-al diffusion length d (see above). This is a model for an experiment in which
temperature is changed with time according to a “temperature history” T(t).7 The
ap-proach yields PN,i, PU,i and PA,i, representing the distribution density of the popula-tion in the native, unfolded and aggregated state of the protein species i. A suggestive
result is presented in FIGURE 7. Here, one can observe the effects of crowding in a mixture of proteins (adenylate kinase, creatine kinase, ATP synthase e and cyto-chrome c) with different thermal stabilities. Steric effects brought by the unfolding
of thermolabile proteins enhance the stability of those proteins in the mixture which have higher melting points. The top curve represents the denturation of cytochrome
c in a homogeneous solution, while the bottom curve describes the course of
dena-turation of this protein in a mixture with the other three proteins. A low probability for aggregation means, implicitly, an increased stability in the native state.
. (4)
ku A ∆Hun
RT --- 1 T
Tm ---–
⎝ ⎠
⎛ ⎞
–
⎝ ⎠
⎜ ⎟
⎛ ⎞
exp =
FIGURE 7. Steric effects brought by the unfolding of thermolabile proteins enhance the stability of those proteins in the mixture that have higher melting points. The time–tem-perature course is the same as the one used in FIGURE 4. The propensity of unfolded proteins
MOLECULAR CROWDING AFFECTS THE BEHAVIOR OF PROTEINS VIA A WATER EFFECT
For a mean protein mass in the cell of ⬃50 kDa, at an assumed protein concen-tration of 300 mg/mL, the fraction of interfacial water would be about 30% (two lay-ers of interfacial water) to 70% (four laylay-ers) of the total water in the cell. Under circumstances in which macromolecular crowding can change the properties of a significant fraction of the water in the cell, crowding effects could exert a strong in-fluence on the behavior of smaller solutes as well as on larger macromolecules.
Recent in vivo expriments21 showed that the interfacial water component
increas-es from 23.5% of total water in the control sample to 25% in yeast heat-shocked at 315K and to 30% in yeast cells at 323K. The heat shock, which causes some proteins
to become unfolded in the cell and, therefore, increases molecular crowding, will change also the hydration of the cell components. An alteration of the properties of the interfacial water could have a direct influence on a range of cellular functions and properties.
Physical properties of water confined in microscopic environments differ from the properties corresponding to bulk water.8 For example, it was shown recently8 that water molecules under hydrophobic confinement move about an order of mag-nitude slower than those in the bulk, and that the dielectric constant of this water lay-er is significantly reduced. Watlay-er’s high dielectric constant is the reason why it is a good solvent for ions: it screens their electrical charges and so prevents them from aggregating. But in the vicinity of hydrophobic residues in a protein chain, the re-duction in dielectric constant means that charged residues will interact much more strongly, potentially helping to fix the protein’s folds in place.
Another example in which the behavior of biomolecules is altered via a water ef-fect is the interaction between hydrophobes. The hydrophobic interaction—the ap-parent attraction between hydrophobic species in water—is considered a key factor in maintaining the correct folded conformation of a protein molecule and also the main cause of protein aggregation. This attraction is thought to result, in a way that is still imperfectly understood, from changes in the arrangement of hydrogen bonds between water molecules surrounding a hydrophobe.8,22 This gives rise to a local polarization of the interfacial water, which is shown to be strong enough to induce long-range attraction between hydrophobic molecules.
SUMMARY
Proteins are three-dimensional structures with conformations dictated by the characteristic amino acid sequences. At body temperature, proteins are in native con-formations that allow them to perform their designated functions. At supraphysio-logical temperatures, proteins are driven towards unfolded conformations. Steric effects increase the stability of the proteins which are in compact, native states.As each type of protein has its own thermal stability, the unfolding of the most thermo-labile proteins will increase the stability of the other proteins. In unfolded
the unfolded state and the rate of aggregation is limited by the diffusion of the un-folded proteins. The net outcome of the steric effects on individual proteins can be modified by diffusional motion of the molecules.
Understanding the regulation of these processes may lead to clinical strategies for limiting the devastating development of the injury after the thermal insult of the tis-sue stopped. Within the computational complexity theory, protein dynamics is de-fined rigorously as NP harda and, so far, we can simulate exactly the in vitro
structural dynamics only for a very limited pool of proteins. However, the volume of data sets is often so big that the efficiency of manipulation and extraction of useful information becomes problematic. Bringing the inherent solvent effects as well as crowding into play increases the complexity of the problem.
Despite difficulties, studying proteins can advance by a profitable utilization of recent statistical mechanical treatments of potential energy surfaces17–20 conjoined with experimental observations on the energetics and stability of proteins.11 Here, we have reviewed several quantitative analyses of the kinetic stability of cellular components confronted with the destabilizing effect of irreversible alteration which are relevant for describing the microscopic mechanism of thermal injuries.
REFERENCES
1. ALBERTS, B., D. BRAY, J. LEWIS, et al. 1983. In Molecular Biology of the Cell.: 111.
Garland. New York & London.
2. ELLIS, J.R. 2001. Macromolecular crowding: an important but neglected aspect of the
intracellular environment. Curr. Opin. Struct. Biol. 11: 114–119.
3. BURG, M.C. 2000. Macromolecular crowding as a cell volume sensor. Cell. Physiol.
Biochem. 10: 251–256.
4. VERKMAN, A.S. 2002. Solute and macromolecule diffusion in cellular aqueous
com-partments. Trends Biochem. Sci. 27: 27–33.
5. HALL, D.& A.P. MINTON. 2003. Macromolecular crowding: qualitative and
semiquanti-tative successes, quantisemiquanti-tative challenges. Biochim. Biophys. Acta 1649: 127–139.
6. CHEUNG, M.S., D. KLIMOV & D. THIRUMALAI. 2005. Molecular crowding enhances
native state stability and refolding rates of globular proteins, Proc. Natl. Acad. Sci. USA 102: 4753–4758.
7. DESPA, F., D.P. ORGILL & R.C. LEE, 2005. Effects of crowding on the thermal stability
of heterogeneous protein solutions. Ann. Biomed. Eng. 33: 1125-1131 .
8. DESPA, F., A. FERNANDEZ & R.S. BERRY. 2004. Dielectric modulation of biological
water. Phys. Rev. Lett. 93: 228104
9. FORD, R.C., S.V. RUFFLE, A.J. RAMIREZ-CUESTA, et al. 2004. Inelastic incoherent
neu-tron scattering measurements of intact cells and tissues and detection of interfacial water. J. Am. Chem. Soc. 126: 4682–4688.
10. DESPA, F., D.P. ORGILL, J. NEWALDER, & R.C. LEE. 2005. The relative thermal stability
of tissue macromolecules and cellular structure in burn injury. Burns 31: 568–577. 11. MAKHATADZE, G.I. & P.L. PRIVALOV. 1995. Energetics of protein structure. Adv.
Pro-tein Chem. 47: 307–425.
12. MINTON, A.P. 2000. Effect of a concentrated “inert” macromolecular cosolute on the
stability of a globular protein with respect to denaturation by heat and by chaotropes: a statistical-thermodynamic model. Biophys. J. 78: 101–109.
aThis is when the number of spatial configurations available for a system increases
13. ZHANG, S. & C. WANG. 2001. Effects of macromolecular crowding on the refolding of
glucose-6phosphate dehydrogenase and protein disulfide isomerase. J. Biol. Chem.
37: 34396–34401.
14. TELLAM, R.L., M.J. SCULLEY, L.V. NICHOL & P.R. WILLS. 1983. Influence of
poly(eth-ylene glycol) 6000 on the properties of skeletal-muscle actin. Biochem. J. 213: 651–
659.
15. DUDICH, I.V,, V.P. ZAV’YALOV, W. PFEIL, et al. 1995. Dimer structure as a minimum
cooperative subunit of small heat-shock proteins. Biochim. Biophys. Acta. 11253:
163–168.
16. YANNAS, I.V., J.F. BURKE, P.L. GORDON, et al. 1980. Design of an artificial skin. II.
Con-trol of chemical composition. J. Biomed. Mater. Res. 12: 7–32.
17. DESPA, F. & R.S. BERRY. 2001. Inter-basin dynamics on multidimensional potential
surfaces: escape rates on complex basin surfaces. J. Chem. Phys. 115: 8274–9278.
18. DESPA, F. & R.S. BERRY. 2003. Inter-basin dynamics on multidimensional potential
surfaces: kinetic traps, Eur. Phys. 24: 203–206.
19. DESPA, F., A. FERNÀNDEZ, R.S. BERRY, et al. 2003. Inter-basin motion approach to
dynamics of conformationally constrained peptides. J. Chem. Phys. 118: 5673–5682.
20. DESPA, F., D.J. WALES & BERRY RS. 2005. Archetypal energy landscapes: dynamical
diagnosis. Escape rates on complex basin surfaces. J. Chem. Phys. 122: 24–103.
21. R.C. FORD, S.V. RUFFLE, I. MICHALARIAS, et al. 2004. Neutron scattering
measure-ments of intact cells show changes after heat shock consistent with an increase in molecular crowding. J. Mol. Recogn. 17: 505–511.