KERAGAMAN GEN PROLAKTIN EKSON EMPAT PADA
ITIK PEKING ITIK MOJOSARI PUTIH DAN ITIK PMP
SERTA ASOSIASINYA DENGAN SIFAT REPRODUKSI
MARLINDA INDRIATI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Keragaman Gen Prolaktin Ekson Empat pada Itik Peking, Itik Mojosari Putih dan Itik PMp serta Asosiasinya dengan Sifat Reproduksi adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Oktober 2014
RINGKASAN
MARLINDA INDRIATI. Keragaman Gen Prolaktin Ekson Empat pada Itik Peking, Itik Mojosari Putih dan Itik PMp serta Asosiasinya dengan Sifat Reproduksi. Dibimbing oleh CECE SUMANTRI dan TRIANA SUSANTI.
Prolaktin merupakan hormon polipeptida yang disekresikan oleh kelenjar pituitary anterior dan memiliki berbagai dampak terhadap aktifitas dan fungsi biologis semua hewan bertulang belakang (vertebrata). Tujuan dari penelitian ini adalah untuk menganalisis polimorfisme wilayah gen prolaktin ekson empat dan sebagian intron 3 dan 4 serta mengasosiasikan polimorfisme gen tersebut dengan sifat-sifat reproduksi itik. Ekstraksi yang dilakukan pada sampel darah menggunakan Genomic DNA Mini Kit (GeneAidTM DNA Isolation Kit) dimana sampel darah diperoleh dari 168 itik yang terdiri dari 19 ekor itik Peking, 36 ekor itik Mojosari putih dan 113 ekor itik silangan Peking Mojosari putih. Sepasang primer ekson 4 disusun berdasarkan database sekuens genom yang telah terpublikasi di Genbank dengan kode akses AB158611.
Analisis pensejajaran terhadap sekuens DNA didapat enam SNP yang terdiri dari dua mutasi transversi yaitu T3941G dan C3975A, satu mutasi transisi yaitu T4110C, dua delesi basa adenin pada posisi pada posisi 3724 pb (INDEL 3724A) dan 4031 pb (INDEL 4031A) dan insersi pada posisi 3939 pb (INDEL 3939A). Khusus untuk INDEL 3939A terjadi pada semua sampel itik penelitian.
Hasil analisis frekuensi alel menunjukan bahwa pada titik mutasi INDEL 3724A, T3941G, C3975A, INDEL 4031A dan T4110A pada itik sampel adalah polimorfik karena terdapat dua alel yang berbeda dengan frekuensi alel < 99%. Lain halnya dengan titik mutasi INDEL 3939A dimana hanya ditemukan satu jenis alel dengan hasil perhitungan frekuensi alel sebesar 100% dengan kata lain bahwa dalam populasi itik sampel INDEL 3939A adalah monomorfik.
Analisis statistik chi square mengenai perbedaan SNP terhadap jenis itik yang berbeda menunjukan bahwa tidak ada hubungan antara perbedaan jenis SNP yang ditemukan dengan ketiga jenis itik Peking, Mojosari putih dan PMp, sedangkan analisis asosiasi SNP terhadap sifat-sifat reproduksi itik dapat menunjukan bahwa lima titik mutasi yang ditemukan pada sekuens gen prolaktin yaitu INDEL 3724A, T3941G, C3975A, INDEL 4031A dan T4110C berpengaruh secara signifikan terhadap sifat produksi telur, persentase fertilitas dan daya tetas telur itik.
SUMMARY
MARLINDA INDRIATI. Prolactin Gene Diversity Exon 4 in Peking, White Mojosari and Peking Mojosari Duck Crossbred and Its Association with Reproduction Traits. Supervised by CECE SUMANTRI dan TRIANA SUSANTI
Prolactin is a polypeptide hormone secreted by the anterior pituitary gland and show has a variety of effects on biological activity and function in all vertebrates. The aim of this study was to characterize the polymorphism in intron regions and coding region of exon four of prolactin gene and gene polymorphisms associate with reproductive traits of ducks. Extraction is performed on a blood sample using the Genomic DNA Mini Kit (GeneAidTM DNA Isolation Kit) where
blood samples obtained from 168 ducks consisting of 19 Peking, 36 White mojosari and 113 Peking white mojosari ducks crossbreeds. Primer pair exon 4 have been prepared on the genome sequence databases have been published in GenBank with accession number AB158611.1.
Alignment analysis resulted 6 SNP there are two transverse mutation T3941G and C3975A and one transition mutation is T4110C. Deletion adenine base at position 3724 bp (INDEL 3724A) and 4031 bp (INDEL 34031). An insertion adenine base at position 3939 bp (INDEL 3939) occured in all duck samples.
Analysis allele frequency result showed that mutation of INDEL 3724A, T3941G, C3975A, INDEL 4031A and T4110A in duck samples are polymorphic because there are two different allele with frequency of allele < 99%. Another mutation of INDEL 3939A only founded one type of allele with resulted calculation of frequency allele is 100%, so that we can sugest that INDEL 3939A is monomorphic.
Statistical analysis of SNP and different types of ducks showed there was no significant relationship between the various types of SNP are found with three different types of duck, whereas analysis association SNP with duck reproduction traits can be shown that five SNPs were found in prolactin gene there are INDEL 3724A, T3941G, C3975A, INDEL 4031A and T4110C significantly was associated with reproductive traits of egg production, fertility and percentages hatchability egg (P <0.005).
© Hak Cipta Milik IPB, Tahun 2014 Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Ilmu Produksi dan Teknologi Peternakan
KERAGAMAN GEN PROLAKTIN EKSON EMPAT PADA
ITIK PEKING, ITIK MOJOSARI PUTIH DAN ITIK PMP
SERTA ASOSIASINYA DENGAN SIFAT REPRODUKSI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2014
Judul Tesis : Keragaman Gen Prolaktin Ekson Empat pada Itik Peking, Itik Mojosari Putih dan Itik PMp serta Asosiasinya dengan Sifat Reproduksi
Nama : Marlinda Indriati NIM : D151120061
Disetujui oleh Komisi Pembimbing
Prof Dr Ir Cece Sumantri, MAgrSc Ketua
Dr Ir Triana Susanti, MSi Anggota
Diketahui oleh
Ketua Program Studi Ilmu Produksi dan Teknologi Peternakan
Dr Ir Salundik, MSi
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
PRAKATA
Puji dan syukur penulis panjatkan kehadirat Allah Subhanahu wa Ta’ala atas segala nikmat dan karunia-Nya sehingga karya ilmiah ini dapat diselesaikan. Shalawat serta salam senantiasa tercurah kepada baginda nabi Muhammad SAW, kepada keluarganya, para sahabat serta umatnya hingga akhir zaman yang berpegang teguh pada tuntunannya.
Penelitian ini berjudul “Keragaman Gen Prolaktin Ekson Empat pada Itik Peking, Itik Mojosari Putih dan Itik PMp serta Asosiasinya dengan Sifat Reproduksi” merupakan penelitian kerjasama dengan Balai Penelitian Ternak Ciawi Bogor yang dilakukan sejak April 2013–Februari 2014. Tujuan pelaksanaan penelitian ini merupakan upaya dalam peningkatan performa produksi itik lokal Indonesia yaitu secara perkawinan dan seleksi ditingkat molekuler.
Penulis sampaikan terima kasih kepada Bapak Prof Dr Ir Cece Sumantri, MAgrSc dan Ibu Dr Ir Triana Susanti, MSi selaku komisi dosen pembimbing atas ilmu, bimbingan dan dukungannya sehingga memudahkan penulis dalam menjalankan penelitian dan penulisan tesis ini. Penulis juga sampaikan terima kasih kepada Dr Ir Niken Ulupi, MS selaku dosen penguji tesis atas masukan, saran, motivasi dan bimbingannya dan kepada Dr Ir Rarah Ratih Adjie Maheswari, DEA (Almarhumah) yang memberikan motivasi, arahan dan bimbingan selama menempuh kuliah.
Disamping itu, penghargaan tertinggi dan terimakasih sebesar-besarnya penulis sampaikan kepada keluarga tercinta Ayahanda Dayat Koyat, Ibu Salmiati Marlin SPd, Suamiku Sibro Malisi SH, adik-adik Nisa, Rizki, Syifa atas kasih sayang, dukungan dan doa yang tak henti, dan terakhir teman-teman seperjuangan pascasarjana ITP 2012, ABGSCi atas dukungan dan kebersamaannya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Oktober 2014
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vi
1 PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 2
Tujuan Penelitian 2
Manfaat Penelitian 2
2 TINJAUAN PUSTAKA 2
Itik Lokal 2
Itik Persilangan Peking Mojosari Putih 4
Gen Prolaktin 6
Penanda Molekuler Gen Prolaktin 7
Hubungan Keragaman Gen Prolaktin dengan Sifat Fenotipe 8
3 METODE 9
Lokasi dan Waktu 9
Materi 9
Prosedur 10
Prosedur Analisis Data 13
4 HASIL DAN PEMBAHASAN 14
Ekstraksi dan Amplifikasi Gen Prolaktin 14
Karakteristik Gen Prolaktin 15
Frekuensi Alel Gen Prolaktin 19
Analisis Asosiasi SNP dengan Sifat Reproduksi Itik 21
5 SIMPULAN DAN SARAN 24
Simpulan 24
Saran 25
DAFTAR PUSTAKA 25
LAMPIRAN 31
DAFTAR TABEL
1 Asosiasi polimorfisme gen prolaktin terhadap sifat reproduksi unggas 9 2 Kandungan nutrisi pakan komplit komersil untuk sampel itik penelitian 9 3 Analisis homologi itik PMp, Peking dan Mojosari putih dengan metode
BLAST 16
4 Analisis komposisi basa nukleotida pada itik PMp, Peking dan Mojosari
putih 16
5 Analisis deteksi mutasi gen prolaktin itik PMp, Peking dan Mojosari
putih 17
6 Analisis frekuensi genotipe gen prolaktin ekson empat 20 7 Analisis Singe Nucleotide Polymorphism terhadap jenis itik yang
berbeda 21
8 Analisis LS Mean ± SE INDEL 3724A terhadap paameter reproduksi 21 9 Analisis LS Mean ± SE T3941A terhadap parameter reproduksi 22 10 Analisis LS Mean ± SE C3975A terhadap parameter reproduksi 22 11 Analisis LS Mean ± SE INDEL 4031A terhadap parameter reproduksi 23 12 Analisis LS Mean ± SE T4110C terhadap parameter reproduksi 23
DAFTAR GAMBAR
1 Beberapa jenis itik lokal Indonesia a) itik Bayang, b) itik Pegagan, c) itik Magelang, d) itik Rambon, e) itik Pitalah dan f) itik Talang
3 2 Skema perkawinan itik tetua peking jantan dengan induk Mojosari putih 5 3 Rekonstruksi struktur gen prolaktin berdasarkan sekuenss gen prolaktin
Anas plathyrhyncos GenBank kode akses AB158611.1 6 4 Posisi penempelan primer pada fragmen gen prolaktin berdasarkan
Genbank kode akses AB158611.1 10
5 Hasil amplifikasi fragmen gen prolaktin ekson empat itik PMp 15 6 Hasil Alignment sekuens gen prolaktin itik PMp dengan Gen Bank
Anas plathyrhyncos (AB158611.1) 18
7 Region amplifikasi partial intron empat dan ekson empat gen prolaktin 18
DAFTAR LAMPIRAN
1 Sekuens gen prolaktin (PRL) Anas plathyrhyncosGen bank kode akses
AB158611.1 31
2 Chromatogram peak mutasi a) INDEL 3724A, b) T3941G, c) C3975A,
d) INDEL 4031A e) T4110A 34
3 Hasil BLAST nukleotida sekuens gen prolaktin itik PMp dengan Anas
plathyrhyncosGen Bank kode akses AB158611.1 35
4 Hasil BLAST nukleotida sekuens gen prolaktin itik Peking dengan
AnasplathyrhyncosGen Bank kode akses AB158611.1 35 5 Hasil BLAST nukleotida sekuens gen prolaktin itik Mojosari putih
6 Hasil Uji kenormalan data bobot pertama bertelur 36 7 Hasil Uji kenormalan data persentase produksi telur 37 8 Hasil Uji kenormalan data persentase fertilitas 37 9 Hasil Uji kenormalan data persentase produksi telur 38 10 Hasil Uji kenormalan data umur pertama bertelur menggunakan SPSS 38 11 Hasil analisis GLM dan LS mean bobot pertama bertelur terhada SNP
C3975A 39
12 Hasil analisis GLM dan LS mean umur pertama bertelur terhadap SNP C3975A
15 Hasil analisis GLM dan LS mean persentase daya tetas telur terhadap SNP C3975A
40 16 Hasil analisis GLM dan LS mean bobot pertama bertelur terhadap SNP
INDEL 3724A 40
17 Hasil analisis GLM dan LS mean umur pertama bertelur terhadap SNP INDEL3724A
20 Hasil analisis GLM dan LS mean persentase daya tetas telur terhadap SNP INDEL 3724A
41 21 Hasil analisis GLM dan LS mean bobot pertama bertelur terhadap SNP
T3941G 41
22 Hasil analisis GLM dan LS mean umur pertama bertelur terhadap SNP T3941G
42 23 Hasil analisis GLM dan LS mean persentase produksi telur terhadap
SNP T3941G
42 24 Hasil analisis GLM dan LS mean persentase daya tetas telur terhadap
SNP T3941G 42
25 Hasil analisis GLM dan Ls mean bobot pertama bertelur terhadap SNP INDEL 4031A
42 27 Hasil analisis GLM dan LSmean umur pertama bertelur terhadap SNP
INDEL 4031A 43
31 Hasil analisis GLM dan LS mean bobot pertama bertelur terhadap SNP T4110C
44 32 Hasil analisis GLM dan LS mean umur pertama bertelur terhadap SNP
T4110C 44
SNP T4110C 44 34 Hasil analisis GLM dan LSmean persentase fertilitas telur terhadap
SNP T4110C 44 35 Hasil analisis GLM dan LS mean persentase daya tetas terhadap
SNP T4110C
45 36 Data analisis transformasi LSMEAN ± SE INDEL 3724A terhadap
parameter reproduksi 45
37 Data analisis transformasi LSMEAN ± SE T3941G terhadap parameter reproduksi
45 38 Data analisis transformasi LSMEAN ± SE C3975A terhadap
parameter reproduksi
45 39 Data analisis transformasi LSMEAN ± SE INDEL 4031A terhadap
parameter reproduksi 46
40 Data analisis transformasi LSMEAN ± SE T4110C terhadap parameter reproduksi
46
1
PENDAHULUAN
Latar Belakang
Seiring dengan perkembangan diversifikasi produk hasil ternak yang sebelumnya bagi masyarakat hanya telur sebagai komoditas utama itik, saat ini masyarakat mulai menggemari daging itik sebagai salah satu sumber protein hewani. Akibatnya permintaan akan daging itik meningkat sepanjang tahun. Hal ini menyebabkan peningkatan permintaan terhadap bibit itik pedaging, namun peningkatan permintaan daging itik tidak diimbangi dengan peningkatan produksi dan ketersediaan bibit unggul.
Sebagai ternak yang berpotensi sebagai penghasil daging, produk daging itik di Indonesia belum begitu tinggi jika dibandingkan dengan negara lain di kawasan Asia. Berdasarkan data produksi daging itik yang dirilis oleh Badan Pangan Dunia (FAO) tahun 2010 menyebutkan bahwa total produksi daging itik di wilayah Asia mencapai 3 330 600 ton (dari delapan negara di Asia). China merupakan produsen daging itik terbesar dengan tingkat produksi 2 736 300 ton. Bila dibandingkan dengan China, Indonesia sangat jauh tertinggal. Indonesia hanya memproduksi 27 900 ton. Hal ini pun masih kalah dari negara asia lainnya yaitu Malaysia 116 300 ton, Bangladesh 41 500 ton, dan negara Asia lain seperti India, Myanmar, Thailand, dan Vietnam, secara berturut-turut memproduksi sebanyak 37 700 ton, 83 600 ton, 80 000 ton, dan 74 600 ton. Dari data tersebut maka Indonesia yang memiliki angka produksi daging itik terkecil (FAO 2012)
Upaya yang dilakukan dalam menghasilkan peningkatan performa itik lokal salah satunya adalah dengan menyilangkan antara itik Peking jantan dengan itik Mojosari putih betina. Persilangan ini dilakukan secara inseminasi buatan menghasilkan keturunan yang dinamakan itik Peking Mojosari putih (PMp). Keunggulan Itik PMp adalah warna bulu putih sehingga warna kulit karkas juga bersih dan cerah, bobot badan 2-2.5 kg pada umur 10 minggu, umur pertama bertelur 5.5-6 bulan dan rataan produksi telur pada umur 6 bulan sebesar 73-80 %. Prolaktin merupakan hormon polipeptida rantai tunggal yang termasuk kedalam family gen hormon pertumbuhan, umumnya disintesis dalam pituitari anterior seluruh hewan vertebrata. Pada kelas aves telah dilaporkan bahwa prolaktin banyak berperan penting dalam regulasi proses fisiologis diantaranya dalam hal produksi telur, memacu dan memelihara sifat mengeram, osmo-regulation, immune-modulation, fungsi dan perkembangan sel-sel gonad (Sharp et al. 1979).
Berdasarkan studi pustaka menunjukan bahwa polimorfisme gen prolaktin banyak ditemukan salah satunya pada sekuens gen prolaktin intron dan ekson empat, analisis asosiasi antara genotipe dengan sifat reproduksi memperlihatkan hasil yang signifikan. Penelitian yang dilakukan oleh Chang et al. (2012) melaporkan bahwa ditemukan dua titik mutasi yang terjadi pada gen prolaktin intron 4. Dua titik mutasi ini mempengaruhi sifat reproduksi pada itik.
Perumusan Masalah
Permintaan pasar akan itik pedaging meningkat sepanjang tahun menyebabkan peningkatan terhadap permintaan bibit itik pedaging. Usaha dalam memenuhi permintaan tersebut dilakukan dengan cara peningkatan mutu bibit itik pedaging dengan cara seleksi dan persilangan agar itik yang dihasilkan memiliki performa produksi dan reproduksi yang baik. Itik Peking Mojosari putih merupakan itik hasil silangan antara itik pejantan Peking dan itik induk Mojosari putih yang terbukti memiliki performa yang lebih baik dibanding performa tetuanya yaitu itik Mojosari putih.
Analisis keragaman gen prolaktin pada itik Peking, itik Mojosari putih dan persilangannya (PMp) dilakukan untuk mengetahui penanda genetik yang merupakan dasar untuk melakukan program seleksi. Selanjutnya dilakukan analisis asosiasi antara keragaman genetik dengan sifat reproduksi itik PMp yang terdiri atas bobot induk pertama bertelur, umur induk pertama bertelur, persentase produktivitas, fertilitas dan daya tetas telur.
Tujuan Penelitian
Tujuan penelitian ini adalah mengidentifikasi keragamangen prolaktin pada itik persilangan Peking Mojosari putih (PMp), Peking dan Mojosari putih serta menginvestigasi asosiasi antara keragamangen prolaktin dengan sifat reproduksi itik Peking Mojosari putih (PMp).
Manfaat Penelitian
Penelitin ini merupakan upaya untuk meningkatkan produktivitas ternak itik lokal adalah dengan melakukan seleksi di tingkat molekuler pada itik Peking Mojosari putih (PMp).
2
TINJAUAN PUSTAKA
Itik Lokal
merupakan bangsa itik Indian Runner yang sangat terkenal sebagai penghasil telur (Srigando 1997). Itik yang dipelihara saat ini disebut Anas domesticus. Itik ini berasal dari domestikasi itik liar (Anas moscha) atau Wild mallard (Suharno dan Setiawan 1999). Taksonomi itik dapat diklasifikasikan menurut Scanes et al. (2004) sebagai berikut:
Filum : Chordata Sub filum : Vertebrata Kelas : Aves Super ordo : Carinatae Ordo : Anseriformes
Spesies : Anas platryhynchos (Mallard dan domestik) Gambar 1 merupakan contoh dari berbagai jenis itik yang ada di Indonesia, penamaan yang diberikan pada itik-itik lokal tersebut berdasarkan asal dan morfologi itik-itik yang dibudidayakan pada suatu daerah. Menurut Setioko et al. (1997) beberapa jenis itik lokal diberi nama sesuai dengan lokasi dan memiliki morfologi yang khas. Sebagai contoh itik Tegal, Alabio, Bali, Cirebon, Tasikmalaya, Tangerang, Medan, Lombok dan Mojosari. Itik-itik tersebut sama halnya dengan itik yang lain diturunkan dari Mallard berkepala hijau (Anas Plathyrhyncos-plathyrhyncos) (Hasse dan Donham 1980; Hatzel 1985).
Akibat proses domestikasi maka terbentuklah beberapa variasi dalam besar tubuh, konformasi dan warna bulu. Perubahan ini juga diperkirakan sebagai akibat
a) b) c)
d) e) f)
campur tangan manusia untuk mengembangkan ternak itik dengan tujuan khusus, juga karena jauhnya jarak waktu domestikasi dengan waktu pengembangan (Chavez dan Lasmini 1978). Itik bersifat omnivorus (pemakan segala) yaitu memakan bahan dari tumbuhan dan hewan seperti biji-bijian, rumput-rumputan, ikan, bekicot dan keong. Itik merupakan unggas yang mempunyai ciri-ciri kaki relatif lebih pendek dibandingkan tubuhnya, jarinya mempunyai selaput renang, paruhnya ditutupi oleh selaput halus yang sensitif, bulu berbentuk cekung, tebal dan berminyak, itik memiliki lapisan lemak di bawah kulit, dagingnya tergolong gelap (dark meat), tulang dada itik datar seperti sampan (Suharno dan Setiawan 1999). Rasyaf (1993) menyatakan bahwa itik merupakan unggas air yang dipelihara untuk diambil telurnya yang mempunyai ciri-ciri umum tubuh ramping, berjalan horizontal, berdiri hampir tegak seperti botol dan lincah sebagai ciri unggas petelur. Itik merupakan hewan monogamus atau hidup berpasangan yang biasa diternakkan untuk diambil daging dan telurnya untuk dikonsumsi manusia. Itik lokal yang terdapat di Indonesia umumnya merupakan itik tipe petelur, mengalami masak kelamin pada umur 20-22 minggu dengan lama produksi sekitar 15 bulan (Hardjosworo dan Rukmiasih 1999).
Sistem pemeliharaan itik selama ini umumnya masih bersifat tradisional, yang lebih kita kenal dengan menggembalakan atau melepaskan di areal persawahan. Kondisi ini yang mengakibatkan produktivitas itik rendah karena kualitas pakan yang rendah dan sedikitnya pakan yang dikonsumsi dilapangan (Prasetyo 1997). Meskipun produktivitasnya rendah, namun bukan berarti potensi produksinya rendah. Potensi produksi dipengaruhi oleh genetik. Beberapa karakteristik genetik yang mempengaruhi produksi telur yaitu 1) masak kelamin dini (semakin cepat bertelur, produksi telurnya akan semakin tinggi dalam satu periode, namun telur yang dihasilkan lebih kecil), 2) intensitas bertelur yang tinggi (karakter ini dapat dilihat dari kemampuan peningkatan produksi yang cepat), 3) panjang masa bertelur (semakin lama bertelur sebelum ternak masuk periode rontok bulu produksi telurnya semakin tinggi), 4) lama istirahat, istirahat antar clutch lebih dari 2-3 hari akan mempengaruhi jumlah telur dalam masa produksi (North 1984).
Menurut Windhyarti (1999) itik dibagi menjadi tiga tipe yaitu tipe pedaging, tipe petelur dan tipe hias (ornamental). Itik tipe pedaging misalnya itik Peking dan itik Rouen. Itik ornamental contohnya itik Blue Swedis. Itik tipe petelur antara lain Indian Runner (Anas javanica) yang terdiri dari itik Karawang, itik Mojosari, itik Tegal, itik Magelang, itik Bali (Anas sp.), itik Alabio (Anas platurynchos borneo), itik khaki Campbell, itik CV 2000-INA serta itik unggul lain yang merupakan hasil persilangan oleh pakar PBT Ciawi-Bogor.
Itik Persilangan Peking Mojosari Putih
ekornya. Itik jantan pada umumnya memiliki satu atau dua lembar bulu yang melengkung ke atas. Selain itu warna paruh dan kaki pada itik jantan lebih hitam dibandingkan dengan itik betina. Itik Mojosari merupakan itik petelur unggul, sehingga itik Mojosari banyak dipelihara secara intensif. Produksi itik Mojosari adalah sekitar 238 butir/tahun (Prasetro et al. 1998). Menurut Suharno dan Amri (1996), rata-rata produksi telur itik Mojosari adalah 200 butir/tahun bila digembalakan. Dipelihara secara intensif dengan sistem kandang tanpa air rata- rata produksi telurnya meningkat sebesar 265 butir per ekor/tahun.
Itik Peking merupakan keturunan itik Mallard (Anas plathyrhynchos). Itik Peking memiliki penampilan yang seragam dengan bulu berwarna putih, paruh dan shank kuning. Pergerakan saat berjalan menyerupai entog, yaitu tubuh agak landai dengan bobot badan jantan berkisar antara 4.0-5.0 kg/ekor, sedangkan betina berkisar antara 2.5-3.0 kg/ekor (Setioko et al. 2004a).
Noor (1996) menyatakan bahwa apabila ternak yang tidak memiliki hubungan kekeluargaan disilangkan, maka keturunannya cenderung menampilkan performa yang lebih baik dari rata-rata performa tetuanya. Fenomena ini disebut hybrid vigor atau heterosis. Persilangan akan menggabungkan dua sifat atau lebih yang berbeda, yang semula terdapat dalam dua bangsa ternak ke dalam satu bangsa hasil silangan. Skema persilangan antara pejantan itik Peking dengan induk itik Mojosari putih tersaji pada Gambar 2.
Upaya yang dilakukan dalam menghasilkan peningkatan performa pada itik Mojosari yaitu dengan persilangan antara itik Peking pejantan dengan itik
♂
♀
Gambar 2 Skema perkawinan itik Peking pejantan dengan induk itik Mojosari putih betina menghasilkan silangan itik Peking Mojosari putih
Mojosari putih betina secara inseminasi buatan. Hasil silangan dari dua jenis itik ini menghasilkan keturunan yang dinamakan itik Peking Mojosari Putih (PMp) yang memiliki performa lebih unggul dari tetuanya karena memiliki dua keunggulan yang berasal dari kedua tetuanya yaitu memiliki produksi telur yang tinggi disertai bobot badan yang besar pula.
Karakteristik produksi itik PMp diantaranya warna bulu putih, sehingga warna kulit karkas juga bersih dan cerah, bobot badan 2-2.5 kg pada umur 10 minggu, jika dikawinkan dengan entog jantan dapat digunakan untuk menghasilkan itik serati dengan bobot badan 3 kg atau lebih pada umur 10 minggu, umur pertama bertelur 5.5-6 bulan, rataan produksi telur 6 bulan 73-78 %. Hasil silangan dari dua galur ini menghasilkan zuriat dengan pola warna bulu yang seragam yaitu putih polos. Mengingat preferensi konsumen senantiasa menggunakan acuan karkas ayam broiler yang memiliki sifat karkas yang putih terkesan bersih, Triyantini et al. (1997) melaporkan bahwa preferensi penampakan warna karkas entog dengan ayam buras adalah sama-sama disukai konsumen. Sementara itu, untuk karkas ayam kampung dan itik cenderung kurang disukai.
Gen Prolaktin
Struktur gen prolaktin pada itik telah berhasil dikarakterisasi oleh (Kansaku et al. 2005). Gen prolaktin memiliki ukuran 6 332 pb yang terdiri dari lima ekson dan empat intron, panjang fragment 5’ flanking region sebesar 242 pb dan mengkode 229 asam amino, adapun rekonstruksi struktur gen prolaktin dapat dilihat pada Gambar 3. Prolaktin itik memiliki kesamaan berturut-turut sebesar 92.0%, 91.7%, dan 91.4% pada ayam, kalkun dan puyuh di level cDNA. Prolaktin itik dewasa secara keseluruhan memiliki kesamaan dengan ayam (95.5%), kalkun (92.5%) dan puyuh (95.5%) (Kansaku et al. 2005).
Rangkaian ekson dan intron gen prolaktin unggas pada umumnya memiliki panjang dari ATG awal dimulainya translasi codon menuju akhir translasi kodon sepanjang 6 kb. Panjang sekuens 5’flanking pada ayam, kalkun
dan itik berturut-turut sebesar 2.6 kb, 2.0kb, dan 0,2kb. Pada manusia prolaktin ditemukan pada kromosom 6 (Goffin et al. 1996), pada tikus terletak di kromosom 13 (Wiemers et al. 2003), pada babi terletak di kromosom 7 (Vincent et al. 1998), dan pada unggas terletak di kromosom 2 (Alipanah et al. 2011). Liu et al. (2009) menyatakan bahwa produk ekspresi gen prolaktin adalah hormon prolaktin, yang tersusun dari 199 asam amino, dengan tiga ikatan disulfida diantara enam cystein, dan memiliki bobot molekul sebesar 23kDa.
Telah dilaporkan bahwa aksi biologis dari prolaktin dijembatani oleh prolaktin reseptor (PRLR) dimana secara struktural memiliki kesamaan yang tinggi dengan growth hormon receptor (GHR) (Kitamura et al. 1994; Bole-Feysot et al. 1998). Aktivasi prolaktin reseptor (PRLR) terjadi karena ikatan prolaktin yang dapat menginisiasi sinyal intraseluler, termasuk aktivasi intraseluler JAK2-STAT5 yang selanjutnya meniginisiasi terjadinya proses transkripsi gen (Bole-Feysot et al. 1998).
Berbeda dengan mamalia yang pengaturan dan pelepasan gen prolaktin di bawah mengaturan inhibitory control, pada unggas pengaturan dan pelepasan prolaktin dibawah pengaturan tonic regulation dan faktor pelepas utama yang disebut dengan vasoactive intestinal polypeptide (VIP) (Kansaku et al. 2008). Hal yang sama juga dikatakan oleh (Kagya et al. 2012) bahwa sekresi prolaktin pada unggas sebagian besar diatur oleh releasing factor yang terdiri vasoactive intestinal peptide (VIP), dopamine (DA) dan serotonine (5-HT). Vasoactive intestinal peptide (VIP) disekresikan oleh hypothalamus, VIP menstimulasi pelepasan dan ekspresi prolaktin baik secara in vitro maupun in vivo pada unggas.
Penelitian yang dilakukan pada ayam menunjukan bahwa gen prolaktin terekspesikan di hypothalamus, pituitary gland, oviduct dan ovarium. Ekspresi gen prolaktin tertinggi terdapat pada pituitary gland (Li et al. 2009). Sama halnya dengan penelitian yang dilakukan pada angsa (Chu et al. 2008) yang mengindikasikan bahwa level prolaktin tertinggi terdapat pada pituitary gland, selanjutnya diikuti oleh hypothalamus, dan yanag terakhir pada ovarium.
Aktivitas hormon prolaktin pada hewan memiliki dampak yang luas. Prolaktin memiliki dampak penting terhadap proses reproduksi pada ikan, unggas dan mamalia (Wang et al 2009). Pada reptil dan amfibi prolaktin berperan sebagai osmoregulator, pada ikan prolaktin berdampak terhadap fungsi organ (insang, usus, ginjal, dan kulit) yang memudahkan ikan beradaptasi pada kondisi lingkungan. Pada mamalia prolaktin menstimulasi pembentukan corpus luteum dan berperan dalam pengaturan mammogenesis, lactogenesis, dan lactopoiesis, menstimulasi hormon progesteron yang merupakan hormon utama dalam pengaturan kebuntingan. Pada unggas prolaktin merupakan hormon penting yang mengatur dan menginisiasi tingkah laku mengeram dan regulasi pembentukan folikel (Reddy et al. 2002).
Penanda Molekuler Gen Prolaktin
DNA, dan diperkirakan bahwa satu SNP terjadi setiap 250-1000 pb. Frekuensi mutasi dan stabilitas yang tinggi menyebabkan SNP sering digunakan sebagai penanda molekuler dalam penelitian genetika populasi dan pemetaan gen untuk penyakit kompleks (Ye et al. 2001) metode yang umum digunakan dalam menganalisis SNP antara lain PCR-RFLP dan PCR-SSCP.
Polimerase chain reaction (PCR) merupakan suatu teknik untuk menggandakan jumlah molekul DNA pada ruas-ruas tertentu dan monomer-monomer nukleotida yang dilakukan secara in vitro. Proses ini berjalan dengan bantuan primer dan enzim polymerase. Primer merupakan oligonukleotida spesifik yang menempel pada bagian sampel DNA yang akan diperbanyak. Enzim polymerase merupakan enzim yang dapat mencetak urutan DNA baru. Hasil dari proses PCR dapat divisualisasikan dengan elektroforesis (Williams 2005).
Ekstraksi DNA yang umum dilakukan adalah dengan mengekstraknya dari darah karena dalam darah terdapat sel darah putih dan sel-sel darah merah yang masih muda. Sampel darah banyak digunakan dalam ekstraksi DNA karena mudah diperoleh serta prosedur isolasi yang relatif mudah.Reaksi yang terjadi dalam mesin PCR secara umum dapat dibagi menjadi tiga tahap yaitu tahap denaturasi DNA cetakan, tahap annealing atau penempelan primer dan tahap extension yaitu pemanjangan primer atau polymerase. Reaksi ini umumnya terjadi dalam 25-35 siklus. Pada tahap denaturasi, DNA dipanaskan hingga 94 oC
sehingga DNA untai ganda berpisah menjadi DNA untai tunggal. Tahapan yang paling menentukan dalam proses PCR adalah tahap penempelan primer, karena tiap pasangan primer memiliki suhu penempelan primer yang spesifik. Tahap pemanjangan primer terjadi pada suhu 72 oC. Pada tahapan ini enzim taq polymerase, buffer PCR, dNTP, dan Mg2+ memulai aktifitasnya memperpanjang primer (Viljoen et al. 2005).
Hubungan Keragaman Gen Prolaktin dengan Sifat Fenotipe
Semenjak Watahiki et al. (1989) berhasil mengkloning dan mensekuenssing gen prolaktin pada ayam, banyak penelitian yang berfokus pada deteksi keragaman gen prolaktin. Saat ini, sejumlah SNP gen prolaktin telah dilaporkan. Cui et al. (2006) menemukan enam SNP diantaranya C2402T, C2161G, T2101G, C2062G, T2054A, G2040A dan 24-pb indel (insertion-deletion) dari direct sequencing dan analisis asosianya menunjukan bahwa 24-pb indel berhubungan dengan produksi telur dan sifat mengeram pada ayam. Liu et al. (2007) menganaliasa tiga mutasi C1607T, C5749T dan T5821C yang menunjukan adanya korelasi antara perbedaan kombinasi haplotipe dengan produksi telur. Dari semua penelitian diatas menunjukan bahwa prolaktin merupakan kandidat gen penting dalam sifat produksi telur.
terhadap parameter jumlah telur dan bobot badan saat pertama bertelur. Berbagai hasil penelitian yang berhasil menganalisis asosiasi antara polimorfisme gen prolaktin dengan sifat reproduksi yang telah terpublikasi, tersaji pada Tabel 1 dibawah ini.
Tabel 1 Asosiasi polimorfisme gen prolaktin terhadap sifat reproduksi unggas
Spesies Lokasi SNP Asosiasi terhadap Sifat Referensi
Ayam In/Del 359 pb Produksi telur Cui et al. 2006
Promotor Sifat mengeram Jiang et al. 2005
Ekson 2 Bobot badan pertama bertelur,
Umur dewasa kelamin
Rashidi et al. 2012
Angsa 5’ untraslated region Produksi telur Chen et al. 2011
5’ flanking region Produksi telur Jiang et al. 2009
Itik Intron 1 Kerabang telur Wang et al. 2011
Ekson 5 Produksi telur, bobot telur
Intron 1 Bobot telur Liu et al. 2012
3
METODE
Lokasi dan Waktu
Penelitian ini dilaksanan di Laboratorium Genetika Molekuler Ternak IPB, dan Balai Penelitian Ternak Ciawi-Bogor pada bulan April 2013-Februari 2014.
Materi Itik
Itik yang digunakan dalam penelitian ini berasal dari kandang penelitian itik Balai Penelitian Ternak Ciawi-Bogor yang terdiri dari itik silangan PMp sebanyak 113 ekor yang terdiri 15 ekor pejantan dan 98 ekor induk betina, 19 ekor itik Peking yang terdiri dari pejantan 4 ekor dan betina 15 ekor, dan 36 itik Mojosari putih yang terdiri dari 12 pejantan dan 24 ekor betina. Itik pejantan Peking dan induk Mojosari putih merupakan tetua dari itik Peking Mojosari putih. Semua itik dipelihara pada kadang individu dalam manajemen perkandangan dan pakan yang sama. Pakan yang diberikan sebanyak 200-230 g/hari dengan kandungan nutrisi yang tersaji pada Tabel 2 sebagai berikut:
Tabel 2 Kandungan nutrisi pakan komplit komersial untuk sampel itik penelitian
Sampel Darah
Sampel darah yang digunakan dalam penelitian adalah darah yang berasal dari itik PMp, itik Peking dan itik Mojosari putih yang dipelihara di kandang penelitian itik Balai Penelitian Ternak Ciawi-Bogor. Sampel darah diambil sebanyak 1 ml dan dikoleksi didalam tube ependorf kapasitas 1.5 ml yang didalamnya dimasukan EDTA (ethylene diamine tetraacetic acid) sebagai koagulan. Sampel sebaiknya diproses pada hari yang sama saat pengambilan untuk isolasi DNA simpan pada suhu 4 oC.
Primer Gen Prolaktin
Primer ekson 4 yang digunakan terdiri dari forward 5- CTG CAT CTG TGG ACA TTG CT -3 dan reverse 5- GAA GCA GGT TTG GGA GTA CG -3. Penyusunan desain primer berdasarkan fragmen gen prolaktin yang sudah terpublikasi (GenBank accession no. AB158611.1). Posisi penempelan primer pada amplifikasi DNA dapat dilihat pada Gambar 4.
Gambar 4 Posisi penempelan primer pada fragmen gen prolaktin berdasarkan Genbank kode akses AB158611.1
Prosedur
Persiapan Ternak dan Teknik Pengambilan Data
Parameter yang diamati pada penelitian kali ini adalah sifat reproduksi yang terdiri dari umur pertama induk bertelur, bobot badan induk pertama bertelur, persentarse produksi ,persentase fertilitas, dan daya tetas telur. Kesemua sifat tersebut akan dianalisis menggunakan polimorfisme–trait association analysis. Nilai fenotipe dari kelima sifat tersebut diamati pada setiap itik betina yang sebelumnya telah dilakukan inseminasi buatan menggunakan polled semen yang berasal dari 15 itik PMp pejantan. Inseminasi buatan dilakukan dengan mengikuti program Balai Penelitian Ternak yaitu dua kali seminggu. Setelah IB dilakukan pengamatan secara individu berupa jumlah telur yang diinkubasi, jumlah telur fertil, jumlah telur fertil saat candling dan jumlah itik hidup setelah menetas. Nilai fertilitas merupakan rasio hasil perhitungan antara jumlah telur fertil saat candling dibagi jumlah telur inkubasi dikali 100. Daya tetas telur dihitung dengan cara membagi jumlah telur yang menetas dibagi dengan jumlah telur fertil.
Uji Kualitatif dan Kuantitatif DNA
murni dapat menyerap cahaya ultraviolet karena keberadaan basa-basa purin dan pirimidin. Pita ganda DNA dapat menyerap cahaya UV pada panjang gelombang 260 nm, sedangkan kontaminan protein atau fenol akan menyerap cahaya pada panjang gelombang 280 nm. Sehingga kemurnian DNA dapat dukur dengan menghitung nilai absorbansi λ 260 nm dibagi dengan nilai absorbansi λ 280 (Å260/Å280), dan nilai kemurnian DNA berkisar antara 1.8-2.0. Serta untuk mengukur konsentrasi DNA digunakan rumus sebagai berikut:
[DNA] = Å260 x 50 x faktor pengenceran Keterangan :
Å 260 : Nilai absorbansi pada λ 260nm
50 : Larutan dengan nilai absorbansi 1.0 sebanding dengan 50 ug untai ganda DNA per mL
Ekstraksi DNA dalam Darah
Ekstrak DNA total berasal dari sampel darah. Metoda ekstraksi DNA dari darah segar dilakukan mengunakan Genomic DNA Mini Kit (GeneAidTM DNA Isolation Kit). Kemurnian DNA dianalisis dua kali yaitu pertama menggunakan alat spektrofotometer dan kedua berdasarkan hasil visualisasi dari elektroforesis.
Metode PCR
Amplifikaasi fragmen DNA akan dilakukan dengan mesin PCR (Polymerase Chain Reaction). Kondisi PCR sangat bergantung pada primer yang digunakan. Pada penelitian ini akan menggunakan primer prolaktin ekson 4 berdasarkan sekuens itik refference Anas platyrhynchos Genbank nomor akses: AB158611.
Larutan cocktail PCR dibuat sebanyak 35 µl yang terdiri dari 0.7 µl larutan primer ekson 4, 1 µl DNA, 17.5 µl KAPA 2G master mix dan 16.8 µl PCR grade water. KAPA 2G master mix merupakan campuran taq DNA polymerase, PCR buffer, MgCl2 dan dNTPs yang siap digunakan.
Amplifikasi DNA berlangsung di dalam mesin PCR Applied Biosystems dengan kondisi suhu predenaturasi 95oC selama lima menit, denaturasi 95oC selama 10 detik, annealing pada suhu 60oC selama 20 detik, dan elongasi pada suhu 72oC selama 30 detik dan elongasi akhir pada suhu 72oC selama lima menit
dilakukan sebanyak 35 siklus. Setelah siklus berakhir diteruskan dengan temperatur penyimpanan 4oC. Hasil amplifikasi PCR dicek dengan elektroforesis
gel agarose 1.5%
Metode Elektroforesis dan Analisis Keragaman DNA
Langkah berikutnya adalah memasukan sampel sebanyak 2µl ke dalam sumur-sumur dalam cetakan dan proses elektroforesis dimulai pada tegangan 100 volt selama 45 menit. Hasil elektroforesis dapat dilihat dalam mesin Gel Doc dan disimpan dalam file, setelah diketahui ukurannya dalam satuan pasang basa (pb).
Masing-masing pola pita yang terbentuk dilihat dibawah sinar dan sampel tersebut akan memperlihatkan pita-pita yang berbeda dalam gel yang sebelumnya telah mengalami amplifikasi dan purifikasi. Selanjutnya dilakukan sequencing menggunakan BIO TRACE 3730 dari perusahaan First Base, Malaysia.
Teknik Pengumpulan Sperma dan Inseminasi Buatan
Teknik perangsangan dan pengumpulan sperma dilakukan dengan menggunakan teknik pengurutan. Cara melakukan rangsangan adalah salah satu tangan peternak memegang pejantan, itik diapit antara tangan dan pinggul. Satu tangan lagi (bagian kanan) lakukan perangsangan dibagian abdomen itik PMp jantan hingga phallaus keluar dan semen siap untuk ditampung. Setelah pejantan terangsang maka terjadi proses ejakulasi yaitu memanjangnya penis dan keluarnya sperma. Pada saat ini peternak siap untuk menyedot dengan alat sedot yang akan menampung sperma dalam tabung gelas. Pengenceran sperma dilakukan dengan cairan garam fisiologis 80% (cairan infus) dengan perbandingan 1:1 atau jika sperma kental dapat 2 cc pengencer : 1 cc sperma (Prasetyo et al. 2010). Semen itik PMp jantan ditampung pagi hari (08.00) dengan maksud agar kondisi lingkungan dingin dan sekit terkena cahaya matahari.
Inseminasi dilakukan kedalam vagina itik PMp betina dengan dosis 108
spermatozoa per straw. Inseminasi dilakukan dengan menggunakan insemination gun yang sudah berisi straw. Masukan insemination gun kedalam vagina itik betina dengan dalaman ±3cm. Itik yang telah diinseminasi dikembalikan ke kandang individu.
Pengukuran Parameter Sifat Reproduksi
Parameter sifat reproduksi yang diamati pada penelitian ini terdiri dari bobot induk pertama bertelur, umur induk pertama kali bertelur, persentase produksi telur, persentase fertilitas telur dan persentase daya tetas telur. Bobot badan pertama bertelur dapat digunakan sebagai salah satu indikator itik telah siap bertelur atau belum. Bobot badan induk ditimbang saat pertama kali bertelur hal ini merupakan salah satu parameter data yang mempengaruhi sifat reproduksi itik.
Umur bertelur yang pertama merupakan tanda yang mudah dikenali bahwa unggas sudah masak kelamin, walaupun sebelumnya sudah terjadi ovulasi. Itik Mojosari mulai bertelur pada umur 145 hari. Umur induk saat pertama kali bertelur dihitung dengan cara mengurangi tanggal pertama induk bertelur dengan tanggal induk menetas.
Informasi tentang kemampuan produksi itik lokal diperlukan untuk peningkatan produktivitasnya. Itik Mojosari pada bulan pertama produksi telurnya mencapai 40 % dan akan mencapai 80% sebagai produksi puncak pada bulan ke empat. Persentase produksi telur dihitung dengan menjumlahkan seluruh telur yang dihasilkan per individu itik dibagi jumlah hari pengamatan dikali 100 persen. Jumlah hari pengamatan produksi telur selama 31 hari.
sejumlah telur yang dieramkan. Angka fertilitas diperoleh dari jumlah telur yang fertil dibagi dengan jumlah telur yang dieramkan dikali 100%.
Kemampuan embrio untuk tetap bertahan sampai menetas disebut dengan daya tetas. Daya tetas telur dipengaruhi faktor genetik, yaitu karena kontribusi gen yang diwariskan. Selain itu, kondisi lingkungan dalam mesin penetasan juga mempengaruhi daya tetas telur. Daya tetas (hatchability) adalah persentase jumlah telur yang menetas terhadap jumlah telur fertil secara keseluruhan dikali 100%.
Prosedur Analisis Data
Analisi Homologi
Analisis homologi antara sekuens DNA itik sampel, itik referensi Anas platyrhynchos dan jenis unggas lain menggunakan metode Basic Local Allignment Search Tool (BLAST) (www.ncbi.nlm.gov.nih). Persentasi homologi memperlihatkan kesamaan komposisi basa nukleotida pada masing-masng sampel.
Analisis Komposisi Basa Nukleotida
Analisis komposisi basa nukleotida masing-masing sampel menggunakan BioEdit Sequence Alignment Editor (Hall 1999) dan Moleculer Evolutionary Genetics Analysis (MEGA4).
Identifikasi Polimorfisme
SNP (Single nucleotide polymorphisms) yang diidentifikasi pada penelitian ini didapat dengan cara membandingkan alignment masing-masing sekuens prolaktin sampel itik dengan sekuens Anas plathyrhynchos (GenBank: AB15861.1; NCBI) menggunakan Moleculer Evolutionary Genetics Analysis (MEGA4) (Tamura et al. 2007) dan hasil kromatogram masing-masing itik menggunakan BioEdit Sequence Alignment Editor (Hall 1999).
Frekuensi Alel Gen Prolaktin
Frekuensi alel adalah rasio suatu alel terhadap keseluruhan alel pada suatu lokus dalam populasi. Frekuensi alel (χi) gen prolaktin yang dihitung berdasarkan Nei
dan Kumar (2000) sebagai berikut:
Keterangan :
χii : frekuensi alel ke-i
nii : jumlah individu bergenotipe ii
nij : jumlah individu bergenotipe ij
Prosedur Analisis Statistik
Uji frekuensi lima SNP sekuens gen prolaktin yang teramati dengan frekuensi harapan dihitung menggunakan uji Chi Square (χ2) menurut Walpole (1993) dengan rumus:
Keterangan :
X 2 : Sebaran Chi Square
oi : Frekuensi teramati
ei : Frekuensi harapan ke-i, dengan uji hipotesis jika :
Ho : χ2 hitung < χ2 tabel, tidak terdapat hubungan yang signifikan antara
SNP terhadap jenis itik yang berbeda
Ha : χ2 hitung < χ2 tabel, terdapat hubungan yang signifikan antara SNP
terhadapt jenis itik yang berbeda
Data fenotifik reproduksi dihitung secara statistik menggunakan prosedur SPSS. Data yang diperoleh dari kegiatan molekuler di laboratorium dengan data pengamatan secara fenotifik reproduksi selanjutnya dianalisis dengan uji statistik menggunakan prosedur GLM persamaannya sebagai berikut :
Yijk = µ + Gi +Hj + eijk
Keterangan:
Yijk : Sifat reproduksi yang diamati
µ : rataan seluruh populasi Gi : Efek genotipe
Hj : Efek tetap
eijk : Galat eror
4
HASIL DAN PEMBAHASAN
Ekstraksi dan Amplifikasi Gen Prolaktin
Kualitas dan kuantitas DNA pada suatu organisme dapat diketahui dengan menggunakan alat spektrofotometer. Prinsip kerja dari spektrofotometer adalah iradiasi sinar ultraviolet yang diserap oleh nukleotida dan protein dalam larutan. Iradiasi sinar ultraviolet oleh nukleotida secara maksimal dicapai pada panjang gelombang 260 nm, sedangkan iradiasi sinar ultraviolet maksimal oleh protein dicapai pada panjang gelombang 280 nm (Sulandari dan Zein 2003).
jika nilai rasio A260/A280 adalah sebesar 1.8-2.0 (Sulandari dan Zein 2003), maka setelah dibandingkan dengan dengan literatur maka dapat disimpulkan bahwa DNA hasil ekstraksi pada penelitian ini adalah murni.
Sangat penting untuk mengetahui berapa banyak konsentrasi DNA yang digunakan untuk perunutan (sequencing) agar keberhasilan sekuenssing terjamin dan tereproduksi. Hasil perhitungan untuk nilai konsentrasi yang didapat pada penelitian ini rata-rata sebesar 263,34 ng/µl. Konsentrasi template DNA genom yang akan digunakan untuk proses perunutan tergantung pada sumber DNA, syarat konsentrasi DNA genom yang digunakan untuk penelitian sebesar 100-250 ng/µl (www.embl.de). Berdasarkan literatur disimpulkan bahwa DNA yang berhasil diekstraksi secara kualitas maupun kuantitas memenuhi syarat untuk dapat dilakukan prosedur penelitian selanjutnya.
Amplifikasi gen prolaktin pada ekson 4 dilakukan menggunakan mesin termal cycler AB system pada suhu annealing 60 0C selama 20 detik. Hasil PCR divisualisasikan pada media agar agarose 1.5 % seperti pada Gambar 5 dibawah ini.
Gambar 5 Hasil amplifikasi gen prolaktin ekson 4 pada gel agarose 1.5%. Keterangan: M = marker DNA. Sampel 1-16 = produk amplifikasi gen prolaktin
Hasil visualisasi menunjukan bahwa panjang produk hasil amplifikasi ruas gen prolaktin ekson 4 dan sebagian intron 3 dan 4 sebesar 496 pb. Hasil ini sesuai dengan target region hasil penempelan primer yang teramplifikasi di posisi 3661-4157 pb. Identifikasi genotipe gen prolaktin pada itik PMp, Peking dan Mojosari putih dilakukan melalui identifikasi penampakan pita dibandingkan dengan marker yang telah diketahui panjang runutan pasang basanya.
Karakteristik Gen Prolaktin
Homologi Gen Prolaktin
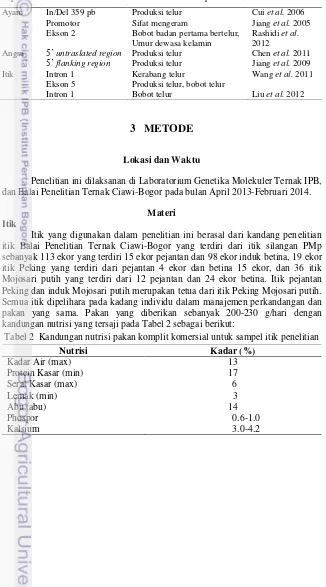
Tabel 3 Analisis homologi itik PMp, Peking dan Mojosari putih dengan metode BLAST
Sampel Kode Akses PMp Peking Homologi (%) Mojosari putih
Itik (Anas platyrhynchos) AB15811.1 99 99 99
Angsa (Anser anser) DQ660983.1 95 95 95
Merak (Pavo cristatus) AB605393.1 87 87 87 Kalkun (Meleagris gallopavo) AB605394.1 83 83 83
Hasil analisis menunjukkan bahwa fragmen gen prolaktin itik PMp, Peking dan Mojosari putih yang diteliti memiliki nilai homologi 99% dengan itik Anas platyrhynchos kode akses Genbank AB15811, 95% pada Angsa (Anser anser) DQ660983.1, 87% pada Merak (Pavo cristatus) AB605393.1 dan 83% pada Kalkun (Meleagris gallopavo) AB605394.1. Kansaku et al. (2005) yang menyebutkan bahwa gen prolaktin itik memiliki kesamaan berturut-turut 92.0%, 91.7% dan 91.4% pada ayam, kalkun dan burung puyuh.
Analisis homologi bertujuan untuk mengetahui tingkat kesamaan fragmen gen prolaktin itik-itik penelitian dengan sekuens fragmen gen itik yang sudah terpublikasi mengacu pada Genbank. Tingginya tingkat homologi sekuens fragmen gen prolaktin menunjukan bahwa DNA itik penelitian yang diuji adalah benar merupakan fragmen gen prolaktin.
Komposisi Basa Nukleotida Gen Prolaktin
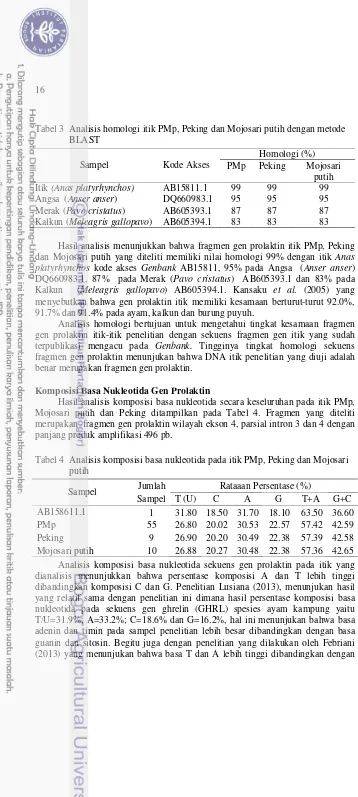
Hasil analisis komposisi basa nukleotida secara keseluruhan pada itik PMp, Mojosari putih dan Peking ditampilkan pada Tabel 4. Fragmen yang diteliti merupakan fragmen gen prolaktin wilayah ekson 4, parsial intron 3 dan 4 dengan panjang produk amplifikasi 496 pb.
Tabel 4 Analisis komposisi basa nukleotida pada itik PMp, Peking dan Mojosari putih
Sampel Jumlah Rataaan Persentase (%)
Sampel T (U) C A G T+A G+C
AB158611.1 1 31.80 18.50 31.70 18.10 63.50 36.60
PMp 55 26.80 20.02 30.53 22.57 57.42 42.59
basa G dan C yaitu analisis komposisi basa nukleotida gen prolaktin pada spesies ayam arab yaitu T/U=32.7%; A=29.1%; C=20.4% dan G=17.8%.
Sebagaimana diketahui bahwa menurut (Muladno 2010) pasangan A dan T akan lebih cepat terpisah menjadi untai tunggal dibandingkan pasangan basa G dan C karena pasangan A dan T memiliki dua ikatan hidrogen sedangkan basa G dan C memiliki tiga ikatan hidrogen. Hal ini mengindikasikan bahwa pada wilayah ekson memiliki komposisi A dan T lebih besar dibanding G dan C karena wilayah ini akan bertranskripsi menjadi untaian tunggal.
Deteksi Mutasi Gen Prolaktin
Mutasi adalah perubahan materi genetik yang bersifat dapat diwariskan. Kesalahan apapun yang terjadi selama replikasi gen didalam molekul DNA pada satu atau lebih basa dapat menyebabkan timbulnya mutasi. Meskipun sel mempunyai suatu mekanisme untuk meningkatkan ketepatan replikasi DNA, terkadang bisa terjadi suatu kesalahan spontan yang menimbulkan perubahan pada DNA yang dapat diwariskan (Stansfield et al. 2003).
Jenis mutasi gen dapat berupa missense mutation, silent mutation, nonsense mutation dan frameshift mutation. Missense mutation yaitu perubahan suatu kode genetik sehingga menyebabkan asam amino terkait (pada polipeptida) berubah. Perubahan pada asam amino dapat menghasilkan fenotipe mutan apabila asam amino yang berubah merupakan asam amino esensial bagi protein tersebut. Jenis mutasi ini dapat disebabkan oleh peristiwa mutasi transisi dan tranversi. Silent mutation, yaitu perubahan suatu pasangan basa dalam gen (pada posisi 3 kodon) yang menimbulkan perubahan satu kode genetik tetapi tidak mengakibatkan perubahan asam amino yang dikode biasanya disebabkan karena terjadinya mutasi transisi dan tranversi. Nonsense mutation, yaitu perubahan kodon asam amino tertentu menjadi kodon stop. Hampir semua mutasi tanpa arti mengarah pada inaktifnya suatu protein sehingga menghasilkan fenotipe mutan. Frameshift mutation, yaitu mutasi yang terjadi karena delesi atau insersi satu atau lebih pasang basa dalam satu gen sehingga ribosom membaca kodon tidak lengkap. Akibatnya akan menghasilkan fenotipe mutan (Stansfield et al. 2003). Hasil analisis alignment pada sekuens prolaktin ekson 4 dan parsial intron 3 dan 4 menunjukan ditemukannya beberapa titik mutasi. Deteksi kejadian mutasi pada ekson 4 dan sebagian intron 3 dan 4 ditampilkan pada Tabel 5.
b = tidak ada basa adenin di 3939 pb pada itik reference AB158611;
c = tidak terjadi mutasi T>C di itik Peking
Berdasarkan hasil analisis pensejajaran (alignment) ekson 4 dan parsial intron 3 dan 4 menunjukkan ditemukannya enam titik mutasi yag dijadikan sebagai SNP diantaranya adalah INDEL 3724A, INDEL 3939A, T3941G, C3975A, INDEL 4031A dan T4110C. Berbagai titik mutasi yang terjadi pada fragmen gen prolaktin ekson 4 dan parsial intron 3 dan 4 dapat dilihat dari hasil analisis alignment yang tersaji pada Gambar 6.
Gambar 6 Hasil analisis alignment nukleotida sekuens gen prolaktin itik sampel dengan itik Anas plathyrhyncosGenbank (AB158611)
Yunita (2009) menyatakan bahwa mutasi tipe delesi terjadi karena pasangan basa tertentu menghilang sehingga terjadi perubahan susunan basa nukleotida. Dari hasil analisis pensejajaran menunjukan bahwa pada posisi 3724 pb dan 4031 pb terjadi kehilangan satu basa adenin sehingga pada posisi ini ditemukan mutasi delesi yang dinyatakan sebagai INDEL 37124A dan INDEL 4031A. Mutasi tipe insersi terjadi karena adanya penyisipan basa tertentu (Camphel et al. 2002). Pada posisi 3939pb terjadi penyisipan satu basa adenin sehingga pada posisi ini ditemukan mutasi yang dinyatakan sebagai INDEL 3939A.
Mutasi tipe tranversi terjadi karena adanya perubahan antara basa purin menjadi pirimidin. Mutasi transisi terjadi karena adanya perubahan antara basa purin (adenin dan guanin) dengan basa purin lainnya atau antara basa pirimidin (timin dan sitosin) dengan pirimidin lainnya (Nei 1987). Pada penelitian ini ditemukan dua titik mutasi jenis transversi, pada posisi 3941pb terjadi perubahan basa pirimidin (timin) menjadi purin (guanin) dinotasikan sebagai T3941G dan di posisi 3975 pb terjadi perubahan basa pirimidin (sitosin) menjadi purin (adenin) yang dinotasikan sebagai C3975A. Sedangkan mutasi transisi ditemukan pada posisi 4110 pb dimana terjadi perubahan basa pirimidin (timin) menjadi pirimidin (sitosin) dan dinotasikan sebagai T4110C. Region gen prolaktin ekson 4 dan parsial intron 3 dan 4 yang teramplifikasi dengan panjang sekuens 496 pb diilustrasikan pada Gambar 7 dibawah ini.
Sebagaimana yang terlihat pada Gambar 7 bahwa diketahui region ekson 4 berada di posisi 3753–3932 pb, sehingga analisis yang dilakukan menunjukan bahwa seluruh mutasi pada penelitian ini terjadi di wilayah intron yaitu wilayah parsial intron 3 dan intron 4. Intron merupakan wilayah sekuens DNA yang tidak bertranskripsi menjadi protein namun demikian kini telah banyak ditemukan hasil penelitian yang menunjukan fungsi intron khususnya pada sel eukariot. Salah satu contoh pentingnya fungsi intron adalah dalam sel eukariot adalah meningkatkan kelimpahan protein. Efek intron pertama kali diamati pada virus simian vacuolating40 yang menghasilkan produk protein yang tidak terdeteksi saat penghilangan sekuens intronnya (Gruss et al.1979; Hamer et al.1979). Penelitian lain menggunakan susunan virus yang sama menunjukan penghilangan intron berdampak pada level RNA. Dalam banyak kasus, mekanisme intron belum diketahui, tetapi pada beberapa kasus menunjukan intron memiliki dampak terhadap proses pematangan mRNA, termasuk inisiasi proses transkripsi, elongasi transkripsi, terminasi transkripsi, poliadenilasi, eksport inti sel dan penstabil RNA (Chorev dan Carmel 2012).
SNP yang berada di region non-coding memiliki dampak terhadap ekspresi gen dengan cara mempengaruhi pengaturan komponen dan beberapa SNP intronik aktif cryptic splice sites yang memnyebabkan reaksi splicing (Goto et al. 2001; Alberobello et al. 2011). Intronik SNP pada gen A-FABP yang ditemukan pada babi menunjukan adanya asosiasi antara level A-FABP dengan perlemakan otot (marbling) yang ditandai dengan berkurangnya genotipe DD dibandingkan dengan genotipe CC (Gao et al. 2011).
Mutasi INDEL 3724A merupakan mutasi yang terjadi pada intron 3. Berdasarkan studi literatur yang telah dilakukan belum ada laporan penelitian yang menunjukan adalanya mutasi pada region intron 3 maupun ekson 3 pada itik. Penelitian yang dilakukan Das et al. (2012) mengkarakterisasi gen prolaktin ekson 3 dan 4 pada sapi. SNP yang ditemukan pada ekson 3 berhubungan erat dengan sifat periode laktasi tinggi, sedangkan SNP pada ekson 4 berhubungan dengan produksi susu yang tinggi.
SNP indel 3939A, T3941G, C3975A indel 4031A dan T4110C ditemukan pada region intron 4. Penelitian yang dilaporkan oleh Chang et al. (2012) bahwa analisis terhadap SNP T3941G dan C3975A yang merupakan mutasi gen yang berada intron 4. Hal ini menunjukan bahwa meskipun region intron tidak terlibat dalam sintesis protein namun variasi dari intron dapat mempengaruhi proses translasi oleh beberapa faktor yang belum diketahui.
Frekuensi Alel Gen Prolaktin
Suatu populasi dinilai beragam jika memiliki dua atau lebih alel dalam satu lokus dengan frekuensi yang cukup (biasanya lebih dari 1%). Suatu alel dikatakan polimorfik jika memiliki frekuensi alel sama dengan atau kurang dari 0.99 (Falconer dan Mackay 1996).
Noor (2008) menjelaskan bahwa dalam Hukum Hardy-Weinberg dinyatakan frekuensi genotipe suatu populasi yang cukup besar akan selalu dalam keadaan seimbang bila tidak ada seleksi, migrasi, mutasi, dan genetic drift. Silang dalam dan silang luar dalam kelompok/populasi yang sama (endogami) juga dapat
Formatted: Font color: Black
Formatted: Font color: Black
Formatted: Font color: Black
mempengaruhi frekuensi alel (Vasconcellos et al. 2003). Menurut Falconer dan Mackay (1996), ukuran populasi juga mempengaruhi perubahan frekuensi alel dari generasi ke generasi. Hasil perhitungan frekuensi alel pada enam titik mutasi disajikan pada Tabel 6.
Tabel 6 Analisis frekuensi alel gen prolaktin ekson 4 dan sebagian intron 3 dan 4
No Mutasi Frekuensi Alel (%)* Keterangan
1 INDEL 3724A 2.67 Polimorfik
2 INSERSI 3939A 100.00 Monomorfik
3 T3941G 25.33 Polimorfik
4 C3975A 32.00 Polimorfik
5 INDEL 4031A 32.00 Polimorfik
6 T4110A 14.67 Polimorfik
Keterangan: (*) Perhitungan frekueasi alel merupakan frekuensi alel basa yang bermutasi pada setiap SNP
Hasil analisis menunjukan bahwa frekuensi alel pada titik mutasi INDEL 3724A, T3941G, C3975A, INDEL 4031A dan T4110A berturut-turut sebesar 2.67%; 25.33%; 32%; 32% dan 14.67%, sedangkan pada titik mutasi INDEL 3939A menunjukan hasil perhitungan frekuensi alel sebesar 100%. Nei (1987) melaporkan bahwa keragaman genetik terjadi apabila terdapat dua alel atau lebih dalam suatu populasi (<99%). Berdasarkan literatur maka dapat disimpulkan bahwa titik mutasi INDEL 3724A, T3941G, C3975A, INDEL 4031A dan T4110A pada itik sampel adalah polimorfik karena terdapat dua tipe genotipe yang berbeda dengan persentase (<99%) sedangkan INDEL 3939A adalah monomorfik karena hanya terdapat satu macam genotipe dimana perhitungan frekuensi genotipe sebesar 100% menunjukan INDEL 3939A terjadi pada semua sampel itik.
Populasi itik pada penelitian ini dikatakan beragam. Informasi keragaman genetik suatu bangsa akan sangat bermanfaat bagi keamanan dan ketersediaan bahan pangan yang berkesinambungan (Blott et al. 2003). Khususnya pada penelitian ini keragaman genetik digunakan sebagai dasar dalam pencarian penanda genetik terhadap fenotipe yang diinginkan yang selanjutnya dilakukan program seleksi pada tingkat molekuler. Keragaman genetik dapat digunakan sebagai parameter dalam mempelajari genetika populasi dan genetika evolusi (Nei dan Kumar 2000).
Analisis Perbedaan Jenis Itik Terhadap SNP
Tabel 7 Analisis singel nucleotide polymorphism terhadap jenis itik yang berbeda
Keterangan: (...) = jumlah sampel
Berdasarkan data yang disajikan pada Tabel 6 terlihat bahwa pada semua titik mutasi (INDEL 3724A, T3941G, C3975A, INDEL 4031A dan T4110C) memiliki nilai χ2
hitungyang yang lebih kecil dibandingkan dengan nilai χ2 tabel(χ2 hitung <χ2 tabel (0.05; 8)). Berdasarkan hipotesis jika χ2 hitung < χ2 tabel maka terima H0,
dimana H0=tidak terdapat hubungan yang signifikan antara perbedaan SNP terhadap jenis itik yang berbeda. Hasil analisis statistik menunjukan bahwa tidak ada hubungan yang signifikan antara perbedaan jenis SNP yang ditemukan dengan ketiga jenis bangsa itik yang berbeda, dengan kata lain bahwa faktor perbedaan ketiga bangsa itik yang digunakan pada penelitian ini tidak berpengaruh terhadap kejadian ditemukannya perbedaan SNP pada penelitian.
Analisis Asosiasi SNP dengan Sifat Reproduksi Itik
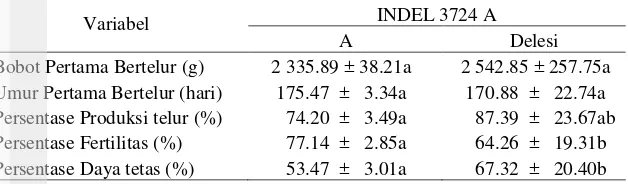
Ketika hubungan antara polimorfisme DNA dan sifat-sifat reproduksi ditemukan, maka polimorfisme ini merupakan potensi penanda genetik untuk program marker assister selection (MAS). Analisis asosiasi SNP gen prolaktin terhadap sifat-sifat reproduksi itik dapat dilihat pada Tabel 8, 9, 10, 11 dan 12. Pada Tabel 8 tersaji analisis asosiasi mutasi INDEL 3724A terhadap sifat-sifat reproduksi.
Tabel 8 Analisis LSMEAN ± SE INDEL 3724 A terhadap parameter reproduksi
Variabel INDEL 3724 A
Keterangan : Angka-angka pada baris yang sama yang diikuti oleh huruf yang sama tidak berbeda nyata pada taraf uji 5%.
terhadap sifat daya tetas telur menunjukan bahwa itik yang mengalami mutasi INDEL 3724A sangat signifikan memiliki persentase daya tetas telur yang lebih besar (P<0.0001) dibandingkan dengan itik yang tidak bermutasi.
Analisis asosiasi mutasi substitusi T3941G terhadap sifat-sifat reproduksi itik tersaji pada Tabel 9.
Tabel 9 Analisis LSMEAN ± SE T3941G terhadap parameter reproduksi
Variabel T3941G
Keterangan : Angka-angka pada baris yang sama yang diikuti oleh huruf yang sama tidak berbeda nyata pada taraf uji 5%.
Hasil analisis Tabel 9 menunjukan bahwa mutasi T3941G berasosiasi dengan sifat produksi, fertilitas dan daya tetas telur. Itik yang mengalami mutasi T3941G memiliki angka persentase produksi telur yang lebih kecil dibandingkan dengan itik-itik yang tidak bermutasi (P<0.05). Analisis terhadap sifat persentase fertilitas dan persentase daya tetas telur menunjukan bahwa itik yang mengalami mutasi T3941G secara signifikan memiliki persentase fertilitas dan daya tetas telur yang lebih besar (P<0.0001) dibandingkan dengan itik yang tidak bermutasi.
Analisis asosiasi mutasi substitusi C3975A terhadap sifat-sifat reproduksi itik, dapat dilihat pada Tabel 10.
Tabel 10 Analisis LSMEAN ± SE C3975A terhadap parameter reproduksi
Variabel C3975A
Keterangan : Angka-angka pada baris yang sama yang diikuti oleh huruf yang sama tidak berbeda nyata pada taraf uji 5%.
Berdasarkan hasil analisis asosiasi Tabel 10 menunjukan bahwa mutasi C3975A berasosiasi dengan sifat produksi telur, fertilitas dan daya tetas telur. Itik yang mengalami mutasi C3975A memiliki angka persentase produksi telur yang lebih kecil dibandingkan dengan itik-itik yang tidak bermutasi (P<0.05). Analisis terhadap sifat fertilitas dan daya tetas telur, itik yang mengalami mutasi C3975A secara sangat signifikan memiliki persentase fertilitas dan daya tetas telur yang lebih besar (P<0.0001) dibandingkan dengan itik yang tidak bermutasi.
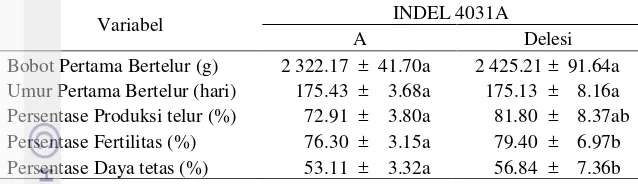
Tabel 11 Analisis LSMEAN ± SE INDEL 4031A terhadap parameter reproduksi
Keterangan : Angka-angka pada baris yang sama yang diikuti oleh huruf yang sama tidak berbeda nyata pada taraf uji 5%.
Berdasarkan hasil analisis asosiasi Tabel 11 menunjukan bahwa mutasi INDEL 4031A berasosiasi dengan sifat produksi telur, fertilitas dan daya tetas telur. Itik yang mengalami mutasi INDEL 4031A memiliki persentase produksi telur yang lebih tinggi dibandingkan dengan itik-itik yang tidak bermutasi (P<0.05). Analisis terhadap sifat fertilitas dan daya tetas telur, itik yang mengalami mutasi INDEL4031A secara signifikan memiliki persentase fertilitas dan daya tetas telur yang lebih tinggi (P<0.0001) dibandingkan dengan itik yang tidak bermutasi.
Analisis asosiasi mutasi substitusi T4110C terhadap sifat-sifat reproduksi itik, dapat dilihat pada Tabel 12 sebagai berikut.
Tabel 12 Analisis LSMEAN ± SE T4110C terhadap parameter reproduksi
Variabel T4110C
Keterangan : Angka-angka pada baris yang sama yang diikuti oleh huruf yang sama tidak berbeda nyata pada taraf uji 5%.
Talbot and Sharp (1994) menyatakan bahwa hormon prolaktin memiliki efek yang krusial terhadap produksi telur, hal ini dikarenakan dimulainya tingkah laku mengeram pada unggas dirangsang oleh peningkatan sekresi hormon prolaktin. Penelitian yang lebih lanjut mengenai mekanisme sifat mengeram menunjukan bahwa meningkatkan produksi telur dapat dilakukan dengan cara menghilangkan sifat mengeram pada unggas. Mengurangi atau menghilangkan sifat mengeram pada unggas dilakukan dengan cara menghambat ekspresi dan sekresi prolaktin (Rozenboim et al. 1996). Cui et al. (2006) pada penelitiannya menganalisis asosiasi polimorfisme region promotor gen prolaktin terhadap sifat produksi telur pada ayam, hasil penelitian menunjukan ditemukan enam SNP pada region tersebut dan hasil analisis kombinasi haplotipe menunjukan bahwa terdapat asosiasi antara kombinasi haplotipe yang disusun dengan sifat produksi telur.
Analisis polimorfisme antara gen prolaktin dengan persentase fertilitas telur pada itik dilaporkan oleh Chang et al. (2012). Penelitiannya dilakukan dengan menyusun kombinasi haplotipe dari 6 SNP yang selanjutnya digunakan untuk analisis kombinasi haplotipe dengan sifat reproduksi. Hasil menunjukan bahwa kombinasi haplotipe (diplotipe) memiliki efek positif yang signifikan terhadap bobot telur, persentase fertilitas dan juga maximum duration fertility (periode fertilitas telur maksimum). Zang et al. (2008) menyatakan bahwa interaksi antara efek dari perbedaan SNP dan asosiasi diplotipe dengan sifat-sifat reproduksi yang lebih akurat dibanding dengan analisis terhadap SNP tunggal.
Faktor yang mempengaruhi jumlah dan pembentukan komponen telur, tingkat produksi, fertilitas dan daya tetas telur bergantung pada berbagai sinkronisasi metabolisme dan proses fisiologi. Sifat-sifat reproduksi ini telah diketahui secara signifikan dipengaruhi oleh berbagai faktor, terutama oleh faktor lingkungan (Brah dan Sandhu 1989). Meskipun begitu, hasil-hasil penelitian terkini menunjukan bahwa parameter produksi dan reproduksi juga dipengaruhi oleh faktor genetik Miazi et al. (2012). Diantara banyak gen yang memiliki efek signifikan terhadap daya tetas telur salah satunya adalah gen prolaktin. Hal ini terbukti dari penelitian yang dilakukan oleh Chang et al. (2007) menyimpulkan cDNA microarray prolaktin dapat mengakibatkan pembentukan ovarian follicular dan juga formasi telur. Hal ini yang mendasari penelitian lanjutan yang dilakukan Chang et al. (2012) yang menginvestigasi hubungan antara SNP prolaktin dengan sifat reproduksi itik. Dari hasil penelitiannya ditemukan enam SNP baru (T233C, T295C, G309T, C381A, G3941T dan A3975C).
5
SIMPULAN DAN SARAN
Simpulan