TINJAUAN PUSTAKA
Tanaman Cabai
Tanaman cabai merupakan tanaman asli dari Amerika yang daerah penyebarannya meliputi Meksiko, Amerika Tengah, dan Amerika Selatan. Columbus adalah orang yang pertama kali memperkenalkan tanaman cabai dan membawa biji yang dijadikan benih ke Spanyol pada tahun 1493, sampai kemudian benihnya banyak ditanam di Eropa. Di Asia, tanaman cabai diperkenalkan oleh bangsa Portugal dan Spanyol pada abad ke - 16, sekarang ini tanaman cabai sudah tersebar di seluruh dunia termasuk Indonesia (Bosland dan Votava 1999).
Di Indonesia cabai (Capsicum annuum L.) adalah sayuran penting yang paling banyak dikenal memiliki nilai ekonomi tinggi. Di Indonesia cabai dapat tumbuh dan berproduksi di dataran rendah dan tinggi, baik di lahan sawah atau tegalan sehingga arealnya mencapai 150,000 ha (Direktorat Jenderal Hortikultura 2008), tetapi produktivitasnya masih sangat rendah. Species Capsicum annuum L. mempunyai keragaman tipe buah yang tinggi untuk (Gambar 2).
Gambar 2. Keragaman tipe buah cabai dalam genotipe species C. annuum L. Sumber : Dremann (2008)
Cabai dalam species Capsicum spp. ini memiliki karakteristik yang luas pada ukuran buah, warna, dan bentuknya, yaitu dengan panjang bervariasi antara 1.5-30 cm, bentuk buah yang sangat lonjong, mengerucut dan bundar, berwarna hijau dan kuning saat belum matang, oranye dan coklat saat sudah matang. Karakteristik tanamannya memperlihatkan keragaman yang tinggi. Keragaman yang tinggi tipe buah (Gambar 2). Banyaknya species cabai dalam genus Capsicum. Genus Capsicum termasuk di antaranya adalah lima spesies yang telah dibudidayakan yaitu Capsicum annuum, C. baccatum, C. chinense, C. frustescens, dan C. pubescens (Greenleaf 1986).
Daftar spesies cabai yang dibudidayakan dan liar serta daerah penyebarannya disajikan pada Tabel 1. Tanaman cabai berbentuk semak dengan batang berkayu dan tipe
percabangan tegak atau menyebar dengan karakter yang berbeda-beda tergantung spesiesnya. Tinggi tanaman cabai berkisar 30–75 cm. Daunnya berwarna hijau dan atau hijau tua, tumbuh pada tunas-tunas samping berurutan, pada batang utama dan tunggal tersusun secara spiral. Daunnya berbentuk hati lonjong atau bulat telur dengan letak yang berselang-seling (Poulus 1994).
Tabel 1. Daftar spesies cabai yang telah dibudidayakan, tipe liarnya serta daerah penyebaran
Spesies Status Daerah sebaran
C.annuum L. Dibudidayakan
Amerika Selatan hingga
Colombia tropik, subtropik dan daerah beriklim sedang
C. chinense Jacq. Dibudidayakan Dataran rendah Amerika Selatan bagian timur
C. frutescens L. Dibudidayakan Amerika tropik
C.baccatum L. Dibudidayakan Peru, Bolivia, Paraguay, Brazil, Argentina
C. praetermisum Heiser & Smith. Liar Brazil Selatan
C. chacoense A. T. Hunz. Liar Argentina Utara, Bolivia Paraguay,
C. galapagoense A. T. Hunz. Liar
Daerah Andes, dataran tinggi Amerika Tengah bagian utara hingga Meksiko
C. pubescens R & P. Dibudidayakan
Daerah Andes, dataran tinggi Amerika Tengah bagian utara hingga Meksiko
C. cardendaii Heiser & Smith. Liar Bolivia
C. eximium A.T.Hunz. Liar Bolivia, Argentina utara C. tovarii Eshbaugh, Smith,
Nickrent. Liar Andes, Peru tengah
C. lanceolatum. Liar Guetamala
Sumber: Greenleaf (1986)
Menurut Kusandriani (1996) bunga cabai termasuk bunga lengkap, yaitu terdiri atas kelopak dan mahkota, Daun-daun mahkota yang berlekatan menjadi satu sehingga digolongkan dalam sub-kelas Sympetalae. Bunga tanaman cabai mempunyai bunga tunggal atau soliter dan tumbuh pada ujung ruas, serta merupakan bunga sempurna. Alat kelamin jantan dan betina terdapat pada satu bunga. Mahkota bunga berwama putih atau ungu tergantung pada kultivarnya, helaian mahkota bunga berjumlah lima atau enam. Pada dasar bunga terdapat daun bunga berjumlah lima helai kadang-kadang bergerigi. Setiap bunga mempunyai satu putik, kepala putik berbentuk bulat. Bunga cabai terdiri dari lima petal, lima sepal, satu putik, dan lima benang sari yang fertile.
Bunga cabai termasuk menyerbuk sendiri tetapi dapat terjadi penyerbukan silang dengan bantuan lebah atau serangga lainnya dengan persentase persilangan berkisar 7.6-36.8%. Bunga tanaman cabai cenderung bersifat protogyny, yaitu kepala putik telah masak sebelum tepung sari keluar dari kotak sari atau sebelum anthesis, dan tepung sari keluar pada saat bunga mekar (Greenleaf 1986).
Menurut Kusandriani (1996) persilangan cabai sering terjadi pada bunga yang mempunyai tangkai putik yang panjang dan kepala putik lebih tinggi dari kotak sari, sedangkan penyerbukan sendiri sering terjadi pada bunga yang memiliki tangkai putik yang pendek sehingga letak kepala putik lebih rendah dari kepala sari. Bunga tunggal terdapat pada setiap ruas dan pada saat antesis tangkai bunga umumnya merunduk. Bunga pertama biasanya terbentuk pada umur 23-31 hari sesudah tanam (HST) dan buah pertama biasanya mulai terbentuk pada umur 29-40 HST.
Benih Cabai
Benih cabai dihasilkan dari buah yang matang dalam waktu 34-40 hari setelah pembuahan. Umumnya memiliki biji cabai berwarna kuning jerami (Hernandez 2002). Proses pemanenan cabai mempengaruhi mutu benihnya baik viabilitas maupun vigornya. Beberapa hal yang perlu diperhatikan pada saat pemanenan cabai adalah ciri dan umur panen, cara panen, periode panen dan perkiraan produksi. Pemanenan pada saat masak fisiologis adalah yang terbaik karena pada saat itu vigor benih yang maksimum. Cabai dipanen pada saat buah memiliki bobot maksimal, buahnya padat dan warnanya 90% tepat merah menyala (Kusandriani 1996).
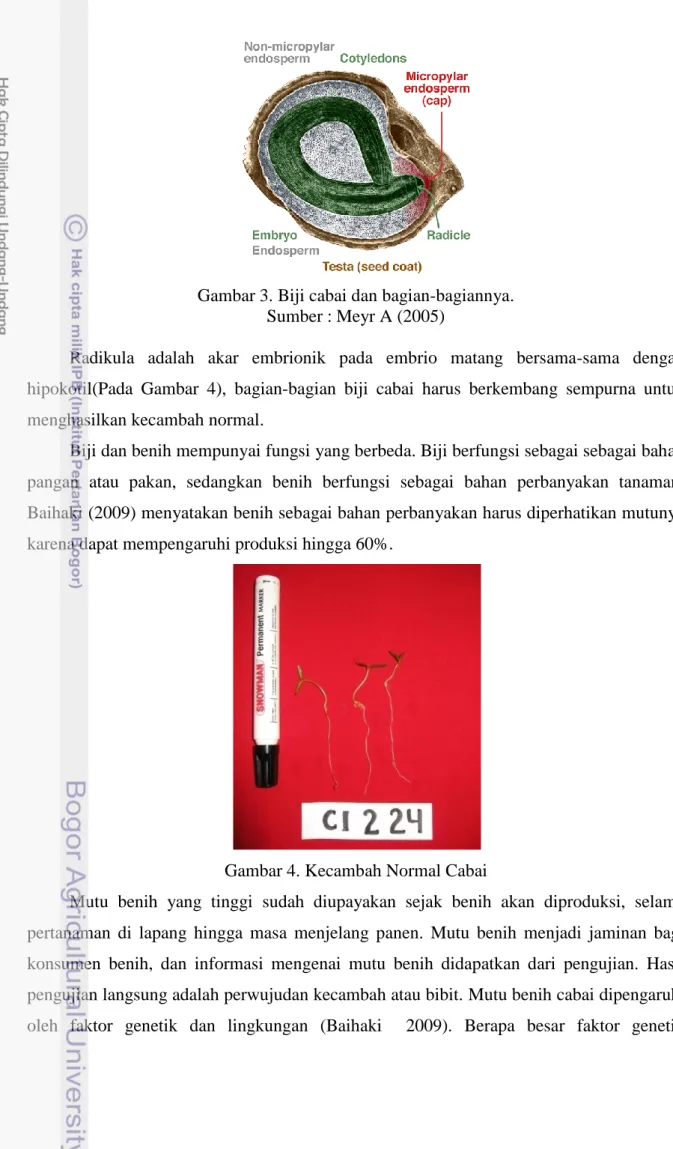
Benih cabai terdiri atas enam bagian (Gambar 3) yaitu endosperm, mikrofil, kotiledon, embrio, testa (seed coat), dan radikula. Endosperm adalah jaringan penyimpan cadangan makanan genomnya berasal dari maternal. Mikrofil adalah saluran atau lubang yang menutup kulit benih, pada nuselus melalui tabung polen yang biasanya dimiliki selama fertilisasi. Pada saat benih matang dan mulai berkecambah mikrofil membantu untuk masuknya air. Biasanya tonjolan radikula melalui mikrofil endosperm. Kotiledon biasanya disebut sebagai daun benih. Embrio adalah sporofit muda hasil fertilisasi. Hipokotil adalah batang yang mirip dengan aksis embrionik di bawah kotiledon. Embrio matang terdiri atas kotiledon, hipokotil dan radikel. Testa adalah lapisan pelindung luar benih yang dikembangkan dari integument pada ovul Meyr (2005).
Gambar 3. Biji cabai dan bagian-bagiannya. Sumber :Meyr A (2005)
Radikula adalah akar embrionik pada embrio matang bersama-sama dengan hipokotil(Pada Gambar 4), bagian-bagian biji cabai harus berkembang sempurna untuk menghasilkan kecambah normal.
Biji dan benih mempunyai fungsi yang berbeda. Biji berfungsi sebagai sebagai bahan pangan atau pakan, sedangkan benih berfungsi sebagai bahan perbanyakan tanaman. Baihaki (2009) menyatakan benih sebagai bahan perbanyakan harus diperhatikan mutunya karena dapat mempengaruhi produksi hingga 60%.
Gambar 4. Kecambah Normal Cabai
Mutu benih yang tinggi sudah diupayakan sejak benih akan diproduksi, selama pertanaman di lapang hingga masa menjelang panen. Mutu benih menjadi jaminan bagi konsumen benih, dan informasi mengenai mutu benih didapatkan dari pengujian. Hasil pengujian langsung adalah perwujudan kecambah atau bibit. Mutu benih cabai dipengaruhi oleh faktor genetik dan lingkungan (Baihaki 2009). Berapa besar faktor genetik
mempengaruhi mutu benih belum banyak informasinya, karena pada umumnya faktor lingkungan banyak mempengaruhi mutu benih.
Terdapat kriteria tipe kecambah normal yang telah ditentukan oleh International Seed Testing Association (ISTA 1999). Pada cabai, kriteria kecambah normal (Gambar 4) yaitu akar primer tumbuh dan berkembang dengan baik, jaringan pembuluh berkembang dengan baik dan tidak terdapat kerusakan; plumula telah tumbuh, plumula harus tumbuh utuh serta berwarna hijau, tumbuhnya boleh melengkung asal tidak busuk; kecambah kelihatan sehat, atau tidak ada kerusakan.
Vigor Benih
Vigor benih adalah kemampuan benih untuk tumbuh dan berkembang menjadi tanaman normal, meski kondisi alam tidak optimum atau suboptimum. Benih yang vigor akan menghasilkan produk di atas normal kalau ditumbuhkan pada kondisi optimum (Sadjad et al. 1999). Kondisi suboptimum adalah kondisi alam terbuka berupa biosfer yang mengganas, cuaca yang tidak akrab, tanah yang tidak subur, pengairan yang tidak menunjang, semua keadaan itu dapat terjadi. Benih vigor yang mampu menumbuhkan tanaman normal pada kondisi alam sub optimum dikatakan memiliki kekuatan tumbuh.
Permasalahan vigor benih
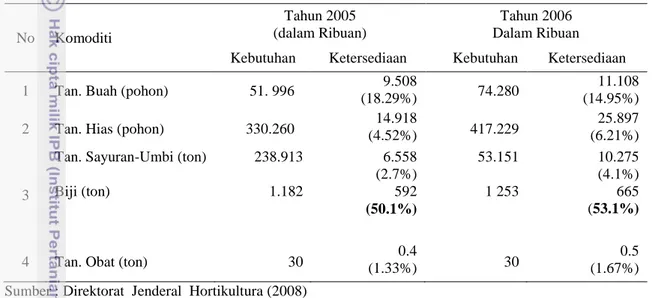
Ketersediaan benih tanaman sayur dan umbi-umbian masih sangat rendah. Yaitu 4.1% (Direktorat Jenderal Hortikultura Departemen Pertanian 2008). Masalah lain adalah rendahnya mutu benih.
Benih tidak selalu segera ditanam, sehingga mengalami penundaan tanam artinya mengalami penyimpanan. Benih yang diproduksi di daerah tertentu mengalami jarak tempuh yang panjang hingga beberapa hari.
Pada tataniaga pertanian benih cabai selalu mengalami penyimpanan pada kondisi sub optimum sebelum sampai ke tangan petani. Penyimpanan pada kondisi sub optimum merupakan kondisi penyimpanan yang kurang baik, karena menyebabkan terjadinya penurunan mutu benih cabai baik viabilitas maupun vigornya sebelum ditanam.
Teknologi penyimpanan yang baik dapat mencegah atau memperlambat kemunduran benih. Benih yang mempunyai viabilitas awal tinggi (mutu benih tinggi) akan memiliki daya simpan yang baik dibandingkan dengan benih yang mempunyai viabilitas awal yang
rendah (mutu benih rendah). Pada benih cabai menurunnya vigor benih ditunjukkan oleh menurunnya (panjang radikula, panjang hipokotil, daya berkecambah benih, kecepatan tumbuh benih, indeks vigor dan terjadi peningkatan nilai daya hantar listrik (Tabel 4). Hal ini didukung oleh Copeland dan McDonald (2001) bahwa menurunnya kualitas benih yang mengakibatkan menurunnya vigor benih dan akhirnya dapat menurunkan hasil.
Tabel 2. Kebutuhan dan Ketersediaan Benih Horikultura Bermutu Tahun 2005-2006
No Komoditi
Tahun 2005 (dalam Ribuan)
Tahun 2006 Dalam Ribuan Kebutuhan Ketersediaan Kebutuhan Ketersediaan
1 Tan. Buah (pohon) 51. 996 9.508
(18.29%) 74.280
11.108 (14.95%) 2 Tan. Hias (pohon) 330.260 (4.52%) 14.918 417.229 (6.21%) 25.897
3
Tan. Sayuran-Umbi (ton)
Biji (ton) 238.913 1.182 6.558 (2.7%) 592 (50.1%) 53.151 1 253 10.275 (4.1%) 665 (53.1%)
4 Tan. Obat (ton) 30 (1.33%) 0.4 30 (1.67%) 0.5 Sumber : Direktorat Jenderal Hortikultura (2008)
Hubungan Vigor benih, Viabilitas dan Deteriorasi
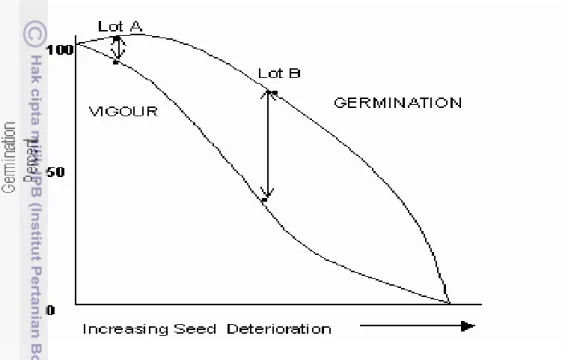
Tiap tahapan benih menggambarkan perubahan pada morfologi dan fisiologi ontogeny yang dapat mengubah potensi penampilan benih. Pada saat benih telah mencapai berat kering maksimum disebut dengan masak fisiologis, pada titik ini, mempunyai potensi yang lebih besar untuk pengecambahan dan vigor yang maksimum (Delouche 1974). Delouche dan Caldwell (1960) menyatakan bahwa pada saat vigor maksimum persentkecambahan mencapai maksimum (100%). Benih lot A adalah benih pada saat masak fisiologis dipanen. Setelah mengalami peningkatan deteriorasi persentase perkecambahan mengalami penurunan secara cepat. Pada umumnya benih mencapai masak fisiologis pada tingkat kadar air tinggi dan tidak aman untuk penyimpanan (Gambar 5).
Benih yang dipanen tidak mencapai masak panen, maka daya simpannya rendah dan tidak dapat meminimalisir kerusakan mekanik. Vigor awal benih mempengaruhi kecepatan deteriorasi dan kecepatan penurunan percent germination. Besarnya sudut penurunan persentase perkecambahan juga berbeda antara benih lot A dan B yang dipanen pada kondisi kemasakan berbeda (Gambar 5).
Pemulia tanaman selama bertahun tahun sangat hati-hati dalam menyeleksi vigor benih. Untuk meningkatkan produksi, pemulia tanaman memperbaiki beberapa karaktetaristik benih seperti integritas mekanik (benih yang keras), resisten terhadap penyakit, kandungan protein, dan ukuran benih. Faktor tersebut berperan pada pertumbuhan di lapang dan sering menghasilkan penambahan daya hasil.
Gambar 5. Hubungan antara vigor benih, viabilitas dan deteriorasi (Delouche dan Caldwell 1960)
Pemulia tanaman telah memperkenalkan ketegaran hibrid benih hibrid vigor untuk kekerasan benih, pengaruh kerusakan benih, dan komposisi benih yang mempengaruhi ekspresi mutu benih. Ketegaran hibrid merupakan komponen heterosis dan menggambarkan pengukuran keunggulan hibrida yang melebihi tetua inbrednya. Keunggulan hibrida sering kali lebih besar pada kondisi cekaman dibandingkan kondisi optimum. Misalnya, benih hibrida jagung dan barley berkecambah dan tumbuh lebih cepat dibandingkan tetua inbrednya (Copeland dan McDonald 2001).
Pemulia tanaman telah menemukan sistem gen yang mengendalikan kualitas nutrisi tetapi tidak diwariskan pada vigor benih. Nass dan Crane (1970) menemukan bahwa berbagai gen untuk ekspresi endosperma mempengaruhi pengecambahan benih pada suhu 15, 20, dan 25oC. Benih dengan gen A1 menghasilkan benih yang lebih vigor dibandingkan dengan benih yang tidak mengandung gen tersebut.
Vigor Daya Simpan Benih
Daya simpan (DS) benih adalah prakiraan waktu berapa lama benih mampu untuk disimpan. DS merupakan parameter lot benih dalam satuan waktu suatu periode simpan (PS). Periode simpan ialah kurun waktu simpan benih, dari benih siap disimpan sampai benih siap ditanam (Sadjad et al. 1999).
Berdasarkan daya simpan benih Robert (1973) menyatakan ada dua tipe benih yaitu benih ortodoks dan rekalsitran. Benih ortodoks adalah benih yang dapat dikeringkan pada kadar air benih (KA) rendah yaitu sampai 5% tanpa kerusakan dan benih orthodiks tersebut toleran pada suhu dingin. Benihrekalsitran adalah benih yang tidak dapat dikeringkan pada KA < 30% tanpa kerusakan dan benih rekalsitran tidak dapat toleran pada suhu dingin.
Sehubungan dengan daya simpan benih terdapat beberapa faktor yang mempengaruhi daya simpan benih, yaitu faktor internal dan eksternal. Menurut Copeland dan McDonald (2001) faktor internal daya simpan benih yaitu ukuran benih, umur simpan benih dan komposisi kimia benih. Faktor eksternal menurut Sadjad (1999) adalah faktor lapangan mulai benih ditanam, pertumbuhan tanaman, pemasakan, pemanenan, pengolahan sampai benih siap disimpan dan kondisi penyimpanan serta lamanya benih disimpan. Kelembaban nisbi dan suhu dapat mempengaruhi daya simpan benih. Pada RH mencapai 80% dan suhu 25–30oC, benih sayuran kehilangan viabilitas dan vigornya.
Copeland dan McDonald (2001) menyatakan benih sayuran dikarakterisasi mempunyai periode simpan pendek contohnya lettuce, bawang dan rye. Benih cabai tersebut termasuk dalam benih yang mempunyai periode simpan pendek. Menurut Hernandez (2002) bahwa daya simpan benih cabai sekitar 3–4 tahun tetapi dalam kondisi penyimpanan optimum pada temperature 10°C dan kelembaban nisbi (RH) 45% dan terkontrol.
Copeland dan McDonald (2001) menurunnya viabilitas dan vigor benih sayuran apabila disimpan pada suhu kamar (kondisi RH 80% dan suhu 25–30oC) akan menyebabkan KA benih sayuran meningkat dan aktifnya peristiwa biokimia seperti aktivitas enzim hidrolik, peningkatan respirasi dan asam lemak bebas, dan cepat terjadi penurunan mutu benih (deteriorasi). Terdapat kaitan cukup erat antara kadar air benih dan kelembaban untuk menurunkan viabilitas dan vigor.
Pengujian Vigor Benih
McDonald (1980) menjelaskan bahwa karakteristik pengujian vigor benih sebaiknya memenuhi kriteria sebagai berikut (1) tidak mahal karena keterbatasan dana untuk pengujian benih, (2) cepat, setiap laboratorium benih mempunyai periode aktivitas maksimum, (3) tidak rumit, prosedur pengujian vigor harus sederhana, (4) obyektif, untuk pengujian vigor lebih mudah dengan standarisasi, (5) dapat diulang, dan (6) korelasi dengan penampilan di lapang.
Standarisasi pengujian vigor benih sangat sulit dilakukan karena kondisi alam yang bervariasi. Vigor daya simpan adalah untuk menduga seberapa lama periode simpan benih. Pengujian vigor daya simpan benih umumnya dilakukan dengan simulasi. Simulasi pengujian vigor daya simpan benih dilakukan dengan metode pengusangan cepat. Benih diperlakukan dalam kondisi suboptimum (cekaman) buatan untuk menduga kondisi simpan sebenarnya misalnya suhu tinggi, kelembaban (RH) tinggi, kimia (etanol, metanol, NaOH, PEG), air panas. Hasil pendugaan akan dihubungkan dengan dugaan lamanya periode simpan benih tersebut (Sadjad et al. 1999).
Benih vigor adalah suatu produk teknologi yang melalui upaya pemuliaan genetik dan pemurnian fisik dapat menghasilkan satu lot benih berisi individu-individu prima yang tinggi tingkat kemurnian genetiknya, bersih penampilan fisiknya, sehat pertumbuhannya, dan homogen kinerja pertumbuhannya di lapang. Usaha pendekatan simulatif untuk menduga vigor benih harus dilakukan melalui jalur ilmu fisiologi, biokimiawi, matematika dan statistika. Semua pendekatan itu dilakukan untuk mendapatkan pendugaan vigor benih yang akurat, karena kompleksnya ilmu tentang vigor benih (Sadjad et al. 1999).
Beberapa metode pengujian vigor daya simpan benih cabai yang dikembangkan pada penelitian ini adalah metode pengujian vigor yang sudah divalidasi ISTA (International Seed Testing Association 2001) adalah (1) Konduktiviti test (Daya hantar listrik) pada benih kacang kapri (Pisum sativum L.) dan (2) metode pengusangan cepat/ accelerated aging (AA) pada benih kedelai (Glycine max L.).
Umumnya hasil pengujian benih di laboratorium dapat dipakai kembali hasilnya pada sampel benih yang sama dalam selang kepercayaan yang dapat diterima. Akan tetapi, pada pengujian yang sama yang dilakukan oleh laboratorium yang berbeda, sering menimbulkan keragaman. Ada beberapa kemungkinan untuk menjelaskan kekurangan standarisasi antara laboratorium (AOSA 1983).
Metode pengujian vigor benih diperlukan metode standar sebagai metode pembanding. Salah satu metode standar pengujian vigor adalah pengujian elektro konduktivitas untuk kacang kapri (Pisum sativum L.). Pengujian konduktivitas test adalah pengukuran terhadap konduktivitas elektrik memberikan penilaian mengenai tingkat kebocoran elektrolit jaringan tanaman. Benih yang mempunyai tingkat kebocoran elektrolit tinggi (konduktivitas tinggi) mempunyai vigor rendah, sedangkan benih yang memiliki kebocoran elektrolit rendah (tingkat konduktivitas rendah) mempunyai vigor tinggi (ISTA 2007).
Lot benih yang mempunyai vigor tinggi akan mampu bertahan pada kondisi ekstrim dan proses deteriorasi (penuaan) lebih lambat dibandingkan dengan lot benih dengan vigor rendah. Sehingga setelah perlakuan pengusangan cepat (AA/Accelerated aging) lot benih yang mempunyai vigor tinggi akan tetap memiliki daya berkecambah tinggi, sedangkan lot benih yang mempunyai vigor rendah daya berkecambahnya akan berkurang. Pengujian AA merupakan suatu pengujian vigor untuk kedelai yang berhubungan dengan daya tumbuh dan daya simpan (Sadjad et al.1999).
Tujuan penyimpanan benih adalah (1) untuk memelihara stok pertanaman dari satu musim sampai musim berikutnya, (2) untuk mempertahankan mutu benih selama periode panjang yang memungkinkan, (3) memberikan jaminan ketersediaan benih pada tahun saat mutu benih masih diterima dan saat produksinya rendah, (4) memungkinkan mempertahankan plasma nutfah melebihi waktunya untuk perbaikan program pemulian tanaman. Kondisi penyimpanan benih kebanyakan spesies mungkin aman disimpan selama beberapa tahun pada suhu dan RH terkontrol. Meskipun kondisi tersebut lebih mahal untuk kebanyakan lot benih pertanian, tetapi sangat berharga untuk memelihara plasma nutfah dan stok benih bernilai tinggi (Copeland dan Mc Donald 2001).
Daya berkecambah, kadar air awal, suhu dan RH lingkungan penyimpanan berpengaruh besar pada derajat deteriorasi benih, karena deteriorasi benih sesuai model persamaan matematik (Roberts 1986). Prinsip umumnya adalah kadar air benih yang rendah, disimpan di bawah penanganan kondisi kering dan dingin mutu benihnya lebih baik dibandingkan kadar air tinggi dengan kondisi lembab dan panas. Robert (1986) mengembangkan persamaan sebagai berikut :
KE = CW log m – CH – CQt2 V = Ki – p/10
V = kemungkinan persentase kemampuan berkecambah setelah periode simpan perhari
Ki = kemungkinan kemampuan berkecambah awal lot benih KE, CW, CH dan CQ = konstanta spesies
m = Kadar Air benih pada bobot basah t = suhu penyimpanan (oC)
Bewley dan Black (1982) mengidentifikasi mutu benih di penyimpanan disebabkan beberapa faktor yaitu (1) kultivar dan keragaman panen, (2) kondisi sebelum dan pasca panen, (3) tekanan oksigen berpengaruh selama penyimpanan, (4) kondisi lingkungan yang fluktuatif atau berubah-ubah.
Wilson dan Mc Donald (1989) memprediksi menggunakan benih Phaseolus vulgaris. Hasilnya bahwa kadar air sangat berpengaruh pada deteriorasi benih dan berhubungan dengan sifat fisiologis benih serta merupakan faktor utama yang menyebabkan benih mengalami deteriorasi selama di penyimpanan.
Parameter Genetik Vigor Benih
Metode Persilangan Dialel
Metode persilangan dialel adalah seluruh kombinasi persilangan yang mungkin diantaranya sekelompok genotipe atau tetua, termasuk tetua itu sendiri lengkap dengan F1 turunannya. Tujuan dari persilangan dialel vigor benih cabai adalah untuk mengevaluasi dan menyeleksi benih tetua yang menghasilkan keturunan terbaik. Genotipe-genotipe tersebut bisa berupa benih, individu, klon atau galur homozigot. Dalam persilangan ini jumlah genotipe yang mungkin dilakukan bisa sangat besar, sehingga membutuhkan ruang, biaya dan tenaga yang lebih besar. Untuk itu maka persilangan tersebut dapat disederhanakan dengan maksud meniru populasi kawin acak (Griffing 1956).
Beberapa metode persilangan dialel yang mungkin dilakukan: metode I (Full diallel) yaitu persilangan yang terdiri dari parent F1 tanpa resiprokal, metode II yaitu persilangan yang terdiri dari tetua dan F1 tanpa resiprokal, metode III yaitu persilangan yang terdiri dari F1 dan resiprokal, metode IV yaitu persilangan yang terdiri dari hanya F1 tanpa resiprokal (Griffing 1956; Roy 2000).
Metode persilangan dialel yang digunakan adalah metode II yaitu persilangan yang terdiri dari tetua dan F1 tanpa resiprokal dengan analisis [n(n+1)/2]. Persilangan setengah
dialel (half diallel atau partial diallel) dibuat agar masing-masing tetua mewakili jumlah persilangan yang sama. Jika terdapat n tetua dan masing-masing tetua meliputi s persilangan, maka jumlah persilangan adalah [(ns)/2] (Griffing 1956).
Semakin banyak tetua pada persilangan dialel, semakin banyak pula jumlah persilangan sehingga kemungkinan kesulitan dalam menangani tempat, waktu, dan tenaga. Penggunaan teknis analisis silang dialel memiliki beberapa keuntungan tersebut yaitu; (1) secara eksprimental merupakan pendekatan sistem sistematik; (2) secara analitik merupakan evaluasi genetik menyeluruh yang berguna dalam mengidentifikasi persilangan bagi potensi seleksi yang terbaik pada awal generasi (Khan dan Habiab 2003). Silang dialel juga dimungkinkan untuk memilih tetua dan memberikan informasi tentang daya gabung tetua dalam hibrida sehingga dapat membantu pemulia untuk meningkatkan dan menyeleksi populasi segregan. Menurut Dudley et al.(1999) analisis dialel kemungkinan dilakukan penilaian daya gabung dan pendugaan komponen ragam serta parameter genetik. Oleh karena itu untuk pendugaan parameter genetik vigor benih cabai digunakan analisis silang dialel.
Dalam analisis silang dialel, pendugaan parameter genetik sudah dapat dilakukan pada F1, tanpa harus membentuk populasi F2, BCP1 ataupun BCP2 seperti pada teknik pendugaan parameter genetik lainya. Akan tetapi dalam pelaksanaanya analisis ini harus memenuhi beberapa asumsi berikut:(1) merupakan segregasi diploid, (2) tidak terdapat pengaruh tetua (tidak ada perbedaan persilangan resiprokal), (3) tidak ada interaksi antara gen-gen yang tidak satu alel (independen), (4) tidak ada peristiwa multiple alel, (5) tetua bersifat homozigot, (6) gen-gen menyebar secara bebas diantara tetua (Hayman 1954; Singh dan Chaudhary 1979; Roy 2000).
Ploidi tanaman cabai adalah diploid (Greenleaf 1986) dengan demikian segregasi gen-gen yang terjadi merupakan segregasi diploid. Perbedaan antar persilangan resiprokal menandakan bahwa ada pengaruh tetua betina. Hal ini merupakan petunjuk bahwa pewarisan suatu karakter diwariskan oleh gen-gen ekstra kromosomal (Mather dan Jinks 1971). Adanya interaksi antara gen-gen yang tidak satu alel dalam analisis silang dialel dapat diuji dengan nilai konfersi regresi b dari garis regresi antara Wr (Peragam antara tetua dan keturunan dari array ke-r) terhadap Vr (ragam di dalam array ke-r). Jika ini b=1 maka tidak ada interaksi antara gen-gen tidak sealel (Singh and Chaudhary 1979). adanya beberapa alel yang mengendalikan suatu karakter akan menyulitkan analisis silang dialel,
Berdasarkan analisis silang dialel menggunakan Metode II Grifing akan diperoleh informasi tentang daya gabung umum (DGU) dan daya gabung khusus (DGK). Selain itu juga dapat diperoleh informasi tentang efek heterosis dan informasi tersebut sangat penting dalam suatu program pemuliaan tanaman (Griffing 1956).
Informasi genetik yang diperoleh dari pengujian DGU dan DGK akan berguna untuk menentukan tetua dan metode pemuliaan yang sesuai dalam rangka perbaikan sifat. Kusandriani (1996) melaporkan adanya efek heterosis sifat kualitas dan daya hasil, serta ketahanan terhadap hama dan penyakit pada tanaman cabai hal ini memberi peluang untuk pembentukan varietas hibrida yang akan menghasilkan sifat yang baik daripada varietas galur murni. Menurut Ahmed et al. (2003): Sujiprihati et al. (2007) metode analisis dialel sudah banyak di manfaatkan untuk mempelajari dasar genetik suatu karakter pada tanaman cabai.
Daya Gabung Galur Murni
Daya gabung merupakan uji keturunan (progeny test), yaitu suatu ukuran kemampuan tanaman dalam persilangan untuk menghasilkan tanaman yang unggul. Evaluasi daya gabung terutama dibuat untuk pembentukan kultivar hibrida F1, yaitu memilih tetua-tetua atau genotipe yang dijadikan tetua hibrida/sintetik (Hermiati 2001).
Daya gabung terdiri atas daya gabung umum (DGU) dan daya gabung khusus (DGK). DGU merupakan ukuran performa keturunan suatu genotipe yang disilangkan dengan contoh acak atau genotipe dengan jumlah besar. DGU diekspresikan pada keturunan persilangan suatu galur murni dengan beberapa genotipe dan terutama merupakan hasil aksi gen aditif. DGK merupakan ekspresi performa di antara dua galur murni dan ditujukan untuk aksi gen dominan, epistasis dan aditif. DGK merupakan ukuran performa keturunan suatu genotipe yang disilangkan dengan genotipe lainnya dan sering diekspresikan sebagai simpangan performa yang diduga dengan rata-rata atau daya gabung umum (Stoskopf et al. 1993). Populasi yang telah diidentifikasi memiliki DGU tinggi sering berpeluang memiliki DGK yang tinggi pula. DGU dan DGK menjadi penting dalam identifikasi tetua yang akan digunakan dalam pembentukan varietas hibrida (Welsh 1981).
Heritabilitas
Heritabilitas adalah potensi suatu individu untuk mewariskan karakter tertentu pada keturunannya. Heritabilitas merupakan rasio ragam genetik terhadap ragam fenotipeik dari suatu karakter. Heritabilitas dibagi menjadi dua, yaitu heritabilitas dalam arti luas (broad sense heritability) dan heritabilitas dalam arti sempit (narrow sense heritability). Heritabilitas arti luas (h2bs) adalah rasio dari ragam total genetik terhadap ragam
fenotipiknya, sedangkan heritabilitas dalam arti sempit (h2ns) adalah rasio ragam genetik
aditif terhadap ragam fenotipe. Heritabilitas dalam arti sempit banyak digunakan karena ragam genetik aditif dipindahkan dari tetua kepada keturunannya (Mangoendidjojo 2003). Heritabilitas suatu faktor perlu diketahui dalam menentukan kemajuan seleksi apakah karakter yang tampil melalui fenotipenya banyak dipengaruhi oleh faktor genetik atau lingkungan (Poehlman 1995). Nilai heritabilitas dapat dinyatakan dalam persen atau desimal. Nilai tertinggi 100% atau 1.0.
Nilai ini menunjukkan bahwa semua variasi disebabkan oleh faktor genetik. Tetapi bila nilainya 0.0 maka tidak ada variasi dalam populasi yang disebabkan oleh faktor genetik. Nilai heritabilitas suatu karakter tidak konstan. Banyak faktor yang mempengaruhi heritabilitas, antara lain karakteristik populasi, sampel genotipe yang dievaluasi, metode pendugaan, adanya pertautan gen (linkage), pelaksanaan percobaan, generasi populasi yang diuji dan lain-lain.
Fenotipe merupakan hasil interaksi antara faktor genetik dengan lingkungan. Dengan demikian kita harus dapat membedakan apakah keragaman yang diamati dari suatu karakter disebabkan oleh faktor genetik atau lingkungannya. Suatu karakter yang dikendalikan oleh sedikit gen (simple genic) disebut karakter kualitatif, dan yang dikendalikan oleh banyak gen (polygenic) disebut karakter kuantitatif. Karakter kualitatif sedikit dipengaruhi oleh lingkungan. Karakter kuantitatif banyak dipengaruhi oleh lingkungan (Mangoendidjojo 2003).
Seleksi pada karakter yang mempunyai nilai heritabilitas rendah dilakukan pada generasi lanjut, sedangkan seleksi pada karakter yang mempunyai nilai heritabilitas tinggi dapat dilakukan pada generasi awal (F2) dan didasarkan pada jenis tanamannya. Kemajuan yang lebih besar dan cepat akan diperoleh apabila seleksi dilakukan pada karakter yang dikendalikan oleh gen aditif (George 1999).
Variabilitas Genetik
Allard (1960) mengemukakan bahwa ragam fenotipik tersusun atas ragam genetik dan ragam lingkungan, sehingga perbedaan fenotipik suatu tanaman tergantung pada genotipe dan lingkungannya serta interaksi keduanya.
Identifikasi dan seleksi awal dari beberapa genotipe melibatkan populasi yang sangat besar. Nilai pemuliaan dari masing-masing individu dipengaruhi oleh beberapa faktor seperti lingkungan maupun kompetisi, sehingga suatu metode analisis genetik kuantitatif khusus sangat diperlukan untuk menghilangkan faktor-faktor yang mengganggu tersebut (Falconer 1976).
Ragam genetik dan ragam lingkungan biasanya dinotasikan dengan σ2G dan σ2E.
Kontribusi ragam genetik dan ragam lingkungan terhadap ragam fenotipik adalah bebas atau tidak saling mempengaruhi. Ragam genetik dan ragam lingkungan yang menyusun ragam fenotipe dapat dipisahkan dengan beberapa metode di antaranya adalah metode lingkungan yang seragam. Dalam metode ini lingkungan dibuat seragam sehingga ragam lingkungan menjadi nol dan ragam genetik sama dengan ragam fenotipe (Falconer 1976).
Setelah dilaporkan adanya faktor mewaris pengendalian sifat oleh Mendel, orang-orang beranggapan bahwa pertumbuhan tanaman semata-mata diatur oleh gen-gen dalam kromosom, sedangkan lingkungan hanya meningkatkan potensi sifatnya. Namun setelah diketahui bahwa tanaman-tanaman tidak berkembang secara teratur menurut perubahan lingkungan, maka mulai disadari adanya interaksi antar genotipe dan lingkungan. Untuk mengetahui seberapa jauh peranan lingkungan pada suatu sifat tanaman, maka didekati dengan usaha untuk memisahkan pengaruh genotipe dan lingkungan serta interaksinya (Poespodarsono 1988).
Heterosis
Hibrida adalah turunan petama (F1) dari persilangan antara dua atau lebih galur
murni. Persilangan antara dua galur murni dapat memberikan hasil yang lebih tinggi dari nilai tengah kedua tetuanya atau bahkan memberikan hasil tertinggi dari nilai salah satu tetua yang paling baik. Keadaan ini dikenal sebagai efek heterosis (Poehlman 1959).
Menurut Welsh dan Mogea (1991), heterosis adalah peningkatan dalam ukuran atau vigor dari suatu hibrida melebihi rata-rata kedua tetuanya. Heterosis berkaitan dengan ekspresi heterozigositas. Adanya akumulasi alel dominan yang baik pada F1 dan sebagian
alel tersebut berasal dari tetuanya, maka pengaruh jelek dari alel homozigot resesif akan tertutupi.
Heterosis dapat dibagi menjadi tiga tipe tergantung genotipe pembanding yang digunakan (Nuruzzaman et al. 2002 dan Virmani et al. 2003). Ketiga tipe heterosis tersebut adalah: (1) Mid-parent heterosis (heterosis) yaitu peningkatan atau penurunan performa hibrida dibandingkan dengan nilai rata-rata kedua tetua; (2) High parent heterosis (heterobeltiosis) yaitu peningkatan atau penurunan performa hibrida dibandingkan dengan nilai tetua terbaik yang digunakan dalam kombinasi persilangan; dan (3) Standard heterosis yaitu peningkatan atau penurunan performa hibrida dibandingkan dengan varietas cek (varietas pembanding).
Menurut Allard (1960), dasar genetik penyebab terjadinya heterosis belum dapat diungkapkan secara jelas. Teori pertama adalah teori epistasis, yaitu interaksi antara alel yang berbeda lokus memberi nilai lebih karena hasil penambahan dan perkalian dari gen dominan pendukung keunggulan sifat.
Teori kedua yaitu hipotesis dominan. Menurut Poehlman (1979), hipotesis ini berdasarkan teori bahwa gen yang menguntungkan untuk tanaman bersifat dominan dan gen yang merugikan bersifat resesif. Gen dominan yang berasal dari satu tetua akan dilengkapi oleh gen dominan dari tetua lainnya sehingga tanaman F1 memiliki kombinasi gen dominan yang menguntungkan dari kedua tetuanya.
Hipotesis over dominan menyatakan bahwa heterozigot (a1a2) lebih vigor dan
produktif dibandingkan homozigot (a1a1 atau a2a2). Alel a1 dan a2 memiliki fungsi yang
berbeda dan penggabungannya a1 dan a2 lebih superior jika dibandingkan homozigotnya
(a1a1 atau a1a1). Semakin berbeda fungsi alel penyusun heterozigot, semakin tinggi efisien
pembentukan superioritasnya (a1a2 < a1a3 atau a1a4) (Allard 1960).
Efek heterosis atau yang dikenal dengan hibrid vigor dapat terekspresi di berbagai bagian tanaman. Pada umumnya pemulia tanaman melihat efek heterosis pada peningkatan pertumbuhan vegetatif atau hasil; seperti ukuran sel, tinggi tanaman, ukuran daun, perkembangan akar, ukuran tongkol, jumlah benih, ukuran biji, dan lainnya (Poehlman 1979).
Keunggulan sifat kuantitatif berkembang dengan peran banyak gen pendukungnya. Masing-masing sifat pendukung ini dapat menunjukkan keunggulannya karena heterosis dari gen pengendalinya. Dengan demikian akan memberi pengaruh pada pertumbuhan
yang lebih baik dan akhirnya akan menunjukkan nilai lebih pada sifat kuantitatif tersebut, misalnya produksi. Hal ini menjelaskan bahwa peran gen masing-masing sifat tidak hanya dari segi morfologisnya tetapi juga kemampuan fisiologisnya. Misalnya biji yang lebih besar, makin cepat berkecambah, makin besar jumlah anakan, makin tahan terhadap lingkungan ekstrim akan memberi kemungkinan berproduksi lebih baik (Sutjahjo et al. 2006).
Penelitian PendugaanParameter Genetik Vigor Benih
Informasi pendugaan parameter genetik sangat diperlukan untuk memperbaiki karakter suatu populasi. Parameter genetik dapat mendukung keberhasilan program pemuliaan tanaman, karena itu sangat penting dilakukan (Baihaki 2009).
Pendugaan parameter genetik yang terkait dengan karakter vigor daya simpan benih cabai belum banyak informasinya. Pendugaan parameter genetik dilakukan untuk menduga nilai heritabilitas arti luas, ragam genetik, ragam fenotipee, koefisien keragaman genetiknya (KKG), koefisisen keragaman fenotipee (KKP) dan heterosis. Pada program pemuliaan tanaman, keragaman genetik pada populasi sangat penting sebagai dasar seleksi. Parameter genetik yang tinggi dapat mendukung keberhasilan program pemuliaan pada karakter yang akan diperbaiki.
Hasil penelitian tentang karakter yang terkait dengan vigor benih mengenai vigor lebih banyak mengenai vigor kekuatan tumbuh benih. Karakter vigor kekuatan tumbuh benih dikendalikan secara genetik dan pendugaan parameter genetiknya telah diteliti pada benih kedelai (Thseng et al. 1995 ; Basra et al. 2000)); benih chickpea (Abbo et al. 2000); benih sorgum (Cisse & Ejeta 2003): dan benih padi (Ali et al. 2006); Okelola et al. 2007; Akram et al. 2007).
Penelitian Priestley (1986) pada benih jagung menghasilkan karakter yang terkait dengan vigor daya simpan benih jagung yang dikendalikan secara genetik adalah karakter ukuran benih. Ukuran benih biasanya mempengaruhi masa hidup benih, karena karakter tersebut dikaitkan dengan kandungan cadangan makanan dan ukuran embrio. Hal ini didukung oleh hasil penelitian Arief et al. (2004), bahwa ukuran benih jagung berpengaruh terhadap daya simpan benih jagung dan vigor daya simpan benih. Penemuan terbaru menemukan gen untuk karakter ukuran benih adalah gen yang mengendalikan ukuran benih dan bimassa hasil oleh Chojecki pada tahun 2008.
Thseng et al. (1995) yang meneliti benih kedelai menghasilkan karakter yang terkait dengan vigor kekuatan tumbuh benih yang dikendalikan secara genetik dan mempengaruhi produktivitas tanaman kedelai pada suhu tinggi yaitu karakter daya berkecambah dan daya simpan benih. Keragaman genetik untuk kedua karakter tersebut sangat tinggi. Thseng meneliti menggunakan 12 genotipe kedelai yang ditaman di Taichung National Chung Hsing University,19 varietas ditanam di Hualien District Agricultural improvement Station dan 46 varietas ditamnam di Seed Improvement and Propagation Station Experimental Farm Taichung.
Abbo et al. (2000) meneliti pada benih chickpea menghasilkan karakter yang terkait dengan vigor kekuatan tumbuh benih yang dikendalikan oleh faktor genetik adalah berat benih, ukuran benih dan konsentrasi kalsium pada benih. Abbo menggunakan 16 genotipe benih chickpea berasal dari 6 negara yaitu Mexico, Israel, Bulgaria, Ethiopia, India, Turkey. Analisis kalsium benih dilakukan pada generasi persilangan F1 dan selfing F1 menghasilkan F2, selfing F2 menghasilkan F3 dan selfing F3 menghasilkan F4, hal ini disebabkan kandungan kalsium dipengaruhi oleh efek maternal pada jaringan seed coat benih.
Penelitian Cisse & Ejeta (2003) pada benih sorgum menghasilkan karakter yang terkait dengan vigor kekuatan tumbuh yang dipengaruhi faktor genetik adalah daya berkecambah dan tinggi kecambah, berat kering kecambah dan berat 100 benih. Keempat karakter tersebut mempunyai nilai yang tinggi untukheritabilitas, dan koefisien keragaman genetik (KKG) nya. Hubungan antara karakter tersebut sangat nyata untuk beberapa genotipe sorgum misalnya genotipe tetua sorgum kaoliang. Genotipe tersebut dapat dipergunakan secara efektif untuk meningkatkan perkecambahan pada kondisi suhu dingin dan pada suhu optimum.
Penelitian Akram et al. (2004) mengenai karakter yang terkait dengan vigor kekuatan tumbuh benih padi. Karakter yang terkait dengan vigor benih padi adalah tolok ukur daya berkecambah, aktivitas α amylase, aktivitas dehidrogenase, panjang akar (PA), bobot kering kecambah normal (BKKN) pada hari ke-14, dan bobot kering benih. Hasilnya menyatakan bahwa 7 (tujuh) genotipe benih padi F1 yang digunakan untuk pengujian vigor di laboratorium dan percobaan di lapang mengindikasikan adanya aksi gen aditif pada hampir semua karakter. Ali et al. (2006) meneliti pada benih padi menghasilkan karakter yang terkait dengan toleransi tanaman padi pada suhu dingin. Karakter yang terkait dengan
toleransi terhadap suhu dingin yang dikendalikan secara genetik yaitu kadar air benih, viabilitas awal, dan daya berkecambah. Ali menggunakan 13 (tiga belas) genotipe padi yang dipanen di Bangladesh Rice Research Institute, 11 (sebelas) genotipe dipanen di Ganzipur dan di lahan pertanian di Barisal.
Okelola et al. (2007) meneliti tentang karakter yang terkait dengan vigor kekuatan tumbuh benih padi yang dikendalikan secara genetik. Karakter tersebut adalah daya berkecambah, kecepatan tumbuh, indeks vigor, energi perkecambahan dan produksi benih. Ketiga karakter tersebut mempunyai nilai koefisien keragaman genetik (KKG), heritabilitas dan kemajuan genetik tinggi. Karakter tersebut kemungkinan dapat menjadi karakter seleksi untuk mengembangkan genotipe padi pada tiap musimnya. Karakter seleksi tersebut dapat digunakan sebagai perbaikan umum padi Afrika Barat.
Chojecki (2008) melaporkan hasil penelitiannya adalah pada benih arabidopsis dihasilkan faktor yang mempengaruhi ukuran benih ditemukan yaitu gen DA1, dengan menggunakan mapping genetik dan studi kloning gen. Gen DA1 menginduksi phytohormon absisic acid (ABA) dan mutannya da1-1 insensitive terhadap ABA dan overexpressi dari da1-1 dapat meningkatkan benih dan organ benih. Hal ini merupakan penemuan baru dalam transgenik tanaman.