kromatografi kemudian diuji aktivitas inhibisinya dengan metode kolorimetri ATPase assay. Beberapa fraksi yang memiliki aktivitas inhibisi yang tinggi digunakan untuk tahapan selanjutnya (Lampiran 3).
Pengujian Inhibisi RNA Helikase Virus Hepatitis C (Utama et al. 2000)
Pengujian ini dapat mengukur jumlah fosfat yang dilepaskan dari hidrolisis senyawa ATP menjadi ADP. Metode ini diawali dengan pembuatan campuran utama yang terdiri dari 38.5 µL akuades steril, 5.0 µL 0.1 mM MOPS, 0.5 µL 0.1 M MgCl2, dan 1 µL 0.1 M ATP. Pengujian kolorimetri ATPase assay dibagi atas tiga bagian, yaitu a. campuran reaksi sebanyak 50 µL sebanyak 3 kali ulangan; b. campuran reaksi dan enzim sebanyak 50 µL dengan 3 kali ulangan; dan c. campuran reaksi, enzim, dan sampel sebanyak 50 µL dengan 3 kali ulangan. Kemudian proses inkubasi dilakukan selama 45 menit pada suhu ruang. Saat 15 menit sebelum waktu inkubasi habis, pewarna dibuat. Pewarna terdiri dari akuades, 0.081% malachite green, 2.3% polyvinil alkohol, dan 5.7% amonium molibdat dalam HCl 6 N dengan perbandingan 2:2:1:1. Sebanyak 100 µL pewarna dimasukkan ke dalam sumur (well). Larutan tersebut diinkubasi lagi selama 5 menit. Setelah waktu inkubasi habis, sebanyak 25 µL natrium sitrat ditambahkan ke dalam microtitter plate untuk menghentikan reaksi warna. Larutan tersebut diukur absorbansinya pada panjang gelombang 620 nm dan panjang gelombang 405 nm sebagai referensi. Satu unit aktivitas RNA helikase virus hepatitis C adalah jumlah mol fosfat bebas (Pi) yang terhidrolisis dari ATP per menit per mg protein.
Karakterisasi Bakteriosin Bakteri Asam Laktat S34
Tahapan karakterisasi Bakteriosin bakteri asam laktat S34 diawali dengan analisis bobot molekul bakteriosin menggunakan elektroforesis gel SDS poliakrilamid (SDS PAGE). Fraksi yang memiliki aktivitas inhibisi RNA helikase virus hepatitis C yang besar dianalisis bobot molekulnya menggunakan elektroforesis gel SDS poliakrilamid 20%. Gel poliakrilamid 20% dibuat dengan mencampurkan sukrosa, poliakrilamid, akuades, TEMED, dan amonium persulfat. Gel dibiarkan memadat. Sampel yang berupa fraksi hasil kromatografi
kolom dicampurkan dengan loading dye dan didenaturasi pada suhu 95ºC selama 10 menit. Perangkat gel elektroforesis dipasang dengan gel poliakrilamid yang telah memadat. Buffer running SDS PAGE dimasukkan kedalam perangkat SDS PAGE. Sampel dimasukkan sebanyak 20 µL ke dalam sumur agar elektroforesis dan marker sebagai pembanding. Setelah selesai elektroforesis SDS poliakrilamid, gel diwarnai dengan pewarnaan perak (silver stainning) hingga muncul pita-pita.
Setelah bakteriosin dianalisis dengan menggunakan SDS PAGE, Kadar protein diukur dengan BCA Protein Assay kit. Sebanyak 0.1 mL sampel ditambahkan 2 mL working reagent. Kemudian campuran tersebut diinkubasi pada suhu 37 ºC selama 30 menit. Absorbansi diukur pada panjang gelombang 562 nm.
HASIL DAN PEMBAHASAN
Hasil Ekspresi dan Purifikasi RNA Helikase Virus Hepatitis C.
Ekspresi RNA helikase virus hepatitis C dilakukan dengan bertujuan mendapatkan RNA helikase virus hepatitis C yang dihasilkan oleh bakteri Escherichia coli BL21(DE3)pLysS sebagai inang dari hasil kloning gen NS3 HCV (Utama 2000).
Penambahan
Isopropyl-β-D-thiogalactopyranoside (IPTG) bertujuan menginduksi ekspresi enzim pada fase logaritmik hingga fase stasioner ( Utama et al. 2000). Setelah penginduksian ekspresi enzim dilakukan pengumpulan pelet. Pengumpulan pelet tersebut berguna untuk mempertahankan stabilitas bakteri Escherichia coli yang telah diinduksi selama penyimpanan dan mengekspresikan RNA helikase untuk pengujian selanjutnya.
Pemurnian RNA helikase virus hepatitis C bertujuan mendapatkan RNA helikase virus hepatitis C yang murni dari hasil ekspresi gen NS3 helikase hepatitis C yang telah diklonkan dalam bakteri Escherichia coli BL21(DE3)pLysS sehingga dapat digunakan dalam penentuan aktivitas inhibisi dari bakteriosin. Proses purifikasi RNA helikase hepatitis C diawali dengan proses pemecahan sel yang terdiri atas dua metode yaitu pengering bekuan dan sonikasi. Proses pengeringbekuan dalam pemurnian RNA helikase hepatitis C dilakukan dengan bertujuan untuk memecahkan sel bakteri Escherichia coli BL21(DE3)pLysS karena
RNA helikase HCV bersifat intraseluler. Sonikasi merupakan tahapan selanjutnya dari proses pemecahan sel. Proses sentrifugasi yang dilakukan pada tahapan setelah pemecahan sel bertujuan memisahkan antara supernatan dan sel debris hasil pemecahan sel. Supernatan yang mengandung beberapa komponen intraseluler dikoleksi sebagian untuk proses identifikasi dengan menggunakan SDS PAGE sehingga dapat dilihat hasil proses pemurnian RNA helikase hepatitis C. Supernatan tersebut dimurnikan lebih lanjut dengan resin TALON metal affinity yang secara spesifik dapat menangkap enzim yang memiliki His-tag. RNA helikase yang telah terekspresi dalam bakteri Escherichia coli BL21(DE3)pLysS ini memiliki label 6 x His-tag yang sehingga dapat terikat secara spesifik oleh resin TALON metal affinity. Proses pengikatan resin terhadap RNA helikase hepatitis C menggunakan proses rotasi dan sentrifugasi sehingga pengikatan resin dengan enzim tersebut dapat diperoleh secara maksimal. Penambahan buffer B (10 mM Tris-HCl buffer (pH 8.5), 100 mM NaCl, 0.25% Tween 20) dilakukan untuk memisahkan enzim RNA helikase dengan komponen intraseluler lainnya. Sebagian kecil hasil pemisahan tersebut disimpan untuk identifikasi dengan SDS PAGE.
Pemurnian dengan TALON metal affinity dilakukan sebanyak 3 kali untuk mendapatkan RNA helikase hepatitis C dengan tingkat kemurnian yang tinggi. Penambahan buffer elusi (imidazola dalam buffer B) berfungsi sebagai eluen dalam proses elusi RNA helikase hepatitis C yang berikatan dengan resin. Imidazola yang merupakan komponen penyusun buffer tersebut dapat memutuskan ikatan antara RNA helikase hepatitis C dengan resin TALON metal affinity sehingga diperoleh enzim yang murni dapat diperoleh.
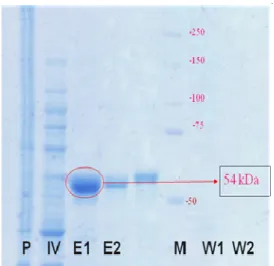
Hasil pemisahan antara debris sel dan komponen intraseluler (inner volume), hasil pemisahan antara elusi dan resin (washing 1 dan washing 2), dan RNA helikase hepatitis C hasil purifikasi (E1 dan E2) yang telah dikoleksi sebagian diidentifikasi dengan SDS PAGE. Hasil SDS PAGE menunjukkan pita tunggal pada elusi 1 dan elusi 2 yang membuktikan bahwa pemurnian RNA helikase virus hepatitis C dilakukan dengan baik. Bobot molekular dari RNA helikase virus hepatitis C adalah 54 kDa (Gambar 4). Hasil penelitian Utama et al. (2000) melaporkan bahwa bobot molekul RNA
helikase virus hepatitis C memiliki bobot molekul 54 kDa (Utama et al. 2000).
Gambar 4 Hasil SDS PAGE RNA helikase virus hepatitis C.
Hasil Identifikasi Bakteri Asam Laktat S34 Identifikasi bakteri asam laktat S34 dilakukan dengan metode identifikasi 16S rRNA. Metode identifikasi filogenetik 16S rRNA banyak digunakan untuk mengidentifikasi mikroorganisme yang berada di tanah, jalur pencernaan, dan sampel klinis (Klijn et al. 1991). Bakteri asam laktat merupakan mikroorganisme yang umumnya diisolasi dari olahan fermentasi (Gambar 5).
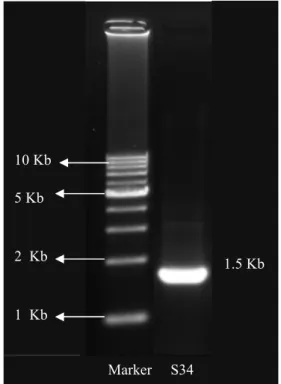
Identifikasi 16S rRNA terdiri atas isolasi DNA, amplifikasi DNA menggunakan teknik PCR, analisis bobot molekul, dan sekuening. Isolasi total genome dengan menggunakan metode Sambrook dan Russel (2001). Hasil isolasi DNA tersebut dianalisis bobot molekulnya dengan menggunakan metode elektroforesis agarosa. Modifikasi dilakukan dengan menggunakan lisozim yang berfungsi sebagai pendegradasi dinding sel bakteri Gram positif yang kaya akan polisakarida. Hasil analisis bobot molekul DNA bakteri hasil isolasi dan amplifikasi PCR menunjukkan bahwa DNA bakteri asam laktat S34 memiliki bobot molekul 1.5 kb (Gambar 6).
Hasil sekuening 16S rRNA bakteri asam laktat menghasilkan sekuen DNA bakteri asam laktat (Gambar 7). Hasil analisis sekuen DNA bakteri asam laktat S34 dengan menggunakan program BLAST menunjukkan bahwa bakteri asam laktat S34 memiliki kesamaan 99% dengan sekuen DNA bakteri Lactobacillus plantarum WCFS1 (Lampiran 4).
Gambar 5 Koloni bakteri asam laktat S34.
Gambar 6 Hasil elektroforesis DNA bakteri asam laktat S34.
Gambar 7 Hasil sekuening DNA bakteri asam laktat S34 dengan primer.
Kurva Pertumbuhan Isolat Bakteri Asam Laktat S34
Bakteriosin merupakan metabolit sekunder yang dihasilkan bakteri asam laktat pada masa pertumbuhan bakteri asam laktat. Peningkatan jumlah biomassa bakteri asam laktat menyebabkan peningkatan bakteriosin yang dihasilkan kemudian menurun pada fase stasioner (Usmiati dan Marwati 2007). Peningkatan jumlah bakteri asam laktat pada waktu tertentu menyebabkan perubahan jumlah nutrisi yang tersedia untuk metabolisme bakteri asam laktat sehingga metabolisme bakteriosin bakteri asam laktat terganggu.
Hasil perhitungan kurva OD bakteri asam laktat yang dilakukan selama 28 jam menunjukkan isolat bakteri asam laktat S34 fase logaritma terjadi setelah tujuh jam inkubasi (Gambar 8). Hasil penelitian Usmiati dan Marwati (2007) melaporkan bahwa fase logaritma bakteri asam laktat Lactobacillus sp terjadi pada jam ke 7. Fase stasioner tercapai pada jam ke-10 sampai jam ke-24 setelah inkubasi (Lampiran 5).
Gambar 8 Kurva pertumbuhan bakteri asam laktat S34.
Hasil Purifikasi dan Pengukuran Aktivitas Inhibisi Bakteriosin Bakteri Asam Laktat
S34 terhadap RNA Helikase Bakteriosin merupakan peptida intraseluler. Teknik sentrifugasi yang dilakukan pada tahapan awal purifikasi bakteriosin bertujuan memisahkan antara debris sel dan komponen intraseluler (supernatan). Supernatan yang dihasilkan masih mengandung banyak komponen intraseluler sehingga diperlukan purifikasi lanjutan.
Bakteriosin dipisahkan dari komponen intraseluler lainnya dengan menggunakan teknik presipitasi amonium sulfat. Teknik
10 Kb 5 Kb 2 Kb 1 Kb 1.5 Kb Marker S34
presipitasi amonium sulfat berdasarkan prinsip salting out. Penambahan amonium sulfat sebagai garam dalam tingkat kejenuhan tertentu dapat mengendapkan protein tertentu. Beberapa bakteriosin dapat diendapkan dengan amonium sulfat pada tingkat kejenuhan yang rendah (Sambrook et al. 1989). Namun ada pula bakteriosin yang dapat diendapkan dengan amonium sulfat pada tingkat kejenuhan yang tinggi. Sehingga diperlukan presipitasi amonium sulfat dengan beberapa rentang tingkat kejenuhan yaitu presipitasi amonium sulfat 60%, 70%, 80%, dan 90% (Lampiran 8). Peptida yang terendapkan melalui proses salting out tersebut dipresipitasi dengan menggunakan teknik sentrifugasi. Hasil presipitasi amonium sulfat dengan kejenuhan 80% diuji aktivitas inhibisinya menggunakan uji kolorimetri ATPase.
Kolorimetri ATPase assay adalah uji yang digunakan untuk beberapa enzim yang bergantung dengan keberadaan ATP/NTP sebagai donor energi untuk aktivitas enzim tersebut (Utama et al. 2000). Kolorimetri ATPase assay merupakan uji kualitatif dan kuantitatif.
Uji ini dapat dijadikan metode untuk mengukur inhibisi RNA helikase virus hepatitis C karena RNA helikase hepatitis C membutuhkan substrat berupa ATP (adenosine trifosfat) untuk membuka untaian ganda pada RNA yang merupakan material genetik dari virus hepatitis C. Larutan campuran utama berguna sebagai blanko dalam pengujian kolorimetri ATPase assay.
Keberadaan MOPS (Asam
4-morfolinopropanafosfat sulfonat) adalah sebagai buffer dalam campuran utama. Keberadaan Mg2+ dan Mn2+ diperlukan sebagai kofaktor RNA helikase sehingga MgCl2 berfungsi sebagai donor kofaktor dalam campuran utama (Utama et al. 2000).
Reaksi RNA helikase dengan ATP akan menghasilkan ADP (Adenosin difosfat) dan fosfat bebas. Fosfat bebas akan membentuk kompleks warna dengan pereaksi warna dari kolorimetri ATPase assay yang memiliki komposisi pewarna hijau malachite, polyvinil alkohol, dan amonium molibdat. Warna yang dihasilkan berkorelasi dengan jumlah fosfat bebas yang dihasilkan dari reaksi RNA helikase dengan ATP. Semakin hijau warna campuran reaksi maka semakin besar aktivitas RNA helikase bekerja (Gambar 9). Bakteriosin yang berfungsi sebagai inhibitor akan menghambat reaksi tersebut.
Kolorimetri ATPase assay juga dapat mengukur konsentrasi dengan menghitung serapan panjang gelombang dari campuran reaksi dengan RNA helikase dan bakteriosin. Panjang gelombang yang digunakan ada dua macam yaitu 620 nm dan 450 nm. Kedua panjang gelombang digunakan agar perhitungan reaksi RNA helikase dengan substratnya akurat. Konsentrasi ATP yang berubah karena adanya inhibitor dapat dihitung dengan membandingkan serapan panjang gelombang RNA Helikase.
Gambar 9 Reaksi pembentukan warna pada kolorimetri ATPase assay.
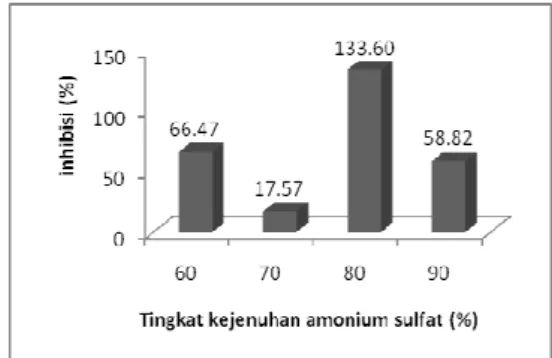
Berdasarkan hasil yang diperoleh melalui kolorimetri ATPase assay menunjukkan bahwa presipitasi amonium sulfat dengan kejenuhan 80% memiliki nilai inhibisi terhadap RNA helikase virus hepatitis C yang tertinggi yaitu 133.60% sedangkan presipitasi amonium sulfat dengan kejenuhan 60%, 70%, dan 90% memiliki nilai inhibisi 66.47 %, 17.57%, dan 58.82% terhadap RNA helikase virus hepatitis C (Gambar 10). Hal tersebut terjadi karena beberapa protein dapat diendapkan dengan konsentrasi tertentu sehingga banyaknya bakteriosin yang terendapkan tidak ekuivalen terhadap aktivitas inhibisi protein terhadap RNA helikase virus hepatitis C.
Gambar 10 Inhibisi fraksi amonium sulfat terhadap aktivitas ATPase RNA helikase virus hepatitis C.
Pemurnian bakteriosin selanjutnya yaitu dengan menggunakan kromatografi gel filtrasi yang menggunakan Sephadex G-50. Matriks Sephadex G-50 dapat memisahkan protein dari ukuran 1.5 kDa-30 kDa. Peptida yang berukuran kecil akan terjerap pada pori matriks Sephadex G-50 sedangkan protein yang berukuran besar akan terelusi terlebih dahulu. Eluen yang digunakan merupakan pelarut polar. Beberapa bakteriosin memiliki kemampuan untuk larut dalam beberapa pelarut seperti akuades, buffer fosfat, metanol dan Tris-HCl (Gautam dan Sharma 2009). Pelarut akuades tidak dapat melarutkan bakteriosin yang dihasilkan bakteri asam laktat S34. Pelarut natrium fosfat buffer tidak dapat digunakan karena mempengaruhi uji ATPase assay yang secara prinsip mengukur jumlah fosfat yang bebas sehingga penggunaan pelarut sodium fosfat buffer dapat menyebabkan kesalahan positif terhadap uji ATPase assay. Metanol dapat mengikat bakteriosin namun dapat pula menghilangkan aktivitasnya sehingga eluen yang digunakan adalah metanol 30%. Metanol dalam konsentrasi rendah tersebut dapat mengelusi bakteriosin serta aktivitas inhibisi bakteriosin terhadap RNA helikase virus hepatitis C tidak menurun.
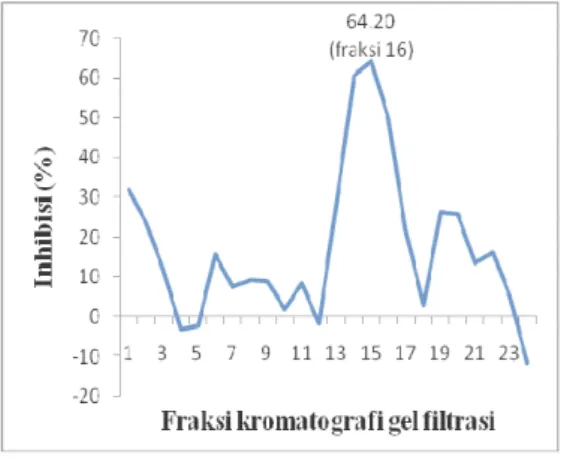
Sebanyak 25 fraksi didapatkan dari proses kromatografi gel filtrasi. Fraksi ke-16 memiliki persentase inhibisi yang lebih besar dari keseluruhan fraksi hasil gel filtrasi. Hasil uji kolorimetri ATPase menghasilkan fraksi ke-14 hingga fraksi ke-16 memiliki aktivitas inhibisi diatas 50% ketika diuji dengan menggunakan metode uji kolorimetri ATPase (Lampiran 9). Persentase inhibisi fraksi ke-14, fraksi ke-15, dan fraksi ke-16 adalah 58.89%, 60.61%, dan 64.20% (Gambar 11). Aktivitas inhibisi yang berada pada fraksi pertengahan mengimplikasikan bobot molekul protein yang memiliki aktivitas yang tinggi terhadap RNA helikase virus hepatitis C adalah protein yang berbobot molekul kecil sesuai dengan literatur yang menyebutkan bahwa semakin kecil molekul protein maka semakin lama terelusi dengan eluen (Scopes 1989).
Beberapa komposisi telah diujikan terhadap proses purifikasi bakteriosin dari bakteri asam laktat S34. Komposisi eluen yang dapat memisahkan bakteriosin bakteri asam laktat S34 yang dapat menginhibisi RNA helikase virus hepatitis C adalah komposisi eluen Metanol 30%. Konsentrasi metanol yang terlalu rendah menyebabkan banyak proein yang tidak terelusi sehingga masih berada pada matriks Sephadex G-50.
Konsentrasi metanol yang terlalu tinggi pada eluen dapat mengelusi protein dengan baik tetapi juga dapat menghilangkan aktivitas inhibisi bakteriosin secara cepat karena terjadi denaturasi (Gautam dan Sharma 2009)
Gambar 11 Inhibisi fraksi filtrasi gel Sephadex G-50 terhadap aktivitas ATPase RNA helikase virus hepatitis C.
Secara umum ringkasan purifikasi bakteriosin bakteri asam laktat S34 menunjukkan bakteriosin bakteri asam laktat S34 memiliki konsentrasi yang besar ekstrak kasar (Tabel 1). Aktivitas spesifik bakteriosin hasil presipitasi amonium sulfat dan kromatografi gel filtrasi sebesar
5.01 x 10
6 U/mg dan2.84 x 10
6 U/mg. Kemurnian bakteriosin yang didapat melalui tahapan presipitasi amonium sulfat 80% adalah 100% sedangkan kemurnian bakteriosin yang didapatkan dari hasil kromatografi gel filtrasi dengan eluen metanol 30% sebesar56.7%
(Tabel 1).Perhitungan konsentrasi protein menggunakan BCA kit assay. Metode ini mengkombinasikan reduksi Cu2+menjadi Cu+ oleh protein pada media alkalin (reaksi biuret). Kation cupro yang terbentuk akan bereaksi dengan asam bicinchoninat membentuk warna ungu yang dapat dideteksi pada panjang gelombang 562 nm.
Perhitungan konsentrasi protein menggunakan kurva standar Bovine Serum Albumin (BSA) yang telah diukur sebelum pengukuran konsentrasi protein (Lampiran 6). Ekstrak kasar yang diperoleh tidak dapat diukur aktivitasnya dan persentasi inhibisinya sebab media kultur bakteri memiliki fosfat bebas yang dapat menimbulkan kesalahan positif saat dilakukan pengujian inhibisi. Hal ini menyebabkan ekstrak kasar tidak dapat dihitung aktivitas spesifiknya.
Tabel 1 Ringkasan purifikasi bakteriosin Tahapan Volume (mL) Kadar Protein (mg/mL) Total protein (mg) Total Aktivitas (U) Aktivitas Spesifik (U/mg) Kemurnian (%) Ekstrak kasar 191 1.057 178.522 - - - Amonium Sulfat 80% 3 1.050 3.150 15.78 x 10 6 5.01 x 106 100 Sephadex G-50 1 0.884 0.884 2.51 x 10 6 2.84 x 106 56.7 Karakteristik Bakteriosin Bakteri Asam
Laktat S34
Bobot Molekul Bakteriosin Bakteri Asam Laktat S34
Analisis bobot molekul bakteriosin bakteri asam laktat S34 dilakukan dengan menggunakan metode elektroforesis gel poliakrilamida sodium dodesil sulfat (SDS PAGE). Teknik ini menganalisis bobot molekul bakteriosin lebih akurat daripada analisis bobot molekul menggunakan teknik kromatografi gel filtrasi sebab teknik ini menggunakan marker yang telah diketahui bobot molekulnya.
Denaturasi sampel diperlukan untuk SDS PAGE. Denaturasi sampel dengan suhu berfungsi untuk membuka struktur tersier dan kuartener dari peptida atau protein sampel. Peptida yang bersifat amfoter (memiliki dua muatan listrik) menjadi bermuatan negatif dengan adanya sodium dodesil sulfat sehingga saat elektroforesis bekerja peptida akan bergerak melalui arus dari muatan negatif (anoda) menuju muatan positif (katoda) berdasarkan dengan bobot molekul (Deyl 1983). Beberapa komponen penting selain SDS membantu proses pemisahan protein berdasarkan bobot molekulnya. Komponen tersebut adalah Tris berguna sebagai buffer dalam SDS PAGE, akrilamida sebagai komponen pembentuk gel, amonium persulfat
dan TEMED (N, N, N',
N'-tetrametiletilenediamin) yang berperan dalam proses polimerisasi akrilamida menjadi poliakrilamid dalam pembentukan gel.
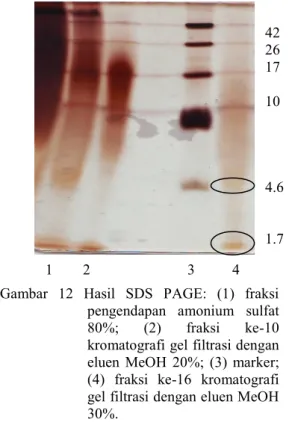
Hasil SDS PAGE dapat diberi warna dengan pewarna biru Coomasie atau pewarna perak. Pewarna perak lebih akurat menunjukkan pita-pita sampel karena memiliki sensitivitas yang lebih tinggi dibanding pewarna biru Coomasie. Hasil analisis bobot molekul bakteriosin bakteri asam laktat S34 menunjukkan pemisahan belum maksimal sebab masih banyak peptida yang muncul dengan berbagai ukuran.
Hasil elektroforesis gel sodium dodesil sulfat poliakrilamid fraksi ke-16 menunjukkan pita-pita yang sudah lebih sedikit dibandingkan dengan hasil elektroforesis gel sodium dodesil sulfat poliakrilamid fraksi amonium sulfat 80%. Terdapat 6 pita bakteriosin pada fraksi gel filtrasi ke-16 yang memiliki aktivitas yang tinggi dalam menginhibisi RNA helikase virus hepatitis C. Bakteriosin yang dapat menginhibisi virus hepatitis C berada pada kisaran 1.7-4.6 kDa (Gambar 12).
Bakteriosin tersebut termasuk kelas tipe 2. Ciri-ciri dari bakteriosin adalah berbobot molekul kecil dan stabil terhadap panas. Peptida yang stabil terhadap panas dapat dimanfaatkan pada industri makanan fermentasi karena peptida tersebut tidak akan rusak meski mengalami pemanasan sehingga makanan fermentasi olahan tidak perlu menggunakan pengawet karena memiliki kemampuan antibakteri dan antivirus.
s
Gambar 12 Hasil SDS PAGE: (1) fraksi pengendapan amonium sulfat 80%; (2) fraksi ke-10 kromatografi gel filtrasi dengan eluen MeOH 20%; (3) marker; (4) fraksi ke-16 kromatografi gel filtrasi dengan eluen MeOH 30%. 1 2 3 4 42 26 17 10 4.6 1.7