PENINGKATAN MUTU POD KAKAO MELALUI AMONlASl DENGAN
UREA DAN BIOFERMENTASI DENGAN
Phanerochaefe
chrysosporium
SERTA PENJABARANNYA KE DALAM
FORMULAS1 RANSUM RUMINANSIA
Oleh
ERIKA BUDlARTl LACONI
PROGRAM PASCA SARJANA
INSTITUT PERTANIAN BOGOR
Erika Budiarti Laconi. Peningkatan Mutu Pod Kakao melalui Amoniasi dengan Urea dan Biofermentasi dengan Phanerochaete chrysosporiurn serta Penjabarannya ke dalam Formulasi Ransum Ruminansia. Toha Sutardi (ketua), Djokowoerjo Sastradipradja, Lily Amalia Sofyan, Triadi Basuki dan Djoko Said Damardjati (Anggota).
Semakin berkurangnya lahan pertanian serta rendahnya mutu hijauan
dan rerumputan menjadi pendorong untuk pengadaan pakan alternatif, antara lain pemanfaatan limbah tanarnan perkebunan seperti kakao melalui aplikasi
teknologi untuk meningkatkan manfaatnya. Keunggulan limbah ini
produksinya berlimpah dan terkonsentrasi dalam wilayah tertentu sehingga
dapat dijadikan titik tolak pertumbuhan agroindustri pakan. Tahun 1997
produksi kakao mencapai 332.929 ton dan sebesar 73% adalah pod kakao
yang belum termanfaatkan secara optimal. Masih banyak kendala untuk menggunakan pod kakao sebagai pakan hijauan ternak ruminansia,
diantaranya konsentrasi amonia rendah (0.74 mM), adanya alkaloid
theobromine dan tingginya kandungan lignin sehingga kecernaan nutrien
menurun. Untuk itu penggunaannya sebagai pakan hijauan rnemerlukan
sentuhan teknologi, seperti arnoniasi dan bioferrnentasi. Selain itu upaya menekan biaya pakan, limbah kakao lainnya seperti kulit biji kakao dan bubuk
kakao bersarna-sama dengan bungkil kelapa sawit juga berpotensi sebagai sumberdaya pakan, hanya saja belum termanfaatkan secara optimal. Untuk itu penelitian ini diharapkan dapat menentukan (a). teknologi pengolahan yang tepat, dan efisien untuk mendayagunakan limbah tanaman kakao. (b). mendapatkan formula ransum komptit dengan bahan baku berasal dari
limbah kakao dan kelapa sawit untuk ternak sapi periode pertumbuhan dan (c). mendapatkan persamaan regresi untuk menduga peubah-peubah tolok
Penetitian dilakukan di Laboratoriurn Makanan Ternak dan
Laboratorium Lapang, Fakultas Peternakan IPB Darmaga, berlangsung selama 10 bulan, yang terdiri dari dua tahap penelitian. Penelitian tahap 1 terdiri dari empat perlakuan pengolahan pod kakao yaitu amoniasi 1.5% urea;
ferrnentasi molases 3.0% ; biofermentasi isi rumen + molases 3.0% dart
biofermentasi dengan kapang Phanerochaete chrysosporium Burdsall ATCC
34541
+
molases3,0%
dan pod kakao tanpa pengolahan sebagai kontrol.Evaluasi dilakukan berdasarkan perubahan kornposisi nutrien dan serat serta
pengamatan perubahan struktur fisik serat biomass pod kakao. Berdasarkan
hasil penelitian tahap 1, dilanjutkan percobaan in vivo pada ternak. Ransum
berbahan dasar kulit biji kakao, bubuk kakao serta bungkil kelapa sawit sebagai konsentrat dan pod kakao sebagai hijauan, juga disuplementasi
dengan CaC03, premix sapi, urea dan garam. Lima macam formula ransum
perlakuan disusun isoprotein dan jumlah TDN yang sama dengan kadar
protein kasar 16% dan TDN 69%, masing-masing terdiri dari R1
=
Konsentrat+ pod kakao tanpa pengolahan; R2
=
Konsentrat + pod kakao amoniasi; R3 =Konsentrat + silase pod kakao; R4
=
Konsentrat + biofermentasi pod kakao(isi rumen) dan R5 = Konsentrat + biofermentasi pod kakao (kapang).
Ransum diujicobakan pada 5 ekor sapi FH jantan berbobot 145+3,6 kg,
dengan rancangan Bujur Sangkar Latin (5x5). Pemberian ransum 2 kali sehari dalarn bentuk pellet, sedangkan air minum disediakan sepanjang hari.
Peubah yang diarnati adalah: (1) konsurnsi dan kecernaan nutrien, (2)
parameter metabolisme rurnen dan afantoin urin. (3) konsentrasi VFA total
dan individual cairan rumen, (4) pertambahan bobot tubuh harian, (5)
Pada proses biofermentasi pod kakao dengan kapang
P.chrysosporiurn, kandungan lignin berkurang sebesar 7.12% dan 9.40% bahan kering yang hilang selama proses berlangsung. Nilai kecernaan bahan kering (KCBK) = 51.70% dan bahan organik (KCBO) = 52.65%. Teknologi arnoniasi pod kakao dengan 1.5% urea, kandungan lignin berkurang sebesar
5.57% dan bahan kering yang hilang hanya 2.64%, sedangkan nilai KCBK =
57.83% dan KCBO
=
55.73%. Perlakuan pengolahan terhadap pod kakaomeningkatkan kecernaannya. Berdasarkan hasil percobaan tahap 1 ini,
biomass pod kakao yang dihasilkan dipergunakan sebagai sumber hijauan
utama dalam ransum sapi pertumbuhan. Hasil penelitian menunjukkan bahwa ransum berbahan baku limbah kakao dan bungkil kelapa sawit cukup
palatabet. Konsumsi bahan kering berkisar 104
-
117 glkg B B ~ , ' ~ / ~ atau 3.0-
3.5% dari bobot badan sapi. Selain itu juga mampu rnenyediakan kondisi lingkungan rumen yang dapat rnenunjang pertumbuhan dan kerja mikroba rurnen, pH rumen berkisar 6.06
-
6.38; konsentrasi amonia 4.18-
6.30 mMdan konsentrasi VFA total adalah 85.50
-
120.62 mM. Terdapatkorelasipositif antara sintesis protein mikroba dengan alantoin yang diekskresikan di dalam urin. Data ekskresi alantoin mencerminkan bahwa
perlakuan ransum R5 mernberikan sumbangan protein asal rnikroba untuk
ternak tertinggi.
Kecernaan nutrien meningkat setelah perlakuan pengolahan terhadap pod kakao. Kecernaan serat ransum R5 (biofermentasi P. chrysosporiurn)
lebih tinggi (Pc0.05), terutama kecernaan NDF (54.20%); ADF (36.14%) dan
energi (56.92%). Kecernaan protein ransum R2 (amoniasi) dan ransum R5 (biofermentasi P.chrysosporium) tidak berbeda nyata yaitu R2 = 51.88% dan R5 = 55.16%, tetapi keduanya lebih tinggi (Pc0.01) dibandingkan dengan
ketiga perlakuan ransum lainnya. Perbaikan kecernaan protein
mengindikasikan terjadinya peningkatan efisiensi penggunaan nitrogen
(PBBH) rnemperkuat indikasi ini. Jumlah nitrogen teretensi untuk ransurn R2
adalah 1.45 glkg BB 0.75/h dan ransurn R5 adalah 1.60 glkg BE3 0.75/h.
Lebih rendahnya jurnlah protein rnikroba yang disintesa dan berkurangnya jurnlah ekskresi alantoin di dalarn urin pada ransum R2,
menunjukkan bahwa ransum R2 bersifat 'by pass' rumen, sehingga lebih banyak sebagai protein murni. Nilai persentase penggunaan nitrogen netto
(NPU) berturut-turut untuk kelima ransum adalah: R1
=
40.58%; R2= 50.14%;R3= 41.38%; R4= 42.49% dan R5= 53.03%.
Kadar glukosa darah sapi percobaan masih dalam kisaran normal, yaitu 68.00
-
78.40 rngIlOOrnl, sehingga fungsi fisiologis organ-organ tubuhtidak terganggu. Teknologi amoniasi dengan 1.5% urea dan bioferrnentasi
dengan kapang P.chrysosporiurn meningkatkan konsentrasi asarn propionat
dan rnenurunkan angka non glugogenik rasio (NGR). Berdasarkan effisiensi penggunaan energi ransurn, fermentasi rurnen yang rnengarah pada
pembentukan asam propionat lebih mengguntungkan, energi yang terbuang
sebaga~ rnetan (CH4) berkurang. Angka NGR berkisar 2.86
-
4.44, rnasihdalam kategori terbaik untuk efisiensi penggunaan energi bagi perturnbuhan
dan penggernukan ternak. Angka NGR terkecil adalah 2.86 untuk ransurn
R5. Ransurn R5 rnenghasilkan produksi panas (PP) yang lebih tinggi
dibandingkan R2 yaitu 0.78 MJIkg BB 0.75/h (R5) dan 0.57 MJIkg BB 0.75/h
(R2). Energi teretensi (RE) untuk ransurn R2 dan R5 lebih baik dari ketiga
ransurn lainnya yaitu R2 = 0.18 M J / ~ ~ B B " ~ ~ / ~ dan R5
=
0.17 M J I ~ ~ B B ~ " ~ ~ ~ , akan tetapi tidak berbeda antar keduanya.Perbedaan jurnlah nitrogen dan energi teretensi berdampak positif
terhadap pertambahan bobot badan harian ternak. Sejalan dengan nilai RN dan RE, angka pertambahan bobot tubuh harian pada ternak yang
rnendapatkan ransum R2 (amoniasi) dan ransum R5 (bioferrnentasi
dan R5
=
1.46 kglh, demikian juga dengan pola deposisi protein dan deposisi lemak.Berdasarkan profil peubah-peubah yang diukur, perlakuan ransurn R2
(arnoniasi) dan ransum R5 (biofermentasi P. chrysosporiurn) mernberikan hasil yang lebih baik, ditinjau dari pasokan nutrien dari mikroba rumen, pola fermentasi rumen, neraca nitrogen dan energi serta pertambahan bobot badan harian ternak. Teknologi arnoniasi dengan 1.5% urea terhadap pod kakao lebih efektif dan efesien untuk diaplikasikan.
Persamaan regresi untuk menduga besarnya perubahan protein dan
energi dengan menggunakan paararneter rnetabolik, pertarnbahan bobot
badan harian (PBBH=kg/h); bobot badan metabolik ( B B M = ~ ~ ' . ' ~ ) ;
konsentrasi amonia (NH3=mM); jumlah nitrogen tercerna (NTC=glh) dan
konsentrasi alantoin dalarn urin (alan=g/h) adalah sebagai berikut :
K-PK
=
-1.844+
0.091 PBBH + 0.058 BBM-
0.012 NH3(kglh) + 0.002 NTC + 0.002 Alan,
dengan nilai RZ
=
0.77K-ME
=
-1 6.122-
0.747 PBBH+
0.487 BBM-
0.1 75 NH3(MJlh)
+
0.324 NTC+
2.586 Alan,dengan nilai R2
=
0.61Persamaan regresi untuk mengevaluasi mutu ransum berdasarkan parameter mutu protein ransum seperti retensi nitrogen (RN=g/h); sintesis protein
mikroba (SPM=g/h); penggunaan nitrogen netto (NPU=%) dan pertarnbahan bobot tubuh harian (PBBH=kg/h) untuk ternak sapi turnbuh adalah sebagai
SPM
=
1073.10-
73.69 PBBH-
26.69 BBM+
3.96 NH3(glh)
+ 5.1 3 NTC+
50.08Alan,dengan nilai R2
= 0.54
NPU
=
141.18-
5.03 PBBH-
3.06BBM
+
1.35 NH3("/.I
-
0.68 NTC-
1.01 Alan, dengan nilai R* = 0.85PBBH
=
-
1.447 + 0.014 BBM+
0.141 NH3+
0.016 NTC(kglh)
+ 0.085 Alan,dengan nilai R2
= 0.61
RN = 3.75 + 1.09 PBBH
-
0.04B B M
+ 0.01 N H 3(glh)
+ 0.94 NTC-
0.54 Alan,Improving of Cocoa Pods Quality Through Urea-Ammonia Treated and Bio-fermented Using Phanerochaete chrysosporium as well as
Its fnclusion i n Ration Formulation for Ruminants
ABSTRACT
Two experiments were carried out to evaluate the utilization of wcoa pods as a feedstuff for ruminants. In the first trial, the effect of urea-ammonia and bio- fermented treated on the quality of wcoa pods was conducted. Treatments were: (i) 1.5 Oh of urea. (ii) 3 O h of mollases, (iii) rumen content plus 3 % of mollases, (iv)
bio-fermented using P.chrysosporium and (v) untreated cocoa pods as a control.
Based on the results found in the previous trial, the feeding study was done using five FH steers in a 5 x 5 Latin-Square design. The ration treatments consisted of offering basal diet added of either, (R,) untreated cocoa pods, (R,) urea-
ammonia treated of cocoa pods, (R,) silage of coma pods, (R,) bio-fermented
of cocoa pods using rumen content, or (R,) with bio-fermented of cocoa pods
using P.chrysosporium. All treatment diets were iso-caloric and iso-nitrogeneous with 16 O h and 69% of crude protein content and total digestible nutrient
respectively. The lignin and dry matter contents were lower respectively 7.12 and 9.4 Oh for cocoa pods treated with bio-fermented using P.chrysosporiurn (&); and
5.57 and 2.64 Ohfor those on urea-ammonia treated (R,). Nutrient digestibility
was greater (P e 0.05) for R, and R, than the others. There was no significantly different on crude protein digestibility between R, and
&,
but the others ( P < 0.01). The improvement of crude protein digestibility implied that the nitrogen utilization of the diet was highly effective. The pattern of nitrogen and energy retention was similar for steer offered R, and R, diets. This was reflected in a greater increase in steer growth rate. Based on data for individual steers found, severaf equation were developed to predict crude protein requirement (equation 1). energy requirement (equation 2), nitrogen retention (equation 3), microbialprotein synthesis (equation 4) and average daily gain (equation 5).
(1) CP =
-
1.844 + 0.091 ADG + 0.058 BW 0.75-
0.012 NH3 +0.002DN+0.002 Alan;
?
= 0.77(2) E =
-
16.122 - 0.747 ADG + 0.487 BW 0.75-
0.175 NH3+ 0.324
D N + 2.586 Alan; r2 = 0.61
(3) NR = 3.75
+ 1.09 ADG
-
0.04 BW 0.75-
0.01 NH3+ 0.94 DN
+
0.54 Alan; r2 = 0.99(4) MPS = 1073.1 - 73.69 ADG
-
26.69 BW 0.75+ 3.96 NH3
+ 5.13 DN+ 50.08 Alan; r2
=
0.54.(5) ADG = - 1.447 + 0.014 BW 07'
+ 0.141 NH3
+ 0.016 DN
+ 0.085PENINGKATAN MUTU POD KAKAO MELALUI AMONIASI DENGAN
UREA DAN BIOFERMENTASI DENGAN
Phanerochaete
chrysosporium
SERTA PENJABARANNYA
K E
DALAM
FORMULAS1 RANSUM RUMlNANSlA
Oleh
ERIKA BUDlARTl LACONI
Disertasi sebagai Salah Satu Syarat untuk
Memperoleh Gelar Doktor pada Program Pascasarjana
lnstitut Pertanian Bogor
PROGRAM PASCA SARJANA
INSTITUT PERTANIAN BOGOR
Judul Disertasi : Peningkatan Mutu Pod Kakao Melalui Amoniasi dengan Urea dan Biofermentasi dengan Phanero-
chaete chrysosporium serta Penjabarannya Ke
dalarn Forrnulasi Ransurn Ruminansia.
Nama Mahasiswa : Erika Budiarti Laconi
Nomor lnduk : 92506, llrnu Ternak (PTK)
1.
Komisi Pembimbing/Prof. Dr. Toha Sutardi. MSc. Ketua
1
-
IProf. Dr. D. ~astradipraci*
A ggota
r
AnggotaDr. Triadi Basuki
Anggota
6r.
Dioko Said Darnardiati AnggotaPenulis dilahirkan di Teluk Betung, Bandarlampung pada tanggal 16
September 1967. Putri kedua dari enam bersaudara, papa bernama
Baharuddin Laconi dan mama Rosna Laconi (Almarhumah).
Pendidikan dasar dt SD Negeri V Teluk Betung, lulus tahun 1973.
Pendidikan Menengah Pertama di SMP Negeri Teluk Betung, lulus tahun
1976. Pendidikan Menengah Atas di SMA Negeri II Tanjung Karang dan
lulus tahun 1980. Tahun yang sama (1980) penulis melanjutkan studi di
lnstitut Pertanian Bogor melalui jalur Perintis II dan rnemperoleh gelar
Sarjana Peternakan tahun 1984.
Penulis menikah dengan lr. Rubi Herman Sutopo pada tahun 1985
dan saat ini telah dikarunia sepasang putra-putri yaitu Panji Hardian ( 7 1
tahun) dan Riskina Juwita (7 tahun).
Tahun 1991 penulis menyelesaikan pendidikan Program Master (S2)
dalam bidang llmu Ternak di Program Pascasarjana lnstitut Pertanian Bogor
dan tahun 1992 melanjutkan pendidikan Program Doktor di tempat yang
sama dalam bidang Makanan Ternak.
Sebelum menjadi staf pengajar penulis pernah bekerja pada
Perusahaan Swasta PT. Anwar Sierad, di Jakarta selama 2 tahun. Sejak
tahun 1986 hingga sekarang penulis menjadi staf pengajar di Jurusan llmu
Nutrisi dan Makanan Ternak, Laboratorium Makanan Ternak Fakultas
KATA PENGANTAR
Puji syukur dipanjatkan kepada Allah SWT yang telah melimpahkan
rahmat dan hidayahNya sehingga penulis dapat rnenyelesaikan pendidikan
program doktor.
Pada kesempatan yang berbahagia ini ijinkanlah penulis
menyampaikan penghargaan dan terima kasih yang sebesar-besarnya
kepada Bapak Prof. Dr. Toha Sutardi MSc., sebagai ketua kornisi
pernbirnbing. Prof. Dr. Djokowoerjo Sastradipradja, Prof. Dr. H. Lily Arnalia
Sofyan MSc., Dr. Triadi Basuki dan Dr. Ir. Djoko Said Damardjati masing-
masing sebagai anggota kornisi pembimbing, atas segala bimbingan, arahan,
serta saran-saran yang diberikan selama penulis mengikuti pendidikan doktor
di Program Pascasarjana IPB.
Kepada Bapak Rektor lnstitut Pertanian Bogor, Direktur Program
Pascasarjana, Dekan Fakultas Peternakan lnstitut Pertanian Bogor serta
Ketua Program Studi llmu Ternak, penulis menyampaikan rasa terima kasih
yang sebesar-besarnya atas segala fasilitas dan dorongan yang diberikan
selarna penulis rnengikuti program ini.
Ucapan terima kasih disampaikan kepada Ketua Tim Manajernen
Program Doktor Ditjen Dikti, suamiku Ir Rubi Herman Sutopo, Yayasan
Supersernar serta Ir. Rachmat Pambudy MS, dan M. Hidayat, SE serta
PT Elanco Animal Health atas bantuan biaya yang diberikan sehingga penulis
dan staf atas bantuan fasilitas penelitian berupa kandang percobaan selama
penulis melakukan penelitian.
Ucapan terimakasih juga disampaikan kepada Ibu Prof. Dr. Lily Amatia
Sofyan dan staf pengajar serta pegawai yang telah memberikan bantuan
fasilitas laboratorium, bimbingan moril dan kerjasama yang sangat
menunjang keberhasilan penelitian ini. Kepada "lbu" yang telah tulus, ikhlas
dan penuh kesabaran memberikan dukungan moril kepada penulis untuk
tetap tabah dan tegar dikala masa-masa sulit, diucapkan terima kasih.
Ucapan terima kasih juga disampaikan kepada penguji luar komisi
Bapak Dr. Ir. Arifin Habibie, MS., Kepala Bagian Kehutanan, Biro Pertanian
dan Kehutanan, BAPPENAS, Jakarta dan Bapak Dr. Ir. Oskari Atmawinata,
Kepala Puslit Kopi dan Kakao, Jember yang telah memberikan saran dan
kritik yang sangat menunjang hasil penelitian ini, sehingga rnenjadi lebih
bermanfaat. Juga kepada adik-adik di Bor:el Computer, terutama Fajar
Gunardi atas bantuan tenaga dan pikiran untuk fasilitas pengetikan dan
pembuatan transparansi pada ujian sidang terbuka.
Akhirnya dengan segala kerendahan hati penulis mempersembahkan
disertasi ini kepada Almarhumah mama tercinta Rosna Laconi dan papa
Baharuddin Laconi yang selalu memberikan motivasi dan dorongan moril
yang tinggi buat ananda. Kepada suamiku tercinta Ir. Rubi Herman Sutopo
serta kedua putra putriku Panji Hardian dan Riskina Juwita yang dengan tulus
hati, penuh pengorbanan, memberikan
kesernpatan dan semangat yang tinggi bagi penulis sehingga dapat
menyelesaikan pendidikan ini dengan baik.
Penulis sadar bahwa tulisan ini rnasih jauh dari kesempurnaan narnun
penulis berharap semoga hasil dan tulisan ini dapat rnernberikan surnbangan
kepada kernajuan ilmu pengetahuan dan teknologi.
Bogor, Pebruari 1998
DAFTAR
IS1
Nomor
HalamanRINGKASAN
...
i ABSTRAK...
vii JUDUL...
LEMBAR PENGESAHAN
...
RIWAYAT HlDUP
...
KATA PENGANTAR
...
DAFTAR IS1
..
.. .
...
.
...
...
... .
... ...
.
..
.
... ...
...,
... ... ... .. . ... ... ... .
DAFTAR TABEL...
DAFTAR GAM BAR... . . .. . ... . . .. . . .
...
.
..
.
. . .
. .
..
.
.
.. .
. . ..
. . .. . . .. . .. . . .. . . .. . . .. . .
DAFTAR LAMPIRAN.. . . ... . . ... . ... . . ... . . .. . .
...
.
....
... .
.. .
...
. .. . . .. . . .. . .
I PENDAHULUAN...
Latar Belakang
...
Tujuan Penelitian...
ix X xi xii xi v xv xvi xvii 1 1 3
Manfaat Penelitian
...
3 II TINJAUAN PUSTAKA...
4 Potensi Limbah Perkebunan Kakao...
4 Struktur Dinding Sel Tanaman...
8 Hemiselulosa...
9 Lignin...
10 Teknologi Pengolahan Pakan Berserat...
$2
Biofermentasi dengan
Phanerochaete chrysosporium
...
...
13
Sistern Pencernaan Ternak Rurninansia
...
Sistern Evaluasi Pakan...
Ill MATERI DAN METODE
...
Percobaan 1...
...
Percobaan 2
...
Metode AnalisaAnalisis Data
...
IV HASlL DAN PEMBAHASAN...
Kornposisi Zat Makanan Pod Kakao
...
Kornposisi Fraksi Serat Pod Kakao...
Konsurnsi dan Kecernaan Ransum...
Metabolisrne Rumen dan Alantoin Urin...
...
Konsentrasi Asarn Lemak Terbang Total dan Individual
...:
Balans Energi dan Protein
...
... Pertarnbahan Bobot Badan dan Komposisi Tubuh TernakPendugaan Kebutuhan Nutrien dan Mutu Ransurn
...
Evaluasi Mutu Ransurn...
Tinjauan Aspek Ekonomi...
V KESIMPULAN DAN SARAN...
VI DAFTAR PUSTAKA...
DAFTAR TABEL
Nomor Judul Tabel Halaman
1. Potensi Produksi Kakao dan Pod Kakao di Indonesia ... 5
2.
Luas Tanam dan Produksi Kakao di Indonesia Tahun 1997 ... 53. Kornposisi Zat Makanan Pod Kakao, Rumput Gajah dan
...
Serat Sawit Berdasarkan Bahan Kering
.
.
...
....
7
4. Komposisi Bahan Makanan dan Zat Makanan dalam
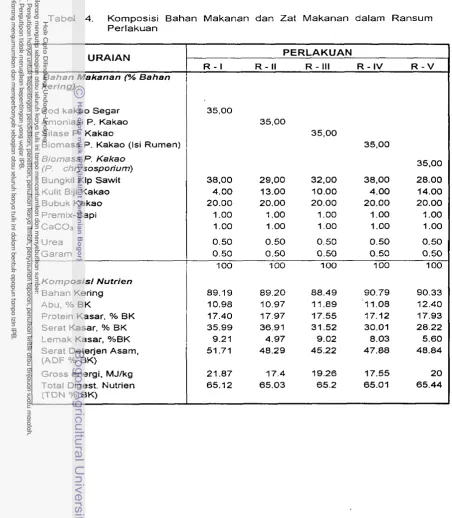
...
Ransurn Perlakuan 33
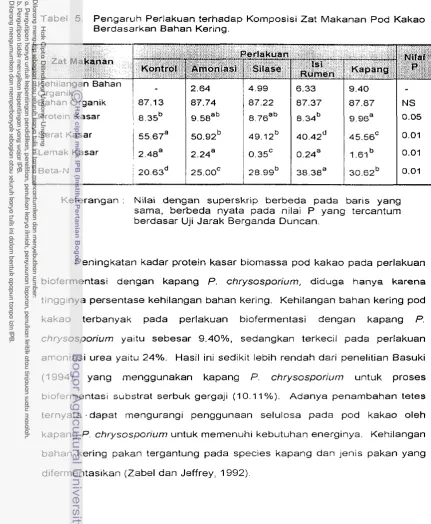
5.
Pengaruh Ransurn Perlakuan terhadap Komposisi Zat MakananPod Kakao Berdasarkan Bahan Kering . . . 44 6. Pengaruh Perlakuan terhadap Komposisi Serat Pod Kakao dan
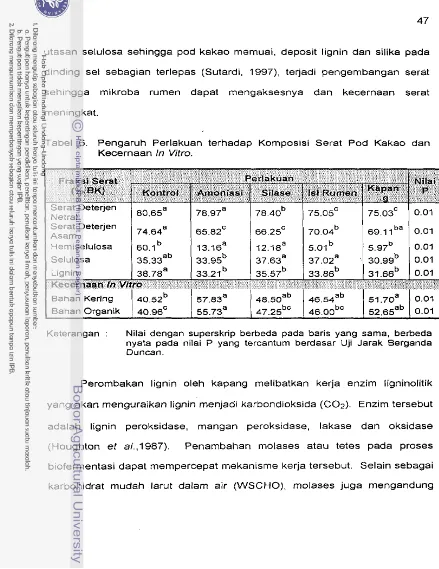
...
Kecernaan In vitro 47
7 . Pengaruh Ransurn Perlakuan terhadap Konsurnsi Zat-Zat
Makanan ... 53
8 . Pengaruh Ransurn Perlakuan terhadap Kecernaan Zat-Zat
...
Makanan
55
9. Pengaruh Ransum Perlakuan terhadap Parameter Metabolisme
Rumen dan Atantoin Urin ...
59
10. Pengaruh Ransum Perlakuan terhadap Konsentrasi VFA Individual Cairan Rumen, Nisbah C2/C3 dan Angka Non
Glukogenik Rasio (NGR)
...
.
.
.
......
... 6511. Pengaruh Ransum Perlakuan terhadap Neraca Energi ... 69
12. Pengaruh Ransurn Perlakuan terhadap Pertambahan Bobot
DAFTAR GAMBAR
Nomor Judul Garnbar H a l a m a n
1. Satuan
-
Satuan Penyusun Lignin ... 112. Skerna Fermentasi Karbohidrat dalam Rumen ... 20 ...
3. Metabotisme Protein pada Ternak Ruminans~a 21
4. Pengaruh Ransurn Perlakuan terhadap Kandungan Serat
Deterjen Netral, Selulosa dan Lignin Pod Kakao
...
495.
Perubahan Struktur Serat Pod Kakao setelah PerlakuanPercobaan ... 50
6.
Pengaruh Ransurn Perlakuan terhadap Kecernaan Protein,...
Serat Deterjen Netral (NDF), dan Serat Deterjen Asam (ADF)
57
7.
Pengaruh Ransurn Perlakuan terhadap Sintesis ProteinMikroba dan Ekskresi Allantoin di Urin ... 64
8. Pengaruh Ransum Perlakuan terhadap Konsentrasi Asam
Prpionat, Nisbah AcetaUPropionat dan Angka Non Glukonik
Rasio (NGR) ... 68
...
9. Pengaruh Ransum Perlakuan terhadap Partisi Energi 71
10. Pengaruh Ransum Perlakuan terhadap Konsumsi Bahan Kering, Pertambahan Bobot Badan Harian (PBBH) dan
DAFTAR
LAMPIRAN
Nomor
Judul Lampiran
Halaman
1. Analisa Ragarn untuk Bahan Organik Pod Kakao (%)
...
932. Analisis Ragarn dan Uji Jarak Duncan untuk Protein Kasar
Pod Kakao
...
933. Analisis Ragam dan Uji Duncan untuk Serat Kasar Pod
Kakao (%)
...
944. Analisis Ragam dan Uji Duncan untuk Lernak Kasar Pod
Kakao (Oh)
...
945.
Analisis Ragam dan Uji Duncan untuk Beta-N Pod Kakao(%)
...
956 . Analisis Ragam dan Uji Duncan untuk Serat Deterjen Netral
...
(NDF)Kakao(%) 95
7. Analisis Ragarn dan Uji Duncan untuk Serat Deterjen Asam
(ADF) Kakao (%)
...
968 . Analisis Ragam dan Uji Duncan untuk Herniselulosa Pod
Kakao (%)
...
969.
Analisis Ragarn dan Uji Duncan untuk Selulosa Pod Kakao(%)
...
97
10. Analisis Ragarn dan Uji Duncan untuk Lignin Pod Kakao (%)
...
971 . Nilai Rataan Kehilangan Bahan Kering Pod Kakao
...
9812.
Analisis Ragarn dan Uji Duncan untuk Pertambahan BobotTubuh Harian (kg/h)
...
9813. Analisis Ragarn dan Uji Duncan untuk Konsumsi Bahan
...
Kering (g/kg0.75/h) 99
14. Analisis Ragarn dan Uji Duncan untuk Konsumsi Bahan
...
Analisis Ragarn dan Uji Duncan untuk Konsurnsi Protein
(ghg B B ~ . ~ ~ / ~ )
...
100 Analisis Ragam dan Uji Duncan untuk Konsurnsi SeratDeterjen Netral (glkg B B ~ ~ ' ~ / ~ )
...
100 Analisis Ragarn dan Uji Duncan untuk Konsumsi SeratDeterjen Asarn
...
101 Analisis Ragarn dan Uji Duncan untuk Konsumsi Protein(g/kg B B ' . ~ ~ / ~ )
...
101 Analisis Ragarn dan Uji Duncan untuk Efisiensi PenggunaanRansurn (EPR)
...
102...
Analisis Ragarn dan Uji Duncan untuk Konversi Ransurn 102
...
Analisis Ragam untuk Kecernaan Bahan Kering (%) 103
...
Analisis Ragam untuk Kecernaan Bahan Organik (%) 103
...
Analisis Ragarn untuk Kecernaan Serat Kasar (%) 103
Analisis Ragam dan Uji Duncan untuk Kecernaan Nitrogen
(%)
...
104Analisis Ragarn dan Uji Duncan untuk Kecernaan Serat
Deterjen Netral (%)
...
104 Analisis Ragam dan Uji Duncan untuk Kecernaan SeratDeterjen Asam (%)
...
.
.
...
105Analisis Ragarn dan Uji Duncan untuk Kecernaan Energi (%)
...
105Analisis Ragam untuk Derajat Keasarnan (pH) Cairan
Rumen
...
106 Analisis Ragam dan Uji Duncan untuk Konsentrasi AmoniaCairan Rurnen (mM)
...
106Analisis Ragam dan Uji Duncan untuk Konsentrasi Asarn
Lemak Terbang Total (mM)
...
107Analisis Ragarn dan Uji Duncan untuk Allantoin Urin (glh)
...
108 Analisis Ra am dan Uji Duncan untuk Konsumsi Energi9
(MJlkg
8
8
'
-
'lh)...
109 Analisis Ragam dan Uji Duncan untuk Energi Tercerna...
109 Analisis Ra am dan Uji Duncan untuk Energi Termetabolis9
(MJlkg BBO- 'lh)
...
1 10Analisis Ra am dan Uji Duncan untuk Produksi Panas
9
(MJI~~ B BO- '/h)
...
1 10Analisis Ra am dan Uji Duncan untuk Retensi Energi
3
...
(MJlkg BB' 'lh) 1 1 ?
Analisis Ragam untuk Rasio MEIDE
...
11 1 Analisis Ragam dan Uji Duncan untuk Glukosa Darah(mg/lOOml)
...
1 12 Analisis Ragam dan Uji Duncan untuk Konsentrasi AsamAcetat Cairan Rumen (mM)
...
1 12 Analisis Ragam dan Uji Duncan untuk Konsentrasi AsamPropionat Cairan Rumen (mM)
...
1 13 Analisis Ragam dan Uji Duncan untuk Konsentrasi AsamButirat Cairan Rumen (mM)
...
1 13 Analisis Ragam dan Uji Duncan untuk Konsentrasi Asam...
Isobutirat Cairan Rumen (mM) 11 3
Analisis Ragarn dan Uji Duncan untuk Konsentrasi Asam
Valerat Cairan Rumen (mM)
...
114 Analisis Ragam dan Uji Duncan untuk Konsentrasi NisbahAcetaffPropionat (C2lC3) Cairan Rumen
...
114 Analisis Ragam dan Uji Duncan untuk Konsentrasi Non49. Analisis Ragam untuk Lemak Tubuh Ternak (%)
...
11650. Analisis Ragam dan Uji Duncan untuk Deposisi Protein
Tubuh Ternak (%)
...
11651. Analisis Ragam dan Uji Duncan untuk Deposisi Lemak
Tubuh Ternak (%)
.. .
...
.
. . ... .. . . .
.
. . . ... .
.. .
.
.
...
...
. .
.. . . ... . .. . . .. .
...
... . . . .. . 1 17PENDAHULUAN
Latar
BelakangSemakin berkurangnya lahan pertanian serta rendahnya mutu hijauan
dan rerumputan, rnenjadi pendorong bagi usaha pengadaan pakan alternatif.
Untuk mengatasi permasalahan tersebut, limbah tanaman perkebunan,
khususnya kakao dan kelapa sawit dapat dijadikan sumberdaya pakan
alternatif untuk ternak ruminansia. Melihat ketersediaannya, limbah tanaman
perkebunan ini dapat dijadikan pakan andalan karena produksinya melimpah
dan terkonsentrasi dalam wilayah tertentu sehingga dapat dijadikan titik tolak
pertumbuhan agroindustri pakan.
Kakao (Theobroma cacao) adalah sa[ah satu tanaman perkebunan
yang penanamannya berkembang dengan cepat. Menurut Direktorat Jendral
Perkebunan produksi coklat di Indonesia tahun 1997 telah mencapai 332.929
ton dengan luas areal 610.876 Ha. Buah kakao terdiri atas 73% kulit buah
kakao atau pod kakao dan 27% isi buah yang terdiri dari kulit biji dan
plasenta. Meningkatnya produksi kakao, akan rneningkatkan pula produksi
limbahnya terutama pod kakao. Pod kakao sangat tinggi kandungan airnya,
sehingga mudah membusuk. Penyebaran pod kakao di sekitar tanaman
kakao dapat menyebabakan infeksi jamur Phytophfora palmivora, juga
dikenal dengan 'Black Pod Diseases'. Jamur ini dapat menyebabkan
Melihat potensi produksi yang demikian besar, perlu dikaji lebih jauh
peluang pemanfaatannya sebagai pakan serat utama sebagai pengganti
rurnput. Beberapa studi telah menunjukkanbahwa pod kakao dapat sebagai
sumber energi tanpa mempengaruhi kinerja ternak. Akan tetapi adanya
alkaloid theobromine (3,7-dimethyl xanthine) membatasi penggunaannya
sebagai pakan. Selain itu pod kakao mengandung serat yang sulit dicerna.
Kandungan ligninnya mencapai 38,75%, sebagian besar terdapat dalam
pelindung isi buah sehingga keras. Selain itu pod kakao juga mempunyai
daya serap air yang tinggi dan cenderung mernbentuk gel, sehingga daya
larutnya rendah.
Upaya untuk meningkatkan nilai guna pod kakao, dilakukan penerapan
teknologi pakan seperti biofermentasi dan amoniasi urea. Bioferrnentasi pod
kakao dengan kapang Phanerochaete chrysosporium diharapkan dapat
mendegradasi ikatan lignoselulosa dan menurunkan kandungan lignin.
Keragaman respon pakan terhadap urea dalam pengolahan amoniasi.
menyebabkan pemakaian urea tidak dapat digeneralisasikan akan tetapi
harus sesuai dengan sifat fisik dan komposisi kimia pakan.
Selain pod kakao, limbah kakao lainnya adalah kulit biji kakao dan
powder (bubuk) kakao. Ditinjau dari komposisi zat makanannya kedua
limbah ini dapat dijadikan pakan konsentrat bersama-sama dengan bungkil
kelapa sawit. Kedua limbah kakao ini juga belum dimanfaatkan untuk pakan
Tujuan Penelitian
Berdasarkan pemikiran di atas, serangkaian percobaan dalam
penelitian ini bertujuan untuk :
1. Mampu meningkatkan nilai nutrisi pod kakao dengan biofermentasi
Phanerochaete chrysosporium dan isi rumen serta amoniasi urea
.
2.
Menentukan teknologi pengolahan yang tepat, efektif dan efisien untukmendayagunakan limbah kakao.
3. Mampu mendapatkan formula ransum komplit dengan bahan baku
limbah kakao dalam ransum pertumbuhan sapi daging dan sapi perah.
4. Memberikan sumbangan data parameter metabolik untuk menduga
kebutuhan nutrien dan sistem evaluasi mutu ransum.
Manfaat Penelitian
Hasil penelitian ini diharapkan dapat memberikan masukan atau
informasi yang sangat berguna bagi PT. Perkebunan (PTP), khususnya
perkebunan kakao tentang pendayagunaan lirnbah kakao yang optimum.
Selain dapat menunjang pengembangan peternakan juga sebagai upaya
mencegah pencemaran lingkungan akibat menumpuknya pod kakao.
Juga diharapkan membuka peluang untuk berkembangnya agroindustri
TINJAUAN PUSTAKA
Potensi Llrnbah Perkebunan Kakao
Tanarnan kakao (Theobroma cacao
L.)
merupakan tanarnan yangterrnasuk dalarn divisi spermafophyta, kelas dicotyledoneae, ordo marvales.
famili sferculiaceae, genus theobroma dan species Theobroma
cacao
L(Siturnorang, 1971). Tanarnan ini dibudidayakan di lndonesia dalam bentuk
perkebunan rnilik rakyat, pernerintah dan perkebunan besar swasta. Masa
depan tanarnan kakao nampaknya cukup cerah dengan semakin
rneningkatnya kebutuhan dunia akan kornoditi kakao.
Limbah perkebunan kakao terdiri dari kulit buah kakao (pod kakao),
kulit biji dan placenta. Berdasarkan hasil penelitian Darwis et a/. (1 988), buah
kakao terdiri dari pod kakao = 75.67%; kulit biji kakao = 21.74% dan placenta
= 2.59%. Luas areal tanaman kakao rneningkat dengan laju peningkatan
sebesar 5.7%/tahun, sedangkan laju peningkatan produksi Rakao sebesar
12.9l%/tahun. Potensi produksi lirnbah kakao terus meningkat sejalan
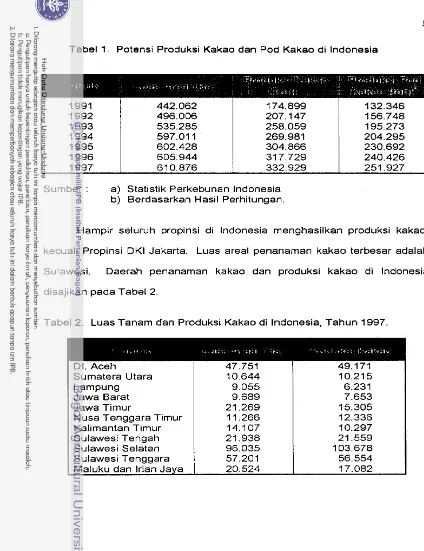
Tabel 1. Potensi Produksi Kakao dan Pod Kakao di lndonesia
Sumber : a) Statistik Perkebunan lndonesia
b) Berdasarkan Hasil Perhitungan.
Hampir seluruh propinsi di lndonesia menghasilkan produksi kakao,
kecuali Propinsi DKI Jakarta. Luas areal penanaman kakao terbesar adalah
Sulawesi. Daerah penanaman kakao dan produksi kakao di lndonesia
disajikan pada Tabel 2.
Tabel 2. Luas Tanam dan Produksi Kakao di Indonesia, Tahun 1997.
Dl. ~ c e h Sumatera Utara Lampung Jawa Barat Jawa Timur
Nusa Tenggara Timur Kalimantan Timur
Sulawes~ Tengah
[image:176.523.40.464.20.571.2] [image:176.523.40.467.38.572.2]Berdasarkan komposisi zat makanannya, pod kakao setara dengan
rumput gajah mengandung TDN 53.0% dan protein 8.75% (Darwis
et
a / . ,1988). Akan tetapi pod kakao tidak dapat dimanfaatkan sebagai pakan serat
secara langsung. Beberapa faktor kendala seperti kandungan lignin tinggi
(38.78%), adanya alkaloid theobromine dan rendahnya kadar amonia
(0.74 mM). Limbah kakao mengandung zat alkaloid yaitu theobromine
(3.7-dimethylxanthine) yang menyebabkan keracunan pada ternak. Kadar
theobromine kulit biji kakao lebih tinggi daripada pod kakao. Pod kakao
mengandung 0.17-0.22% theobrornine (Wong dan Hasan, 1988). Pada level
rendah zat ini dapat menstimulasi myocardial dan diuresis pada ternak
unggas. Alkaloid ini diduga dapat menghambat pertumbuhan mikroba rumen
ternak ruminansia, akibatnya kemampuan mencerna juga menurun.
Cendawan mikotoksin Phytophthora palmivora (Butler) dapat tumbuh dan berkembang pada pod kakao yang tertimbun di bawah pohon kakao
dalam kondisi kelembaban tanah dan udara yang tinggi. Cendawan ini dapat
menjadi hama dan penyakit busuk buah, hawar daun dan kanker batang
pada tanaman kakao. Akan tetapi jamur ini tidak dapat bertahan hidup jika
melewati saluran pencernaan sapi (Lopez et
a/.,
1984).Pod kakao sebagai pakan serat dengan kandungan protein rendah,
lebih tepat digunakan sebagai sumber energi bagi ternak ruminansia (Smith
dan Adegbola, 1982). Hal senada juga dikatakan (Sukri et at., 1986), bahwa
pod kakao dapat sebagai sumber energi tanpa mempengaruhi kinerja ternak.
Hasilnya menunjukkan bahwa pod kakao tanpa pengolahan yang diberikan
bersama konsentrat pada 12 ekor sapi pedaging selama 84 hari,
mengakibatkan terjadinya penurunan bobot badan ternak dengan pernberian
di atas 40% , meskipun konsumsinya rneningkat (Smith dan ~ d e g b o l a , 1982).
Percobaan pada sapi laktasi telah dilakukan. Ternyata pod kakao
cukup palatabel. Penggunaannya sarnpai taraf 15% tidak berpengaruh nyata
terhadap produksi dan kualitas air susu. Produksi 4 % Fat Corrected Milk
(FCM) dan produksi lemak tertinggi dicapai pada taraf 5 % penggunaan pod
kakao (Gumanizar, 1993). Pod kakao setara dengan rurnput gajah
mengandung Total Digestible Nutrient (TDN) 53% dan protein 7.16% (Sutardi
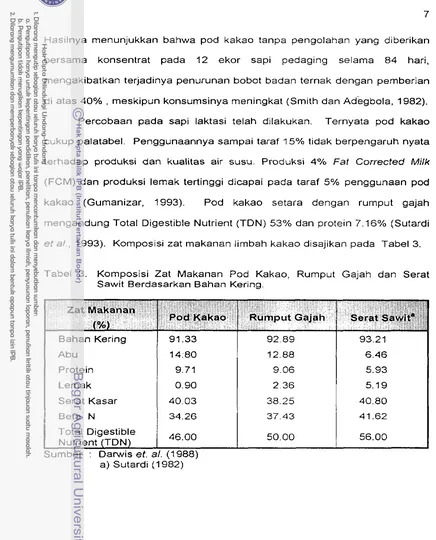
e t a l . , 1993). Kornposisi zat makanan lirnbah kakao disajikan pada Tabel 3.
Tabel 3. Komposisi Zat Makanan Pod Kakao, Rurnput Gajah dan Serat Sawit Berdasarkan Bahan Kering.
Bahan Kering Abu
Protein Lemak
Serat Kasar Beta N
Zat
Makanan
(Oh)
a) Sutardi (1 982)
Pod
KakaoTotal Digestible Nutrient (TDN)
Rumput Gajah Serat Sawita
46.00
'.
[image:178.528.40.476.35.575.2]Penggunaan pod kakao sebagai pengganti rumput pada ransum sapi
Holstein jantan sebesar 0; 15; 30 dan 45% memberikan indikasi bahwa
semua parameter konsumsi meningkat sejalan kenaikan penggunaan pod
kakao. Akan tetapi semua parameter kecernaan menurun. Kecernaan in
vitro
ransum menurun dari 58.3% menjadi 52.0%. Kemungkinan penurunan ini
disebabkan oleh kenaikan kadar lignin ransum.
Struktur Dinding
Sel
Tanaman
Struktur sel tanaman terdiri dari isi sel dan dinding sel. Sebagian
besar komponen penyusun dinding sel adalah fraksi karbohidrat. Fraksi
karbohidrat dalam pakan dikelompokkan menjadi dua bagian yaitu
karbohidrat non struktural (fraksi yang mudah tersedia) dan karbohidrat
struktural (fraksi serat). Fraksi karbohidrat dibagi menjadi monosakarida dan
turunannya (glukosa, fruktosa dan silosa); oligosakarida yang terdiri dari 2-1 0
unit sakarida (sukrosa, fruktosa rantai pendek dan raffinosa) serta
polisakarida (pati, fruktosa, selulosa dan hemiselulosa). Sebagian besar
fraksi karbohidrat struktural menjadi komponen penyusun dinding sel pada
pakan serat, di antaranya adalah selutosa, hemiselulosa, lignin dan silika.
Akan tetapi selulosa seringkali berikatan dengan lignin sehingga membentuk
ikatan lignoselulosa. lkatan komplek ini sulit dipecah oleh mikroba rumen.
bagi ternak rurninansia. Pod kakao rnengandung ikatan lignoselulosa yang
tersusun dari selulosa terikat dengan lignin. Komponen ini dapat dikonversi
menjadi produk lebih sederhana dengan menggunakan kapang dan arnoniasi
dengan urea (Lynch. 1987).
Karakteristik struktur polimer dinding sel tanaman lebih efektif
dipelajari dengan rnetode fisik dibandingkan dengan metode kimia. Metode
kirnia seperti Analisa Proksionat dan Van Soest hanya rnengukur kornposisi
serat dinding sel. Pengamatan secara visual perubahan struktur serat
dinding sel tanaman rnenggunakan mikroskop elektron. Ada dua macam
mikroskop elektron yaitu Scanning electrone Microscopy (SEM) dan
Transmision Electrone Microscopy ( T E M ) . Metode S E M dapat dipergunakan
untuk rnernpelajari struktur anatorni tanarnan, perubahan struktur polirner
dinding sel hijauan karena perlakuan pengolahan secara kirnia dan biologis,
degradasi dan kecernaan dinding set hijauan oteh mikroorganisme di dalam
rurnen (Van der rneer, 1989).
Hemiselulosa dan Selulosa
Struktur dan kornposisi kimia dinding sel tanaman bervariasi rnenurut
species, umur, varietas dan tipe sel tanarnan. Secara urnurn dinding set
tanaman terdiri dari dinding primer, dinding sekunder dan lamella. Dinding
primer tanaman monokotil dan dikotil merupakan serat-serat selulosa sebagai
polisakarida dengan kandungan glikoprotein (Orphin, 1984). Selulosa adalah
unsur utarna pembentuk kerangka tanaman dan penyusun dinding sel
berantai lurus dan panjang dengan ikatan
p
-1.4 unit glukosa. Selulosarnerupakan sumber energi yang sangat potensiat bagi ternak rurninansia.
Adanya mikroorganisme anaerobik di dalam rumen membantu proses
pencernaan selulosa untuk dapat rnembebaskan sejurnlah energi (Arora,
1982). Mikroba rurnen mencerna selulosa di dalarn rumen dan retikulum
dengan bantuan enzirn selulase yang dihasilkan oleh jasad renik di dalam
saluran pencernaan, menjadi selubiosa dan selanjutnya oleh enzirn selubiose
diubah menjadi glukosa. Hasil akhir adalah asam-asam lemak terbang yang
terdiri dari asarn acetat, asam propionat dan asarn butirat serta gas
karbondioksida (Con) dan methan (CH4)
Herniselulosa adalah bagian dinding sel yang lebih rnudah didegradasi
dibandingkan dengan selulosa dan lignin. Hemiselulosa merupakan
kelornpok polisakarida yang berantai lurus yaitu hornopolisakarida dan
heterosakarida yang terdiri dari xilosa, manosa, galaktosa, arabinosa dan
gtukosa (Puls dan Poutanen, 1989).
Lignin
Lignin rnerupakan senyawa polimer korniferil alkohol yang mernbentuk
ikatan ether dengan selulosa dan herniselosa dalam jaringan tanaman dan
selalu terdapat dalam senyawa kompleks dinding sel (Nolan et.
a/.,
1988).Setelah selulosa, lignin merupakan bahan .organik terbesar di burni.
Lignin disusun oleh unit-unit fenil propen, yaitu korniferil, sinafil dan para
dihubungkan satu sama lain dalam berbagai macam ikatan tidak beraturan
atom C-C dan C-0-C (Gambar 1).
[image:182.526.44.463.20.578.2]a Korniferil Alkohol b. Sinafil Alkohol c . Para Kumaril Alkohol
Gambar 1. Satuan-Satuan Penyusun Lignin
Lignin memiliki kandungan energi yang sangat potensial, akan tetapi
sangat sulit dirombak oleh rnikroba rumen, terutama pada pemecahan cincin
aromatiknya. Cincin ini hanya dapat dipecah pada keadaan aerob,
sedangkan rumen dalam kondisi anaerob (Orpin, 1984). Degradasi lignin
masih mungkin terjadi dengan proses degradasi oleh jamur (Leisola dan
Garcia, 1989). Hanya sebagian kecil kapang yang mempunyai kemampuan
merornbak lignin, khususnya kapang dari kelas Basidiornycetes yaitu kapang
Phanerochaete chrysosporium.
Perombakan lignin oleh mikroorganisme melibatkan enzim ligninolitik
yaitu lignin peroksidase, mangan peroksidase, Iakase dan oksidase. Enzim
lignolitikk ini akan menguraikan lignin menjadi karbondioksida (Houghton et
Teknologi Pengolahan Pakan Berserat
Kelompok pakan yang tinggi fraksi seratnya memerlukan pengolahan
terlebih dahulu untuk rneningkatkan fermentabilitasnya
.
Terutama untuk memutuskan ikatan lignoselulosa yang sulit dicerna oIeh mikroba rumen.Peningkatan ferrnentabilitas pakan serat dapat dilakukan dengan pengolahan
kimia (amoniasi dengan urea, alkali dengan abu dan NaOH); pengolahan fisik
(extruding, pelleting dan pemanasan) serta pengolahan biologis (ferrnentasi
dengan mikroorganisme).
Fermentasi pakan serat di dalam rurnen merupakan suatu sistem yang
komplek dan dipengaruhi oleh adanya interaksi dinamik antara faktor ternak,
pakan dan populasi mikroba. Faktor pakan diantaranya struktur kimia dan
fisik serat. Fraksi serat yang sukar sekali dicerna adalah lignin. Adanya
proses delignifikasi baik secara kimia, fisik dan biologis, diharapkan akan
meningkatkan laju fermentasi pakan serat di dalam rumen. Untuk mencerna
serat, ternak ruminansia sepenuhnya tergantung pada peranan mikroba
rumen. Ternak ruminansia tidak memproduksi enzim yang dapat
menghidrolisis selulosa atau hemiselulosa.
Upaya peningkatan nilai guna pod kakao dan mengatasiberlimpahnya
produksi limbah kakao perlu dilakukan penerapan teknologi pakan seperti
amoniasi urea dan biofermentasi dengan bantuan mikroorganisme
.
Salah satu mikroorganisme tersebut yaitu kapang Phanerochaete chrysosporiurnBiofermentasi Kapang P. chrysosporium Burdsall ATCC 34541
Peningkatan kualitas dan ferrnentabilitas pakan serat dapat dilakukan
dengan beberapa pengolahan diantaranya biofermentasi dengan isi rumen
dan kapang pendegradasi serat. Bioferrnentasi rnerupakan proses
perubahan kimia pada substrat sebagai hasil kerja enzirn dari
rnikroorganisme dengan rnenghasilkan produk tertentu. Proses ini berjalan
tergantung pada jenis sustrat, kapang dan kondisi lingkungan yang
rnempengaruhi pertumbuhan dan rnetabolisrne kapang. Selama ferrnentasi
berlangsung, terjadi perubahan terhadap pH, kelembaban dan aroma serta
perubahan komposisi zat makanan seperti protein, lemak, serat kasar.
karbohidrat, vitamin, dan mineral.
Winarno, (1980) rnengatakan bahwa substrat yang mengalami
fermentasi biasanya memiliki nilai gizi yang lebih tinggi daripada bahan
asalnya. Hal ini dikarenakan sifat katabolik dan anabolik rnikroorganisrne
sehingga mampu rnernecah kornponen yang lebih kompleks menjadi rnudah
tercerna. Proses biofermentasi diharapkan akan rnerombak struktur jaringan
kirnia dinding sel, pernutusan ikatan lignosellulosa dan penurunan kadar
lignin. Pakan serat yang mengalami fermentasi dengan kapang akan
meningkat kecernaan nutriennya (Puls dan Poutanen, 1989).
Kapang Phanerochaete chrysosporium adalah kapang pendegradasi
lignin dari kias Basidiomycetes, membentuk sekumpulan miselia dan
Kapang ini adalah kapang "White rot" yang mempunyai kemampuan kuat
merombak lignin secara efektif dengan cara menghasilkan enzim
peroksidase ekstraseluler, berupa lignin peroksidase (LIP) dan mangan
peroksidase (MnP) (Vallie et a/., 1992) serta pelapukan kayu secara mikrobial
(Erikson dan Vallander, 1980). .Enzim ligninolitik ini dapat memutuskan ikatan
lignoselulosa. Kapang ini juga mendegradasi berbagai senyawa organik
pencemar ltngkungan (Bumpus dan Aust, 1987), sehingga memberikan
harapan untuk digunakan dalam proses delignifikasi pakan dan proses
pengolahan limbah yang mengandung derivat lignin dan senyawa toksik.
Sistern kerja enzim peroksidase ekstraseluler yaitu tidak memisahkan
serat-serat dengan cara melarutkan lignin yang ada dalam lamela tengah,
akan tetapi dengan cara melunakkan dan memecahkan dinding-dinding serat
dan terkadang juga melepaskan pita-pita serat mikrofibrilnya. Lebih lanjut
(Totter, 1990) menjelaskan bahwa kunci reaksi degradasi lignin oIeh kapang
P. chrysosporium adalah biokatalis ligninase yang mengkatalis oksidasi cincin
aromatik lignin untuk membentuk radikal-radikal kation. Selanjutnya senyawa
ini akan melepaskan ikatan-ikatan inti pada cincin aromatik.
Kapang 'White Rot" lainnya yang dapat dipergunakan untuk proses
biofermentasi adalah Trichoderma viride dan Fusarium sp. Nilai gizi pod
kakao dapat ditingkatkan melalui proses fermentasi dengan menggunakan
kapang Trichoderma viride sehingga setara dengan hijauan yang umum
digunakan sebagai pakan ternak ruminansia, akan tetapi kecernaan nutrien
biologis telah dilakukan sebagai upaya meningkatkan nilai gizi pod kakao.
Kornbinasi perlakuan alkali dengan menggunakan 6% NaOH dan ensilase
menghasilkan penurunan daya cerna terkecil yaitu 5.3% (Wong Hee Kum
ef
a/., 1988). Silase pod kakao dengan penambahan bahan 1% urea dan 6%
tetes dengan lama penyimpanan 1 minggu menunjukkan kualitas silase yang
terbaik dengan produksi amonia (NH3) sebesar 6.072 mM (Sa'diyah, 1992).
Amoniasi
Pakan BerseratPeningkatan nilai gizi pakan serat juga dapat dilakukan dengan
pengolahan secara kimia, seperti pengolahan alkali dengan NaOH dan
Ca(OH)2 serta amoniasi dengan amonia dan prekusor amonia (urea).
Perlakuan kimia yang telah banyak dilakukan adalah dengan urea, dikenal
sebagai proses amoniasi. Dibandingkan dengan perlakuan kimia lainnya,
penggunaan urea lebih murah, tidak berbahaya dan mudah dilakukannya.
Urea digunakan sebagai sumber amonia pada perlakuan kimia untuk jerami
di negara berkembang. Urea mengalami dekomposisi menjadi COz dan NH3
dengan adanya tekanan panas. Efektifitas perlakuan amoniasi terhadap
limbah pertanian berserat tinggi, dipengaruhi oleh tingkat pemberian amonia,
suhu, lama perlakuan dan kadar air, serta tipe dan kualitas bahan yang
diperoses (Davis, 1983). Umumnya pakan berserat rnempunyai kandungan
nitrogen rendah dan kandungan serat yang tinggi. Proses amoniasi
diharapkan dapat rnengatasi kendala tersebut. Selain meningkatkan
kecernaan serat, juga meningkatkan degradasi protein yang berikatan
Arnoniasi dengan urea juga rnerupakan perlakuan alkali. Urea yang
ditambahkan ke dalarn pakan rnengalarni ureolitik rnenjadi arnonia ( N H 3 ) dan
C 0 2 dan enzirn urease dihasilkan oleh bakteri.yang terdapat dalarn pakan.
Bersama air pakan, N H 3 rnembentuk basa N H 4 0 H . Kelebihan arnoniasi
dengan perlakkuan lain adalah rnarnpu rnemasok nitrogen untuk rnikroba
rumen, dan juga proses pernbuatannya yang rnudah, rnurah, dan cepat.
Amoniasi jerarni padi dengan 5% urea rneningkatkan kecernaannya
dari 45% rnenjadi 57%. Kombinasi perlakuan penggilingan dan arnoniasi
pakan jerarni jagung dengan 3-4% urea dapat rneningkatkan konsumsi dan
daya cerna bahan kering
Berdasarkan penelitian sebelumnya amoniasi dengan berbagai tingkat
penggunaan urea untuk pod kakao kakao, tingkat pernakaian urea terbaik
adalah 1.5%. Penggunaan urea di atas level tersebut cenderung
menurunkan kecernaan in vitro dan degradasi in situ pod kakao (Sutardi
et
a/.,
1994). Level urea 1.5% dipergunakan dalarn arnoniasi pod kakao padapercobaan ini.
Sistem Pencernaan Ternak Rurninansia
Proses pencernaan pada ternak rumniansia dapat terjadi secara
rnekanis di rnulut, ferrnentatif oleh mikroba rumen dan secara hidrolisis oleh
enzirn-enzirn pencernaan. Ternak ruminansia rnempunyai lambung majemuk
yang terdiri dari retikulo rumen, ornasum dan abornasurn. Proses ferrnentasi
bantuan rnikroba rurnen (Satter dan Roffler, 1981 ). Rumen adalah ternpat
untuk proses ferrnentasi makanan yang masuk serta menyediakan energi
dan protein rnikroba untuk kebutuhan proses rnetabolisrne.
Jenis mikroorganisme yang terda~at dalam rumen adalah bakteri,
protozoa, fungi dan virus. Kecernaan pakan sangat tergantung dari peranan
-
rnikroba rurnen. Bakteri rnembentuk kotoni pada pakan serat selarna 5 menit,
protozoa selama 15 rnenit dan selama 2 jam untuk sporangia dan rizhoid
kapang (Derneyer, 1981). Oteh karena itu ternak yang rnendapatkan ransurn
dengan serat tinggi, kehadiran fungi sangat berperanan sekali dalarn
rnencerna pakan serat tersebut. Fungi rnernbentuk koloni pada jaringan
ikatan lignoselulosa partikel pakan (Fonty et a/., 1990). Adanya benang-
benang rhizoid pada fungi rnernungkinkan fungi dapat rnenernbus dinding
partikel, sehingga menciptakan akses bagi bakteri. Setelah itu bekerjasama
dengan bakteri selulolitik rnencerna serat.
Proses ferrnentasi di dalarn rumen oleh rnikroba yaitu rnenghidrolisa
karbohidrat menjadi rnonosakarida dan disakarida yang kernudian
difermentasi lebih lanjut rnenjadi asarn lernak terbang atau 'volatil fatty acid'
(VFA) terutarna asam acetat, asam propionat serta asarn butirat dan
kernudian diserap melalui dinding rurnen serta gas rnetan (CH4) dan gas
karbondioksida (Con) (Mc. Donald et a/., 1988). Asarn lemak terbang (VFA)
rnerupakan sumber energi utarna bagi ternak rurninansia. Menurut
mencapai 60
-
80% kebutuhan energi pada ternak rurninansia. Asarn lernakterbang (VFA) diserap diserap di rurnen, retikulurn dan ornasum (Fangel dan
Wagner, 1984). Kadar VFA cairan rurnen antara 80 - 160 rnM telah
mencukupi kebutuhan untuk sintesis protein rnikroba rumen yang optimal
(Sutardi
e t a / . ,
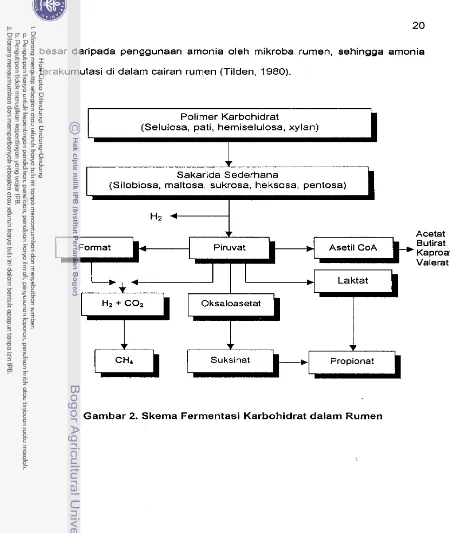
1983). [image:189.526.41.474.31.575.2]Ferrnentasi karbohidrat pakan di dalarn rurnen diperlihatkan pada
Gambar 2. Produksi volatil fatty acid utarna dalarn cairan rurnen adalah 65%
asarn acetat (C2), 25% asarn propionat (C3) dan 10% asarn butirat (C4).
Energi pakan dalam bentuk asarn lernak terbang rnencapai 75%. Sisanya
sebagai produksi gas rnetan (CH4) sebesar 12.4%, panas ferrnentasi 6.4%
dan sekitar 6.2% dipergunakan rnikroba rurnen sebagai sumber energi dalarn
bentuk adenosin triphosfat (ATP) (Clark dan Davis, 1983
dalam
Erwanto,1995). Kornponen asarn lernak terbang yang termasuk glukogenik yaitu
asarn propionat, sedangkan asarn asetat dan asarn butirat tidak terrnasuk
kelornpok metabolit glukogenik. Menurut Orskov (1 977) suatu nilai konstanta
yang disebut sebagai non
glukogenic ratio
(NGR) berhubungan erat denganproduksi gas rnethan. Terdapat korelasi positif antara keduanya. Makin
tinggi angka NGR rnaka produksi gas rnethan rnakin besar. Perhitungan nilai
angka NGR secara sederhana dirumuskan sebagai berikut :
A c e t a t
+
2 B u t i r a t + V a l e r a tN G R =
Untuk mensintesa protein mikroba yang optimal diperlukan
keseimbangan energi (VFA) dan nitrogen dalam bentuk N-NH3. Kekurangan
salah satu unsur ini dapat menghambat pertumbuhan mikroba rumen.
Pembentukan protein mikroba yang memerlukan energi,
memperlihatkan adanya keteroantungan antara metabolisrne protein dan
energi. Sumbangan protein mikroba sangat berarti untuk menyediakan
kebutuhan protein untuk induk semang (ternak). Orskov (1 982) mengatakan
sumber nitrogen untuk mikroba rurnen berasal dari a) degradasi protein
pakan, b) daur ulang urea melalui saliva, c) daur ulang urea melalui darah,
d) nitrogen endogen dari dinding rumen. Sumber nitrogen pakan berupa
protein murni dan nitrogen bukan protein (NPN). Urea adalah salah satu
sumber NPN. Proses pencernaan urea terjadi di dalam rumen. Urea
dihidrolisis rnenjadi NH3 dan Con. Proses hidrolisis ini berlangsung karena
adanya aktivitas enzim urease yang diproduksi oleh mikroba rumen (Cullison.
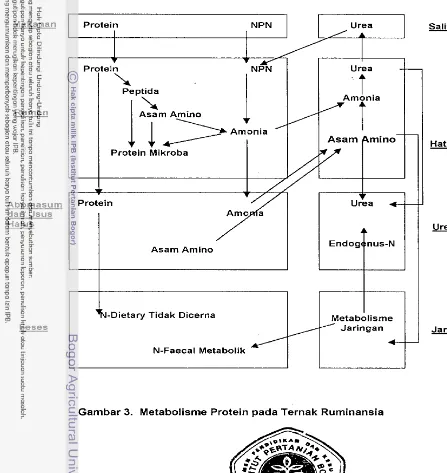
1982). Fermentasi protein di dalam rurnen menghasilkan asam amino,
peptida dan amonia. Skerna metabolisme protein pada ternak ruminansia
disajikan pada Garnbar 3.
Proses metabolisme protein di dalam rumen cukup kompleks. Arnonia
selain berasal dari protein, juga berasal dari senyawa nitrogen bukan protein.
Hasil ini dipergunakan lagi untuk mensintesa protein rnikroba. Jumlah
amonia yang dapat digunakan oleh bakteri, tergantung dari jurnlah bakteri
besar daripada penggunaan amonia oleh rnikroba rurnen, sehingga arnonia
terakumulasi di dalam cairan rumen (Tilden, 1980).
Polimer Karbohidrat
(SeluIosa, pati, herniselulosa, xylan)
-
.
Sakarida Sederhana
(Silobiosa, maltosa, sukrosa, heksosa, pentosa)
Acetat
Piruvat Asetil CoA Butirat Kaproat
[image:191.523.43.493.20.553.2]I Valerat