www.elsevier.com / locate / bres
Short communication
Motor neuron cell death in a mouse model of FALS is not mediated
by the p53 cell survival regulator
Neurologische Klinik, Universitat des Saarlandes, 66421 Homburg /Saar, Germany
b
¨ ¨
Institut f ur Med. Biometrie, Epidemiologie und Med. Informatik, Universitat des Saarlandes, 66421 Homburg /Saar, Germany
c
¨ ¨
Institut f ur Anatomie, Universitat des Saarlandes, 66421 Homburg /Saar, Germany
d
¨ ¨
Institut f ur klinisch-experimentelle Chirurgie,Universitat des Saarlandes, 66421 Homburg /Saar, Germany
e
¨
Abteilung Virologie, Haus 47, Universitat des Saarlandes, D-66421 Homburg /Saar, Germany
Accepted 25 July 2000
Abstract
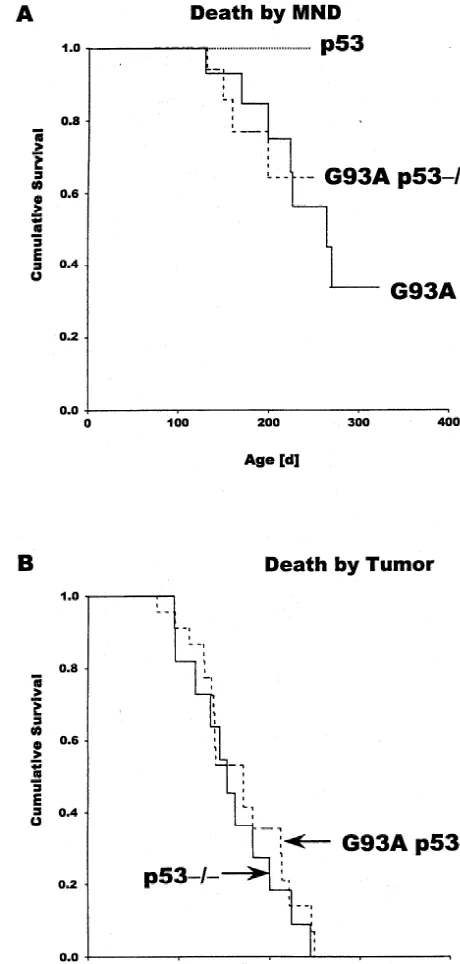
Mutant Cu / Zn superoxide dismutase (SOD1) associated with familial amyotrophic lateral sclerosis (FALS) causes selective motor neuron loss through unknown mechanisms of cell damage. Damaged neurons frequently undergo apoptosis mediated by the p53 cell survival regulator. We therefore studied whether motor neuron disease (MND) in mice expressing the human SOD1 mutant G93A is dependent on p53 by crossing G93A mice with p53-knockout mice. Since p532/2 mice’s life expectance is usually shorter (160649 days, n511) than the time at which the G93A mice die from MND (212650 days, n57), only a few of the G93A / p532/2 double transgenics were expected to live to experience MND. Nevertheless, four of the 22 G93A / p532/2 mice succumbed to MND after 160628 days, as expected under these conditions of competing death risks if the absence of p53 fails to protect from MND. Thus, MND in mice expressing G93A does not require p53. This conclusion is supported by histology: pre-symptomatic G93A mice display disease-associated vacuoles within the dendrites of motor neurons regardless of p53 status. 2000 Elsevier Science B.V. All rights reserved.
Theme: Disorders of the nervous system
Topic: Degenerative disease: other
Keywords: ALS; FALS; Apoptosis; Motor neuron disease; P53
Expression of mutated forms of the Cu / Zn superoxide transgenic mice [17], several other reports have suggested dismutase (SOD1*) is the primary cause of approximately that FALS may involve neuronal apoptosis [10,11,14,18]. 20% of FALS. While earlier work had suggested that Apoptosis as a consequence of cell damage and stress is SOD1* may cause selective motor neuron degeneration frequently mediated by the p53 tumor suppressor protein through the interference with normal cellular SOD1 func- [4] and rise in p53 level upon damage with the conse-tions or through oxidative reacconse-tions catalyzed by the quence of apoptosis is also frequently observed with mutant enzyme, more recent studies on transgenic mice neurons. For instance, accumulation of b-amyloid protein have indicated that SOD1* acts through mechanisms that in a transgenic mouse model of Alzheimer’s disease operate independently of the endogenous wild-type SOD1 correlates with p53 accumulation and nuclear DNA frag-and that might instead involve the aggregation of mis- mentation characteristic of apoptosis [15]. Along the same folded SOD1* protein (for a recent review, see [8]). line, elevated p53 levels cause apoptosis in cultured rat Although histologic studies have so far failed to demon- pyramidal hippocampal neurons [13]. Gamma-irradiation strate neurons with apoptotic morphology in SOD1* induces cerebellar granule cell death in normal but not in p53-null mice [20], and neurotoxicity of cytosine arabino-side to postmitotic sympathetic neurons [2] and MPTP *Corresponding author. Tel.:149-6841-163-983; fax:1
49-6841-163-toxicity to dopamine neurons [19] are the result, at least in 980.
E-mail address: [email protected] (K. Roemer). part, of p53-dependent apoptosis. Not least, even the death
of neurons that are regulated by the TrkA and p75 Ultrathin sections were stained with lead citrate and neurotrophin receptors during the development of the examined with a Zeiss EM 10 TEM. Two independent peripheral nervous system has been demonstrated to raters counted the numbers of vacuoles in four semithin require the presence of p53 [1]. With these observations in sections of the upper, intermediate and lower lumbar mind, we decided to study the effects of expression in region, respectively, at 20003 magnification. Four micro-transgenic mice of the Cu / Zn SOD1 mutant G93A in a scopic fields of the left and right ventral horn were
p53-null background. analyzed per semithin section.
Transgenic mice of the strain B6SJL-TgN(SOD1-dl
G93A)1Gur (stock[2300) that express ubiquitously the G93A mutant of the human Cu / Zn SOD1 were purchased from The Jackson Laboratory, Maine, USA. SOD gene status was determined on toe DNA by PCR using the primer pair 59-CATCAGCCCTAATCCATCTGA-39 and 59-CGCGACTAACAATCAAACTGA-39that amplifies, in a standard PCR cycle (958C, 30 s; 608C, 30 s; 728C, 45 s; repeated 35 times), a 236 bp product from exon 4 of the human SOD1 gene. G93A mice of stock [2300 were incapacitated by MND after 6 to 8 months rather than the published 4 to 5 months [12]. P53 knockout mice of the strain C57Bl / 6Tac BR-(KO)p53 N4 were obtained from the Bomholtgard Breeding and Research Center, Ry, Denmark. For genotyping, toe DNA was subjected to PCR analysis with a primer pair that amplifies 320 bp of the wild-type p53 gene (59 -GTGTTTCATTAGTTCCCCACC-TTGAC-39 and 59 -ATGGGAGGCTGCCAGTCCTAA-CCC-39) and, in a separate reaction, a primer pair that amplifies 280 bp of the disrupted p53 gene (59 -GTGGGA-GGGACAAAAGTTCGAGGCC-39 and 59 -TTTACGGA-GCCCTGGCGCTCGATGT-39). The PCR conditions were as described for the SOD1 primers. A standard mating strategy was applied to control for strain background effects.
Disease progresses in G93A mice from an initial and characteristic paralysis of the hindlimbs to paralysis of the forelimbs, and finally to complete paralysis. Motor neuron disease was first diagnosed as the inability of the mice to hold on to the rim of the cage with their hindlimbs. Mice unable to right themselves were killed; this time point was arbitrarily chosen as the time of death. For competing risk analysis, 14 G93A mice, 11 p53 2/2 mice, and 22 G93A / p532/2mice were followed up for the competing risk endpoints (i) death by MND (caused by the G93A genotype) and (ii) death by tumor (caused by the p532/2
genotype).
Histologic analysis was performed in a blinded study on seven littermates (age: 140 d) of three genotypes: (i) G93A; (ii) p532/2 and (iii) G93A, p532/2. Animals were anesthetized with chloralhydrate (0.4 mg / g) and transcardially perfused with a solution containing 1% formaldehyde and 1% glutaraldehyde in 0.1 M phosphate buffer pH 7.4. The spinal cords were removed; the upper,
Fig. 1. Kaplan-Meier survival analysis of G93A mice (n57) and G93A / intermediate and lower lumbar spinal cord were prepared,
P53-null (p532/2) mice were crossed to mice harbor- Lab) was available that was known to succumb to MND at ing the G93A gene. At the time this study was initiated, 6 to 8 months of age. In agreement with this report, our only one G93A mouse strain (stock[ 2300, The Jackson G93A mice (with normal p53 alleles, n57) died of
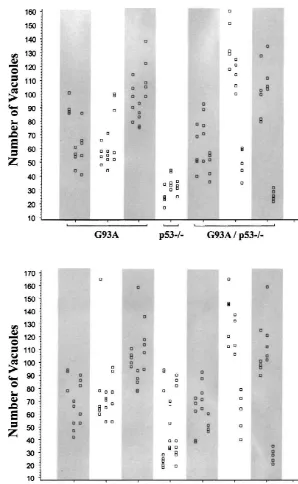
Fig. 2. Lumbar spinal cord of 140 day old pre-symptomatic transgenic mice. A–D: G93A mouse; E, F: G93A / p532/2mouse. (A) Low magnification of the ventral spinal cord (bar5100mm); square5location of picture B. (B) The vacuoles are more concentrated close to the limit between grey and white matter (arrow; bar5100mm). (C) Anterior horn with normal motor neuron and vacuoles located in the neutropil (bar510mm). (D) Small area of the anterior horn close to the border of white matter showing four motor neurons, two of them with signs of degeneration (‘dark cells’, see arrows) (bar52.5
paralysis at 212650 days (mean6S.D.), with a mean time and indeed, only four of the 22 double transgenics lived to of 20 days from the first signs of hindlimb weakness to experience MND. Their mean survival time was 160628 complete paralysis. This contrasted with the mean survival days (Fig. 1A), and there was no detectable difference in time of p53-deficient mice. These have been shown to die the interval between disease onset and death time when of tumors on average at 140 days of age [3,6]. Our compared with the G93A-only mice. These numbers (4 / 22 p532/2 mice (n511) died on average at 160649 days. animals) match the numbers that would be expected under With this discrepance in survival time between G93A and the described conditions of competing death risks if the p532/2mice, we expected that most of the G93A / p532/ absence of p53 fails to protect from MND (G93A / p532/
2 double transgenic animals would succumb to tumors, 2 vs. G93A-only: P50.84 (log rank test, competing risk
as regulated by the TrkA and p75 neurotrophin receptors, J. Cell analysis)). Thus, motor neurons damaged by G93A do not
Biol. 143 (1998) 1691–1703. die via p53-dependent apoptosis, unlike damaged resting
[2] C.N.G. Anderson, A.M. Tolkovsky, A role for MAPK / ERK in thymocytes and unlike, for instance, araC-damaged sympa- sympathetic neuron survival: Protection against a p53-dependent, thetic neurons that die in the presence and live in the JNK-independent induction of apoptosis by cytosine arabinoside, J. absence of p53 [2,7,16]. In accord with these results, Neurosci. 19 (1999) 664–673.
[3] L.D. Attardi, T. Jacks, The role of p53 in tumour suppression: pre-symptomatic 140 days old G93A mice displayed
lessons from mouse models, Cell. Mol. Life Sci. 55 (1999) 48–63. disease-associated vacuoles [9] within the dendrites of
[4] S. Bates, K.H. Vousden, Mechanisms of p53-mediated apoptosis, motor neurons regardless of p53 status (Fig. 2). There Cell. Mol. Life Sci. 55 (1999) 28–37.
were no significant differences between the numbers of [5] K. Blyth, A. Terry, M. O’Hara, E.W. Baxter, M. Campbell, M. vacuoles present in neurons from the lumbar spinal cord of Steward, L.A. Donehower, D.E. Onions, J.C. Neil, E.R. Cameron, Synergy between a human c-myc transgene and p53 null genotype G93A-only and G93A / p532/2 animals (Fig. 3).
in murine thymic lymphomas: contrasting effects of homozygous In the recent past, several studies have revealed that
and heterozygous p53 loss, Oncogene 10 (1995) 1717–1723. apoptosis effectors such as Bcl-2 or caspase-1 that act [6] J. Choi, L.A. Donehower, p53 in embryonic development: maintain-further downstream in the apoptosis cascades relative to ing a fine balance, Cell. Mol. Life Sci. 55 (1999) 38–47. p53, can indeed influence SOD1*-induced MND. For [7] A.R. Clarke, C.A. Purdie, D.J. Harrison, R.G. Morris, C.C. Bird,
M.L. Hooper, A.H. Wyllie, Thymocyte apoptosis induced by p53-instance, overexpression of the anti-apoptotic protein Bcl-2
dependent and independent pathways, Nature 362 (1993) 849–852. in the neurons of transgenic mice producing SOD1* delays
[8] D.W. Cleveland, From Charcot to SOD1: Mechanisms of selective the disease onset and increases the lifespan of the mice by motor neuron death in ALS, Neuron 24 (1999) 515–520. approximately 20% [14]. Furthermore, targeted disruption [9] M.C. Dal Canto, M.E. Gurney, Development of central nervous of caspase-1, one of the executing enzymatic mediators of system pathology in a murine transgenic mouse model of human amyotrophic lateral sclerosis, Am. J. Pathol. 145 (1994) 1271–1279. apoptosis, fails to affect disease onset or mortality but
[10] H.D. Durham, J. Roy, L. Dong, D.A. Figlewicz, Aggregation of slows down the progression of paralysis in transgenic mice
mutant Cu / Zn superoxide dismutase proteins in a culture model of harboring SOD1 mutant G93R [11]. Combined, these ALS, J. Neuropathol. Exp. Neurol. 56 (1997) 523–530.
observations suggest that apoptosis may play a role in [11] R.M. Friedlander, R.H. Brown, V. Gagliardini, J. Wang, J. Yuan, FALS but involve other elements of the cell death machin- Inhibition of ICE slows ALS in mice, Nature 388 (1997) 31.
[12] M.E. Gurney, H. Pu, A.Y. Chiu, M.C. Dal Canto, C.Y. Polchow, ery than p53. However, before a final conclusion on the
D.D. Alexander, J. Caliendo, A. Hentati, Y.W. Kwon, H.-X. Deng, W. role of p53 is reached, the results should be confirmed in a
Chen, P. Zhai, R.L. Sufit, T. Siddique, Motor neuron degeneration in larger study utilizing a SOD1 mutant mouse strain showing mice that express a human Cu,Zn superoxide dismutase mutation,
MND at an earlier age. Science 264 (1994) 1772–1775.
Crosses of p53 knockouts to other transgenic mice has [13] J. Jordan, M.F. Galindo, J.H.M. Prehn, R.R. Weichselbaum, M. Beckett, G.D. Ghadge, R.P. Roos, J.M. Leiden, R.J. Miller, p53 helped in the past to study whether the dysregulating or
expression induces apoptosis in hippocampal pyramidal neuron damaging overproduction of certain proteins can promote
cultures, J. Neurosci. 17 (1997) 1397–1405.
tumor development. It has been documented, for example, [14] V. Kostic, V. Jackson-Lewis, F. de Bilbao, M. Dubois-Dauphin, S. that overexpression of the c-Myc oncoprotein in a p53-null Przedborski, Bcl-2: Prolonging life in a transgenic mouse model of background can increase the incidence of T-cell lymphoma familial amyotrophic lateral sclerosis, Science 277 (1997) 559–562. [15] F.M. LaFeria, C. Kappel Hall, L. Ngo, G. Jay, Extracellular as compared to p53-null-only animals [5]. In this study, the
deposition of b-amyloid upon p53-dependent neuronal cell death in mean time to death of the G93A / p532/2 double
trans-transgenic mice, J. Clin. Invest. 98 (1996) 1626–1632.
genics that did not develop MND (n518) was 162650 [16] S.W. Lowe, E.M. Schmitt, S.W. Smith, B.A. Osborne, T. Jacks, p53 days, whereas the mean time to death of the p532/2only is required for radiation-induced apoptosis in mouse thymocytes, mice (n511) was 160649 days (Fig. 1B). This shows that Nature 362 (1993) 847–849.
[17] A. Migheli, C. Atzori, R. Piva, M. Tortarolo, M. Girelli, D. Schiffer, both groups share an equal risk of premature (non-MND)
C. Bendotti, Lack of apoptosis in mice with ALS, Nature Med. 5 death and suggests that the presence of the ubiquitously
(1999) 966–967.
expressed G93A protein does not increase the tumor [18] S. Rabizadeh, E.B. Gralla, D.R. Borchelt, R. Gwinn, J.S. Valentine, incidence in p53-null mice. S. Sisodia, P. Wong, M. Lee, H. Hahn, D.E. Bredesen, Mutations associated with amyotrophic lateral sclerosis convert superoxide dismutase from an antiapoptotic gene to a proapoptotic gene: Studies in yeast and neural cells, Proc. Natl. Acad. Sci. USA 92 Acknowledgements
(1995) 3024–3028.
[19] P.A. Trimmer, T.S. Smith, A.B. Jung, J.P. Bennett Jr, Dopamine This work was supported by the Deutsche Gesellschaft neurons from transgenic mice with a knockout of the p53 gene resist ¨
fur Muskelkranke e.V. and in part by grant SFB399(B5) MPTP neurotoxicity, Neurodegeneration 5 (1996) 233–239. [20] K.A. Wood, R.J. Youle, The role of free radicals and p53 in neuron from the German Research Foundation (DFG).
apoptosis in vivo, J. Neurosci. 15 (1995) 5851–5857.
References