Seisaibo keiko imejingu ni yoru ERK MAPK kasseika dotai no 1saibo teiryo keisoku
Bebas
6
0
0
Teks penuh
(2) 修 士 論 文 ダ イ ジ ェ ス ト 2013年 度 ( 平 成 25年 度 ) 生細胞蛍光イメージングによるERK MAPK活性化動態の1細胞定量計測 政 策 •メディア 研 究 科 修 士 課 程 2 年 新 土 優 樹. 57.
(3) 1 . 序論 M APK. (m ito g e n -a ctiva te d. protein k i n a s e ; 分 裂 促 進 因 子 活 性 イ 匕 タ ン パ ク 質 キ ナ ー ゼ ; M A P キ ナ ー. ゼ ) は 酵 母 か ら 哺 乳 類 ま で 高 度 に 保 存 さ れ た セ リ ン /ス レ オ ニ ン キ ナ ー ゼ で あ り , 増 殖 や 分 化 , ア ポ ト ー シ ス な ど の 細 胞 運 命 決 定 を 制 御 し て い る ( Q i and E lio n , 2 0 05 ) . 哺 乳 類 細 胞 に お い て は , これ ま で に E R K 1 / 2 , J N K , p 38, E R K 3 / 4 , E R K 5 の 5 種 類 の M A P K フ ァ ミ リ ー が 同 定 さ れ て い る ( R o u x and B le n is, 2 0 0 4 ) . そ の 中 で も 最 も 有 名 な の は E R K (extracellular signal-regulated k in a s e ) 1/2で あ り , 古 典. 的 M A P K と も 呼 ば れ て い る ( C h a n g and K a r in ,2 0 01 ) . E R K M A P K カ ス ケ ー ド は R a f iT V E K / E R K か ら な り, 増 殖 因 子 や サ イ ト 力 イ ン , ホ ル ボ ー ル エ ス テ ル 等 に よ っ て 活 性 化 が 誘 導 さ れ る . EGF. (e p id e rm a l. g ro w th. f a c t o r ;上 皮 成 長 因 子 ) 刺 激 に 対 す る E R K の 応 答 は 特 に よ く 調 べ ら れ て お. り, シ ス テ ム 生 物 学 に お け る モ デ ル シ ス テ ム と し て も し ば し ば 用 い ら れ る . E G F の 結 合 に よ っ て E G F R が 活 性 化 さ れ る と , ア ダ プ タ ー タ ン パ ク 質 で あ る G rb 2 ,. G T P 交 換 促 進 因 子 で あ る Sosとの相互. 作 用 を 介 し , Rasが 活 性 化 さ れ る . Rasの 活 性 化 に よ り R a fは 細 胞 膜 へ と 局 在 変 化 し て 活 性 化 さ れ , R a f/M E K /E R K の カ ス ケ ー ド が 順 に 活 性 化 さ れ る .活 性 化 さ れ た E R K は 細 胞 質 か ら 核 へ と 局 在 変 化. し, 転 写 因 子 を は じ め と す る 核 内 の タ ン パ ク 質 を リ ン 酸 化 す る こ と で 細 胞 増 殖 に 導 く. (以 降 , 上. 記 の 経 路 を E G F -E R K 経 路 と 表 記 す る ) .. X 洲 o /m oocytesに お い て は , ERK の リ ン 酸 化 は 入 力 シ グ ナ ル の 上 昇 に 対 し て switch-like に 起 こ る こ と が 知 ら れ て い る (図 1 ) . こ の 非 線 形 性 を 生 み 出 す メ カ ニ ズ ム の 1つ は 2 重 リ ン 酸 化 ;モ チ ー フ で あり, 2 回 の リ ン 酸 化 反 応 が 独 立 ( distributive) に 起 こ る こ と に 起 因 す る ( Ferrell Jr. and Bhatt, 1997;. Huang and Ferrell, 1 9 9 6 ) . さ ら に ,. oocytesの 場 合 は , ERK か ら MAPKKK へ の ポ ジ テ ィ ブ フ. ィ ー ド バ ッ ク が 存 在 す る こ と で , そ の 応 答 は 非 常 に 急 峻 な も の と な っ て い る . ER K は 増 殖 や 分 化 , 発 生 と い っ た 一 般 的 に 不 可 逆 な 過 程 を 制 御 し て い る こ と か ら , そ の 応 答 性 が switch-like で あ る と い う事実は合目的的であると考えられる. 他 方 , 免 疫 染 色 や F A C S に よ る 定 量 解 析 の 結 果 か ら , 酵 母 や 哺 乳 類 細 胞 に お い て は , E R K のリン 酸 化 が 入 力 シ グ ナ ル に 対 し て g ra d e d に 起 こ る こ と が 報 告 さ れ て い る ( C o lm a n -L e m e r e? a/., 2 0 0 5; M a c K e ig a n. a/., 2 0 0 5 ; W h it e h u r s t 以 a/.,2 0 0 4 ) .. E R K の2重 リ ン 酸 化 モ チ ー フ は 哺 乳 類 細 胞 で も 保 存. さ れ て い る こ と か ら , E R K の リ ン 酸 化 ;が s w it c h -lik e で は な く g ra d e d に 起 こ る こ と は 必 ず し も 自 明 で は な い .実 際 の と こ ろ , 足 場 タ ン パ ク 質 ( s c a f f o ld )や 細 胞 内 の 分 子 混 雑 の 影 響 な ど が 示 唆 さ れ て いるが, そのメカニズムは現在でも完全には明らかになっていない( A o k i が 况 ,2 0 1 1 ,2 0 1 3 ). と こ ろ で , ERK は 細 胞 増 殖 や 分 化 と い っ た 細 胞 運 命 決 定 の 制 御 を 担 う 分 子 で あ り , そ の 情 報 は 最 終 的 に は 0 か 1の 情 報 に 変 換 さ れ る 必 要 が あ る と 考 え ら れ る . す な わ ち , gradedな ERK の リ ン 酸 化 も, そ の 下 流 の い ず れ か で switch-like な シ グ ナ ル に 変 換 さ れ る 必 要 が あ る と 考 え ら れ る . し か し な が ら , 実 際 に そ の よ う な メ カ ニ ズ ム が 存 在 す る の か は 明 ら か に な っ て い な い .そ こ で 本 研 究 で は , ERK の リ ン 酸 化 ; 後 に 起 こ る 核 へ の 局 在 変 化 に 着 目 し , そ の ダ ^ ナ ミ ク ス を 生 細 胞 蛍 光 イ メ ー ジ ン グ に よ り 1細 胞 レ ベ ル で 計 測 し た .. 58.
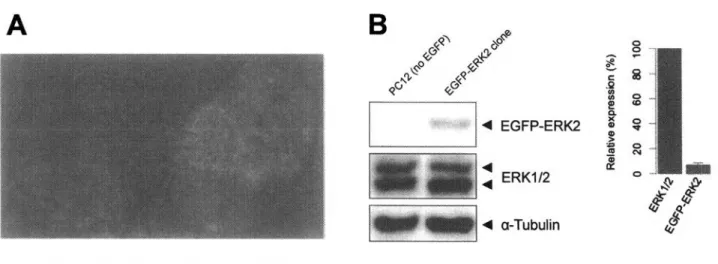
(4) Switch-like response. Graded response. Xenopus oocytes, In Vitro. Yeast, Mammalian cells. Inpyt. Input. 図 1 : E R K の2 重 リ ン 酸 化 の 入 力 シ グ ナ ル へ の依存性 及 /?〇/?奶oocytesや w v / 加 に お い て は , E R K のリン酸化が入力シグナルに対して switch-like に起こる . 一方,酵 母 や 哺 乳 類 細 胞 に お い て は , E R K のリン酸化は入力シグナルの上昇に graded に依存する .. 2•結果と議論 3 .2 .1. E G F P -E R K 2 の 生 細 胞 蛍 光 イ メ ー ジ ン グ. 生 き た 細 胞 内 に お け る E R K の 動 態 を 観 察 す る た め に , E R K 2 にE G F P を 融 合 さ せ た タ ン パ ク 質 (E G F P -E R K 2 )を 用 い た .. また,核 マ ー カ ー と し て TagR FP を 融 合 さ せ た ヒ ス ト ン H 2 B を用 い , こ. れ ら を 安 定 発 現 す る P C 1 2 細 胞 を 使 用 し て 実 験 を 行 な っ た . P C 1 2 は ラ ッ ト の 副 腎 髄 質 ( a d re n a l m e d u lla )由 来 の 褐 色 細 胞 腫 ( P h e o c h ro m o c y to m a )で あ り , E R K を 介 し た 細 胞 運 命 決 定 研 究 の モ デ. ルとして用いられてきた実績がある( G re e n e a n d T is c h le r, 1976; V a u d r y 扣 《/.,2 0 0 2 ). ま ず 始 め に , ウ ヱ ス タ ン プ ロ ッ テ イ ン グ に よ り 内 在 性 E R K とE G F P - E R K 2 の 発 現 量 比 を 見 積 も っ た.. こ れ は , E R K の 過 剰 発 現 は 細 胞 内 局 在 を 変 化 さ せ る こ と が 知 ら れ て お り ( B u ra ck. and. Shaw ,. 2 0 0 5; F u k u d a 以 虬 ,1 9 9 7 ) , 導 入 し た E G F P - E R K 2 の 発 現 量 が 内 在 性 E R K に 比 べ て 十 分 に 低 い こ と を. 確 認 す る 必 要 が あ る た め で あ る . 発 現 量 比 を 見 積 も っ た 結 果 , E G F P -E R K 2 の 発 現 量 は 内 在 性 E R K の 1 0 % 未 満 で あ っ た (図 2 B ) .. 内 在 性 E R K の 発 現 量 は 約 1 u M で あ る こ と か ら ( F u jio k a. 2 0 0 6 ) , E G F P - E R K 2 の 発 現 量 は 100 n M 未 満 で あ る と 見 積 も ら れ る .. d. a/.,. これは, 導 入 し た E R K の発現量. が 150 n M 程 度 以 下 で あ れ ば , 本 来 の 細 胞 内 動 態 に ほ と ん ど 影 響 を 与 え な い と い う 報 告 の 範 囲 内 で ある( C o s t a W f l/ ., 2 0 0 6 ) . 以 上 よ り , E G F P - E R K 2 の 発 現 が 生 細 胞 に お け る E R K の 動 態 を 乱 す 可 能 性 は低いと結論付けた. 次 に , E G F 刺 激 に 対 す る E R K の 核 移 行 応 答 を 調 べ る た め に , 0 .0 00 5 -5 0 n g / m l の E G F で 細 胞 に 刺 激 を 与 え , E G F P - E R K 2 の 核 移 行 夕 V ナ ミ ク ス を イ メ ー ジ ン グ し た . E G F 濃 度 は , 50 , 5 , 1 , 0. 5,. 0.1,. 0.08, 0.06, 0 .05, 0.04, 0 .0 3 , 0.01, 0.005, 0 .0005, 0 n g/m l の 14条 件 で 実 験 を 行 な っ た . 核 マ ー カ ー で あ る T a g R F P -H 2 B の 画 像 か ら 各 細 胞 の 核 領 域 を 決 定 し , 核 内 に お け る E G F P - E R K 2 の 平 均 笛 光 強 度 を 1細. 胞 レ ベ ル で 定 量 化 し た . 核 内 に お け る E G F P -E R K 2 の 蛍 光 強 度 値 は , E G F 刺 激 前 の 値 で 規 格 化 し た (C o h e n -S a id o n e W / . ,2 0 0 9 ) . 各 E G F 濃 度 条 件 に お い て 最 低 170 個 以 上 , 14 条 件 合 計 で 3 ,2 73 個 の 細 胞 に. お け る 核 内 E G F P -E R K 2 の 時 系 列 デ 一 夕 を 取 得 し た .. 59.
(5) A. 図 2 : E G F P -E R K 2 の 観 察 に 使 用 し た P C12 細胞 E G F P -E R K 2 とT a g R F P -H 2 B を 安 定 発現する P C 12細 胞 . ( B ) ウヱ スタンプロッテイングによるE G F P -E R K 2. (A ). と内在性 E R K の 発 現 量 比 の 見 積 も り .. 0 min. 8 min. 2 0 min. 図 3 : E G F P -E R K 2 の生細胞 笛 光 イ メ ー ジ ン グ E G F P -E R K 2 を 安 定 発 現 す る P C12 細 胞 を 50 ng/mlのE G F で刺 激 し , E G F P -E R K 2 の細胞内局在を共焦点レーガ. 一 顕 微 鏡 に よ り 可 視 化 計 測 し た . E G F 刺 激 によ っ て E G F P -E R K 2 が 細 胞 質 か ら 核 へ と 局 在 変 化 す る 様 子 が 1相 胞 レベルで捉 え ら れ て い る . 3.2.2. ER K の 核 移 行 応 答. 論文投稿準備中のため省略.. 3• 蠢 論 論文投稿準備中のため省略.. 4.. 対象と手法. 省 略 .詳 細 は 修 士 論 文 を 参 照 さ れ た い .. 60.
(6) 参考文献 A o k i, K ., Yam a d a , M ., K u n id a , K ., Yasuda, S., and M atsuda, M . (2 0 1 1 ). Processive phosphorylation o f E R K M A P kinase in m a m m a lia n cells. P roc. N a tl. A c a d . Sci. U . S. A .. 108,. 1 2 6 7 5 -1 2 6 8 0 .. A o k i, K ., Takahashi, K . , K a iz u , K ., and M atsuda, M . (2 0 1 3 ). A Q uantitative M o d e l o f E R K M A P K inase Phosphorylation in C ro w d e d M e d ia . S ci. R e p . 1 -8 .. 3,. B u ra ck , W .R ., and Shaw , A .S . (2 0 0 5 ). L iv e C e ll Im a g in g o f E R K and M E K : sim ple b in d in g e q u ilib riu m. 280, 3 8 3 2 -3 8 3 7 . C h a n g , L ., and K a rin , M . (2 0 0 1 ). M a m m a lia n M A P kinase sig n a llin g cascades. N a tu re 410, 3 7 -4 0 . explains the regulated nucle o cyto pla sm ic distribution o f E R K . J. B io l. C h e m .. C o h e n -S a id o n , C ., C o h e n , A . a, Sigal, A . , L iro n , Y ., and A lo n , U . (2 0 0 9 ). D y n a m ic s and v a ria b ility o f E R K 2 response to E G F in in d iv id u a l liv in g cells. M o l. C e ll 8 8 5 -8 9 3 .. 36,. C o lm a n -L e m e r, A . , G o rd o n , A . , Serra, E ., C h in , T ., Resnekov, 0 . , E n d y , D ., Pesce, C .G ., and B re n t, R . (2 0 0 5 ). Regulated c e ll-to -c e ll variation in a cell-fate decision system. N a tu re. 437, 6 9 9 -7 0 6 .. Costa, M ., M a rc h i, M ., C a rd a re lli, F., R o y , A . , B e ltra m , F., M a ffe i, L . , and Ratto, G . M . (2 0 0 6 ). D y n a m ic regulation o f E R K 2 nuclear translocation and m o b ility in liv in g cells. J. C e ll Sci.. 119, 4952-^4963.. Ferrell Jr., J .E ., and B hatt, R .R . (1 9 9 7 ). M ech anistic Studies o f the D u a l Pho sphorylation o f M ito g e n activated Protein K ina se . J. B io l. C h e m .. 272,. 1 9 0 0 8 -1 9 0 1 6 .. Fujioka , A ., Te ra i, K . , Ito h , R .E ., A o k i, K ., N a ka m u ra , T ., K u ro d a , S ., N is h id a , E ., and M atsuda, M . (2 0 0 6 ). D yn a m ics o f the R a s / E R K M A P K cascade as m onitored b y fluorescent probes. J. B io l. C h e m . 8926.. 2 8 1 ,8 9 1 7 -. Fukuda, M ., G o to h , Y ., and N is h id a , E . (1 9 9 7 ). Interaction o f M A P kinase w ith M A P kinase kinase: its possible role in the control o f n ucleocyto plasm ic transport o f M A P kinase. E M B O J .. 1 6 , 1 9 0 1 -1 9 0 8 .. Greene, L . A . , and Tis ch le r, A .S . (1 9 7 6 ). Establishm ent o f a noradrenergic clonal line o f rat adrenal pheochrom ocytom a cells w h ic h respond to nerve g ro w th factor. P roc. N a tl. A c a d . Sci. U . S. A .. 73, 2 4 2 4 -. 2428. H u an g , C .Y ., and F e rre ll, J .E . (1 9 9 6 ). U ltra s e n s itivity in the m itogen-activated protein kinase cascade. Proc. N a tl. A c a d . Sci. U . S. A .. 9 3 , 1 0 0 7 8 -1 0 0 8 3 .. M a c K e ig a n , J.P., M u rp h y , L . O . , D im itr i, C . A . , and B le n is , J. (2 0 0 5 ). G ra d e d m itogen-activated protein kinase activity precedes s w itch -lik e c -F o s induction in m a m m a lia n cells. M o l. C e ll. B i o l . 2 5 ,4 6 7 6 -4 6 8 2 . Q i, M ., and E lio n , E . a (2 0 0 5 ). M A P kinase pathw ays. J. C e ll Sci.. 118, 3 5 6 9 -3 5 7 2 .. R o u x , P.P., and B le n is , J. (2 0 0 4 ). E R K and p38 M A P K -A c t iv a t e d P rotein K in a s e s : a F a m ily o f Protein Kinases w ith D ive rs e B io lo g ic a l Functions. M ic ro b io l. M o l. B io l. R e v.. 68, 3 2 0 -3 4 4 .. V audry, D ., Stork, P .J.S ., L a z a ro v ic i, P., and Eid e n , L . E . (2 0 0 2 ). S ig n a lin g pathw ays fo r P C 12 cell differentiation: m a k in g the rig h t connections. Science. 1 6 4 8 -1 6 4 9 .. 296,. W hitehurst, A . , C o b b , M . H ., and W h ite , M . A . (2 0 0 4 ). S tim u lu s -C o u p le d Spatial Restriction o f E xtra ce llular Signal-R egulated K in a se 1/2 A c t iv it y Contributes to the S p e cificity o f Signal-Response. M o l. C e ll. B i o l . 1 0 1 4 5 -1 0 1 5 0 .. 61. 24,.
(7)
Gambar
Dokumen terkait
Yuntho, Emerson, et al., 2014, Hasil Penelitian Penerapan Unsur Merugikan Keuangan Negara Dalam Delik Tindak Pidana Korupsi Policy Paper Indonesia Corruption Watch 2014,
Girsang, Juniver, 2010, Implementasi Ajaran Sifat Melawan Hukum Materiel Dalam Tindak Pidana Korupsi Dihubungkan Dengan Putusan Mahkamah Konstitusi Republik Indonesia Nomor:
Penerjemahan setia dari kalimat TSu This is why orthodox Islam has never had a Church or a Synod that could dictate what others should and should not believe
Merujuk pada pertimbangan tersebut, maka lokasi yang dapat diwujudkan untuk kebutuhan ruang terbuka hijau publik bagian wilayah kota, diantarnya adalah : Ruang Terbuka
Namun, kata Rizal, ditegaskan kepada Dirut Bank Mandiri ICW Nelloe agar tak boleh ada dana keluar (penarikan dana) sedikit pun.”Dan ternyata dalam waktu
Banyak dana talangan bank century yang tidak sesuai dengan keadaan yang sebenar nya, dalam masalah century ini sebenar nya adalah masalah krisis keuangan yang dialami oleh
(a) Deputi Gubernur BI bidang Pengawasan Perbankan Dra Sti Chalimah Fadjriah, MM sudah mengemukakan di depan rapat KSSK tanggal 20 November 2008 itu bahwa sebaiknya Bank Century
Manajemen perubahan tidak akan berhasil tanpa adanya implemtasi organisasi pembelajaran yang menyediakan ruang untuk selalu improvement, diharapakan menjadi atau
