ASCENDING RETICULAR ACTIVATING SYSTEM
(ARAS)
Oleh:
dr. Ida Ayu Sri Wijayanti, M.Biomed, Sp.S
BAGIAN/SMF ILMU PENYAKIT SARAF
FK UNUD/RSUP SANGLAH DENPASAR
i
Om Swastyastu,
Puja dan puji syukur penulis panjatkan ke hadirat Tuhan Yang Maha Esa karena berkat rahmat-Nya tinjauan kepustakaan dengan judul “Ascending Reticular Activating System (ARAS)” ini dapat selesai pada waktunya.
Pada kesempatan ini penulis ingin mengucapkan terima kasih kepada berbagai pihak yang telah membantu penyelesaian tinjauan kepustakaan ini. Ucapan terima kasih penulis sampaikan kepada:
1. dr. AA Bagus Ngurah Nuartha, Sp.S(K) selaku Kepala Bagian/Kepala SMF di Bagian Ilmu Penyakit Saraf Fakultas Kedokteran Universitas Udayana/RSUP Sanglah Denpasar yang telah memberikan saya kesempatan untuk menyelesaikan karya tulis ini;
2. Dr.dr. Thomas Eko Purwata, Sp.S(K) selaku Kepala divisi Nyeri di Bagian Ilmu Penyakit Saraf Fakultas Kedokteran Universitas Udayana/RSUP Sanglah Denpasar yang telah memberikan pengarahan, kritik, dan saran dalam pembuatan tinjauan kepustakaan ini;
3. Seluruh pihak yang tidak dapat penulis sebutkan satu persatu yang telah membantu dalam penyusunan tinjauan kepustakaan ini.
Penulis menyadari laporan ini masih jauh dari kata sempurna sehingga saran dan kritik yang bersifat membangun sangat penulis harapkan untuk kesempurnaan tinjauan kepustakaan ini. Akhir kata, semoga tulisan ini dapat bermanfaat bagi pembaca.
Om Shanti, Shanti, Shanti Om.
Denpasar, Desember 2014
ii
DAFTAR ISI
KATA PENGANTAR……… i
DAFTAR ISI……….. ii
DAFTAR GAMBAR………. iii
DAFTAR TABEL………. iv
BAB 1 PENDAHULUAN………... 1
BAB 2 NEUROANATOMI ARAS……… 3
BAB 3 NEUROFISIOLOGI ARAS……….. 7
3.1. Neurotransmiter dalam Pengaktifan ARAS……… 9
3.2. ARAS sebagai Penggalak Kesadaran …………... 14
3.2.1 Patologi Gangguan Kesadaran………... 16
3.3 Peranan ARAS dalam Fisiologi Bangun Tidur………….……… 19
3.3.1 Patologi Tidur………. 25
BAB 4 RINGKASAN ……….. 29
iii
DAFTAR GAMBAR
Gambar 2.1 Permukaan Medial Otak...5
Gambar 2.2 Formasio Retikularis... 6
Gambar 3.1 Nukleus Raphae pada Batang Otak... 9
Gambar 3.2 Distribusi Norepinephrine pada Sistem Saraf Pusat... 12
Gambar 3.3 Distribusi Serotonin pada Sistem Saraf Pusat... 13
Gambar 3.4 Hubungan Timbal Balik Neurotransmiter Pengatur Kesadaran... 14
iv
DAFTAR TABEL
BAB I PENDAHULUAN
Susunan retikularis adalah susunan traktus-traktus yang bersifat nonspesifik, multineuron (polineuron), multisinaps (polisinaps), secara relatif difus dan mengintegrasi serta menghantarkan sejumlah impuls-impuls yang bersifat asenden. Susunan retikularis bersifat nonspesifik (secara primer tidak berhubungan dengan modalitas spesifik). Susunan ini terdapat di seluruh susunan saraf pusat, pada medulla spinalis, batang otak, serebellum, diensefalon dan hemisfer serebrum. Formasio retikularis merupakan substrat anatomi dan fungsional bagi banyak susunan retikuler (Noback, 1993).
Secara embriologi formasio retikularis merupakan salah satu unit fungsional tertua dari sistem saraf pusat, menempati bagian tengah dari batang otak, membentang ke arah rostral meliputi daerah midline, nukleus thalamikus retikularis dan intralaminaris, dan zona insersa dari subthalamus. Neuron retikularis menerima kolateral dari jalur asenden dan desenden, kecuali dari lemnikus medialis. Formasio retikularis pada bagian ventromedial dibatasi oleh traktus piramidalis dan lemnikus medialis, sedangkan bagian dorsolateral oleh jalur sensorik sekunder. Regulasi fungsi motorik saraf somatik dan saraf visceral (otonom) serta modulasi aktivitas elektrokortikal didukung oleh hubungan neuron retikularis dan pusat otonom pada otak dan medula spinalis. Fungsi tambahan dari formasio retikularis meliputi pengaturan ekspresi emosi, transmisi nyeri dan regulasi aktivitas reflek yang berhubungan dengan saraf kranialis (Arslan, 2001).
Susunan retikularis berhubungan dengan luas ekspresi perilaku kewaspadaan, perhatian hingga siklus tidur. Hilangnya pengaruh susunan retikuler asenden dapat menimbulkan seseorang tertidur. Pusat tidur diperkirakan terdapat di dalam formasio retikularis yaitu diantara pons bagian bawah dan bagian atas medula oblongata. Pada akhirnya, dipostulasikan bahwa tidur disebabkan secara aktif oleh aktivitas perangsangan terhadap pusat tidur di dalam batang otak (deaktivasi retikuler aktif) atau secara pasif oleh penekanan pengaruh susunan retikuler asenden (deaktivasi retikuler pasif). Suatu keadaan koma permanen, akibat adanya cedera otak, mungkin disebabkan oleh kerusakan
pada formasio retikularis (hilangnya pengaruh asenden) di dalam jalur retikuler asenden (Noback, 1993).
Kesadaran memperlihatkan dua segi yaitu derajat kesadaran dan kualitas kesadaran. Derajat kesadaran adalah kewaspadaan (alertness), tergantung dari jumlah impuls aferen yang sampai di korteks serebri. Sedangkan kualitas kesadaran seseorang tergantung dari cara pengelolaan impuls afferen oleh korteks serebri, yang akhirnya akan menghasilkan pola pemikiran yang mendasari kata-kata yang diucapkan atau hal yang dikerjakan seseorang. Kerusakan pada ARAS akan menimbulkan gangguan derajat kesadaran. Bila derajat kesadaran menurun, maka selalu akan tampak pula adanya gangguan dari kualitas kesadaran. Tapi sebaliknya, penderita dengan kualitas kesadaran menurun, tidak selalu memperlihatkan gangguan derajat kesadaran. Bila kita perhatikan fungsi dari Ascending Reticular Activating System (ARAS), maka susunan ini dapat kita namai Susunan penggalak kewaspadaan (Ngoerah, 1991).
BAB 2
NEUROANATOMI ARAS
Impuls-impuls aferen sampai di korteks serebri melalui dua lintasan, yaitu lintasan sensorik spesifik dan lintasan sensorik nonspesifik. Lintasan sensorik spesifik adalah lintasan-lintasan: traktus spinotalamikus, lemniskus medialis, lemniskus lateralis, radiasio optika dan lain-lain. Lintasan-lintasan ini mengantar impuls dari suatu titik alat reseptor ke suatu titik tertentu di korteks perseptif primer dari SSP. Sedangkan yang dimaksud dengan lintasan sensorik nonspesifik adalah serabut-serabut dalam formasio retikularis. Dalam perjalanannya, semua lintasan sensorik spesifik memiliki kolateral-kolateral ke formasio retikularis, dan serabut-serabut afferent dalam formasio retikularis inilah yang disebut dengan ARAS (Ngoerah, 1991).
ARAS merupakan suatu konsep fungsional yang terdiri dari formatio retikularis batang otak, subthalamus, hipothalamus dan thalamus medialis. Bagian otak ini diperlukan untuk mencetuskan dan mempertahankan keadaan sadar yang berwaspada. Serat retikular asenden ini sebagian besar berasal dari nukleus giganto cellularis, sebagian kecil dari nukleus reticularis ventralis dan lateralis. Serat ini menanjak di daerah traktus tegmentalis centralis, sebagian besar tidak menyilang garis median dan menuju ke bagian nukleus intralaminaris dan retikularis thalami. Sehingga impuls dapat disebarkan ke korteks serebri melalui traktus thalamokortikalis. Serabut dan lintasan dari nukleus retikularis thalamus ini dapat mengaktivasi korteks serebri tanpa bergantung kepada sistem sensorik spesifik atau sistem neural lainnya yang mengaktivasi korteks serebri (Chusid, 1990).
Secara embriologi formasio retikularis merupakan salah satu unit fungsional tertua dari sistem saraf pusat, menempati bagian tengah dari batang otak, membentang ke arah rostral meliputi daerah midline, nukleus thalamikus retikularis dan intralaminaris, dan zona insersa dari subthalamus. Neuron retikularis menerima kolateral dari jalur asenden dan desenden, kecuali lemnikus medialis. Formasio retikularis pada bagian ventromedial dibatasi oleh traktus piramidalis dan lemnikus medialis, dan bagian dorsolateral oleh jalur sensorik sekunder (Arslan, 2001).
Formasio retikularis adalah suatu struktur atau susunan neuron-neuron yang membentang secara berkesinambungan dengan variasi histologik minimum sepanjang medula spinalis, batang otak, daerah basal diensefalon dan telensefalon. Susunan retikular bersifat non-spesifik (secara primer tidak berhubungan dengan modalitas spesifik). Batas-batas formasio retikularis tidak dapat ditentukan secara tepat, karena serat-serat penyusunnya tidak dikelompokkan secara umum dalam traktus atau nukleus kompak. Substrat struktural formasio retikularis yang paling tetap adalah adanya neuron-neuron isodendrit. Namun, neuron-neuron ini juga ditemukan di daerah-daerah di luar formasio retikularis klasik. Akson-akson neuron retikuler bercabang menjadi cabang ascenden panjang dan cabang descenden panjang, cabang-cabang ini serta kolateralnya merupakan cabang akson transit. Cabang-cabang panjang membentuk susunan jalur yang menghantarkan output sel-sel ini ke rostral dan kaudal. Jalur-jalur ini adalah susunan-susunan difus yang diselingi oleh sejumlah besar hubungan sinaps dan diatur sebagai susunan neuron yang teratur. Formasio retikularis di dalam medula spinalis terletak pada zona intermedia (lamina VII) dengan perluasan-perluasan ke bagian anterior dan posterior. Jalur formasio retikularis medula spinalis adalah fasciculus proprius. Kemudian berlanjut ke arah rostral, menuju tegmentum batang otak. Sehingga formasio retikularis pada batang otak disebut reticular core, yang dibagi menjadi beberapa bagian, yaitu: nukleus-nukleus raphae dan kelompok nukleus retikuler paramedian yang terletak pada garis tengah batang otak. Kelompok nukleus retikuler sentral yang terletak pada 2/3 medial tegmentum. Kelompok nukleus retikularis lateral yang terletak pada 1/3 lateral tegmentum (Noback, 1993).
Ada dua sumber pengaruh pada sistem aktivasi retikuler asenden, yaitu: pengaruh impuls-impuls perifer yang mula-mula diantarkan melalui serat-serat sensorik spesifik; dan pengaruh yang datang dari korteks serebri melalui fibrae kortikoretikularis. Fibrae kortikoretikularis ini, yang berasal dari semua bagian korteks serebri, dapat memiliki pengaruh fasilitasi maupun inhibisi terhadap neuron-neuron formasio retikularis. Penelitian oleh Rossi dan Brodal (1956) menunjukkan bahwa sebagian besar fibrae kortikoretikularis ini berasal dari korteks motorik dan premotorik (Sukardi, 1985).
Jaras retikularis desenden (traktus retikulospinalis lateral dan ventral) berasal dalam nukleus yang mempunyai pengaruh aktivasi dan inhibisi pada neuron motorik spinalis. Nukleus-nukleus ini dipengaruhi oleh korteks serebral (korteks frontalis), serebelum dan ganglia basalis. Impuls aktivasi berasal dari bagian lateral formasio retikularis, terutama pada pons dan mesensefalon. Impuls inhibisi berasal dari nukleus retikularis bagian ventromedial dari medula oblongata (Duus,1996).
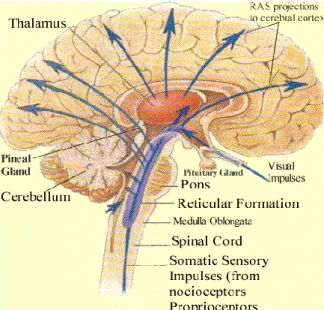
Gambar 2.1: Permukaan medial otak
Sumber:http://medicinembbs.blogspot.com 2009/10/reticular-activating-system.html
Berikut ini adalah inti-inti pada batang otak dan diencephalon yang merupakan komponen dari ARAS yang berperan dalam mekanisme kesadaran, yaitu:
1. Nukleus kawasan paramedian, medial dan lateral pons
2. Locus ceruleus (LC) atau blue spot atau nukleus pigmentosus pontis 3. Nukleus pedunkulopontin (PPN)
4. Nukleus retikularis mesensefalik
5. Hipotalamus posterior, dengan 4 nukleusnya yaitu:
- Nukleus Tubero-mammilary (TMN) yang bersifat histaminergik - Ventrolateral preoptik (VLPO) bersifat GABA-ergik
- Perifornical nuclei yang mensekresi orexin (hypocretin)
6. Sistem limbik yang terdiri dari amygdala, nukleus accumbens, nukleus basalis Meynert yang nukleinya bersifat glutamatergik yang diaktifkan oleh nuklei dari midpons.
7. Sistem Mesolimbik yang bersifat dopaminergik dan tampaknya lebih bertanggung jawab dalam hal respon kesadaran terhadap stimulus yang spesifik daripada mempertahankan keadaan bangun dari tidur.
8. Basal forebrain (substansia innominata) bersifat kolinergik, terutama di dalamnya adalah nukleus basalis, diagonal band of Broca, dan nukleus septal medial.
9. Thalamus, yaitu nukleus retikularis dan nukleus relay thalamic
Gambar 2.2: Formasio retikularis Sumber: Nieuwenhuys et al, 2007.
BAB 3
NEUROFISIOLOGI ARAS
Sistem retikuler (non spesifik) asenden adalah sistem aferen kompleks dengan hubungan luas dalam seluruh susunan saraf. Sistem ini menyediakan substrat struktural dan fungsional yang bekerja untuk mempengaruhi, memodulasi dan mengatur keadaan organisme dalam siklus tidur-bangun. Sistem ini menerima input dari bebagai sumber, diantaranya traktus spinoretikularis, jalur anterolateral-lemnikus spinalis, jalur trigeminus, formasio retikularis bagian lateral batang otak, jalur pendengaran, jalur optik dan jalur prenghidu. Berbagai input ini akhirnya diintegrasikan dalam ekspresi fungsional sistem retikuler, yang mungkin memperketat dan memperkecil efek rangsang. Neuron sistem retikuler dipengaruhi oleh rangsang dari banyak tempat di dalam tubuh (kulit, sendi, retina, organ spiral korti). Dalam hal ini, sistem retikuler terdiri dari satuan-satuan kompleks yang masing-masing berespon terhadap rangsangan dari banyak modalitas (Noback, 1993).
Impuls asenden aspesifik dari berbagai sumber dipancarkan secara difus ke korteks serebri. Komponen ascending reticular activating system (ARAS) berhubungan dengan perubahan derajat kesadaran. ARAS adalah anyaman polisinaptik tegmentum rumit, yang tidak hanya berhubungan dengan derajat kesadaran, tetapi juga berkaitan dengan memori, emosi dan motivasi. Susunan ini memiliki komponen ekstrinsik dan instrinsik. Komponen ekstrinsik terdiri dari neuron pada medula oblongata dan pons yang berespon terhadap stimulasi dari nervus kranialis dan nervus spinalis. Komponen instrinsik meliputi neuron pada mesencephalon yang menunjukkan aktivitas siklus (contoh: diurnal) yang berkaitan dengan proyeksi hipothalamus anterior (area suprachiasma) ke bagian otak tengah melalui medial forebrain bundle (Arslan, 2001).
Sistem retikuler di dalam batang otak dan hypothalamus sangat penting dalam aktivasi tonik korteks serebri. Hal ini secara obyektif terungkap dalam pemeliharaan siklus tidur-bangun dalam jangka waktu yang lama. Nukleus-nukleus retikuler thalamus sangat penting di dalam aktivasi fasik korteks serebri. Dorongan terakhir terungkap dalam respon terhadap perubahan-perubahan dalam intensitas masukan sensorik dengan
pergeseran segera terhadap perhatian. Jalur asenden utama pada sistem retikuler adalah traktus spinothalamikus tidak langsung (jalur paleospinothalamikus). Jalur ini menerima masukan tambahan di dalam formasio retikularis batang otak dari saraf otak dan cabang kolateral jalur asenden seperti traktus neospinthalamikus. Traktus tegmental sentral adalah jalur asenden utama dari formasio retikularis batang otak ke hypothalamus dan
thalamus. Sistem dengan susunan nonsomatotopis mengandung jalur nyeri
palaeospinothalamikus multineuron yang menghantarkan nyeri difus dengan lokalisasi tidak jelas. Rasa nyeri ini dirasakan setelah masa laten yang panjang setelah rangsang dan mungkin tetap bertahan untuk jangka waktu yang lebih panjang setelah rangsang dihentikan (Noback, 1993).
. Formasio retikularis terdiri dari sekelompok nukleus yang belum teridentifikasi keseluruhannya dan tersebar pada batang otak. Formasio retikularis berfungsi sebagai pusat pencetus aktivitas motorik (seperti berjalan dan berlari), mengatur regulasi konjugasi pergerakan bola mata, regulasi sistem respirasi dan sistem kardiovaskular. Neuron retikularis dikelompokkan menjadi raphae median, paramedian, nukleus medial dan lateral. Regulasi fungsi motorik saraf somatik dan saraf visceral (otonom) serta modulasi aktivitas elektrokortikal didukung oleh hubungan neuron retikularis dan pusat otonom pada otak dan medulla spinalis. Fungsi tambahan dari formasio retikularis meliputi pengaturan ekspresi emosi, transmisi nyeri dan regulasi aktivitas reflek yang berhubungan dengan saraf kranialis (Arslan, 2001).
Formasio retikularis adalah bangunan pengintegrasi yang paling bermakna, karena menjadi daerah dimana impuls-impuls dari modalitas sensorik maupun yang berasal dari sumber-sumber serebri dan serebellum bertemu dan berintegrasi. Daerah ini mampu memodifikasi aktivitas neuron-neuron dari input dan mampu menekan atau mendorong eksitabilitas banyak neuron, sehingga dapat menghambat, memfasilitasi atau memodifikasi transmisi informasi saraf bahkan melalui jalur-jalur spesifik. Perangsangan formasio retikularis mungkin meningkatkan sensibilitas nyeri. Susunan retikuler asenden dihubungkan dengan mekanisme integrasi saraf yang berhubungan dengan banyak segi aktivitas perilaku termasuk emosi, persepsi, motivasi, perangsangan, kesiagaan, tidur dan habituasi. Habituasi adalah suatu mekanisme saraf dimana suatu organisme menjadi tidak
memperhatikan rangsangan-rangsangan berulang secara monoton. Hal ini berhubungan dengan penurunan sensitivitas terhadap pola rangsang yang berulang. Sistem retikuler berfungsi memfasilitasi aktivitas sistem lemnikus. Sistem retikuler menyebabkan seseorang menjadi sadar terhadap rangsang yang diterima, bukan untuk menyampaikan dan melaporkan modalitas spesifik (Noback, 1993).
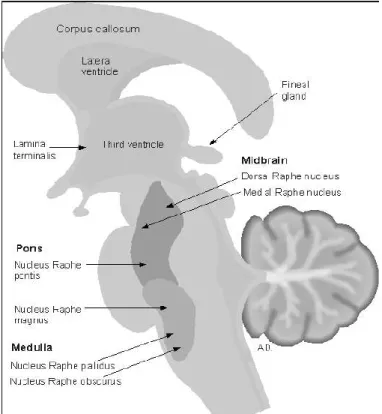
Gambar 3.1: Nukeus raphae pada batang otak Sumber: Arslan, 2001.
3.1 Neurotransmitter dalam pengakitfan ARAS
Mekanisme ARAS mengaktifkan sistem thalamokortikal disebabkan oleh adanya jalur glutamatergik dorsal dan jalur kolinergik ventral serta jalur lain yang berperan dalam pengaktifan ARAS, yaitu jalur dopaminergik, serotonergik, serta histaminergik (Guyton, 2007 ; Posner et al, 1980).
Glutamat adalah salah satu neurotransmitter asam amino yang disekresi oleh terminal presinaps pada jalur-jalur sensoris yang memasuki sistem saraf pusat. Jalur ini bersifat mengaktifkan reseptor eksitasi. Glutamat merupakan neurotransmitter utama
dalam proyeksi talamokortikal dan proyeksi kortikostriatal. Nukleus-nukleus pada hipokampus terutama menghasilkan neurotransmiter ini.
Banyak neuron tambahan yang berproyeksi ke nuklei relay, garis tengah dan intralaminar thalamus. Kebanyakan dari neuron ini adalah neuron glutamatergik yang memperkuat sinyal kewaspadaan yang datang dari tegmentum mesopontin. Di lain pihak tampaknya neuron-neuron tersebut tidak mampu mempertahankan status kesadaran pada kasus-kasus kehilangan pengaruh dari neuron mesopontin secara akut.
Pada level mesopontin batang otak terdiri dari setidaknya tiga kelompok monoamin (norepinefrin, serotonin, dan asetilkolin) yang berbeda yang akson-aksonnya berproyeksi menuju thalamus kemudian ke korteks serebri secara difus.
Sumber utama input aferen mesopontin yang terentang ke talamus adalah kumpulan dari neuron-neuron kolinergik yang membentuk dua kelompok besar yaitu nukleus pedunkulopontin (PPN) dan nukleus tegmental laterodorsal (LDT). Neuron-neuron dari kedua nukleus ini berproyeksi menuju formatio retikularis paramedian mesensefalon kemudian ke pusat relai di thalamus yaitu nukleus garis tengah dan intralaminar yang menginervasi korteks serebri secara difus dan juga nukleus retikularis thalamus yang memainkan peranan penting dalam transmisi thalamokortikal dengan menghiperpolarisasi neuron-neuron relai thalamik melalui reseptor GABAB. Neuron-neuron lain pada PPN dan LDT mengirim akson-akson ke lateral hipotalamus, dimana mereka akan berhubungan dengan kumpulan neuron yang berproyeksi ke kortikal secara difus. Neuron-neuron pada PPN dan nukleus LDT paling aktif saat terjaga dan tidur REM, dua kondisi yang menghasilkan gambaran EEG voltage rendah, cepat (desinchronized), dan melambat selama fase tidur NREM saat EEG didominasi oleh gelombang lambat voltage tinggi.
Sekelompok neuron kolinergik di nukleus basalis Meynert berjalan ke sistem limbik dan korteks serebri. Sedangkan neuron kolinergik tambahan pada formatio retikularis yaitu pada sel-sel kolinergik tegmentum terutama pada nukleus kuneiformyang kemudian berjalan ke hipotalamus, talamus, sistem limbik dan korteks serebri.
Dalam perjalanan proyeksi neuron monoaminergik komponen ARAS melalui basal forebrain dan melalui jalurnya ke korteks serebri, mereka bergabung dan
memperbanyak diri dengan adanya populasi neuron-neuron kolinergik nukleus magnoseluler pada basal forebrain. Neuron kolinergik ukuran besar ini menerima aferen dari seluruh jalur neuron komponen ARAS pada hipotalamus dan neuron monoaminergik yang merupakan komponen ARAS pada batang otak dan menyertai perjalanannya dalam menginervasi korteks serebri secara difus. Namun pola terminasi neuron kolinergik pada korteks serebri lebih spesifik jika dibandingkan dengan input neuron monoaminergik. Akson-akson neuron monoaminergik individual secara tipikal menyebar secara luas pada korteks serebri. Sementara akson-akson dari neuron kolinergik basal forebrain masing-masing menginervasi korteks serebri dengan diameter yang hanya beberapa milimeter. Berdasarkan rekaman basal forebrain binatang coba Rodentia, pada percobaan siklus bangun tidur mengindikasikan bahwa neuron kolinergik ini memiliki pola aktivitas yang luas. Aktivitas neuron-neuron basal forebrain berkorelasi dengan status behavioral yang kompleks sehingga diduga neuron-neuron ini amat berpengaruh pada aspek fungsi luhur yaitu kesadaran atau kewaspadaan seperti memfokuskan perhatian (Posner et al, 1980).
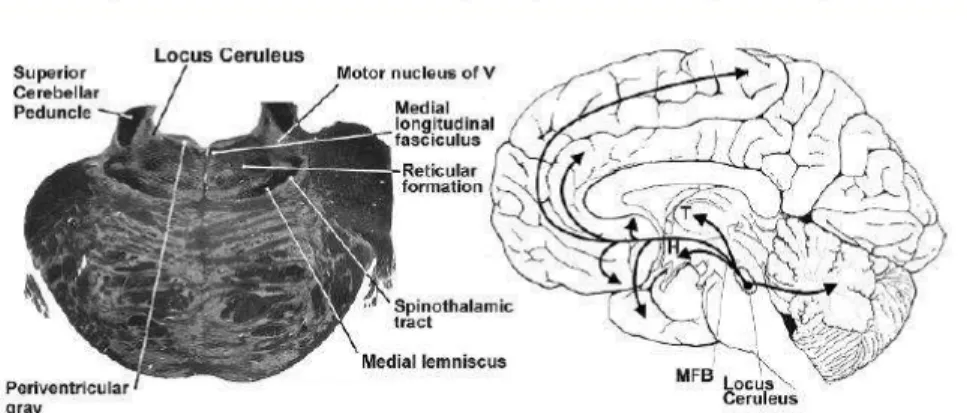
Pusat utama untuk neuron-neuron yang menghasilkan norepinefrin adalah nuklei pontin khususnya LC dan nukleus raphae bagian kaudal. Norepinefrin disekresi pada bagian terminal neuron-neuron yang berlokasi pada LC. Serat-serat saraf asenden dari LC kemudian berproyeksi ke talamus, hipotalamus, sistem limbik dan korteks frontal. Sedangkan proyeksi serat saraf dari raphe nuklei bagian kaudal adalah ke daerah amygdala. Adanya pengaktifan jalur neurokimia norepinefrin ini membantu mengontrol seluruh aktifitas dan mood, dengan mengaktivasi reseptor eksitasi seperti misalnya meningkatkan derajat kewaspadaan walaupun beberapa area dapat bersifat inhibisi (Ganong, 2002 ; Guyton, 2007).
Gambar 3.2: Distribusi norepinefrine pada sistem saraf pusat Sumber: Mendoza, 2008
Tempat utama untuk badan sel serotonergik adalah pons bagian atas dan mesensefalon. Secara spesifik yaitu nuklei raphe dorsalis, locus ceruleus caudal, area postrema dan area interpedunkular. Neuron tersebut berhubungan ke ganglia basalis, sistem limbik, lalu korteks serebri. Berhubungan dengan aspek prilaku dan siklus bangun tidur. Dapat terjadi insomnia bila terjadi kerusakan di sel-sel nuklei raphe atau bila cadangan serotonin berkurang oleh karena obat. Sebaliknya peningkatan level serotonin menurunkan sensitivitas terhadap nyeri. Sedangkan neuron-neuron serotonergik yang asalnya dari raphe nuklei median berproyeksi melalui jalur yang sama.
Sejumlah kecil neuron-neuron yang bersifat dopaminergik yang berasal dari ventral tegmental sepanjang midline dari mesensefalon bergabung dengan neuron serotoninergik menuju area di bawah akuaduktus serebri. Neuron dopamin ini juga berproyeksi melalui formatio retikularis paramedian mesensefalon yang kemudian menuju ke nuklei garis tengah dan intralaminar thalamus, sementara sebagian lagi melalui hipotalamus lateral menuju ke basal forebrain dan korteks prefrontal. Penelitian mutakhir membuktikan neuron-neuron monoaminergik ini aktif saat kondisi bangun, kurang aktif pada tidur gelombang lambat dan inaktif saat tidur REM.
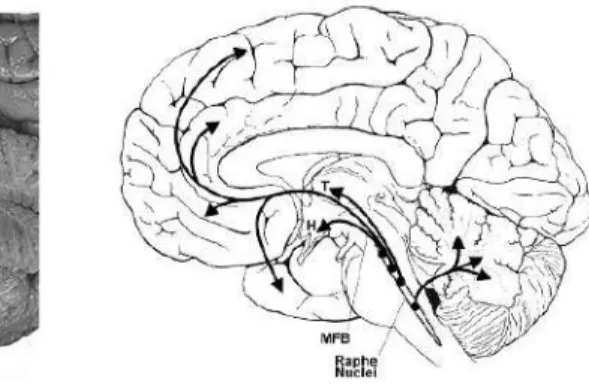
Gambar 3.3: Distribusi serotonin pada sistem saraf pusat Sumber: Mendoza, 2008
Neuron yang melepaskan histamin sebagai neurotransmitternya berlokasi di hipotalamus dan kemudian berjalan ke korteks serebri, sistem limbik dan talamus. Sebagaimana halnya salah satu jalur proyeksi ARAS yang menuju daerah hipotalamus, terdapat sejumlah neuron pada hipotalamus yang memproyeksikan impuls kesadaran ke daerah basal forebrain dan korteks serebri. Neuron-neuron ini termasuk neuron histaminergik pada nukleus TMN demikian juga dengan neuron dari daerah hipotalamus lateral yang menginervasi nukleus intralaminar dan nukleus garis tengah lalu memproyeksikan impuls secara difus ke korteks serebri. Terdapat banyak bukti bahwa input histaminergik sangat penting dalam hal memelihara status terjaga (wakefulness). Histamin H1 dapat memblokade gangguan status terjaga percobaan dengan binatang ataupun manusia. Pada tikus transgenik yang kekurangan histidin decarboxilase menunjukkan defisit pada status keterjagaan dan bila tikus ini diinjeksi dengan inhibitor histamin maka menunjukkan adanya penurunan kesadaran (Zeman, 2001 ; Posner et al, 1980).
Beberapa neuron hipotalamus lateral mengandung orexin/hipocretin, yaitu suatu peptida yang diasosiasikan dengan keadaan waspada dan terjaga sedangkan neuron lainnya melanin-concentrating hormone ataupun GABA. Neuron yang menghasilkan orexin/hipocretin berproduksi secara aktif selama status terjaga dan menurun selama periode tidur gelombang lambat dan tidur REM.
Sehingga dapat dinyatakan bahwa ARAS terdiri dari jalur-jalur ascendens yang multipel, yang berasal dari tegmentum mesopontin, tapi diperbanyak oleh input-input
tambahan pada setiap level, dimana jalur ascenden tersebut akan melalui basal forebrain, thalamus dan korteks serebri. Jalur-jalur yang berbeda ini mungkin aktif secara independen dibawah kondisi yang bervariasi, yang memodulasi kapasitas fungsional dari neuron-neuron kortikal.
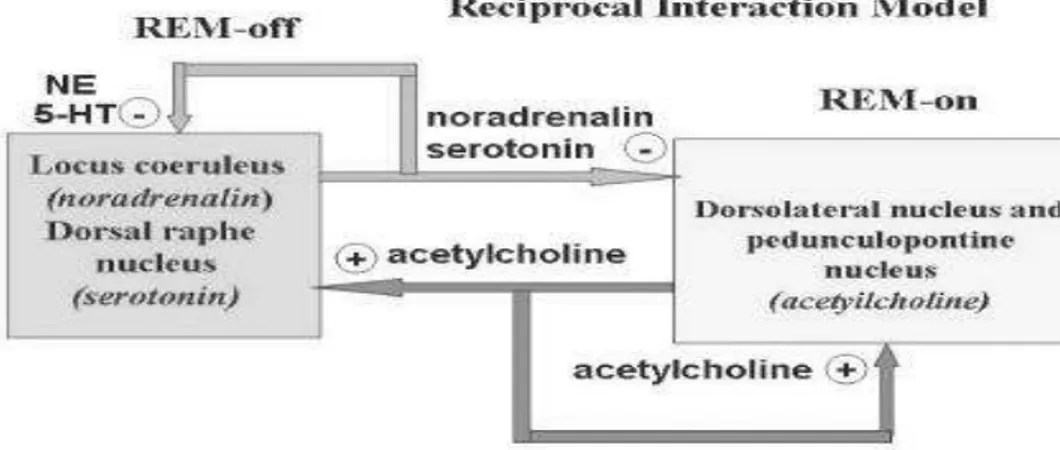
Gambar 3.4: Hubungan timbal balik neurotransmitter pengatur kesadaran Sumber: Parkers, 1996
3.2 ARAS sebagai Penggalak Kesadaran
ARAS memiliki komponen anatomi dan neurokimia yang multipel pada rostral tegmentum batang otak dan diensefalon yang bertanggung jawab terhadap kesiapsiagaan yang merupakan prasyarat untuk suatu kesadaran yang maksimal (Youbg and Pigott, 1999).
Konsep kesadaran terbagi menjadi dua yaitu kesadaran sebagai suatu pengalaman dan kesadaran sebagai status terjaga (waking state) yang terdiri dari keadaan terjaga, mengantuk, tidur, tidur REM, perubahan derajat kesadaran patologis seperti koma, halusinasi, dan meregulasi korelasi elektrik neuron-neuron yang menyusunnya (Zeman, 2001 ; Guyton, 2007).
Kesadaran yang adekuat disebut compos mentis, yaitu pada mana aksi dan reaksi terhadap apa yang dilihat, didengar, dihidu, dikecap, dialami, dan perasaan
keseimbangan, nyeri, suhu, raba, gerak, getar, tekan, dan sikap, bersifat adekuat yaitu tepat dan sesuai (Mardjono dan Sidharta, 2008).
Seseorang dikatakan compos mentis bila derajat dan kualitas kesadaran penderita adalah baik. Derajat kesadaran adalah kewaspadaan, tergantung dari jumlah impuls eferen yang sampai di korteks serebri yang akhirnya akan menghasilkan pola-pola pemikiran yang mendasari kata yang diucapkan dan hal-hal yang dikerjakan seseorang menentukan kualitas kesadaran seseorang.
Korteks serebri tidak hanya distimulasi secara sepihak oleh ARAS tapi juga secara resiprokal menstimulasi dan memodulasi aktivitas ARAS, menghasilkan suatu mekanisme umpan balik. Namun mekanisme fisiologis secara detail proyeksi nukleus-nukleus yang menyusun ARAS pada batang otak ke arah rostral menuju korteks serebri belu sepenuhnya dipahami. Setidaknya dapat diidentifikasi tiga jalur proyeksi ARAS ke korteks serebri, yaitu:
1. Proyeksi ARAS melalui nukleus spesifik retikularis thalami ke korteks serebri 2. Proyeksi ARAS melalui hipothalamus untuk memberi pengaruh pada
struktur-struktur basal forebrain termasuk sistem limbik
3. Proyeksi aksonal dari neuron-neuron serotonin yang menyusun ARAS yang terdapat pada nukleus raphae (direct afferent system)
Studi anatomik yang dilakukan oleh Scheibels menunjukkan bahwa nukleus retikular thalami berproyeksi hanya pada thalamus dan efek fisiologis difus pada korteks dimediasi oleh nukleus thalamik yang spesifik. Stimulasi pada area retikularis mesensefalon menurunkan atau bahkan menghilangkan pengaruh inhibisi tonik dari nukleus retikularis thalami seperti halnya yang terjadi dengan stimulasi jalur frontal-thalamik yang membawa impuls kembali ke thalamus melalui sistem thalamofrontal kortikal. Melalui peningkatan atau penurunan mekanisme inhibisi thalamik pada korteks serebri, ARAS akan menghasilkan suatu mekanisme “gate” yang membatasi atau meningkatkan pengaruh pada korteks serebri (Guyton, 2007 ; Posner et al, 1980).
Mekanisme relay sistem limbik oleh ARAS mempengaruhi korteks serebri merupakan hal yang penting dalam fungsi kesadaran. Hipotalamus memiliki banyak serat
saraf yang berproyeksi mencapai amygdala dan area septal. Proyeksi tersebut pun mencapai komponen lain dari sistem limbik yakni hipokampus serta dorsal nukleus thalamus. Khususnya yang terpenting adalah proyeksi mayor ke area neokorteks lobus prefrontal melalui medio dorsal talamus. Nauta berspekulasi bahwa hubungan resiprokal retikular-hipotalamik-neokortikal memodulasi tidak hanya pada level kesadaran organisme, tapi juga respon-respon eksternal terhadap vegetatif instingtual dan pengaturan emosional. Kenyataannya bahwa setiap area korteks sensoris berproyeksi balik melalui mekanisme relay yang multipel ke arah sistem limbik (Posner et al, 1980). 3.2.1 Patologi Gangguan Kesadaran
Gangguan derajat kesadaran dapat dianggap sebagai gangguan fungsi neuron-neuron kortikal sebagai pengemban kesadaran. Menurunnya kewaspadaan dapat pula berarti menurunnya kesadaran. Bila fungsi korteks serebri secara bilateral dan menyeluruh terganggu maka kewaspadaan penderita akan terganggu. Bila ARAS terganggu fungsinya maka kewaspadaan penderita akan terganggu pula. Bila pengelolaan impuls-impuls yang sampai pada korteks serebri terganggu, sedangkan fungsi dari ARAS baik, maka kualitas kesadaran penderitalah yang terganggu.
Ganguan kesadaran yang berat oleh karena gangguan neuron-neuron pengemban kesadaran disebut dengan koma kortikal bihemisferik. Sedangkan gangguan kesadaran yang disebabkan oleh disfungsi ARAS disebut dengan koma diensefalik yang dapat ditimbulkan oleh adanya proses patologis supratentorial dan infratentorial.
Koma Kortikal Bihemisfer
Gangguan kehidupan neuron-neuron sehingga tidak berfungsi kembali, dapat ditinjau secara menyeluruh bilamana struktur dan metabolismenya dipahami. Neuron merupakan satuan fungsional susunan saraf. Neuron berbeda dalam struktur, metabolisme, dan fungsinya dengan sel-sel tubuh yang lain oleh karena neuron tidak bermitosis. Neuron hanya menggunakan oksigen dan glukosa saja untuk metabolismenya. Beberapa kondisi yang menyebabkan terganggunya kondisi optimal neuron-neuron hemisfer serebri untuk melaksanakan fungsinya, misalnya:
- Gangguan oksigenasi atau respirasi dapat menimbulkan anoksemia - Gangguan keseimbangan elektrolit misalnya pada diare
- Toksemia atau intoksikasi dapat menyebabkan penurunan konsentrasi enzim dan substrat lain
Koma yang timbul oleh karena adanya gangguan metabolisme sel-sel neuron di korteks serebri pada kedua hemisfer dinamakan koma kortikal bihemisfer. Pada suatu permulaan koma ini akan tampak suatu permulaan sindroma otak organik yang memberi gambaran psikiatrik misalnya delirium, dan gangguan fungsi luhur seperti gangguan memori, gangguan orientasi, atau pengertian. Disamping itu tampak gejala-gejala gangguan saraf misalnya gangguan ketangkasan gerakan, gerakan involunter, ataupun bangkitan epilepsi (Ngoerah, 1991).
Destruksi bilateral korteks serebri yang bersifat difus atau substansia alba yang barada dibawahnya menyebabkan hilangnya substrat metabolik seperti oksigen, glukosa, ataupun gangguan pada aliran darah yang membawanya. Kondisi ini sering terjadi pada pasien yang mengalami hipoksia oleh karena kegagalan pulmonal atau pada pasien dengan hipoglikemia yang lama. Kekurangan substrat metabolik tersebut menyebabkan neuron-neuron pada lapisan korteks III dan V dan pada CA1 dan CA3 formasi hipocampal mengalami kerusakan yang diperkirakan menyebabkan toksisitas asam amino yang bersifat eksitasi. Pengeluaran neurotransmitter yang bersifat eksitasi yang berlebihan pada reseptor N-metyl-D-aspartat (NMDA) menyebabkan perpindahan intraseluler ion kalsium sehingga terjadi apoptosis sel. Oleh karena asam amino eksitasi digunakan secara luas dalam komunikasi kortikal-kortikal, neuron-neuron yang paling berisiko adalah yang menerima hubungan tersebut. Hubungan neuron satu dengan yang lain terputus sehingga tidak mampu memberikan suatu respon behavioral yang bermakna (Posner et al, 1980).
Pada pasien dengan koma dalam, status pupil menjadi satu-satunya kriteria penting yang dapat membedakan antara kelainan metabolik dengan lesi struktural. Refleks pupil yang bertahan, sekalipun terdapat depresi nafas, okulosefalik yang tidak responsif, rigiditas deserebrasi, atau flasiditas motorik, mungkin menunjukkan suatu koma metabolik.
Bola mata sering kali bergerak kemana-mana pada koma metabolik yang ringan dan terfiksasi pada koma dalam. Adanya deviation conjugee atau pergerakan bola mata diskonjugat menunjukkan adanya lesi struktural. Namun gerakan konjugat ke arah bawah atau ke atas dapat terjadi pada koma metabolik maupun koma diensefalik.
Pasien dengan koma metabolik secara umum menunjukkan dua abnormalitas motorik yaitu gangguan tenaga, tonus refleks yang nonspesifik seperti kejang fokal atau general, yang kedua adalah gerakan yang tidak disengaja yang khas. Gerakan bola mata dan aktivitas motorik inipun kurang adekuat untuk membedakan koma metabolik dengan diensefalik (Posner et al, 1980).
Koma Diensefalik
Pada koma diensefalik fungsi korteks serebri adalah baik tetapi karena terdapat gangguan pada ARAS menyebabkan kesadaran tidak bisa dibangkitkan. Koma diensefalik yang dapat ditimbulkan oleh adanya proses patologis supratentorial dan infratentorial. Proses patologis supratentorial akan menimbulkan “pressure cone” yaitu inkarserasi uncus pada incisura tentorial, dimana proses ini dapat disebabkan oleh adanya tumor, hematoma, ataupun abses.
Pada proses patologis infratentorial dapat menimbulkan terjepitnya tonsil serebelli pada foramen magnum. Hal ini biasanya disebabkan oleh infark batang otak rostral, kontusio serebri, tumor serebelli, atau arachnoiditis yang akhirnya akan menimbulkan sumbatan lintasan CSF.
Pada koma diensefalik supratentorial dapat dilihat gejala-gejala sebagai berikut: - Bangkitan epilepsi
- Nyeri kepala
- Sindroma lobus temporalis/ sindroma lobus frontalis - Papil edema
Pada koma diensefalik infratentorial dapat ditemui gejala-gejala sebagai berikut: - Peningkatan tekana intrakranial
- Penurunan kesadaran
- Paralisi nervus kranialis dan defisit neurologi lain yang menunjukkan adanya lateralisasi
- Tidak ditemui gejala sindroma otak organik
3.3 Peranan ARAS dalam Fisiologi Bangun Tidur
Susunan retikularis berhubungan dengan luas ekspresi perilaku kewaspadaan, perhatian hingga siklus tidur. Hilangnya pengaruh susunan retikuler asenden dapat menimbulkan seseorang tertidur. Pusat tidur diperkirakan terdapat di dalam formasio retikularis yaitu diantara pons bagian bawah dan bagian atas dari medulla oblongata. Pada akhirnya, dipostulasikan bahwa tidur disebabkan secara aktif oleh aktivitas perangsangan terhadap pusat tidur di dalam batang otak (deaktivasi retikuler aktif) atau secara pasif oleh penekanan pengaruh susunan retikuler asenden (deaktivasi retikuler pasif). Suatu keadaan koma permanen, akibat adanya cedera otak, mungkin disebabkan oleh kerusakan pada formasio retikularis (hilangnya pengaruh asenden) di dalam jalur retikuler asenden. Lesi pada bagian bawah batang otak mungkin menimbulkan koma yang dalam, sedangkan lesi formasio retikularis batang otak bagian atas mungkin tidak menyebabkan terjadinya koma yang terlalu dalam (Noback, 1993).
Ditinjau dari sudut derajat kesadaran, tidur adalah suatu keadaan dimana derajat kesadaran berada di bawah awas waspada. Namun demikian tidur bukanlah suatu manifestasi gangguan kesadaran. Tidur merupakan keadaan fisiologis yang ditentukan oleh aktivitas bagian-bagian tertentu dari substansia retikularis. Pada saat tidur terjadi suatu proses aktif yang merupakan aktivitas sinkronisasi bagian ventral substansia retikularis medula oblongata. Sedangkan aktivitas bagian rostral batang otak (ARAS) menghilangkan aktivitas tersebut, dengan diadakannya desinkronisasi keadaan tidur diubah menjadi awas waspada.
Walaupun belum ada definisi pasti mengenai tidur, namun hal ini dapat dijelaskan melalui pengamatan sebagai suatu kondisi dengan karakteristik berikut:
b. Munculnya secara spontan aktivitas ritmis otak c. Hilangnya persepsi eksternal
d. Tertutupnya mata e. Hilangnya tonus otot
Pada manusia tidur merupakan hal yang penting untuk memelihara fungsi motorik dan Kognitif. Orang-orang yang mengalami insomnia akan mengalami kesulitan untuk konsentrasi, gangguan daya ingat dan mudah lupa, lelah, depresi, bahkan kadangkala ketidakstabilan emosi. Sebagai tambahan siklus tidur bangun juga mempengaruhi sistem kekebalan tubuh (Shneerson, 2005).
Tidur merupakan aktivitas susunan saraf pusat yang berperanan sebagai “lonceng biologis” yang memperlihatkan irama kehidupan yang sesuai dengan masa rotasi bumi yang dinamakan irama sirkadian (circadian rhythm). Diduga penyebab tidur adalah proses penghambatan aktif. Sebuah teori lama menyebutkan bahwa area eksitatori pada batang otak bagian atas, yang disebut sebagai sistem aktivasi retikuler, mengalami kelelahan setelah seharian terjaga dan oleh sebab itu menjadi tidak aktif. Keadaan ini disebut dengan teori pasif dari tidur. Percobaan lain membuktikan sebuah teori baru bahwa tidur disebabkan oleh suatu mekanisme penghambatan aktif. Hal ini dibuktikan dengan suatu percobaan dengan melakukan pemotongan pada batang otak setinggi regio midpontin dan berdasarkan hasil perekaman listrik otak, ternyata otak tidak pernah tidur. Sehingga dapat disimpulkan bahwa ada beberapa struktur yang terletak di bawah midpontin pada batang otak yang diperlukan untuk menyebabkan tidur dengan cara menghambat bagian-bagian otak yang lain. (Guyton, 1997)
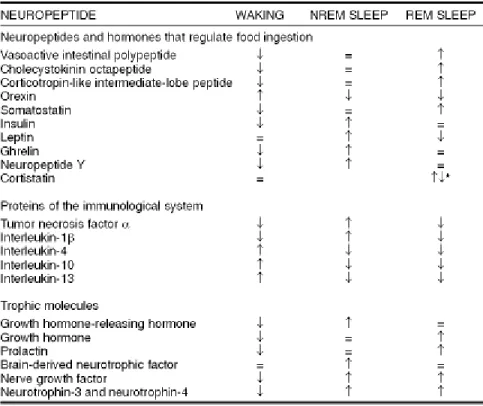
Tabel 1: Neuropeptida dan siklus tidur-bangun Sumber: www. Journals.prous.com
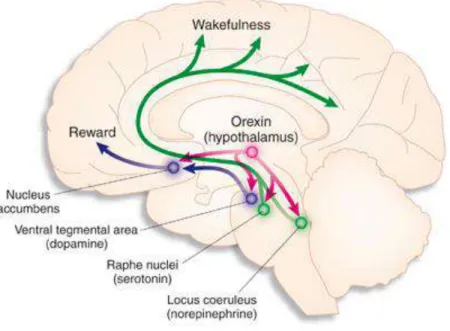
Adapun struktur-struktur yang berkaitan dengan siklus bangun tidur ini, yaitu: 1. Suprachiasmatic nuclei (SCN)
2. ARAS pada midpons dan mesensefalon dan neuron korteks serebri
3. Jalur serotonin yang dihasilkan oleh sistem raphe nuklei yang aktif saat mengantuk dan fase NREM
4. Area parabrachial pons yang menghasilkan asetilkolin yang aktif pada fase REM 5. Locus Ceruleus (LC) sebagai yang merupakan tempat dari susunan saraf
adrenergik yang menghambat susunan saraf serotonergik dari sistem raphe sehingga timbul kembali keadaan terjaga
Suprachiasmatic Nuclei (SCN) merupakan struktur yang sangat kecil berbentuk
sayap, terdiri atas sepasang area sebesar kepala paku yang masing-masing berisi sekitar 10.000 sel saraf. Para peneliti mengatakan bahwa setiap sel saraf pada nukleus ini berfungsi sebagai jam yang menimbulkan letupan iram bertanggung jawab terhadap irama sirkadian dan mempromosikan suatu keadaan bangun dari tidur. Lesi yang mengenai SCN berhubungan dengan gangguan mengantuk yang hebat. Letupan irama sirkadian pada SCN ini mempengaruhi siklus bangun tidur melalui dua proses yaitu:
1. SCN mengatur pelepasan hormon melatonin yang merupakan penginduksi tidur dari pineal body dengan irama sirkadian, dimana hormon tersebut sangat sensitif terhadap kondisi lingkungan, terutama terhadap cahaya. Saat malam, penghambatan SCN terhadap sintesa melatonin menurun sehingga hormon ini akan banyak dikeluarkan dalam sirkulasi darah. Akibatnya melatonin akan menekan aktivitas saraf pada SCN yang terkait dengan aktivasi kortikal dan kondisi bangun.
2. Sedangkan neuron pada SCN yang menjaga kondisi bangun, normalnya saat siang sampai sore hari dengan memproduksi suatu peptida yaitu prokinetisin yang mengaktifkan jalur hypocretin/orexin.
Seseorang mampu tetap berada dalam keadaan terjaga berkat adanya aktivitas sel-sel neuron di sel-seluruh korteks serebri yang secara berkesinambungan oleh penggalak kesadaran yaitu ARAS. Pada malam hari atas pengaruh SCN terjadi pelepasan melatonin oleh glandula pinealis dengan hasil antaranya yaitu serotonin. Serotonin sendiri digunakan oleh sistem nukleus raphae untuk menghambat aktivitas ARAS sehingga timbul rasa mengantuk dan dimulai dengan fase tidur NREM. Serotonin akhirnya memacu sistem kolinergik sehingga tidur memasuki fase REM. Aktivitas kolinergik yang berlebihan dapat memacu kegiatan susunan saraf adrenergik. Manakala aktivitas adrenergik cukup intens maka dapat menghambat kegiatan aktivitas serotonergik dan kolinergik sehingga kegiatan ARAS meningkat kembali dan timbullah keadaan terjaga (Goetz, 1999).
Tidur umumnya terjadi dalam siklus yang teratur dan dipengaruhi oleh dua proses internal yang utama yaitu mekanisme homeostatik dan irama sirkadian. Selama siklus
sirkadian berlangsung, saklar ini bertanggung jawab atas terbuka atau tertutupnya “gerbang tidur”. Dengan bekerjanya nukleus VLPO saat tidur, mereka akan menghambat kerja nukleus yang membangunkan tuberomamillary. Demikian pula yang terjadi sebaliknya pada kondisi bangun nukleus tuberomamillary menghambat kerja VLPO. Hubungan timbal balik ini disebut dengan istilah ‘flip-flop”, dimana posisi sumbu saklar berada di tengah-tengah sehingga kondisi bangun-tidur senantiasa stabil dengan masa transisi yang minimal. Aktivitas otak saat tidur pun dapat direkam dengan EEG. Perubahan gambaran EEG tersebut sesuai dengan tahapan-tahapan tidur.
Gambar 3.5: Struktur otak yang berperan dalam siklus tidur-bangun Sumber:: www. Journals.prous.com
Tahap pertama sesuai dengan keadaan dimana seseorang baru saja terlena. Seluruh otot menjadi lemas, kelopak mata menutupi mata, dan kedua bola mata bergerak bolak-balik ke kedua samping. EEG pada tahap tidur pertama ini memperlihatkan voltage gelombang alpha yang makin menurun frekuensinya (Goetz, 1999).
Keadaan tidur memasuki tahap kedua bila timbul sekelompok gelombang yang berfrekuensi 14-18 siklus per detik pada aktivitas dasar yang berfrekuensi 3-6 siklus per
detik yang dikenal sebagai gelombang tidur (sleep spindle). Bola mata berhenti bergerak tetapi tonus otot masih terpelihara.
Pada tahap tidur yang ketiga, EEG memperlihatkan gelombang dasar yang lambat (1-2 siklus per detik) dengan sekali-kali timbul sleep spindles.
Pada tahap tidur keempat, terlihat hanya gelombang lambat saja tanpa sleep
spindles. Keadaan fisik pada tahap ketiga dan keempat adalah lemah lunglai karena tonus
otot sangat menurun.
Pada tahap kelima, tonus otot kembali meninggi, terutama rahang bawah. Bahkan otot-otot anggota gerak dapat berkejang yang disertai dengan munculnya kembali gerakan bola mata dengan kecepatan yang lebih tinggi. Karena itu tahapan ini dinamai sebagai rapid eye movement sleep (REM sleep) atau paradoxal sleep. Sedangkan tahap tidur pertama sampai keempat gerakan bola mata tidak secepat tahap kelima sehingga dinamakan non-rapid eye movement sleep (NREM sleep). Pada tidur malam yang berlangsung rata-rata 7 jam, kedua macam tidur diatas dapat berselingan 4-6 kali dengan perbandingan 75-80% tidur lelap atau tidur NREM dan 20-25% tidur mimpi atau tidur REM (Goetz, 1999; Guyton, 1997)
Tidur NREM mempunyai kaitan dengan metabolisme amine terutama 5-hydroxitryptamine (serotonin) atau serotonergik, sedangkan REM diatur oleh mekanisme adrenergik (Mardjono dan Sidharta, 2008).
Apabila seseorang kurang cukup menjalani tidur REM, maka esok harinya kecenderungan tingkah laku menjadi hiperaktif, nafsu makan, serta libido yang meningkat. Namun bila fase tidur NREM yang kurang maka esok harinya akan menjadi kurang gesit.
Lesi pada pusat-pusat pencetus tidur dapat menyebabkan keadaan siaga yang terus menerus. Lesi berbatas tegas di nuklei raphae menimbulkan keadaan siaga yang ekstrem. Keadaan ini juga timbul bila ada lesi bilateral pada bagian mediorostral suprakiasma pada hypothalamus anterior. Pada kedua keadaan ini, nuklei retikular eksitatori pada mesensefalon dan pons bagian atas tampaknya terlepas dari hambatan. Sebaliknya, kadang-kadang lesi di bagian anterior hypothalamus dapat menyebabkan timbulnya
keadaan yang sangat siaga sehingga dapat menyebabkan kematian karena kelelahan (Guyton, 1997).
Seseorang yang tidur memiliki sejumlah ciri yang menyerupai penurunan kesadaran yang patologis seperti somnolen, stupor, bahkan koma. Hal-hal tersebut mencakup menguap, menutupnya kedua palpebra, deviasi ke atas, divergensi atau gerakan bola mata, kehilangan tonus muskular, penurunan refleks-refleks tendon, pernapasan yang tidak teratur. Walaupun terbangun dari tidur dalam, seseorang yang normal mungkin kebingungan untuk beberapa saat. Namun demikian seseorang yang tidur masih berespon terhadap stimulus yang tidak biasa, dan terdapat aktivitas mental dalam bentuk mimpi yang meninggalkan jejak ingatan, dimana hal inilah yang membedakannya dengan penderita stupor atau koma.
Hal terpenting yang membedakan kedua kondisi tersebut adalah, pada seseorang yang tidur, jika diberi stimulus dapat terbangun kembali ke kesadaran normal. Ambilan oksigen serebral tidak menurun selama seseorang tertidur, tidak seperti halnya yang terjadi pada koma (Victor M & Ropper,2001).
3.3.1 Patologi Tidur
Berdasarkan pembagian dari Asosiasi Pusat Kajian Gangguan Tidur dan Kajian Psikofisiologi Tidur, gangguan tidur dikelompokan menjadi empat bagian, yaitu:
1. Insomnia, ditandai dengan keluhan sulit untuk memulai tidur, mudah terjaga saat tidur dan bangun pagi ssebelum waktunya. Sehingga sulit untuk memulai dan mempertahankan tidur.
2. Excessive Sleepiness, Excessive Daytime Sleep (EDS), ditandai dengan tidur siang berlebihan, sesekali didapatkan dengan dengan jumlah waktu tidur yang berlebihan (hipersomnia), kesadarannya terganggu, kesiapsiagaan dan koordinasi gerakan menurun setelah bangun tidur.
3. Circadian Sleep-Wake Rhythm Disorders, pada kelainan ini pola tidur dan bangun tidak sesuai dengan irama sirkadian siang-malam. Akibatnya timbul rasa kelelahan,
mengantuk tidak pada waktunya, kesiapsiagaan dan penampilannya menurun setelah bangun tidur.
4. Parasomnia, bentuk kelainan gerakan selama tidur atau kegiatan berlebihan dari fungsi sistem saraf otonom saat tidur atau bangun tidur.
Insomnia
Kebutuhan tidur setiap orang bervariasi dan sesekali waktu mengalami kekurangan tidur yang disebabkan oleh berbagai faktor. Kekurangan tidur yang berlangsung dalam waktu lama dapat dikatakan sebagai suatu insomnia. Insomnia sementara waktu (transient) menghilang dengan sendirinya (kembali normal) dalam waktu kurang dari tiga minggu. Gangguan tidur dapat berupa kesulitan memulai dan mempertahankan tidur secara wajar. Penyebab yang tersering dari transient insomnia ini adalah kecemasan akibat stress mendadak, penyakit fisik dan perjalanan jauh. Insomnia yang sebenarnya adalah gangguan berlangsung lama atau kronis, dibagi menjadi 4 bagian:
1. Insomnia akibat penyakit fisik yang menimbulkan rasa nyeri, sesak, batuk, nokturia dan lain-lain. Keadaan ini sering dialami oleh para lansia.
2. Insomnia akibat masalah psikologik dan psikiatri yang merupakan bagian uatama dan terbesar dari kasus insomnia. Terutama pada penderita dengan gangguan afektif dan gangguan cemas.
3. Insomnia primer, suatu keadaan insomnia yang tidak diketahui secara jelas penyebabnya, namun sepanjang hidupnya penderita mengalami tidur yang terbagi-bagi dalam waktu yang singkat, disertai kelelahan di siang hari, mudah tersinggung, tegang dan depresi somatik. Namun perlu diketahui, ada orang yang kebutuhan tidurnya memang sedikit dan orang tersebut tidak mengalami keluhan meskipun tidur dalam waktu yang singkat.
4. Insomnia akibat penyakit organik., misalnya sleep apneau, parasomnia.
Insomnia yang dapat menimbulkan kematian dilaporkan sebagai fatal familial insomnia, ditandai oleh gejala insomnia yang tidak terobati, terdapat kelainan sistem saraf otonom, kelainan motorik dan didapatkan adanya atrofi inti thalamus. Umur rata-rata penderita
adalah 49 tahun dan lamanya gangguan ini berlangsung hingga meninggal adalah 13 bulan.
Narkolepsi
Berdasarkan kriteria DSM IV, narkolepsi adalah suatu keadaan episode tidur singkat yang rekuren dan tidak terkontrol yang sering disertai halusinasi hipnagogik atau hipnopompik, katapleksi (tonus dan kekuatan otot yang menurun atau menghilang) dan paralisis tidur.
Narkolepsi disebabkan adanya gangguan kontrol dan pengaturan tidur REM, yaitu pada rekaman otak saat tidur didapatkan adanya pemendekan REM Latency yaitu hanya berlangsung selama 10 menit, sedangkan siklus pada orang normal berlangsung hingga 90 menit. Keadaan ini sering diderita oleh sekelompok orang dari satu keluarga yang diduga memilki kelaianan genetika. Pada semua kasus yang dilaporkan, didapatkan setiap kasus memiliki genotip HLA-DR2 dan HLA-DQWI yang berlokasi pada kromosom 6, dengan sedikit pengecualian. Hal ini disebabkan karena sepertiga penduduk di dunia memiliki genotip tersebut, sehingga pemeriksaan tissue typing hanya memiliki nilai eksklusi, bukan untuk menghasilkan diagnosis.
Gangguan Irama Sirkadian Tidur
Sebagian besar keluhan dari penderita gangguan irama sirkadian tidur adalah tidak dapat tidur pada waktunya yang sesuai, sehingga penderita akan mengeluh tidak bisa tidur (insomnia) atau bahkan terus meneus mengantuk. Penyebab yang tersering adalah perubahan jadwal waktu bekerja, kesulitan menyesuaikan dnegan lingkungan baru dan jet
lag. Pergantian jadwal waktu bekerja menimbulkan kesulitan tidur, yaitu pemendekan
waktu tidur dan sering terbangun sehingga menimbulkan sulit berkonsentrasi, kelelahan dan peningkatan angka kecelakaan kerja. Kemampuan penyesuaian dengan jam kerja baru memang sangat individual, namun rata-rata terjadi penurunan setelah usia 40 tahun. Perjalanan jauh melintasi meridian waktu juga dapat menimbulkan gangguan tidur yang serupa dengan perubahan jadwal kerja dan keadaan ini akan diperburuk dengan mengonsumsi alkohol.
Parasomnia
Gangguan tidur ini kebanyakan diderita oleh anak-anak, amat jarang terjadi pada usia dewasa. Gangguan ini dapat dalam bentuk gangguan gerakan tidur (hypnic jerks,
bruxisme dan myoclonus) atau kegiatan berlebihan dari fungsi saraf otonom (detak
jantung yang meningkat, berkeringat, enuresis) ketika tertidur ataupun saat terbangun.
Hypnic jerk adalah gerakan atau hentakan dari anggota gerak atau tubuh pada saat
mulai tidur, dapat disertai dengan menangis, rasa seperti terjatuh atau melihat kilatan cahaya. Gangguan ini tidak sampai mengganggu siklus tidur penderita, tetapi kemungkinan dapat menghambat permulaan tidur (sleep latency). Hypnic jerk sering terjadi sesudah berolahraga dan stress emosi. Keadaan ini masih dianggap normal dan tidak diperlukan pengbatan khusus.
Gangguan perilaku tidur REM terjadi pada saat fase tidur REM dan sebagian besar (60%) bersifat idiopatik, sisanya kemungkinan disebabkan oleh gangguan sistem saraf pusat. Tonus otot yang seharusnya menghilang pada fase tidur REM, pada keadaan ini cenderung meningkat sehingga penderita akan mendadak meloncat dan berlari dari tempat tidur, menggigit atau bertingkah laku kasar. Penderita gangguan ini biasanya adalah manula dekade ke 7 dan pria lebih sering daripada wanita.Kelainan sistem saraf pusat yang mendasari antara lain demensia, gangguan pembuluh darah otak, kerusakan pada batang otak ataupun multipel sklerosis. (Parkers, 1996)
BAB 4 RINGKASAN
- Susunan retikularis adalah susunan traktus-traktus yang bersifat nonspesifik, multineuron, multisinaps), secara relatif difus dan mengintegrasi serta menghantarkan sejumlah impuls-impuls yang bersifat asenden
- Susunan ini terdapat di seluruh susunan saraf pusat, pada medulla spinalis, batang otak, serebellum, diensefalon dan hemisfer serebrum
- Mekanisme ARAS mengaktifkan sistem thalamokortikal disebabkan oleh adanya jalur glutamatergik dorsal dan jalur kolinergik ventral serta jalur lain yang berperan dalam pengaktifan ARAS, yaitu jalur dopaminergik, serotonergik, serta histaminergik - Mekanisme relay sistem limbik oleh ARAS mempengaruhi korteks serebri
merupakan hal yang penting dalam fungsi kesadaran
- Kesadaran memperlihatkan dua segi yaitu derajat kesadaran dan kualitas kesadaran - Ganguan kesadaran yang berat oleh karena gangguan neuron-neuron pengemban
kesadaran disebut dengan koma kortikal bihemisferik
- Gangguan kesadaran yang disebabkan oleh disfungsi ARAS disebut dengan koma diensefalik yang dapat ditimbulkan oleh adanya proses patologis supratentorial dan infratentorial
- Tidur merupakan aktivitas susunan saraf pusat yang berperanan sebagai “lonceng biologis” yang memperlihatkan irama kehidupan yang sesuai dengan masa rotasi bumi yang dinamakan irama sirkadian (circadian rhythm)
- “Lonceng biologis” yang mengatur irama sirkadian diduga terletak pada daerah nuklues suprakiasma hypothalamus yang berkaitan dengan mata.
- Lesi pada pusat-pusat pencetus tidur dapat menyebabkan keadaan siaga yang terus menerus
- Berdasarkan pembagian dari Asosiasi Pusat Kajian Gangguan Tidur dan Kajian Psikofisiologi Tidur, gangguan tidur dikelompokan menjadi empat bagian, yaitu: insomnia, excessive day-time sleepiness, circadian sleep-wake rhythm disorders, parasomnia
DAFTAR PUSTAKA
Arslan, 2001. Neuroanatomical Basis of Clinical Neurology. New York: The Parthenon Publishing Group.
Chusid, J.G. 1991. Neuroanatomi Korelatif dan Neurologi Fungsional. Yogyakarta: Gadjah Mada University Press.
Duus, P. 1996. Diagnosis Topik Neurologi: Anatomi, Fisiologi, Tanda, Gejala. Edisi 2. Jakarta: EGC.
Ganong, W.F. 2002. Buku Ajar Fisiologi Kedokteran. 20th Ed. Jakarta: Penerbit Buku Kedokteran EGC.
Guyton, A.C., Hall, J.E. 1997. Buku Ajar Fisiologi Kedokteran. 9th Ed. Jakarta: Penerbit Buku Kedokteran EGC.
Goetz, CG. 1999. Textbook of Clinical Neurology. Philadelphia: W.B Saunders Company.
Mardjono, M., Sidharta, P. 2006. Neurologi Klinis Dasar. Jakarta: Penerbit Dian Rakyat.
Mendoza E, Foundas. 2008. Clinical Neuroanatomy: A Neurobehavioral Approach. New York: Springer Science&Bussiness Media.Inc.
Nieuwenhuys, et al. 2007. The Human Central Nervous System. Fourth edition. Berlin: Auflage-Springer.
Noback, C.R., Demarest, R.J. 1993. Anatomi Susunan Saraf Manusia. Edisi 2. Jakarta: EGC.
Ngoerah, I.G.N.G. 1991. Dasar-dasar Ilmu Penyakit Saraf. Surabaya: Airlangga University Press.
Parkers, J D.1996. Neurology in Clinical Practice. Principle of Diagnosis and Management Vol.II. Boston: Butter Worth-Hinemann.
Plum, F., Posner, J.B. 1980. The Diagnosis of Stupor and Coma. Third Edition. Philadelphia: F.A.Davis Company.
Shneerson, Jhon M. 2005. Sleep Medicine: A Guide to Sleep and It’s Disorders. 2nd Ed. Oxford: Blackwell Publishing Ltd.
Sukardi, E. 1985. Neuroanatomia Medica. Jakarta: Penerbit Universitas Indonesia.
Victor, Maurice., Ropper, Allan H. 2001. Adam’s and Victor’s Principles of Neurology. 7th Ed. United States of America: The Mcgraw-Hill Companies Inc.