DETEKSI GEN KETAHANAN PENYAKIT HAWAR
DAUN BAKTERI PADA POPULASI SILANG
GANDA F1 (DCF1) PADI CIHERANG
HERMANTO
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa skripsi berjudul Deteksi Gen Ketahanan Penyakit Hawar Daun Bakteri pada Populasi Silang Ganda F1 (DCF1) Padi Ciherang adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Agustus 2014
ABSTRAK
HERMANTO. Deteksi Gen Ketahanan Penyakit Hawar Daun Bakteri pada Populasi Silang Ganda F1 (DCF1) Padi Ciherang. Dibimbing oleh LAKSMI AMBARSARI dan FATIMAH.
Xanthomonas oryzae pv. oryzae penyebab penyakit hawar daun memiliki banyak strain dan mudah berubah membentuk strain baru. Akibatnya varietas padi yang awalnya tahan dapat berubah menjadi peka. Perbaikan varietas Ciherang menggunakan pyramiding gen ketahanan penyakit hawar daun bakteri akan membawa ketahanan yang berbeda dan memiliki spektrum yang luas. Penelitian ini bertujuan mendeteksi 207 individu DCF1 Ciherang hasil pyramiding gen dengan menggunakan marka SSR dan STS. Diperoleh empat Individu memiliki gen Xa4+xa5+Xa7+Xa21 yaitu individu DCF1Ch|CA-42, DCF1Ch|CA-69, DCF1Ch|CA-162, DCF1Ch|CA-163, 11 individu memiliki gen xa5+Xa7+Xa21, 46 individu memiliki gen xa5+Xa21, 31 individu memiliki gen Xa7+Xa21, dan 45 individu memiliki gen xa5+Xa7. Karakter agronomi jumlah malai DCF2 Ciherang tidak berbeda nyata dengan tetuanya dan panjang malai DCF2 Ciherang tidak berbeda signifikan dengan tetuanya. Bobot gabah isi DCF2 Ciherang lebih besar secara nyata dibandingkan tetuanya. Bobot gabah hampa DCF2 Ciherang juga berbeda nyata dengan tetuanya, lebih besar dari tetua Code, Angke, Ciherang, dan IRBB21.
Kata kunci: hawar daun bakteri, padi Ciherang, pyramiding gen, silang ganda.
ABSTRACT
HERMANTO. Detection of Resistance Genes Bacterial Leaf Blight on Population Double Cross F1 (DCF1) Rice Ciherang. Supervised by LAKSMI AMBARSARI and FATIMAH.
Xanthomonas oryzae pv. oryzae causes leaf blight has many strains and volatile form a new strain. A result that was initially resistant rice varieties may turn out to be sensitive. Improvement Ciherang using gene pyramiding bacterial leaf blight resistance will carry different resistance and has a broad spectrum. This research aims to detect the 207 individual DCF1 Ciherang results gene pyramiding using SSR markers and STS. Four individuals have acquired genes Xa4+xa5+Xa7+Xa21 that is DCF1Ch|CA-42, DCF1Ch|CA-69, DCF1Ch|CA-162, DCF1Ch|CA-163, 11 individuals have genes xa5+Xa7+Xa21, 46 individuals have xa5+Xa21 gene, 31 individuals have Xa7+Xa21 gene, and 45 individual have genes xa5+Xa7. Agronomic characters panicle number DCF2 Ciherang not significantly different from the parent and panicle length DCF2 Ciherang did not differ significantly with parent. Weight of filled grain DCF2 Ciherang significantly greather than the parent. Weight of empty grains DCF2 Ciherang also significantly different from the parent, greather than elders Code, Angke, Ciherang, and IRBB21.
DETEKSI GEN KETAHANAN PENYAKIT HAWAR
DAUN BAKTERI PADA POPULASI SILANG
GANDA F1 (DCF1) PADI CIHERANG
HERMANTO
Skripsi
sebagai salah satu syarat memperoleh gelar Sarjana Sains
pada
Departemen Biokimia
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
PRAKATA
Puji syukur penulis panjatkan kepada Allah SWT atas segala rahmat dan karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Karya ilmiah yang berjudul “Deteksi Gen Ketahanan Penyakit Hawar Daun Bakteri pada Populasi Silang Ganda F1 (DCF1) Padi Ciherang” merupakan bagian dari proyek penelitian Dr Fatimah SP MSi dan dilakukan di Laboratorium Biologi Molekuler, Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian (BB-Biogen), Jalan Tentara Pelajar nomor 3A, Cimanggu, Bogor, Jawa Barat. Dilaksanakan sejak bulan Desember 2013 sampai Mei 2014.
Terima kasih penulis ucapkan kepada Dr Laksmi Ambarsari MS dan Dr Fatimah SP MSi atas bimbingan, motivasi, serta arahan yang telah diberikan. Penulis mengucapkan banyak terima kasih kepada kedua orang tua dan adik tercinta (Yusi Suciati dan Fitria Novian), Ayu Kartika, serta seluruh keluarga atas doa dan kasih sayangnya. Penulis juga mengucapkan terima kasih kepada Mba Muslihatun Baroya, Staf di Laboratorium Biologi Molekuler BB-Biogen, rekan selama penelitian, teman-teman Biokimia angkatan 2010, dan rekan sejawat atas segala bantuan, motivasi dan ilmu yang diberikan.
Penulis menyadari masih banyak kekurangan dalam karya ilmiah ini. Oleh karena itu, penulis mengharapkan kritik dan saran yang membangun untuk perbaikan penelitian selanjutnya. Semoga karya ilmiah ini bermanfaat.
Bogor, Agustus 2014
DAFTAR ISI
DAFTAR TABEL viii
DAFTAR GAMBAR viii
DAFTAR LAMPIRAN viii
PENDAHULUAN 1
METODE 2
Bahan dan Alat 2
Prosedur Penelitian 3
HASIL 6
Deteksi Gen Xa4 Menggunakan Primer MP1+MP2 6
Deteksi Gen xa5 Menggunakan Primer RM611 7
Deteksi Gen Xa7 Menggunakan Primer RM20582 7
Deteksi Gen Xa21 Menggunakan Primer pTA248 8
Karakter Agronomi Hasil Panen 9
Representasi Data Genotipe 9
PEMBAHASAN 10
Deteksi Gen Xa4, xa5, Xa7, dan Xa21 10
Karakter Agronomi Hasil Panen 12
Representasi Data Genotipe 13
SIMPULAN DAN SARAN 15
Simpulan 15
Saran 16
DAFTAR PUSTAKA 16
LAMPIRAN 19
DAFTAR TABEL
1. Genotipe 207 individu DCF1 Ciherang 8
2. Karakter agronomi hasil panen tetua dan DCF2 Ciherang 9
DAFTAR GAMBAR
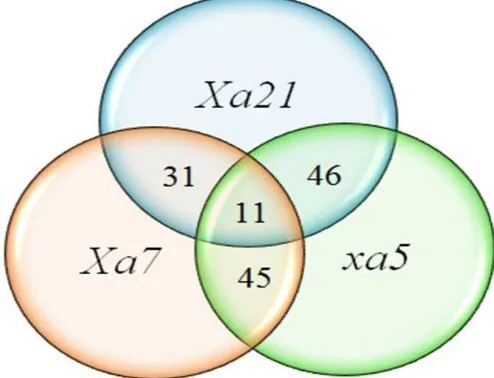
1. Elektroforegram hasil validasi gen Xa4 dengan primer MP1+MP2 6 2. Elektroforegram hasil validasi gen xa5 dengan primer RM611 7 3. Elektroforegram hasil validasi gen Xa7 dengan primer RM20582 7 4. Elektroforegram hasil validasi gen Xa21 dengan primer pTA248 8 5. Perkiraan heterozigositas gen ketahanan hawar daun bakteri 10 6. Diagram venn 3 gen ketahanan HDB padi DCF1 Ciherang 11
7. Keterkaitan gen resisten HDB dan bakteri Xoo 15
DAFTAR LAMPIRAN
1. Diagram alir penelitian 19
2. Diagram persilangan 19
3. Elektroforegram hasil amplifikasi gen Xa4 dengan primer MP1+MP2 20 4. Elektroforegram hasil amplifikasi gen xa5 dengan primer RM611 20 5. Elektroforegram hasil amplifikasi gen Xa7 dengan primer RM20582 20 6. Elektroforegram hasil amplifikasi gen Xa21 dengan primer pTA248 20 7. Hasil skoring elektroforegram DCF1 Ciherang dan tetuanya 21
8. Karakter agronomi hasil panen 28
PENDAHULUAN
Padi (Oryza sativa L.) merupakan tanaman pangan penghasil beras yang paling penting dan dikonsumsi oleh setengah populasi penduduk dunia, termasuk Indonesia (Yang et al. 2003). Kebutuhan akan padi terus meningkat sebagai makanan pokok seiring dengan semakin meningkatnya laju pertumbuhan penduduk. Berdasarkan estimasi laju pertumbuhan penduduk 1.3 persen pada rentang waktu 2005-2010, tingkat konsumsi beras bruto 139.5 kilogram perkapita maka kebutuhan beras 32.49 juta ton. Asumsi lain laju pertumbuhan penduduk 0.92 persen yang diperkirakan pada rentang waktu 2025-2030 dengan tingkat konsumsi 139.5 kilogram perkapita maka kebutuhan beras mencapai 39.8 juta ton (Kementrian Pertanian 2012).
Penggunaan padi varietas unggul bagi sebagian besar petani sangat berperan dalam meningkatkan produktivitas. Hal ini tercermin dari luas tanaman padi yang dewasa ini telah didominasi oleh varietas unggul (Pusat Penelitian dan Pengembangan Tanaman Pangan 2006). Salah satu varietas unggul tersebut ialah Ciherang. Penyebaran varietas Ciherang terus berkembang di Indonesia sejak dilepas pada tahun 2000, dan terus mendominasi luas lahan padi di Jawa Barat dari tahun 2009 hingga tahun 2011. Dinas Pertanian dan Tanaman Pangan Provinsi Jawa Barat (2013) menyebutkan bahwa pada masa tanam 2010/2011 persentase luas lahan untuk varietas Ciherang yaitu 55% atau sekitar 195.455 hektar dari luas lahan padi varietas unggul lainnya. Akan tetapi, peningkatan produksi padi sering mengalami banyak kendala karena adanya cekaman abiotik maupun biotik. Cekaman abiotik berupa kekeringan, banjir atau keracunan aluminium, besi, dan logam-logam lainnya, sedangkan cekaman biotik meliputi serangan hama dan penyakit seperti penyakit Hawar Daun Bakteri (HDB).
2
antara Ciherang dan padi galur IRBB21. Hasil persilangan dari masing-masing varietas (generasi pertama/F1) akan disilangkan kembali dengan cara double cross. Double cross atau silang ganda merupakan persilangan antara dua silang tunggal (F1) yang melibatkan empat galur murni yang tidak berhubungan satu sama lain. Double cross dari generasi F1 tersebut diharapkan memiliki pyramiding gen ketahanan terhadap hawar daun bakteri yaitu gen ketahanan Xa4, xa5, Xa7, dan Xa21 yang dideteksi dengan marka molekuler.
Menurut Balai Besar Bioteknologi dan Pengembangan Sumberdaya Genetik (2007), analisis molekuler melalui marka DNA sangat diperlukan untuk membantu seleksi galur baru tanaman yang benar-benar mengandung gen yang ditargetkan. Blair et al. (2003) menggunakan primer RM611 dengan tipe SSR (Simple Sequence Repeat) untuk mengidentifikasi gen xa5 pada kromosom 5. Primer RM20582 dengan marka SSR digunakan untuk mengidentifikasi gen Xa7 pada kromosom 6 (Chen et al. 2008). Singh et al. (2001) menggunakan primer pTA 248 dengan marka STS (Sequence-Tagged Sites) untuk mengidentifikasi gen Xa21 pada kromosom 11, dan primer MP1+MP2 dengan marka tipe STS untuk mengidentifikasi gen Xa4 (Ma et al. 1999).
Penelitian ini bertujuan mendeteksi keberadaan gen ketahanan Xa4, xa5, Xa7, dan Xa21 melalui teknik marka molekuler pada padi hasil silang ganda atau double cross F1 (DCF1) Ciherang. Penelitian ini diharapkan dapat mempermudah dan mempersingkat pemilihan genotipe ketika melakukan pemuliaan varietas unggul secara berkelanjutan guna meningkatkan produktivitas tanaman padi Ciherang yang tahan terhadap hawar daun bakteri (HDB).
METODE
Bahan dan Alat
Bahan tanaman yang digunakan untuk mendeteksi adanya gen ketahanan Xa4, xa5, Xa7, dan Xa21 yaitu: tetua Code, Angke, Ciherang, IRBB21, dan 207 tanaman yang hidup dari 250 tanaman hasil silang ganda F1 yang diduga memiliki pyramiding gen dari persilangan tetua Code × Angke dan Ciherang × IRBB21. Bahan yang digunakan untuk media tanam padi, yaitu: kapas, akuades, tanah, pupuk (urea, TSP, KCL), dan air. Bahan yang digunakan untuk isolasi DNA adalah daun padi yang masih muda, es, nitrogen cair, bufer ekstrak (Tris-HCl 100 mM pH 8.0, EDTA 50 mM, NaCl 500 mM, SDS (Sodium Dodecyl Sulphate) 1.25%, NaOH 8.3 mM, dan 0.38 gram sodium bisulfit/100 mL), chisam 24:1 (kloroform:isoamil v/v), Etanol absolut, Etanol 70%, NaOAc (natrium asetat), dan ddH2O. Bahan yang digunakan untuk amplifikasi DNA dengan PCR adalah DNA
hasil isolasi, es, PCR mix (ddH2O, 10x PCR bufer, dNTPs 2.5 mM, GCRich
3
digunakan untuk visualisasi hasil elektroforesis yaitu EtBr (ethidium bromida), silver staining (silver nitrat, ddH2O, NaOH, dan formaldehida).
Alat-alat yang digunakan dalam penelitian ini yaitu: cawan petri, inkubator benih, pinset, bak plastik, karton, bolpen, gunting, box es, sudip, tabung mikro, neraca analitik, termos, gelas ukur, sentrifuse Backman MicrofugeTM 12, gelas piala, inkubator, mesin autoklaf, labu Erlenmeyer, pipet mikro (1000µ, 200µ, 20µ, dan 2µ), tabung Eppendorf, tip, PCR plate, stirer, dan beberapa perangkat alat seperti air flow cabinet, elektroforesis horizontal, PS500XT DC power supply, mesin PCR Bio Rad T100TM Thermal Cycler, oven microwave, minivortex Bio Rad, Nanodrop Thermo Scientific, lemari asam, kulkas, spektrofotometer Bio Rad SmartSpecTM plus, serta perangkat untuk analisis hasil elektroforegram yaitu Chemidoc UV-Illuminator Bio-Rad.
Prosedur Penelitian
Penumbuhan Padi DCF1 Ciherang x IRBB21//Code x Angke
Benih tetua padi dan 250 benih DCF1 Ciherang dioven terlebih dahulu pada suhu 50°C selama semalam. Selanjutnya benih tersebut disemai di cawan petri yang telah dilapisi kapas basah. Setelah akar dan batang tanaman padi bertunas, kemudian tanaman padi dipindahkan ke dalam bak plastik sebagai masa adaptasi. Setelah dua minggu berada dalam bak plastik, tanaman dipindahkan ke dalam ember dan dibiarkan tumbuh selama kurang lebih satu bulan agar dapat diisolasi DNA dari daun mudanya.
Isolasi DNA Padi Metode Miniprep
4
Analisis Kualitatif DNA Padi
Kualitas DNA padi diuji menggunakan elektroforesis gel agarose 0.8% (Sambrook & Russell 2001). Agarose 0.8% dibuat dengan menambahkan 0.8 gram agarose dengan 100 mL TBE 0.5x dan dipanaskan dalam microwave 2-4 menit hingga larut. Selanjutnya larutan dituang ke dalam cetakan agar yang sudah disiapkan dan dipasangkan sisir untuk sumur sesuai dengan banyaknya sampel. Setelah gel agarose memadat, gel dimasukkan ke dalam bak elektroforesis yang berisi bufer TBE 0.5x. Sebanyak 1 μL sampel DNA ditambahkan dengan 1 μL loading dye dan dicampur sempurna, kemudian dimasukkan ke dalam sumur gel. Selanjutnya, sampel DNA dialiri arus listrik dengan daya 100 volt selama ± 90 menit. Hasil elektroforegram divisualisasi dengan Chemidoc UV-Transiluminator.
Analisis Kuantitatif DNA Genomik dengan Nanodrop
Kuantitas DNA dianalisi dengan Nanodrop (Thermo Fisher Scientific 2009). Sebanyak 1 μL ddH2O dimasukkan ke dalam lubang ukur. Setelah itu, tutup
nanodrop dan tekan tombol read blank pada komputer. Kertas tissue digunakan untuk membersihkan sisa ddH2O tersebut. Masing-masing sampel DNA
dimasukkan sebanyak 1 μL ke dalam lubang ukur secara bergantian dan dipilih menu read sample. Hasil pengukuran nilai kemurnian sampel DNA akan muncul dalam satuan konsentrasi ng/μL. DNA yang murni memiliki rasio serapan pada 260 nm dan 280 nm (A260/A280) sebesar 1.8 atau lebih.
Amplifikasi DNA Padi dengan PCR
Pembuatan mix PCR untuk mengamplifikasi lokus gen ketahanan Hawar Daun Bakteri dengan sampel padi Code, Angke, Ciherang, individu DCF1 Xa21); sehingga volume total menjadi 20 μL.
Proses amplifikasi DNA dilakukan dengan menggunakan mesin PCR Bio Rad T100TM. Amplifikasi gen Xa21 terdiri atas beberapa tahapan reaksi, yaitu predenaturasi pada suhu 94°C selama 5 menit, denaturasi pada suhu 94°C selama 1 menit, annealing (penempelan primer) pada suhu 60°C selama 1 menit, extension (pemanjangan rantai DNA yang telah ditempeli primer)pada suhu 72°C selama 1 menit, dan pasca pemanjangan primer pada suhu 72°C selama 10 menit. Proses amplifikasi untuk gen Xa4, xa5 dan gen Xa7 terdiri atas predenaturasi pada suhu 94°C selama 5 menit, denaturasi pada suhu 94°C selama 45 detik, annealing pada suhu 55°C selama 45 detik, extension pada suhu 72°C selama 1 menit 45 detik, dan pasca pemanjangan primer pada suhu 72°C selama 10 menit.
Elektroforesis Hasil Amplifikasi
5
larutan dituang ke dalam cetakan. Setelah gel agarose memadat, gel dimasukkan ke dalam bak elektroforesis yang berisi bufer TBE 0.5x. Sebanyak 6 μL loading dye ditambahkan pada hasil amplifikasi, kemudian 5 μL sampel dimasukkan ke dalam sumur gel. Elektroforesis dilakukan dengan daya 95 volt selama 1-2 jam. Hasil elektroforegram divisualisasi dengan Chemidoc UV-Transiluminator.
Analisis gen xa5, Xa4 dan Xa7 menggunakan PAGE. Gel poliakrilamid dibuat dengan mencampurkan 50 mL poliakrilamid 8%, 800 μL APS (ammonium persulfat) 10%, dan 50 μL TEMED (tetrametilen-etilendiamin). Campuran diaduk hingga merata lalu dituangkan ke dalam cetakan kaca dan didiamkan hingga memadat selama 15 menit. Kemudian gel poliakrilamid dimasukkan ke dalam tangki elektroforesis dan dimasukkan bufer TBE 0.5x. Setelah gel siap, produk PCR yang telah ditambahkan 5 μL loading dye dimasukkan ke dalam sumur gel sebanyak 3 μL dan disertakan DNA marker 100 bp ladder sebanyak 3 μL sebagai pembanding pada sumur pertama untuk melihat ukuran DNA. Selanjutnya sampel DNA dialiri arus listrik 70 volt selama 120 menit. Gel poliakrilamid yang telah selesai dirunning, kemudian dicuci dengan ddH2O. Gel diwarnai dengan larutan silver staining dengan komposisi 0.2 gram perak nitrat dan 200 mL ddH2O,
digoncangkan menggunakan mesin Roto Mix 5-8 menit, kemudian dihilangkan pewarnaannya dengan ddH2O. Gel dicuci kembali dengan larutan yang terdiri atas
200 mL ddH2O, 3 gram NaOH, 1.5 mL CH2O (formaldehida) dalam Roto Mix
selama 8-10 menit, kemudian gel dicuci dengan ddH2O dan didokumentasikan.
Evaluasi Karakter Agronomi Komponen Hasil Panen
Evaluasi karakter agronomi dilakukan dengan pengamatan secara kualitatif dan kuantitatif ketika padi mencapai masa panen (percobaan dilakukan 3 kali ulangan). Evaluasi hasil panen secara kualitatif meliputi pengamatan panjang malai. Sementara evaluasi secara kuantitatif meliputi pengamatan jumlah malai, bobot gabah isi, dan bobot gabah hampa. Panjang malai diukur dari ujung malai sampai pangkal sumbu utama. Jumlah malai dihitung berdasarkan banyaknya malai pada satu rumpun padi. Bobot gabah isi dan bobot gabah hampa dihitung dari setiap rumpun padi.
Analisis Data Karakter Agronomi Hasil Panen
Data evaluasi karakter agronomi hasil panen dianalisis secara statistik menggunakan program SPSS dengan metode percobaan rancangan acak lengkap (RAL) dan menggunakan metode ANOVA (Analysis of Variance) (Mattjik 2002) serta uji lanjut menggunakan Duncan.
Model rancangannya yaitu Yij = μ + τi + εij pada selang kepercayaan 95%. Keterangan:
i = 1,2,3,...7 dan j= 1,2,3
Yij = Pengamatan pada karakter agronomi ke-i dan ulangan ke-j μ = Pengaruh rataan umum
τi = Pengaruh karakter agronomi ke-i
Representasi Data dengan Graphical Genotypes
6
Genotypes 2.0 (Berloo 2007) dengan skoring pita H untuk sampel DNA yang memiliki pita Heterozigot, A untuk sampel yang mengikuti pola pita tetua Code, Angke, dan IRBB21. B untuk sampel DNA yang mengikuti pola pita Ciherang.
HASIL
Dua ratus tujuh individu DCF1 Ciherang hasil pyramiding gen dan 4 galur tetua, yaitu Code, Angke, Ciherang, dan IRBB21 dideteksi dengan marka mikrosatelit (SSR) dan STS yang berbasis PCR untuk mengetahui adanya lokus 4 gen ketahanan HDB. Individu homozigot ditunjukkan oleh hasil amplifikasi yang mengikuti satu pola pita tetua, sedangkan individu heterozigot ditunjukkan oleh hasil amplifikasi yang mengikuti dua pola pita tetua. Terdapat 4 individu yang memiliki empat gen tahan hawar daun bakteri, yaitu individu dengan kode DCF1Ch|CA-42, DCF1Ch|CA-69, DCF1Ch|CA-162, DCF1Ch|CA-163.
Deteksi Gen Xa4 Menggunakan Primer MP1+MP2
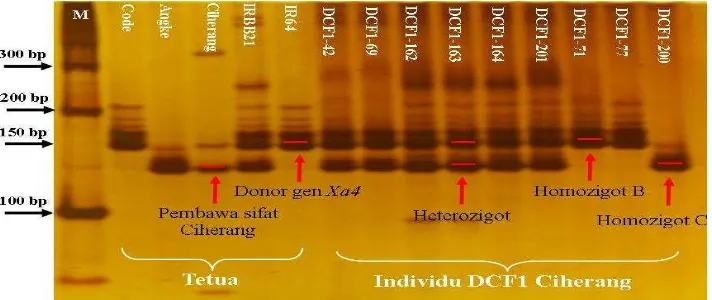
Amplifikasi individu DCF1 Ciherang menggunakan primer MP1+MP2 bertipe STS yang terkait dengan lokus gen Xa4 menunjukkan adanya pita heterozigot mengikuti pola tetua Ciherang sebagai inti tetua betina pembawa sifat Ciherang dan IR64 sebagai donor gen Xa4 dengan ukuran 150 bp yang merupakan penanda sifat resistant (tahan) (Lampiran 3). Elektroforegram hasil amplifikasi DNA dari 207 individu DCF1 Ciherang menunjukkan bahwa terdapat 4 individu yang mengikuti kedua pola tetuanya (heterozigot), sementara individu yang lain mengikuti salah satu pola tetuanya (homozigot) (Lampiran 7). Validasi gen Xa4 menunjukkan individu DCF1 Ciherang nomor 42, 69, 162, dan 163 terdeteksi pada elektroforegram gen ketahanan lainnya (gen xa5, Xa7, dan Xa21) pada penelitian ini (Gambar 1).
7
Deteksi Gen xa5 Menggunakan Primer RM611
Amplifikasi gen resesif xa5 menggunakan primer RM611 tipe mikrosatelit (SSR) menunjukkan adanya pita heterozigot mengikuti fragmen tetua Ciherang sebagai tetua betina pembawa sifat Ciherang dan Angke sebagai donor gen xa5 dengan ukuran 213 bp yang merupakan penanda sifat tahan HDB (Lampiran 4). Elektroforegram hasil amplifikasi DNA menunjukkan 103 individu memiliki pola pita heterozigot penanda sifat resistant (Lampiran 7). Validasi gen xa5 menunjukkan individu DCF1 Ciherang nomor 42, 69, 162, dan 163 terdeteksi pada elektroforegram gen ketahanan lainnya (Gambar 2).
Gambar 2 Elektroforegram hasil validasi gen xa5 dengan primer RM611; (M) marker 100 bp DNA ladder, (Code) kontrol positif gen Xa7, (Angke) kontrol positif gen xa5, (Ciherang) tetua betina pembawa sifat Ciherang, (IRBB21) kontrol positif gen Xa21, (IR64) kontrol positif gen Xa4, (B) mengikuti pola pita Ciherang
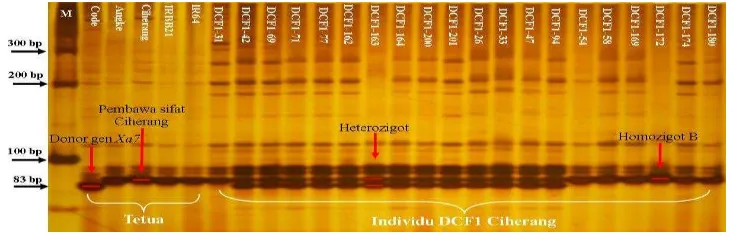
Deteksi Gen Xa7 Menggunakan Primer RM20582
Amplifikasi menggunakan primer RM20582 tipe SSR terkait lokus gen Xa7 menunjukkan adanya pita heterozigot mengikuti pola tetua Ciherang sebagai tetua betina pembawa sifat Ciherang dan Code sebagai donor gen Xa7 dengan ukuran 83 bp yang merupakan penanda sifat tahan (Lampiran 5). Elektroforegram hasil amplifikasi 207 individu DCF1 Ciherang menunjukkan bahwa terdapat 89 individu yang mengikuti kedua pola tetuanya (Lampiran 7). Validasi gen Xa7 menunjukkan individu DCF1 Ciherang nomor 42, 69, 162, dan 163 terdeteksi pada elektroforegram gen ketahanan lainnya (Gambar 3).
8
Deteksi Gen Xa21 Menggunakan Primer pTA248
Amplifikasi lokus gen Xa21 dengan primer pTA248 tipe STS menunjukkan adanya pita heterozigot mengikuti pola tetua Ciherang sebagai tetua betina pembawa sifat Ciherang dan IRBB21 sebagai donor gen Xa21 dengan ukuran 1000 bp sebagai penanda sifat resistant dan 650 bp penanda sifat susceptible (rentan) (Lampiran 6). Elektroforegram hasil amplifikasi DNA dari 207 individu DCF1 Ciherang menunjukkan 68 individu memiliki pola pita heterozigot yang sejajar dengan Ciherang dan IRBB21 sebagai penanda sifat resistant (Lampiran 7). Validasi gen Xa21 menunjukkan individu DCF1 Ciherang nomor 42, 69, 162, dan 163 terdeteksi pada elektroforegram gen ketahanan lainnya (Gambar 4).
Gambar 4 Elektroforegram hasil validasi gen Xa21 dengan primer pTA248; (M) marker 100 bp DNA ladder, (Code) kontrol positif gen Xa7, (Angke) kontrol positif gen xa5, (Ciherang) tetua betina pembawa sifat Ciherang, (IRBB21) kontrol positif gen Xa21, (IR64) kontrol positif gen Xa4, (B) mengikuti pola pita Ciherang
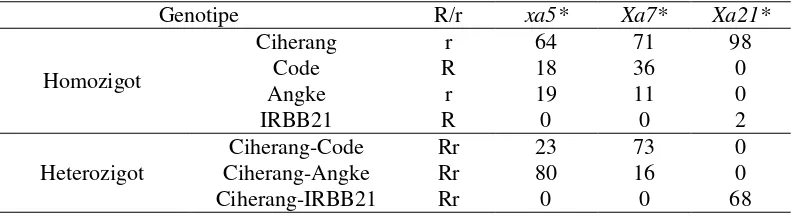
Genotipe DCF1 Ciherang
Genotipe 207 individu DCF1 Ciherang dari hasil deteksi empat gen ketahanan diperoleh bahwa individu yang heterozigot mengikuti tetua Ciherang-Code sebanyak 23 individu, dan mengikuti tetua Ciherang-Angke sebanyak 80 individu untuk gen xa5. Genotipe individu untuk gen Xa7 diperoleh 73 individu yang heterozigot mengikuti tetua Ciherang-Code, dan 16 individu mengikuti tetua Ciherang-Angke. Genotipe individu DCF1 Ciherang untuk gen Xa21 yang mengikuti tetua Ciherang-IRBB21 sebanyak 68 individu. Genotipe individu yang memiliki gen Xa4 tidak ditampilkan dalam tabel 1 karena gen Xa4 dianggap sudah diperoleh dari gen tetua induk Ciherang yaitu IR64. Tabel genotipe ini menunjukkan rekombinasi yang terjadi selama proses meiosis antar kromosom. Tabel 1 Genotipe 207 individu DCF1 Ciherang
9
Karakter Agronomi Hasil Panen
Karakter agronomi pada dasarnya dikendalikan oleh banyak gen. Karakter agronomi baik kualitatif maupun kuantitatif merupakan hasil akhir dari proses pertumbuhan dan perkembangan yang berkaitan langsung dengan karakter fisiologis dan morfologis. Diantara kedua karakter tersebut, karakter morfologis akan lebih mudah diamati sebagai perbandingan produktivitas antara individu DCF2 Ciherang dengan tetuanya. Karakter morfologis yang diamati meliputi jumlah malai, panjang malai, bobot gabah isi, dan bobot gabah hampa (Tabel 2). Karakter agronomi komponen panen secara lengkap terdapat pada lampiran 8. Hasil pengamatan menunjukkan jumlah malai individu DCF2 Ciherang memiliki karakter yang tidak berbeda nyata dengan tetuanya. Panjang malai DCF2 Ciherang 24.10 cm tidak berbeda nyata dengan Code 24.92 cm dan IRBB21 25.51 cm, serta tidak berbeda signifikan dengan tetua Angke 28.98 cm dan Ciherang 23.40 cm. Bobot gabah isi padi DCF2 Ciherang memiliki hasil yang lebih besar yaitu 34.49 gram dibandingkan tetua IRBB21 yaitu 24.05 gram, tetua Code 26.62 gram, dan tetua Ciherang 25.51 gram yang berbeda nyata pada uji selang berganda Duncan taraf 5%, serta tidak berbeda signifikan dengan tetua Angke yang memiliki bobot gabah isi 37.36 gram. Bobot gabah hampa DCF2 Ciherang juga berbeda nyata dengan tetuanya, bobot gabah hampa individu ini lebih besar dari tetua Code, Angke, Ciherang, dan IRBB21. Meskipun bobot gabah isi individu DCF2 Ciherang relatif lebih besar dari tetua Code, Ciherang, dan IRBB21. Akan tetapi apabila bobot gabah hampa individu DCF2 Ciherang ini memiliki massa yang lebih besar dan berbeda nyata dengan tetuanya maka dapat dijadikan pertimbangan dalam pemuliaan varietas Ciherang selanjutnya.
Tabel 2 Karakter agronomi hasil panen tetua dan DCF2 Ciherang
Varietas JM PM BGI BGH berganda Duncen taraf 5%, JM: Jumlah malai (malai), PM: Panjang malai (cm), BGI: Bobot gabah isi (gram), BGH: Bobot gabah hampa (gram).
Representasi Data Genotipe
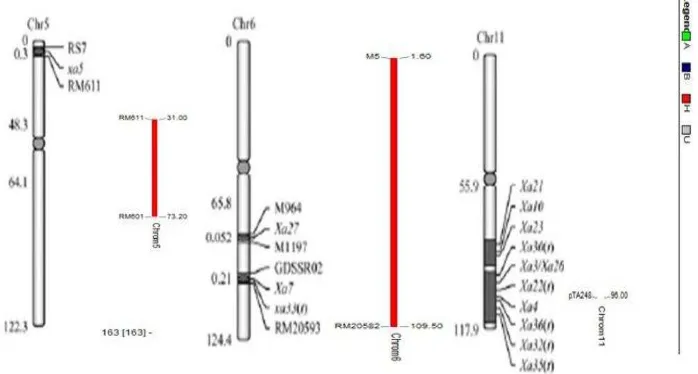
10
di posisi 95.95 cM menggunakan penanda pTA248. Peta kromosom diambil dari gambar perkiraan gen ketahanan hawar daun bakteri oleh Chun et al. (2012).
Gambar 5 Perkiraan heterozigositas gen ketahanan hawar daun bakteri pada kromosom menggunakan software Graphical Genotypes 2.0
PEMBAHASAN
Deteksi Gen Xa4, xa5, Xa7, dan Xa21
Gen resisten terhadap penyakit hawar daun bakteri tidak begitu efektif apabila strategi pengembangan gen resisten HDB tidak dilakukan dengan tepat. Sebagaimana diketahui bahwa penerapan gen tunggal resisten HDB dapat mengendalikan penyakit yang disebabkan oleh bakteri hawar daun. Akan tetapi, bakteri Xoo memiliki patotipe yang tinggi sehingga lebih cepat berkembang untuk mematahkan resistensi dari gen tunggal. Xa4 dan Xa3, dua gen resisten HDB padi hibrida di China dan negara-negara Asia selatan, juga gen resistensi luas seperti Xa21 telah berhasil mengendalikan penyebaran bakteri hawar daun di dunia dalam dua dekade terakhir, tetapi gen resisten tunggal tersebut dapat dipatahkan oleh ras virulen baru dalam beberapa tahun terakhir di Filipina, India, Korea, dan China (Marella et al. 2001; Lee et al. 1999; Zeng et al. 2002). Oleh karena itu, penelitian ini menggunakan gen resisten HDB berbasis pyramiding gen dalam program pemuliaan padi. Pyramiding gen resisten berdasarkan MAS (Marker Assisted Selection) telah menjadi metode yang efektif untuk mengendalikan penyakit bakteri hawar daun (Singh et al. 2001; Zhang et al. 2006; Loida et al. 2008; Gopalakrishna et al. 2008).
11
(Sequence-Tagged Sites). Marka STS juga digunakan dalam penelitian ini untuk mendeteksi gen Xa21 menggunakan primer pTA248 (Singh et al. 2001). STS merupakan sekuen unik pendek yang mengidentifikasi satu atau lebih lokus dan dapat teramplifikasi dengan PCR. Marka STS dapat digunakan dalam pemetaan genetik karena bersifat kodominan (mampu membedakan alel homozigot dan heterozigot), dan menghasilkan amplifikasi yang stabil dan berulang (Azrai 2005). Primer MP1 dan MP2 berhasil mendeteksi gen Xa4 dengan peluang sebesar 1.93% dan primer pTA248 berhasil mendeteksi gen Xa21 dengan peluang sebesar 32.85% pada DCF1 Ciherang.
Deteksi gen resesif xa5 pada penelitian ini mengacu pada penelitian Blair et al. (2003) menggunakan primer RM611, dan deteksi gen Xa7 mengacu pada penelitian Chen et al. (2008) menggunakan primer RM20582. Kedua primer tersebut merupakan marka SSR. Mikrosatelit atau SSR merupakan marka yang digunakan untuk menentukan genotipe individu dengan mendeteksi segmen DNA yang mengandung pola perulangan sederhana dari basa nitrogen yang kemudian dikenali dengan teknik PCR (Mullis dan Faloon 1987). Menurut Prihatin (2006) mikrosatelit dapat dimanfaatkan untuk mendeteksi induk suatu individu serta menguji hasil persilangan individu. Peluang primer RM611 dalam mendeteksi gen xa5 sebesar 49.76% dan peluang primer RM20582 dalam mendeteksi gen Xa7 sebesar 42.99% pada individu DCF1 Ciherang.
Peluang mendapatkan empat gen ketahanan dalam satu individu masih sangat kecil sekitar 1.93%, hal ini disebabkan adanya rekombinasi yang cukup tinggi pada individu hasil persilangan. Individu DCF1 Ciherang yang memiliki pyramiding gen ketahanan terhadap hawar daun bakteri xa5+Xa7+Xa21 sebanyak 11 individu, 46 individu memiliki gen xa5+Xa21, 31 individu memiliki gen Xa7+Xa21, dan 45 individu memiliki gen xa5+Xa7 (Gambar 6). Nisbah tersebut merupakan suatu probabilitas atau kemungkinan dari pemindahan gen tetuanya. Probabilitas dipengaruhi oleh keterkaitan antara dua lokus, jika tidak ada keterkaitan antara dua lokus maka besarnya probabilitas genotipe tertentu untuk setiap generasi adalah sama dengan probabilitas genotipe pada awal generasi. Sebaliknya, jika ada keterikatan antara dua lokus maka probabilitas genotipenya tidak sama untuk setiap generasi (Wijayanto et al. 2013).
12
Rekombinasi terjadi selama meiosis sebagai pindah silang antara kromosom yang berpasangan. Hasil genotipe dari Populasi DCF1 Ciherang homozigot tidak mengikuti 25% Code, 25% Angke, 25% Ciherang, 25% IRBB21 artinya pada populasi DCF1 ini telah terjadi pindah silang yang ditandai dengan tetua Angke sebagai donor xa5 pada DCF1 Ciherang tersisipkan sebesar 77.67% dari total individu heterozigot. Code sebagai donor Xa7 pada DCF1 Ciherang tersisipkan dari total individu yang heterozigot sebesar 82.02%. IRBB21 sebagai donor Xa21 untuk DCF1 Ciherang tersisipkan 100% dari total individu yang heterozigot. Menurut BB-Biogen (2007), varietas Angke dan Code dibentuk dengan metode silang balik dengan IR64 sebagai tetua berulang dan IRBB5 dan IRBB7 sebagai tetua donor masing-masing untuk gen ketahanan terhadap HDB xa5 dan Xa7, sementara donor gen Xa21 diperoleh dari galur isogenik IRBB21.
Karakter Agronomi Hasil Panen
Karakter agronomi hasil panen merupakan indikator dari pencapaian tujuan seleksi untuk meningkatkan produktivitas tanaman padi yang dinyatakan dengan indeks panen. Indeks panen dipengaruhi oleh keragaman genetik. Keragaman genetik pada tanaman menyerbuk silang umumnya cukup besar dibandingkan dengan tanaman menyerbuk sendiri sehingga dalam menentukan kriteria seleksi diutamakan pada sifat ekonomi, terutama bobot gabah isi. Komponen hasil dapat diduga melalui karakter lain yang berkorelasi dengan karakter hasil yang dinamakan seleksi tidak langsung (Rachmadi 2000).
Pertumbuhan, perkembangan, dan hasil produksi tanaman ditentukan oleh dua faktor yaitu genetik dan lingkungan. Faktor genetik dalam hal ini berupa rekombinasi genetik yang merupakan proses pertukaran elemen genetik yang dapat terjadi antara untaian DNA yang berlainan (interstrand), atau antara bagian-bagian gen yang terletak dalam satu untaian DNA (intrastrand). Rekombinasi genetik berkontribusi terhadap perbedaan genetik pada populasi.
Faktor lingkungan yang sangat berperan dalam karakter agronomi tanaman yaitu tersedianya unsur hara. Siklus nitrogen merupakan salah satu siklus hara paling penting karena digunakan dalam asimilasi nitrat membentuk asam amino. Reduksi nitrat berlangsung di sitosol, enzim yang mengkatalisis adalah nitrat reduktase, enzim ini memindahkan dua elektron dari NADPH2, hasilnya adalah
nitrit, NAD (NADP) dan H2O. Nitrat reduktase adalah suatu enzim besar dan
kompleks yang terdiri dari FAD, satu sitokrom, dan Molibdenum (Mo) yang semuanya akan tereduksi dan teroksidasi pada waktu elektron diangkut dari NADH2 ke atom nitrogen dalam NO3. Reduksi nitrit berlangsung di kloroplas atau
13
tetuanya dimungkinkan karena adanya pengaruh efisiensi penggunaan nitrogen dalam bentuk nitrat oleh padi tersebut dalam membentuk asam amino sehingga produktivitasnya berbeda.
Bobot gabah dan jumlah malai berkorelasi positif dengan jumlah anakan dan ukuran malai, dimana pertumbuhannya dipengaruhi oleh pemupukan sebagai penyedia unsur hara dan cahaya matahari untuk proses fotosintesis. Fotosintesis terjadi pada reaksi gelap dan reaksi terang. Klorofil dan pigmen lain pada sel fotosintetik akan menyerap energi cahaya pada reaksi terang dan mengubahnya menjadi ATP (adenosin trifosfat) dan NADPH (nikotinamida adenin dinukleotida fosfat), bersamaan dengan itu oksigen ikut dibebaskan. Senyawa berenergi tinggi (ATP dan NADPH) yang dibentuk pada reaksi terang digunakan untuk mereduksi karbon dioksida membentuk glukosa. Karbon dioksida juga dapat dirubah menjadi pati yang merupakan polimer dari glukosa melalui jalur asimilasi CO2 di dalam
kloroplas. Pati yang dihasilkan dapat dirubah kembali menjadi triosa fosfat yang kemudian dapat dikeluarkan sebagai sukrosa dan disimpan dalam vakuola sebagai cadangan makanan berupa biji. Pati merupakan penyusun utama endosperm atau bagian utama dari bulir beras. Sintesis pati terjadi di kloroplas dengan substrat triosa fosfat, disintesis melalui ADP-glukosa ketika konsentrasi fosfat anorganik (Pi) di sitosol rendah. Sementara itu, sintesis sukrosa melalui UDP-glukosa terjadi di sitosol ketika konsentrasi Pi tinggi (Lehninger et al. 2008).
Representasi Data Genotipe
Data genotipe merupakan representasi hasil skoring pita DNA empat gen ketahanan yang diidentifikasi pada 3 kromosom yaitu kromosom 5, 6, dan 11. Kromosom tersebut berasosiasi dengan penanda molekuler gen ketahanan HDB pada posisi rekombinasi tertentu. Posisi rekombinasi diukur berdasarkan jarak relatif antara dua gen atau lebih. Gen Xa4 terletak pada kromosom 11 (Suh et al. 2013). Xa4 adalah gen hasil eksploitasi dari generasi turunan padi yang paling banyak dimanfaatkan dalam program pemuliaan karena memberikan resistensi dalam jangka waktu yang lama pada kultivar padi komersial (Mew et al. 1992).
Gen resesif xa5 sangat efektif tahan terhadap strain Xoo, sehingga gen ini potensial digunakan dalam penanggulangan penyakit hawar daun bakteri di Indonesia (Yunus 1998). Gen xa5 terletak pada lengan pendek kromosom 5 dalam suatu daerah 0.5 cM, sekitar 70 kb, yang diapit oleh SNP marker RS7 dan SSR marker RM611 (Sidhu et al. 1978; Blair et al. 2003). Pada penelitian ini, gen xa5 berasosiasi dengan penanda RM611 pada posisi rekombinasi 31 cM. Gen ini dapat membedakan alel rentan melalui satu kodon, terdiri dari 4 ekson dan 3 intron, mengkode subunit gamma dari faktor transkripsi eukariotik (TFIIAγ) yang mengandung 106 asam amino (Hampsey 1998).
Gen ketahanan Xa7 sangat penting dalam mengendalikan hawar daun bakteri karena memiliki resistensi yang cukup lama (durable) disebabkan gen Xa7 memiliki protein mirip dengan gen ketahanan HDB lainnya, termasuk Xa1, Xa3, Xa13, Xa21, Xa26, dan Xa27 (Iyer dan McCouch 2004; Chen et al. 2008). Xa7 pada genom padi terdapat pada kromosom 6 yang dalam penelitian ini berasosiasi dengan penanda RM20582 pada posisi rekombinasi 109.5 cM.
14
Wang et al. (1996) yaitu gen Xa21. Gen ini telah diidentifikasi oleh Khush et al. (1990) dari Oryza longistamina dan diisolasi dari individu isogenik IRBB21 oleh Song et al. (1995) menggunakan kolonisasi gen berdasarkan pemetaan. Gen Xa21 pada penelitian ini terpetakan pada kromosom 11 berpaut di posisi 95.95 cM menggunakan penanda pTA248.
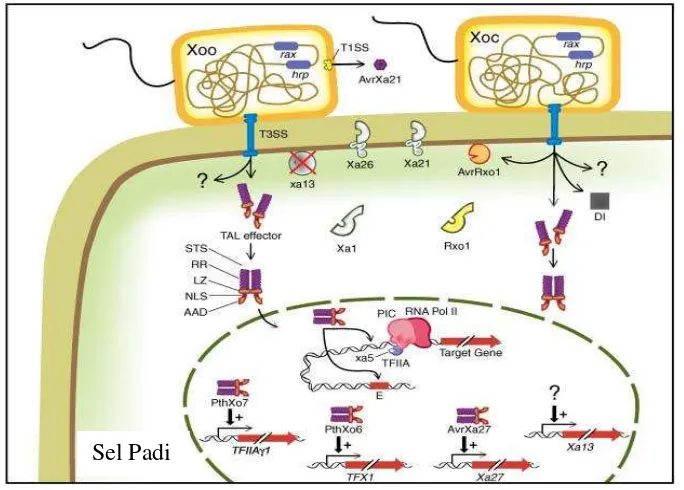
Reaksi tanaman padi terhadap infeksi patogen Xoo sangat bervariasi. Varietas padi yang telah memiliki pyramiding gen ketahanan akan cenderung lebih resisten dan tidak mudah dipatahkan ketahanannya dibandingkan dengan varietas yang hanya memiliki satu gen ketahanan, karena pyramiding akan menghasilkan reaksi berantai sebagai bentuk pertahanan tanaman padi sesuai konsep gen ke gen. Gen ketahanan pada tanaman padi yang berperan pertama kali menangkal serangan patogen Xoo adalah gen Xa21. Gen ini mengkode protein reseptor kinase (RLK) yang berperan dalam sejumlah lintasan sinyal pada tanaman, termasuk kekebalan alami (Morillo dan Tax 2006). RLK memiliki struktur multidomain, termasuk domain ekstraseluler Leucine rich repeat (LRR) yang dapat mengenali patogen Xoo. Ketika pertahanan awal tersebut mampu ditembus oleh patogen Xoo, maka diperlukan gen ketahanan lain seperti halnya xa5 yang terdapat pada inti sel tanaman. Gen xa5 dalam interaksinya dengan bakteri Xoo akan menghasilkan protein termodifikasi dari penyandi subunit transkripsi pada preinitiation complex (PIC), sehingga dapat menurunkan avirulensi xa5 (Liu et al. 2006).
Virulensi Xoo umumnya dipengaruhi oleh dua gen, yaitu gen rax dan hrp (Liu et al. 2006). rax akan menyandikan protein sekresi tipe 1 yaitu T1SS (type 1 secretion system). Elisitor (protein virulen Xoo) yang melalui sekresi tersebut adalah avrXa21, avrXa26, avrXa1, avrxa5, dan avrxa13 (Burdman et al. 2004). Sementara hrp (hypersensitivity reaction and for pathogenesis) akan menyandikan sistem sekresi T3SS (type III secretion system yang bersifat hidrofobik). Elisitor Xoo yang melalui sekresi tersebut adalah avrXa2, avrXa7, avrXa10 (Schornack et al. 2006). Elisitor-elisitor tersebut disandikan oleh kelompok gen AvrBs3/PthA seperti gen PthXo06, PthXo07 yang menyandikan protein serupa transcription activator-like (TAL) yang mempengaruhi proses transkripsi (Liu et al. 2006).
Keterkaitan gen resisten pada tanaman padi dan efektor bakteri Xoo diawali dengan diaktifkannya gen Xa21 dalam apoplast dengan avrXa21 dari hrp bakteri Xanthomonas oryzae pv. oryzae. avrXa21 adalah gen yang tergantung pada molekul rax yang dibawa oleh patogen. Termasuk gen dengan peran yang diduga dalam regulasi gen, seperti peptida sulfat dan T1SS. Sementara sinyal avirulen untuk xa1 dan xa5 belum teridentifikasi (Liu et al. 2006). Kemudian didalam sel tumbuhan, TAL efektor didimerisasi dan diangkut ke inti yang selanjutnya menunjukkan mungkin gen xa5 menonaktifkan virulensi TAL efektor dengan mengganggu atau memodifikasi interaksi TAL dan promotor DNA target dengan PIC (Gambar 7). avrXa7 ditunjukkan untuk mengikat double strand DNA (Yang et al. 2000). Gen avrXa7 merupakan anggota AvrBs3/PthA dan berisi urutan sinyal lokalisasi inti dan aktivasi domain asam (AAD) di ujung 3’ dan wilayah tengah yang berisi sekuen berulang, dimana keduanya penting bagi avirulensi dan agresifitas (Ponciano 2004). Garis keturunan yang mengandung gen Xa7 tidak bertahan dalam waktu lebih dari 2 tahun.
15
dilakukan dengan kombinasi gen ketahanan yang tepat. Hal ini sesuai dengan penelitian Susanto dan Sudir (2012), dimana padi IRBB64 yang memiliki gen ketahanan Xa4+xa5+Xa7+Xa21 dapat dijadikan sebagai sumber ketahanan terhadap patotipe III, namun ketika kombinasi Xa4+xa5 dengan gen lain seperti xa13 terhadap strain III menjadi agak rentan. Begitu juga dengan kombinasi Xa4 dan xa5 dengan Xa21 tanpa Xa7 tidak mampu memberikan ketahanan terhadap HDB strain III. Strain III merupakan salah satu patotipe Xoo yang dominan pada sentra produksi padi di Jawa dengan komposisi 23.5% (Sudir et al. 2009).
Gambar 7 Keterkaitan gen resisten HDB dan bakteri Xoo (Liu et al. 2006)
SIMPULAN DAN SARAN
Simpulan
Deteksi gen ketahanan Xa4, xa5, Xa7, dan Xa21 menggunakan marka SSR dan STS pada 207 individu DCF1 Ciherang secara pyramiding gen diperoleh 4 individu memiliki gen Xa4+xa5+Xa7+Xa21, 11 individu memiliki gen xa5+Xa7+Xa21, 46 individu memiliki gen xa5+Xa21, 31 individu memiliki gen Xa7+Xa21, dan 45 individu memiliki gen xa5+Xa7. Gen Xa4 teramplifikasi pada 150 bp dengan primer MP1+MP2. Gen xa5 teramplifikasi pada 213 bp menggunakan primer RM611. Gen Xa7 teramplifikasi pada 83 bp dengan primer RM20582. Gen Xa21 teramplifikasi pada 1000 bp menggunakan primer pTA248. Karakter agronomi jumlah malai DCF2 Ciherang tidak berbeda nyata dengan tetuanya. Panjang malai DCF2 Ciherang tidak berbeda nyata dengan tetua Code dan IRBB21, serta tidak berbeda signifikan dengan tetua Angke dan Ciherang. Bobot gabah isi DCF2 Ciherang lebih besar secara nyata dibandingkan tetuanya. Bobot gabah hampa DCF2 Ciherang juga berbeda nyata dengan tetuanya, lebih besar dari tetua Code, Angke, Ciherang, dan IRBB21.
16
Saran
Memperbesar jumlah individu dalam populasi pada penelitian gen ketahanan terhadap hawar daun bakteri berbasis pyramiding gen pada individu DCF1 Ciherang, sehingga dapat memperbesar peluang mendapatkan individu yang memiliki keempat gen ketahanan dengan produktivitas yang tetap tinggi. Selain itu, diperlukan deteksi gen ketahanan berbasis marka molekuler yang berkelanjutan pada generasi kedua dan generasi seterusnya sebagai konfirmasi bahwa keberadaan pyramiding gen ketahanan hawar daun bakteri tetap ada dalam padi hasil silang ganda Ciherang pada setiap generasinya.
DAFTAR PUSTAKA
Azrai M. 2005. Pemanfaatan markah molekuler dalam proses seleksi pemuliaan tanaman. J Agro Biogen 1(1): 26-37. 2003. High resolution genetic mapping and candidate gene identification at the xa5 locus for bacterial blight resistance in rice (Oryza sativa L.). Theor. Appl. Genet. 107:62-73.
Burdman S, Shen Y, Lee SW, Xue Q, Ronald P. 2004. RaxH/RaxR: a two-component regulatory system in Xanthomonas oryzae pv. oryzae required for AvrXa21 activity. Mol Plant Microbe Interact. 17:602-612.
Chen S, Huang Z, Zeng L, Yang J, Liu Q, Zhu X. 2008. High-resolution mapping and gene prediction of Xanthomonas oryzae pv. oryzae resistance gene Xa7. Mol Breed. 22:433–441.
Chun et al. 2012. Identification, mapping, isolation of the genes resisting to bacterial blight and breeding application in rice. Molecular Plant Breeding. 13(12):121-123.
Dellaporta Sl, Jonathan W, James BH. 1983. A plant DNA minipreapration: version II. Plant molecular Biology Reporter. 1(4):19-21.
[Dinas Pertanian dan Tanaman Pangan Propinsi Jawa Barat]. 2013. Penyebaran Varietas Padi 2010. Diperta [Internet]. [diunduh 26 Desember 2013]. Tersedia pada: http://diperta.jabarprov.go.id/index.php/subMenu/931. [Direktorat Perlindungan Tanaman Pangan]. 2011. Penyakit Bakteri pada padi.
Litbang [Internet]. [diunduh 17 September 2013]. Tersedia pada: http://pangan.litbang.deptan.go.id/berita/penyakit-bakteri-pada-padi.
Hampsey M. 1998. Molecular genetics of the RNA polymerase II general transcriptional machinery. Microbiol. Mol. Biol. Rev. 62:465–503.
Iyer AS, McCouch SR. 2004. The rice bacterial blight resistance gene xa5 encodes a novel form of disease resistance. Mol Plant Microbe Interact. 17:1348–1354.
17
Kementrian Pertanian. 2012. Indonesia Butuh Tambahan Tujuh Juta Ton Beras. Mentan [Internet]. [diunduh 26 Desember 2013]. Tersedia pada: http://www.antaranews.com/berita/341347/indonesia-butuh-tambahan tujuh-juta-ton-beras.
Khush GS, Bacalangco E and Ogawa T. 1990. A new gene for resistance to bacterial blight from O. longistaminata. Rice Genet Newslett. 7:121-122. Lehninger AL, David LN, Michael MC. 2008. Principles of Biochemistry 5th
edition. New York (US): W.H. Freeman and Company.
Liu DON, Pamela CR, Adam JB. 2006. Xanthomonas oryzae pathovars: model pathogen of a model crop. Mol Plant Path. 7:303-324.
Ma BJ, Wang WM, Zhao B, Zhou YL. 1999. Studies of PCR markers for the rice bacterial blight resistance gene Xa-4. Hereditas 21: 9-12.
Marella LS, Vera CM, George LC, Bernardo MA, Nelson RJ, Lenug H, Reddy JN, Sridhar R. 2001. Idenification of resistance genes effective against rice bacterial bight pathogen in Eastern India. Plant Dis. 85(5): 506-512.
Morillo SA, Tax FE. 2006. Functional analysis of receptor-like kinases in monocots and dicots. Curr Opin Plant Biol. 9:460–469.
Mew TW, Vera Cruz CM, and Medalla ES. 1992. Changes in race frequency of Xanthomonas oryzae pv. oryzae in response to rice cultivars planted in the Philippines. Plant Dis. 76: 1029–1032.
Muliis KB, Faloona FA. 1987. Specific synthesis of DNA in vitro by a polymerase catalysed chain reaction. Enzymology. 155:335-350.
Ponciano G, Webb K, Bai J, Cruz CV, Leach JE. 2004. Molecular characterization of the avrXa7 locus from Xanthomonas oryzae pv. oryzae field isolates. Physiol. Mol. Plant Pathol. 64:145–153.
Prihatin I, Taryono, Rimbawanto A. 2006. Penggunaan penanda mikrosatelit untuk analisis induk Acacia manginum WILD. UGM, Pusat Litbang Hutan Tanaman. 3:139-148.
Pusat Penelitian Pengembangan Tanaman Pangan. 2006. Padi Ciherang Makin popular. Warta Penelitian dan Pengembangan Pertanian Indonesia 28:2. Rachmadi M. 2000. Pengantar Pemuliaan Tanaman Membiak Vegetatif. Bandung
[ID]: Universitas Padjajaran.
Sambrook J, DW Russell. 2001. Molecular Cloning: A Laboratory Manual. Edisi 3. New York (US): Coldspring Harbor Laboratory Pr.
Schornack S, Meyer A, Romer P, Jordan T, Lahaye T. 2006. Gene for gene mediated recognition of nuclear target AvrBs3-like bacterial efector proteins. J Plant Physiol. 163:256-272.
Sidhu GS, Khush GS, and Mew TW. 1978. Genetic analysis of bacterial blight resistance in seventy-four cultivars of rice (Oryza sativa L.). Theor Appl Genet. 53:105-111.
Simanjuntak BH, Suprihatin MP, Isjwara MR. 2000. Pengaruh perbandingan nitrat dan amonium terhadap pertumbuhan dan hasil tanaman selada (Lactusa sativa L.) yang dibudidayakan secara hidroponik. Prosiding. Seminar Nasional Pengembangan Teknologi Hortikultura Memasuki Indonesia Baru. ISBN:979-9458-88-9
18
Susanto U, Sudir. 2012. Ketahanan genotipe padi terhadap Xanthomonas oryzae pv. oryzae patotipe III, IV, dan VIII. Penelitian Pertanian Tanaman Pangan. 31(2):108-116.
Gopalakrishnan S, Sharma RK, Anand K, Joseph M, Singh VP, Singh AK, Bhat KV, Singh NK, Mohapatra T. 2008. Integrating marker assisted background analysis with foreground selection for identification of superior bacterial blight resistant recombinants in Basmati rice. Plant Breed. 27: 131-139. Singh S, JS Sidhu, N Huang, Y Vikal, Z Li, DS Brar, HS Dhaliwal, GS Khush.
2001. Pyramiding three bacterial blight resistance genes (xa5, xa13, and Xa21) using marker-assisted selection into indica rice cultivar PR106. Theor Appl Genet. 102:1011-1015.
Song W, Wang G, Chen L, Kim H, Pi L, Holsten T, Gardner J, Wang B, Zhai W, Zhu L. 1995. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science. 270:1804–1806.
Suh et al. 2013. Development of breeding lines with three pyramided resistance genes that confer broad-spectrum bacterial blight resistance and their molecular analysis in rice. Rice. 6:5.
Thermo Fisher Scientific. 2009. Nanodrop 2000/200c Spectrpphotometer V1.0 User Manual. Wilmington (US): Thermo Fischer Scientific.
Wang G, Song W, Ruan D, Sideris S, Ronald PC. 1996. The cloned gene, Xa21, confers resistance to multiple Xanthomonas oryzae pv. oryzae isolates in transgenic plants. Mol Plant Microbe Interact. 9:850–855.
Wijayanto DA, Hidayat R, Hasan M. 2013. Application of difference equations model in determining genotype probability offspring with two different charavteristic. JurnalIlmu Dasar. 14:79-84.
Yang Z, X Sun, S Wang, and Q Zhang. 2003. Genetic and physical mapping of a new gene for bacterial blight resistance in rice. Theor. Appl. Genet 106: 1467−1472.
Yang B, Zhu W, Johnson LB, and White FF. 2000. The virulence factor AvrXa7 of Xanthomonas oryzae pv. oryzae is a type III secretion pathway-dependent nuclear-localized double-stranded DNA-binding protein. Proc. Natl Acad. Sci. 97: 9807–9812.
19
LAMPIRAN
Lampiran 1 Diagram alir penelitian
Lampiran 2 Diagram persilangan
Pemilihan benih DCF1 Ciherang yang akan ditanam
Persiapan media dan penyemaian benih padi pada cawan petri
Perawatan benih padi yang dikecambah selama 1-2 minggu
Pemindahan tanaman padi pada media sesuai urutan nomor
Perawatan tanaman padi ± 1 bulan
Identifikasi gen ketahanan Xa4, xa5, Xa7, dan Xa21
Isolasi DNA daun padi
Analisis data molekuler: nomor tanaman yang memiliki 4 gen dan Graphical Genotyping
Amplifikasi DNA dengan PCR
Elektroforesis dengan agarose 2% dan PAGE, dianalisis elektroforegramnya menggunakan Chemidoc
Evaluasi karakter agronomi
20
Lampiran 3 Elektroforegram hasil amplifikasi gen Xa4 dengan primer MP1+MP2
Lampiran 4 Elektroforegram hasil amplifikasi gen xa5 dengan primer RM611
Lampiran 5 Elektroforegram hasil amplifikasi gen Xa7 dengan primer RM20582
21
Lampiran 7 Hasil skoring elektroforegram DCF1 Ciherang dan tetuanya
Nama Galur/individu Kode Primer
RM611 (xa5) RM20582 (Xa7) pTA248 (Xa21) MP1+MP2 (Xa4)
Code Cd Cih Cd Cd Cd
Angke AK Ak Ak Ak Ak
Ciherang Cih Cih Cih Cih Cih
IRBB21 I21 Cih I21 I21 I21
DCF1 CiherangxIRBB21//Code X Angke-1 DCF1Ch|CA-1 Ak Cd - -
DCF1 CiherangxIRBB21//Code X Angke-2 DCF1Ch|CA-2 H Ak - -
DCF1 CiherangxIRBB21//Code X Angke-3 DCF1Ch|CA-3 Cih Cd H -
DCF1 CiherangxIRBB21//Code X Angke-4 DCF1Ch|CA-4 H H - -
DCF1 CiherangxIRBB21//Code X Angke-5 DCF1Ch|CA-5 H H - -
DCF1 CiherangxIRBB21//Code X Angke-6 DCF1Ch|CA-6 H Cih - -
DCF1 CiherangxIRBB21//Code X Angke-7 DCF1Ch|CA-7 H H - -
DCF1 CiherangxIRBB21//Code X Angke-8 DCF1Ch|CA-8 H H H -
DCF1 CiherangxIRBB21//Code X Angke-9 DCF1Ch|CA-9 Cd Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-10 DCF1Ch|CA-10 Cd Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-11 DCF1Ch|CA-11 H H Cih -
DCF1 CiherangxIRBB21//Code X Angke-12 DCF1Ch|CA-12 H H Cih -
DCF1 CiherangxIRBB21//Code X Angke-13 DCF1Ch|CA-13 Ak Cd H -
DCF1 CiherangxIRBB21//Code X Angke-14 DCF1Ch|CA-14 Cd H H -
DCF1 CiherangxIRBB21//Code X Angke-15 DCF1Ch|CA-15 Ak Cih H -
DCF1 CiherangxIRBB21//Code X Angke-16 DCF1Ch|CA-16 Cd H H -
DCF1 CiherangxIRBB21//Code X Angke-17 DCF1Ch|CA-17 Cih H H -
DCF1 CiherangxIRBB21//Code X Angke-18 DCF1Ch|CA-18 H H Cih
DCF1 CiherangxIRBB21//Code X Angke-19 DCF1Ch|CA-19 Cd Cd H -
DCF1 CiherangxIRBB21//Code X Angke-20 DCF1Ch|CA-20 H H Cih -
DCF1 CiherangxIRBB21//Code X Angke-21 DCF1Ch|CA-21 Cih H H -
DCF1 CiherangxIRBB21//Code X Angke-22 DCF1Ch|CA-22 Cd H H -
DCF1 CiherangxIRBB21//Code X Angke-23 DCF1Ch|CA-23 Cd Ak H -
DCF1 CiherangxIRBB21//Code X Angke-24 DCF1Ch|CA-24 H Ak H -
DCF1 CiherangxIRBB21//Code X Angke-25 DCF1Ch|CA-25 H Cd H -
DCF1 CiherangxIRBB21//Code X Angke-26 DCF1Ch|CA-26 Cih H H -
22
(lanjutan dari lampiran 7)
Nama Individu Kode Primer
RM611 (xa5) RM20582 (Xa7) pTA248 (Xa21) MP1+MP2 (Xa4)
DCF1 CiherangxIRBB21//Code X Angke-28 DCF1Ch|CA-28 H Cih H -
DCF1 CiherangxIRBB21//Code X Angke-29 DCF1Ch|CA-29 Ak Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-30 DCF1Ch|CA-30 H H - -
DCF1 CiherangxIRBB21//Code X Angke-31 DCF1Ch|CA-31 Cih Cih - -
DCF1 CiherangxIRBB21//Code X Angke-32 DCF1Ch|CA-32 Cd H H -
DCF1 CiherangxIRBB21//Code X Angke-33 DCF1Ch|CA-33 H H H -
DCF1 CiherangxIRBB21//Code X Angke-34 DCF1Ch|CA-34 Cd Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-35 DCF1Ch|CA-35 Cih Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-36 DCF1Ch|CA-36 H H - -
DCF1 CiherangxIRBB21//Code X Angke-37 DCF1Ch|CA-37 Cih H Cih -
DCF1 CiherangxIRBB21//Code X Angke-38 DCF1Ch|CA-38 Cd Ak H -
DCF1 CiherangxIRBB21//Code X Angke-39 DCF1Ch|CA-39 Cd Cd Cih -
DCF1 CiherangxIRBB21//Code X Angke-40 DCF1Ch|CA-40 Ak Cd Cih -
DCF1 CiherangxIRBB21//Code X Angke-41 DCF1Ch|CA-41 Cd H Cih -
DCF1 CiherangxIRBB21//Code X Angke-42 DCF1Ch|CA-42 H H H H
DCF1 CiherangxIRBB21//Code X Angke-43 DCF1Ch|CA-43 H H Cih -
DCF1 CiherangxIRBB21//Code X Angke-44 DCF1Ch|CA-44 H Cd Cih -
DCF1 CiherangxIRBB21//Code X Angke-45 DCF1Ch|CA-45 H Cd Cih -
DCF1 CiherangxIRBB21//Code X Angke-46 DCF1Ch|CA-46 H H - -
DCF1 CiherangxIRBB21//Code X Angke-47 DCF1Ch|CA-47 Cih H Cih -
DCF1 CiherangxIRBB21//Code X Angke-48 DCF1Ch|CA-48 H Cd Cih -
DCF1 CiherangxIRBB21//Code X Angke-49 DCF1Ch|CA-49 H H Cih -
DCF1 CiherangxIRBB21//Code X Angke-50 DCF1Ch|CA-50 H Cd Cih -
DCF1 CiherangxIRBB21//Code X Angke-51 DCF1Ch|CA-51 H Cd - -
DCF1 CiherangxIRBB21//Code X Angke-52 DCF1Ch|CA-52 H Cd Cih -
DCF1 CiherangxIRBB21//Code X Angke-53 DCF1Ch|CA-53 Cih Cd - -
DCF1 CiherangxIRBB21//Code X Angke-54 DCF1Ch|CA-54 H Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-55 DCF1Ch|CA-55 Cd H Cih -
DCF1 CiherangxIRBB21//Code X Angke-56 DCF1Ch|CA-56 Cd Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-57 DCF1Ch|CA-57 H Cih Cih -
23
(lanjutan dari lampiran 7)
Nama Individu Kode Primer
RM611 (xa5) RM20582 (Xa7) pTA248 (Xa21) MP1+MP2 (Xa4)
DCF1 CiherangxIRBB21//Code X Angke-59 DCF1Ch|CA-59 Cih H Cih -
DCF1 CiherangxIRBB21//Code X Angke-60 DCF1Ch|CA-60 H H Cih -
DCF1 CiherangxIRBB21//Code X Angke-61 DCF1Ch|CA-61 Cih Cd Cih -
DCF1 CiherangxIRBB21//Code X Angke-62 DCF1Ch|CA-62 H Cd Cih -
DCF1 CiherangxIRBB21//Code X Angke-63 DCF1Ch|CA-63 Cih Cd Cih -
DCF1 CiherangxIRBB21//Code X Angke-64 DCF1Ch|CA-64 H Cd Cih -
DCF1 CiherangxIRBB21//Code X Angke-65 DCF1Ch|CA-65 Cih Cih H -
DCF1 CiherangxIRBB21//Code X Angke-66 DCF1Ch|CA-66 H Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-67 DCF1Ch|CA-67 Cih Ak Cih -
DCF1 CiherangxIRBB21//Code X Angke-68 DCF1Ch|CA-68 Cih Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-69 DCF1Ch|CA-69 H H H H
DCF1 CiherangxIRBB21//Code X Angke-70 DCF1Ch|CA-70 Ak H H -
DCF1 CiherangxIRBB21//Code X Angke-71 DCF1Ch|CA-71 H H Cih Cd
DCF1 CiherangxIRBB21//Code X Angke-72 DCF1Ch|CA-72 H H Cih -
DCF1 CiherangxIRBB21//Code X Angke-73 DCF1Ch|CA-73 Ak Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-74 DCF1Ch|CA-74 H Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-75 DCF1Ch|CA-75 H H - -
DCF1 CiherangxIRBB21//Code X Angke-76 DCF1Ch|CA-76 H Cih - -
DCF1 CiherangxIRBB21//Code X Angke-77 DCF1Ch|CA-77 H Cd H Cih
DCF1 CiherangxIRBB21//Code X Angke-78 DCF1Ch|CA-78 H H Cih -
DCF1 CiherangxIRBB21//Code X Angke-79 DCF1Ch|CA-79 H Cd Cih -
DCF1 CiherangxIRBB21//Code X Angke-80 DCF1Ch|CA-80 H H Cih -
DCF1 CiherangxIRBB21//Code X Angke-81 DCF1Ch|CA-81 H Cd H -
DCF1 CiherangxIRBB21//Code X Angke-82 DCF1Ch|CA-82 H Cih H -
DCF1 CiherangxIRBB21//Code X Angke-83 DCF1Ch|CA-83 H Cih H -
DCF1 CiherangxIRBB21//Code X Angke-84 DCF1Ch|CA-84 Cih Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-85 DCF1Ch|CA-85 Cih Cih H -
DCF1 CiherangxIRBB21//Code X Angke-86 DCF1Ch|CA-86 H H Cih -
DCF1 CiherangxIRBB21//Code X Angke-87 DCF1Ch|CA-87 Ak Cih H -
DCF1 CiherangxIRBB21//Code X Angke-88 DCF1Ch|CA-88 Cih H H -
24
(lanjutan dari lampiran 7)
Nama Individu Kode Primer
RM611 (xa5) RM20582 (Xa7) pTA248 (Xa21) MP1+MP2 (Xa4)
DCF1 CiherangxIRBB21//Code X Angke-90 DCF1Ch|CA-90 H H Cih -
DCF1 CiherangxIRBB21//Code X Angke-91 DCF1Ch|CA-91 Cih H Cih -
DCF1 CiherangxIRBB21//Code X Angke-92 DCF1Ch|CA-92 H H Cih -
DCF1 CiherangxIRBB21//Code X Angke-93 DCF1Ch|CA-93 H H H -
DCF1 CiherangxIRBB21//Code X Angke-94 DCF1Ch|CA-94 Ak H H -
DCF1 CiherangxIRBB21//Code X Angke-95 DCF1Ch|CA-95 Cih Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-96 DCF1Ch|CA-96 Cih Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-97 DCF1Ch|CA-97 H Cih H -
DCF1 CiherangxIRBB21//Code X Angke-98 DCF1Ch|CA-98 H Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-99 DCF1Ch|CA-99 H Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-100 DCF1Ch|CA-100 - Ak - -
DCF1 CiherangxIRBB21//Code X Angke-101 DCF1Ch|CA-101 Cih Cih H -
DCF1 CiherangxIRBB21//Code X Angke-102 DCF1Ch|CA-102 Cih Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-103 DCF1Ch|CA-103 Cih Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-104 DCF1Ch|CA-104 Ak Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-105 DCF1Ch|CA-105 - H Cih -
DCF1 CiherangxIRBB21//Code X Angke-106 DCF1Ch|CA-106 Cih H Cih -
DCF1 CiherangxIRBB21//Code X Angke-107 DCF1Ch|CA-107 Cih Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-108 DCF1Ch|CA-108 Ak Cd Cih -
DCF1 CiherangxIRBB21//Code X Angke-109 DCF1Ch|CA-109 Cih Cd Cih -
DCF1 CiherangxIRBB21//Code X Angke-110 DCF1Ch|CA-110 Cih Cd H -
DCF1 CiherangxIRBB21//Code X Angke-111 DCF1Ch|CA-111 Cih Cih H -
DCF1 CiherangxIRBB21//Code X Angke-112 DCF1Ch|CA-112 H Cih H -
DCF1 CiherangxIRBB21//Code X Angke-113 DCF1Ch|CA-113 Cih Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-114 DCF1Ch|CA-114 H Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-115 DCF1Ch|CA-115 Cih Cih H -
DCF1 CiherangxIRBB21//Code X Angke-116 DCF1Ch|CA-116 H Ak H -
DCF1 CiherangxIRBB21//Code X Angke-117 DCF1Ch|CA-117 H Cd H -
DCF1 CiherangxIRBB21//Code X Angke-118 DCF1Ch|CA-118 H H Cih -
DCF1 CiherangxIRBB21//Code X Angke-119 DCF1Ch|CA-119 Cih Cih Cih -
25
(lanjutan dari lampiran 7)
Nama Individu Kode Primer
RM611 (xa5) RM20582 (Xa7) pTA248 (Xa21) MP1+MP2 (Xa4)
DCF1 CiherangxIRBB21//Code X Angke-121 DCF1Ch|CA-121 Cih H Cih -
DCF1 CiherangxIRBB21//Code X Angke-122 DCF1Ch|CA-122 H Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-123 DCF1Ch|CA-123 H Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-124 DCF1Ch|CA-124 Cih H H -
DCF1 CiherangxIRBB21//Code X Angke-125 DCF1Ch|CA-125 Ak Cih H -
DCF1 CiherangxIRBB21//Code X Angke-126 DCF1Ch|CA-126 Cih Cd Cih -
DCF1 CiherangxIRBB21//Code X Angke-127 DCF1Ch|CA-127 Cih H H -
DCF1 CiherangxIRBB21//Code X Angke-128 DCF1Ch|CA-128 H Ak - -
DCF1 CiherangxIRBB21//Code X Angke-129 DCF1Ch|CA-129 H Cd Cih -
DCF1 CiherangxIRBB21//Code X Angke-130 DCF1Ch|CA-130 Cih Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-131 DCF1Ch|CA-131 H Cd Cih -
DCF1 CiherangxIRBB21//Code X Angke-132 DCF1Ch|CA-132 H Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-133 DCF1Ch|CA-133 Cih Cd H -
DCF1 CiherangxIRBB21//Code X Angke-134 DCF1Ch|CA-134 H Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-135 DCF1Ch|CA-135 H Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-136 DCF1Ch|CA-136 Cih H Cih -
DCF1 CiherangxIRBB21//Code X Angke-137 DCF1Ch|CA-137 H Cih H -
DCF1 CiherangxIRBB21//Code X Angke-138 DCF1Ch|CA-138 H Cih H -
DCF1 CiherangxIRBB21//Code X Angke-139 DCF1Ch|CA-139 Cih H Cih -
DCF1 CiherangxIRBB21//Code X Angke-140 DCF1Ch|CA-140 Ak Cih H -
DCF1 CiherangxIRBB21//Code X Angke-141 DCF1Ch|CA-141 H Cd H -
DCF1 CiherangxIRBB21//Code X Angke-142 DCF1Ch|CA-142 Cih H Cih -
DCF1 CiherangxIRBB21//Code X Angke-143 DCF1Ch|CA-143 H H H -
DCF1 CiherangxIRBB21//Code X Angke-144 DCF1Ch|CA-144 Cih Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-145 DCF1Ch|CA-145 H Ak Cih -
DCF1 CiherangxIRBB21//Code X Angke-146 DCF1Ch|CA-146 Ak H H -
DCF1 CiherangxIRBB21//Code X Angke-147 DCF1Ch|CA-147 - H Cih -
DCF1 CiherangxIRBB21//Code X Angke-148 DCF1Ch|CA-148 Cih H Cih -
DCF1 CiherangxIRBB21//Code X Angke-149 DCF1Ch|CA-149 Cih Cih H -
DCF1 CiherangxIRBB21//Code X Angke-150 DCF1Ch|CA-150 H Cih Cih -
26
(lanjutan dari lampiran 7)
Nama Individu Kode Primer
RM611 (xa5) RM20582 (Xa7) pTA248 (Xa21) MP1+MP2 (Xa4)
DCF1 CiherangxIRBB21//Code X Angke-152 DCF1Ch|CA-152 Cih H Cih -
DCF1 CiherangxIRBB21//Code X Angke-153 DCF1Ch|CA-153 Cih Cih H -
DCF1 CiherangxIRBB21//Code X Angke-154 DCF1Ch|CA-154 H Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-155 DCF1Ch|CA-155 H Cd Cih -
DCF1 CiherangxIRBB21//Code X Angke-156 DCF1Ch|CA-156 H Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-157 DCF1Ch|CA-157 H H H -
DCF1 CiherangxIRBB21//Code X Angke-158 DCF1Ch|CA-158 Ak Cd H -
DCF1 CiherangxIRBB21//Code X Angke-159 DCF1Ch|CA-159 - Cd H -
DCF1 CiherangxIRBB21//Code X Angke-160 DCF1Ch|CA-160 Cih H - -
DCF1 CiherangxIRBB21//Code X Angke-161 DCF1Ch|CA-161 H H Cih -
DCF1 CiherangxIRBB21//Code X Angke-162 DCF1Ch|CA-162 H H H H
DCF1 CiherangxIRBB21//Code X Angke-163 DCF1Ch|CA-163 H H H H
DCF1 CiherangxIRBB21//Code X Angke-164 DCF1Ch|CA-164 H H H Cih
DCF1 CiherangxIRBB21//Code X Angke-165 DCF1Ch|CA-165 Cih H H -
DCF1 CiherangxIRBB21//Code X Angke-166 DCF1Ch|CA-166 Cih H Cih -
DCF1 CiherangxIRBB21//Code X Angke-167 DCF1Ch|CA-167 Cih H Cih -
DCF1 CiherangxIRBB21//Code X Angke-168 DCF1Ch|CA-168 Cih H H -
DCF1 CiherangxIRBB21//Code X Angke-169 DCF1Ch|CA-169 H Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-170 DCF1Ch|CA-170 Cih H Cih -
DCF1 CiherangxIRBB21//Code X Angke-171 DCF1Ch|CA-171 Ak H H -
DCF1 CiherangxIRBB21//Code X Angke-172 DCF1Ch|CA-172 H Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-173 DCF1Ch|CA-173 H Cd Cih -
DCF1 CiherangxIRBB21//Code X Angke-174 DCF1Ch|CA-174 H Cih H -
DCF1 CiherangxIRBB21//Code X Angke-175 DCF1Ch|CA-175 H Cd H -
DCF1 CiherangxIRBB21//Code X Angke-176 DCF1Ch|CA-176 H H - -
DCF1 CiherangxIRBB21//Code X Angke-177 DCF1Ch|CA-177 Cih Cih I21 -
DCF1 CiherangxIRBB21//Code X Angke-178 DCF1Ch|CA-178 - H I21 -
DCF1 CiherangxIRBB21//Code X Angke-179 DCF1Ch|CA-179 H H - -
DCF1 CiherangxIRBB21//Code X Angke-180 DCF1Ch|CA-180 Cih Cih - -
DCF1 CiherangxIRBB21//Code X Angke-181 DCF1Ch|CA-181 H Ak - -
27
(lanjutan dari lampiran 7)
Nama Individu Kode Primer
RM611 (xa5) RM20582 (Xa7) pTA248 (Xa21) MP1+MP2 (Xa4)
DCF1 CiherangxIRBB21//Code X Angke-183 DCF1Ch|CA-183 Cih Cd - -
DCF1 CiherangxIRBB21//Code X Angke-184 DCF1Ch|CA-184 H H - -
DCF1 CiherangxIRBB21//Code X Angke-185 DCF1Ch|CA-185 Cih Cih - -
DCF1 CiherangxIRBB21//Code X Angke-186 DCF1Ch|CA-186 H Ak - -
DCF1 CiherangxIRBB21//Code X Angke-187 DCF1Ch|CA-187 H Cih - -
DCF1 CiherangxIRBB21//Code X Angke-188 DCF1Ch|CA-188 Cih H - -
DCF1 CiherangxIRBB21//Code X Angke-189 DCF1Ch|CA-189 H Cih - -
DCF1 CiherangxIRBB21//Code X Angke-190 DCF1Ch|CA-190 H H - -
DCF1 CiherangxIRBB21//Code X Angke-191 DCF1Ch|CA-191 Ak H - -
DCF1 CiherangxIRBB21//Code X Angke-192 DCF1Ch|CA-192 Cih Cih - -
DCF1 CiherangxIRBB21//Code X Angke-193 DCF1Ch|CA-193 Cih Cd - -
DCF1 CiherangxIRBB21//Code X Angke-194 DCF1Ch|CA-194 Cih H - -
DCF1 CiherangxIRBB21//Code X Angke-195 DCF1Ch|CA-195 H H Cih -
DCF1 CiherangxIRBB21//Code X Angke-196 DCF1Ch|CA-196 H H - -
DCF1 CiherangxIRBB21//Code X Angke-197 DCF1Ch|CA-197 H H - -
DCF1 CiherangxIRBB21//Code X Angke-198 DCF1Ch|CA-198 H Cih Cih -
DCF1 CiherangxIRBB21//Code X Angke-199 DCF1Ch|CA-199 Cih H H -
DCF1 CiherangxIRBB21//Code X Angke-200 DCF1Ch|CA-200 Cih H H Cih
DCF1 CiherangxIRBB21//Code X Angke-201 DCF1Ch|CA-201 H H H Cih
DCF1 CiherangxIRBB21//Code X Angke-202 DCF1Ch|CA-202 H H Cih -
DCF1 CiherangxIRBB21//Code X Angke-203 DCF1Ch|CA-203 Cih H H -
DCF1 CiherangxIRBB21//Code X Angke-204 DCF1Ch|CA-204 Cih H Cih -
DCF1 CiherangxIRBB21//Code X Angke-205 DCF1Ch|CA-205 Ak Cih H -
DCF1 CiherangxIRBB21//Code X Angke-206 DCF1Ch|CA-206 H H - -
DCF1 CiherangxIRBB21//Code X Angke-207 DCF1Ch|CA-207 H Cih Cih -
28
Lampiran 8 Karakter agronomi hasil panen
29
Lampiran 9 Hasil analisis sidik ragam (ANOVA) karakter agronomi hasil panen
ANOVA
30
31