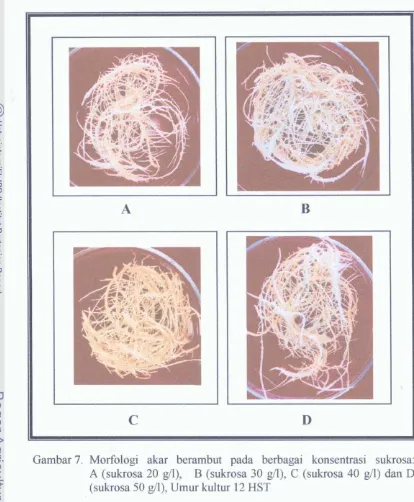
Kultur akar transgenik dari Trichosanthes cucumerina L.: beberapa faktor yang berpengaruh terhadap produksi biornassa dan hasil protein total, serta aktivitas anticendawan dari protein asal akar transgenik
Bebas
218
0
0
Teks penuh
Gambar
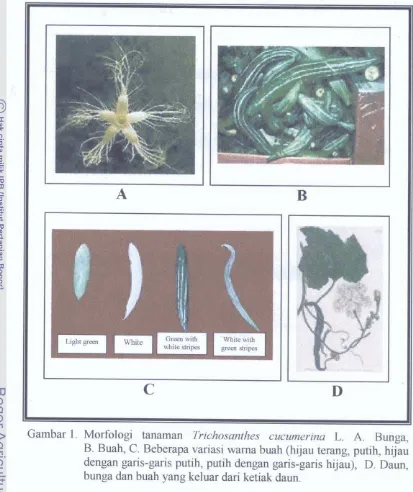

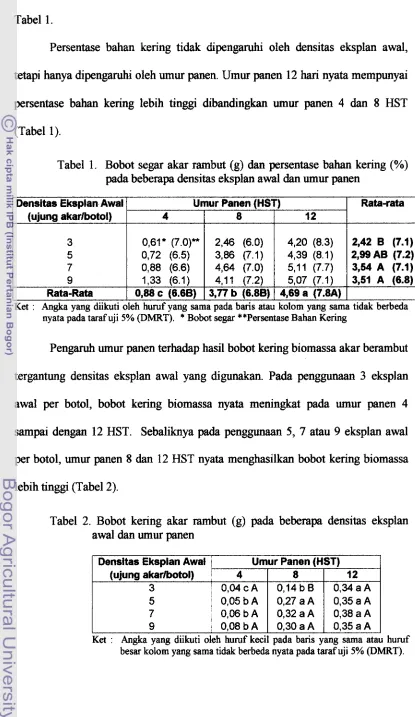
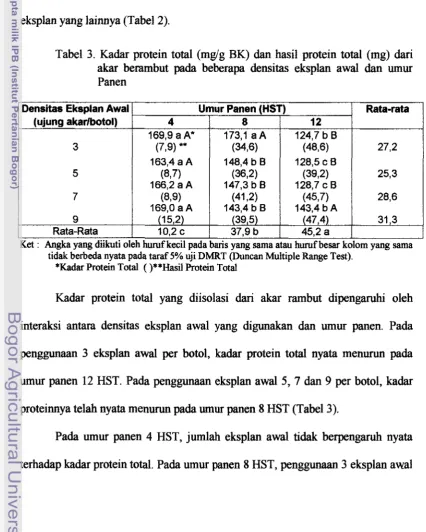
+7
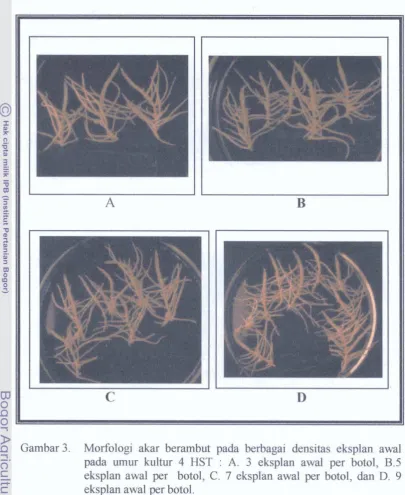
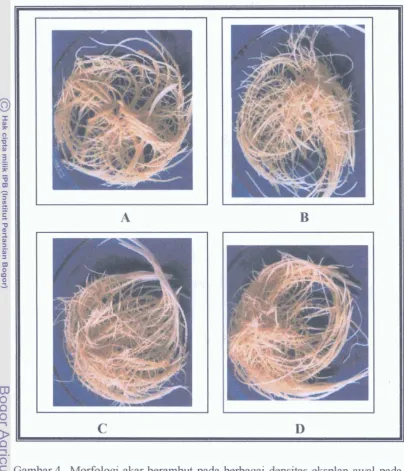
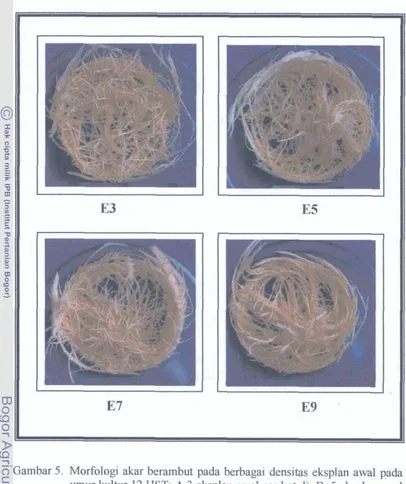
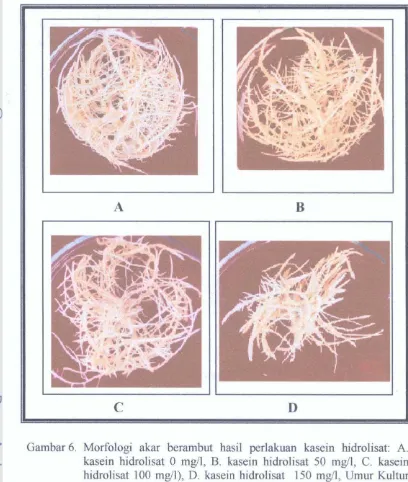
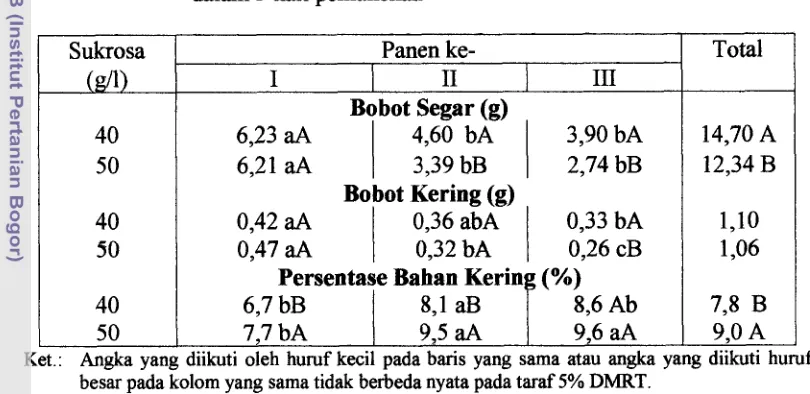
Dokumen terkait
