KOMPLEMEN
6.1. PENDAHULUAN
Definisi: Komplemen, adalah senyawa yang mampu melisis sel yang diselimuti Ab, labil panas (rusak, jika dipanaskan pada suhu 56°C, selama 30 menit).
Protein Sistem Komplemen
Protein sistem komplemen terdiri dari lebih 25 protein yang berbeda (Tabel 1), diproduksi jaringan dan sel yang berbeda, termasuk hepatosit, makrofag dan sel epitel gut. Protein-protein ini diaktifkan oleh bermacam-macam agen dan aktivasi mereka terbentuk dalam kaskade yang menyebabkan lisis. Sebagai akibat, dengan tidak adanya salah satu dari kornponen dalam kaskade, dapat mengganggu kaskade dan hasil akhir reaksi tersebut.
Tabel 1 : Protein Sistem Komplemen
Kaskade klasik Kaskade lektin Kaskade alternatif Kaskade lisis Protein fungsional: Clqrs C2 C3 C4 Protein kontrol : Cl-INH C4-BP Mannan binding Protein (MBP) Mannan-associated Serine protease (MSAP, MASP2) C3, Faktor B & D*, Faktor Properdin
Fator I*, Faktor H, DAF, CR1 C5 C6 C7 C8 C9 Protein S BAB 6
Aktivasi Kaskade Komplemen
Aktivasi komplemeji dapat dibagi menjadi 4 kaskade : 1. Kaskade Klasik
2. Kaskade Lektin 3. Kaskade Alteraatif 4. Kaskade Lisis
Kaskade 1 dan 2, menyebabkan aktivasi C5 convertase, yang menghasilkan protein pecahan komplemen C5b, penting untuk aktivasi kaskade penyerang membran.
6.2. KASKADE KLASIK
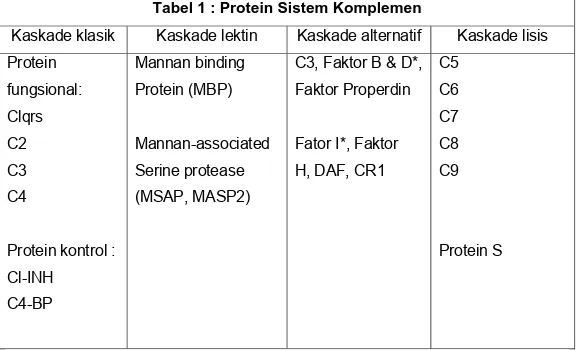
Kaskade klasik (Gambar 1), pada keadaan normal memerlukan kompleks Ag-Ab, protein komplemen Cl, C4, C2, C3 dan kation Ca+ and Mg++
Aktivasi Cl
Ikatan Clqrs (kompleks tergantung Ca), terdapat dalam serum normal, dengan kompleks Ag-Ab menghasilkan autokatalisis Clr. Clr memecah Cls dan pecahannya menjadi suatu enzim (C4-C2 convertase) yang mampu memecah C4 maupun C2.
Aktivasi C4 dan C2 (terbentuknya C3 convertase): Cls yang aktif memecah C4 menjadi C4a dan C4b.
C4b mengikat partikel antigen atau membran sel, sedangkan C4a tetap sebagai peptida aktif biologik pada sisi reaksi. C4b yang mengikat C2 yang mudah dipengaruhi Cls dan dipecah menjadi C2a dan C2b.
C2a tetap sebagai kompleks dengan C4b., sedangkan C2b dilepas ke dalam lingkungan mikro.
Kompleks C4b2a dikenal sebagai C3 convertase dan C2a merupakan bagian enzim.
Aktivasi C3 (terbentuknya C5 convertase): C3 convertase, dengan adanya
Mg++, memecah C3 menjadi C3a dan C3b.
C3b terikat pada membran, membentuk kompleks C4b2a3b, sedangkan C3a tetap berada dalam lingkungan mikro. Kompleks C4b2a3b, berfungsi sebagai C5 convertase yang memecah C5 menjadi C5a dan C5b.
Terbentuknya C5 convertase ini, menandai akhir dari kaskade klasik.
6.3. KASKADE LEKTIN
Adanya kaskade lektin, aktivasi C4 dapat dicapai tanpa partisipasi antibodi dan Cl (Gambar 2). Kaskade ini diinisiasi oleh 3 protein, yaitu : MBL (mannan-binding lectin), juga dikenal sebagai MBP(mannan-binding protein), yang berinteraksi dengan dua mannan-binding lectin-associated serine proteases (MASP dan MADSP2) yang analog dengan Clr dan Cls.
Interaksi ini menghasilkan kompleks yang analog dengan Clqrs dan menyebabkan aktivasi kaskade klasik yang tidak tergantung antibodi.
Clq juga mengikat beberapa agen, misalnya beberapa retrovirus, mikoplasma, asam poli-inosinat dan agregat IgG dan menginisiasi kaskade klasik.
Gambar 2 : Kaskade Lektin
6.4. KASKADE ALTERNATIF
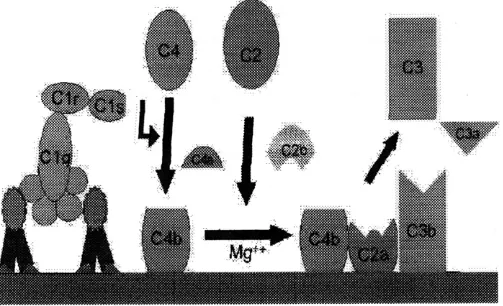
Kaskade alternatif dimulai dengan aktivasi C3 dan memerlukan faktor B dan Mg, yang kesemuanya terdapat dalam serum normal.
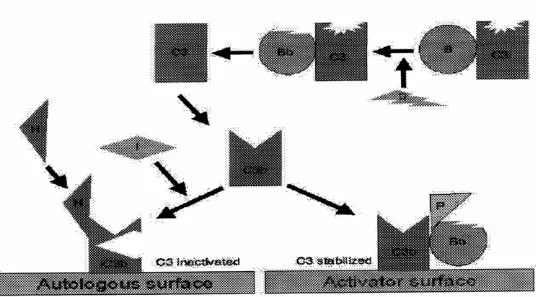
Aktivasi Spontan C3: suatu molekul metastabil seperti C3b (C3i) dihasilkan oleh adanya hidrolisis lambat dari C3 native. C3i mengikat faktor B yang dipecah oleh faktor D untuk menghasilkan C3iBb. Kompleks C3iBb memecah C3 menjadi C3a dan C3b (Gambar 3).
C3b mengikat factor B, yang juga dipecah lagi oleh faktor D untuk menghasilkan C3bBb (C3 convertase).
C3 convertase (juga dihasilkan dari kaskade klasik : C4b2a ) apabila tidak diaktifkan akan meneruskan aksinya pada C3 dan menyebabkan deplesi total C3.
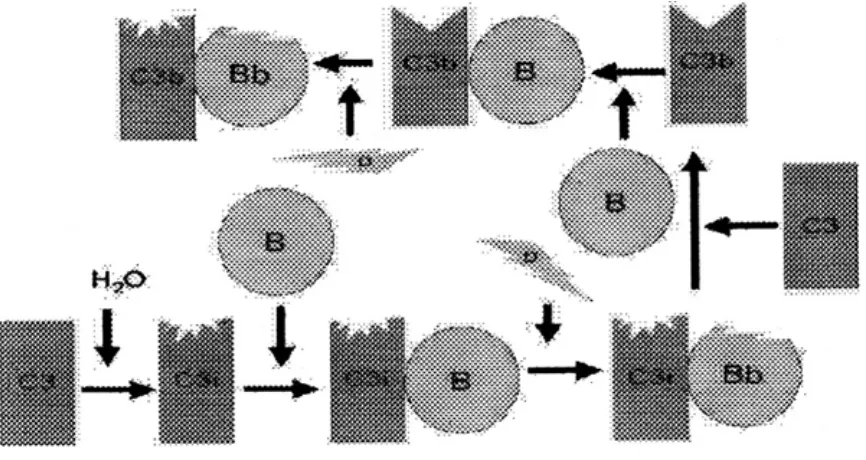
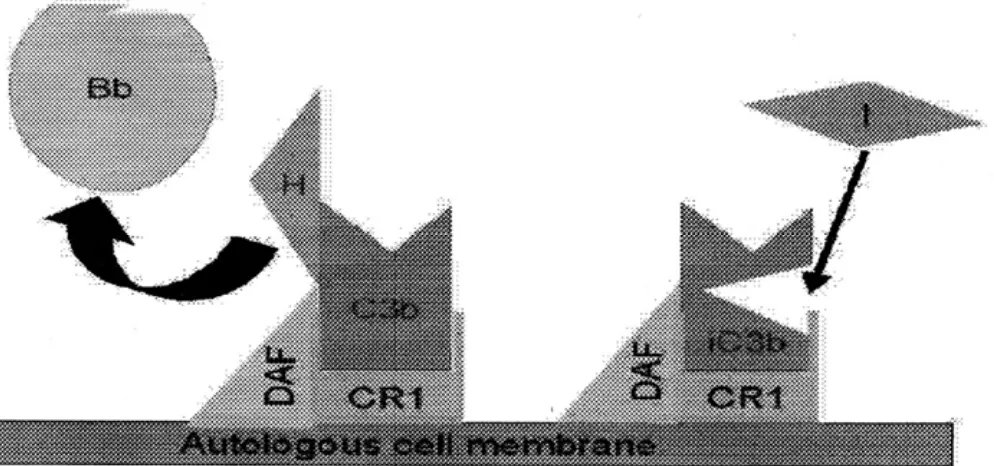
Pengaturan Normal C3 convertase: C3b, dalam fase cairan, mempunyai kehidupan yang pendek, kecuali menemukan membran atau sel sebagai penstabil yang cocok. (aktivator C3). Dengan tidak adanya patogen eksogenus, C3b akan mengikat dengan cepat sel darah merah autologus melalui reseptor C3b, CR1 pada sisi yang dekat dengan DAF ( mencegah adanya pengikatan faktor B). Ikatan dengan CR1 juga membuat C3b mudah dipengaruhi oleh faktor I (Gambar 4) yang memecahnya ke dalam beberapa fragmen (iC3b, C3d, C3e).
Gambar 4 : Pengaturan Aktivasi C3 oleh CR1
C4b, yang dihasilkan dari kaskade klasik, juga diatur oleh DAF, CR1 dan faktor I (Gambar 5).
Adanya kemsakan atau kekurangan DAF dapat menyebabkan Lisis sel dan anemia, yang akan menyebabkan tidak adanya aktivasi komplemen lebih lanjut dan menyebabkan kaskade penyerangan membrane dan lisi sel.
Protein serum yang lain, faktor H, dapat memindahkan faktor B dan mengikat C3b. Dcatan faktor H tersebut menyebabkan C3b mudah dipengaruhi faktor I (lihat Gambar 4).
C3 convertase yang dihasilkan kaskade klasik juga diatur oleh faktor DAF, CR1 dan factor I dengan cara yang mirip. Bedanya hanya bahwa protein yang mengikat C4b (C4b-BP, bukan faktor H) yang membuat mudah dipengaruhi faktor I.
Keadaan yang disebabkan oleh adanya defisiensi faktor I (atau faktor H) mengakibatkan aktivasi C3 tidak terkontrol dan menjadi penyebab utama defisiensi C3 yang dapat diturunkan (diwariskan).
Stabilisasi C3 convertase: Bakteri tertentu atau produknya (peptidoglikan, polisakarida dll) menyediakan aktivator permukaan untuk C3b. Jadi, C3b yang terikat pada permukaan seperti itu relatif resisten terhadap aksi dari faktor I (Gambar 6). Bahkan C3bBb yang terikat pada membran terdisosiasi dengan cepatnya. Akan tetapi, ikatan dengan protein yang lain, properdin, menstabilkan kompleks ini. Hal ini yang mendasari alasan, bahwa kaskade alternatif juga dinamakan kaskade properdin.
Terbentuknya C5 convertase : C3 convertase yang stabil memecah lebih banyak C3 dan menghasilkan kompleks C3bBb3b (analog dengan C4b2a3b dari kaskade klasik), dan C5 convertase yang dihasilkan, memecah C5 menjadi C5a dan C5b (Gambar 6).
C5b menginisiasi kaskade penyerangan membran yang dapat menyebabkan lisis sel. Sementara aktivasi kaskade C3 diinisiasi oleh mekanisme yang berbeda, analog satu sama lain dan keduanya dapat menyebabkan lisis membran. Kaskade alternatif menyediakan resistensi non spesifik terhadap infeksi tanpa adanya partisipasi antibodi dan juga menyediakan garis pertahanan pertama terhadap beberapa agen infeksi. Beberapa bakteri Gram - dan Gram +, virus tertentu, parasit, sel darah merah heterologus, agregat Ig (khususnya, IgA) dan beberapa protein yang lain dari kaskade alternatif (contohnya protease, produk kaskade kloting ). Protein, yaitu cobra venom factor (CVF) telah dipelajari secara ekstensif kemampuannya mengaktivasi kaskade ini.
6.5. KASKADE LITIK
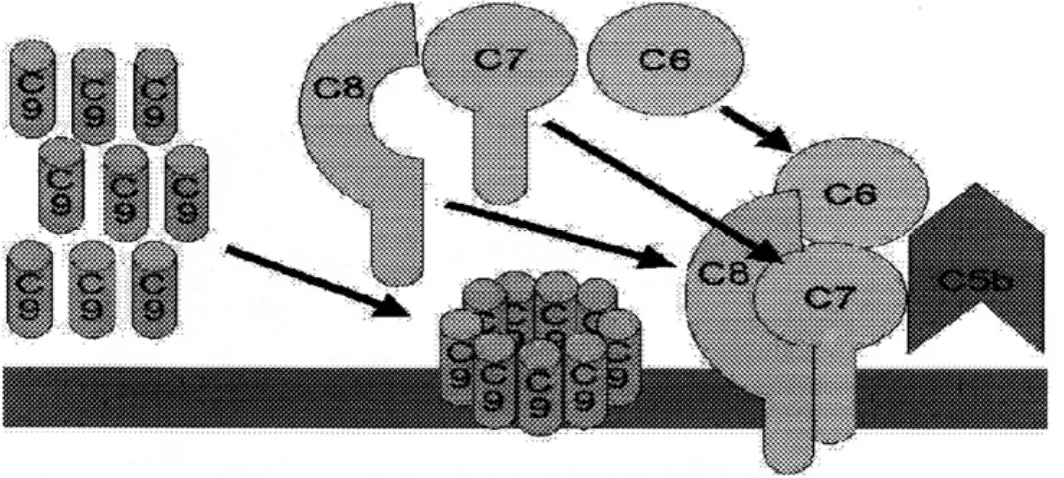
Kaskade litik (penyerang membran) melibatkan komponen C5, C6, C7, C8 dan C9.
C5 convertase yang diturunkan dari kaskade klasik maupun alternatif memecah C5 menjadi C5a dan C5b. C5b mengikat C6 dan selanjutnya C7 menghasilkan kompleks C5b67 yang hidrofobik, yang menyerang dengan cepat membrane plasma (Gambar 7)
Kemudian, C3 terikat pada kompleks ini dan menyebabkan masuknya beberapa molekul C9, terikat pada kompleks ini dan menyebabkan pembentukan pori-pori pada membran dan menghasilkan lisis sel. Lisis target sel oleh kompleks C5b6789 adalah non enzimatik dan dipercaya tergantung pada bentuk fisik membran plasma. C5b67 dapat terikat juga pada membran sel manapun yang mengakibatkan lisis sel. Tidak adanya diskriminasi seperti ini merusak sel yang dekat, dicegah oleh protein S (vitronektin) yang mengikat kompleks C5b67 dan membloknya untuk tidak terikat pada sel tanpa diskriminasi, selain target utama.
6.6. PRODUK AKTIF BIOLOGIK AKTIVASI KOMPLEMEN
Aktivasi komplemen menghasilkan produk yang mempunyai beberapa aktivitas biologik yang mendukung adanya resistensi anafilaksis dan inflamasi.
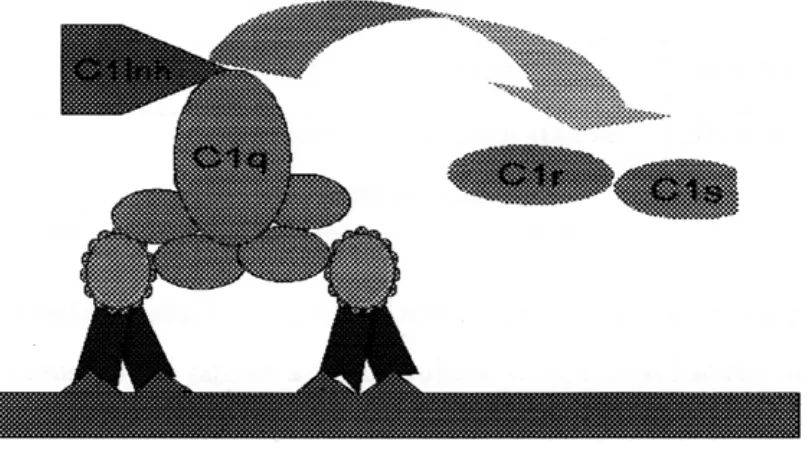
Produk kinin: C2b yang dihasilkan selama aktivasi komplemen kaskade klasik adalah prokinin yang menjadi aktif karena proses pengubahan enzimatik oleh plasmin. Produk C2b yang berlebihan dicegah oleh aktivasi C2 yang dipengaruhi oleh Cl inhibitor (Cl-INH) yang juga dikenal sebagai serpin, memindah Clqrs dari kompleks Clqrs (Gambar 8).
Defisiensi secara genetik Cl-INH menyebabkan adanya over produksi C2b dan mengakibatkan terjadinya edema hereditari angioneurotik.
Kondisi ini dapat diobati dengan Danasol yang meningkatkan produksi Cl-INH atau dengan asam ε-amino kaproat yang menurunkan aktivitas plasmin.
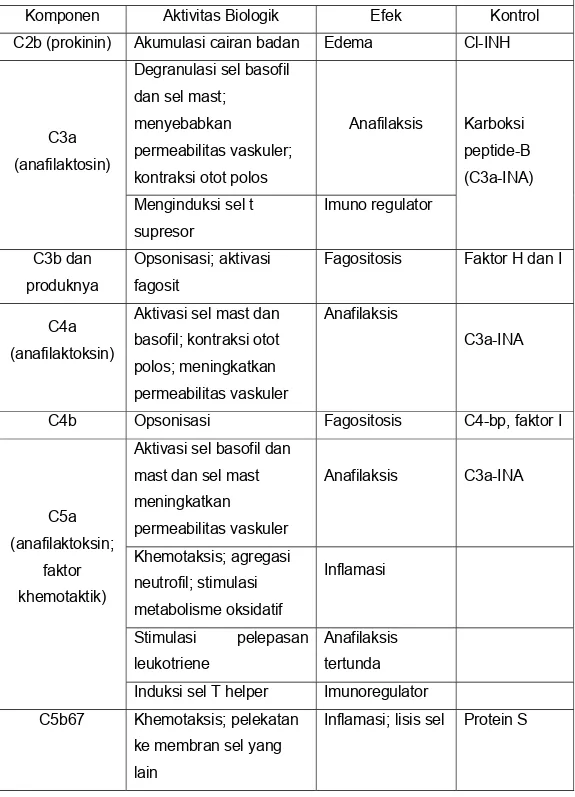
Tabel 2 : Sifat-sifat Biologik Produk Aktivasi Komplemen Komponen Aktivitas Biologik Efek Kontrol C2b (prokinin) Akumulasi cairan badan Edema Cl-INH
C3a (anafilaktosin)
Degranulasi sel basofil dan sel mast;
menyebabkan
permeabilitas vaskuler; kontraksi otot polos
Anafilaksis Karboksi peptide-B (C3a-INA) Menginduksi sel t supresor Imuno regulator C3b dan produknya Opsonisasi; aktivasi fagosit
Fagositosis Faktor H dan I
C4a (anafilaktoksin)
Aktivasi sel mast dan basofil; kontraksi otot polos; meningkatkan permeabilitas vaskuler
Anafilaksis
C3a-INA
C4b Opsonisasi Fagositosis C4-bp, faktor I
C5a (anafilaktoksin;
faktor khemotaktik)
Aktivasi sel basofil dan mast dan sel mast meningkatkan permeabilitas vaskuler Anafilaksis C3a-INA Khemotaksis; agregasi neutrofil; stimulasi metabolisme oksidatif Inflamasi Stimulasi pelepasan leukotriene Anafilaksis tertunda
Induksi sel T helper Imunoregulator C5b67 Khemotaksis; pelekatan
ke membran sel yang lain
Anafilaktoksin: C4a, C3a dan C5a (meningkat karena adanya aktivitas kaskade komplemen) adalah anafilaktoksin yang menyebabkan degranulasi sel basofil / sel mast dan kontraksi otot polos. Efek peptida ini dikontrol oleh karboksi peptidase B (C3a-INA)
Faktor khemotaktik: C5a dan MAC (C5b67), keduanya adalah faktor khemotatik. C5a juga merupakan activator yang poten untuk neutrofil, basofil dan makrofag dan menyebabkan induksi molekul adesi pada sel indotelial vaskuler.
Opsonin: C3b dan C4b pada permukaan organisme melekat pada reseptor-C (CR1) pada sel fagosit dan mengakibatkan fagositosis.
Produk biologik aktif yang lain: Produk degradasi C3 (iC3b, C3d dan C3e) juga terikat pada sel yang berbeda oleh reseptor yang berbeda dan memodulasi fangs! mereka.
6.7. KESIMPULAN
Sistem komplemen mengambil bagian dalam produk biologik spesifik dan non spesifik dan patofisiologik yang signifikan (Tabel 2). Ada beberapa defisiensi genetik komponen komplemen, C3, yang paling serius dan fatal. Defisiensi komplemen juga dijumpai dalam penyakit kompleks imun (misal SLE) dan bakterial akut dan kronik, infeksi viral dan parasit.
Anda telah mempelajari:
1. Protein sistem komplemen
2. Perbedaan dan persamaan aktivasi C3 pada kaskade yang berbeda 3. Perbedaan yang signifikan pada kaskade yang berbeda dalam imunitas
Tugas
a. Berikan jawaban dari pertanyaan-pertanyaan berikut
1. Komplemen diperlukan untuk apa saja dalam sistem imun?
2. Penderita SLE, menderita kekurangan protein komplemen apa saja?
3. Sebutkan fungsi biologik fragmen C5a
4. Dalam sistem yang mana DAF mengatur aktivasi komplemen? 5. Apa pengaruh kompleks imun Ag-Ab pada pasien dengan
defisiensi C3?
b. Carilah melalui internet, publikasi ilmiah dalam bahasa yang ada hubungannya dengan sistem komplemen, dan buatlah ringkasannya dalam bahasa Inggris