ISOLASI DAN PEMURNIAN PROTEIN INHIBITOR RNA
HELIKASE VIRUS HEPATITIS C DARI
KAPANG ENDOFIT CgKTm 5 F
ERVIAN HADI RAMDANI
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2011
ISOLASI DAN PEMURNIAN PROTEIN INHIBITOR RNA
HELIKASE VIRUS HEPATITIS C DARI
KAPANG ENDOFIT CgKTm 5 F
ERVIAN HADI RAMDANI
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada
Departemen Biokimia
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2011
Judul : Isolasi dan Pemurnian Protein Inhibitor RNA Helikase Virus Hepatitis C
dari Kapang Endofit CgKTm 5 F
Nama : Ervian Hadi Ramdani
NIM : G84062350
Disetujui
Komisi Pembimbing
Ir.Eman Kustaman
A.Zaenal Mustopa, M.Si
Ketua
Anggota
Diketahui
Dr. I Made Artika, M.App.Sc
Ketua Departemen Biokimia
ABSTRAK
ERVIAN HADI RAMDANI. Isolasi dan Pemurnian Protein Inhibitor RNA
Helikase Virus Hepatitis C dari Kapang Endofit CgKTm 5 F. Dibimbing oleh
EMAN KUSTAMAN dan A. ZAENAL MUSTOPA.
Hepatitis C merupakan penyakit yang disebabkan oleh infeksi virus hepatitis C
(HCV), dan menjadi masalah kesehatan yang belum teratasi karena pengobatan
yang dilakukan memiliki efiseinsi yang rendah dan mahal. Pengobatan penyakit
hepatitis C berdasarkan molekul target banyak dikembangkan akhir-akhir ini,
diantaranya mengarah pada target RNA helikase virus hepatitis C. RNA helikase
berfungsi dalam replikasi HCV, sehingga penghambatannya dapat memutus siklus
hidup virus ini. Penghambatan terhadap RNA helikase dapat dilakukan oleh
protein dari kapang endofit CgKTm 5 F. Penelitian ini bertujuan melakukan
isolasi dan pemurnian protein inhibitor dari kapang endofit CgKTm 5 F yang
mempunyai aktivitas inhibisi terhadap RNA helikase HCV. Isolasi dilakukan
dengan pengendapan amonium sulfat 90% dan pemurnian dilakukan dengan
kromatografi gel filtrasi Sephadex G-50 dengan pelarut metanol 40%. Uji ATPase
digunakan untuk menghitung aktivitas inhibisi RNA helikase HCV. Fraksi ke- 8
hasil kromatografi gel filtrasi memiliki aktivitas tertinggi dalam menghambat
RNA helikase HCV. Analisis SDS-PAGE terhadap fraksi tersebut menunjukkan
bahwa terdapat empat pita protein dan semuanya memiliki bobot molekul > 17
kDa. Aktivitas spesifik protein inhibitor pada fraksi ke- 8 sebesar 5.92x10
-4U/mg
dengan kemurnian 5.38.
ABSTRACT
ERVIAN HADI RAMDANI. Isolation and Purification Protein Inhibitor of RNA
Helicase Hepatitis C Virus from Endophytic Fungi CgKTm 5 F. Under the
direction of EMAN KUSTAMAN and A. ZAENAL MUSTOPA.
Hepatitis C is a disease caused by hepatitis C virus (HCV) infection. It is still
unresolved health problem because of low efficiency and high cost of current
therapy. Treatment of hepatitis C based on the target molecule has been
developed recently, and RNA hepatitis C virus helicase targeting is among of
them. RNA helicase is highly involved in HCV replication, any interruption to
RNA helicase could disrupt HCV life cycle. It has been reported that endophytic
fungi protein, namely CgKTm 5 F, has ability to inhibit RNA helicase. Thus, the
aim of the study is to isolate and purify CgKTm 5 F. Isolation was carried
out with 90% ammonium sulphate precipitation and purification was done using
Sephadex
G-50
gel
filtration
chromatography
with
40%
ethanol
solution. ATPase assay was used to calculate the inhibitory activity of HCV
RNA helicase. The results showed that the 8
thfraction of gel filtration
chromatography exhibits the highest activity in inhibiting HCV RNA
helicase. SDS-PAGE analysis indicated that
there
were four bands of
protein and all of them had a molecular weight of > 17 kDa. Moreover the specific
activity was 5.92x10
-4U / mg with fraction purity of 5.38.
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT, yang telah melimpahkan
karunia-Nya sehingga usulan penelitian ini dapat terselesaikan dengan
sebaik-baiknya. Penelitian ini berjudul Isolasi dan Pemurnian Protein Inhibitor RNA
Helikase Virus Hepatitis C Dari Kapang Endofit CgKTm 5 F. Penelitian ini
dilakukan di Laboratorium Bakteriologi dan Virologi molekuler, Puslit
Bioteknologi LIPI Cibinong.
Penulis menyampaikan terima kasih kepada Bapak Eman Kustaman selaku
ketua pembimbing, Bapak A. Zaenal Mustopa selaku anggota pembimbing, Ibu
Laksmi Ambarsari, Bapak I Made Artika, dan Ibu Titik K. Prana yang telah
memberi izin penggunaan isolat kapang endofit CgKTm 5 F. Ucapan terima kasih
juga penulis sampaikan kepada bapak, mamah, adikku Anis, Mbah, Umi, Pak
Idun, Mak Idun, Prabawati Hyunita Putri dan semua keluarga yang telah
memberikan bantuan dan doa selama ini. Tidak lupa penulis mengucapkan terima
kasih kepada staf laboratorium (Mas Ridwan, Mas Erik, dan Mas Kukun),
teman-teman di laboratorium (Mas Melki, Soleh, Skotia, Tya, Ika, dan Marcel),
Biokimia 43 (Adju, Tiwi, Nestri, Taufiq, Herdit, Mike, Danang, Umul, Haya, dan
Himajaman), dan D’Sabars (Diki, Dedi, Soleh, Imam, dan Rido) atas bantuannya
selama ini.
Akhir kata, penulis mengharapkan kritik dan saran yang membangun atas
ketidaksempurnaan dari skripsi ini sehingga tulisan ini dapat berguna bagi penulis
sendiri maupun semua pihak yang membutuhkan demi kemajuan ilmu
pengetahuan.
Bogor, Februari 2011
RIWAYAT HIDUP
Ervian Hadi Ramdani dilahirkan di Sukabumi pada tanggal 11 April 1989 dari
ayah Karna Sukarna dan ibu E. Nurhasanah. Penulis merupakan anak pertama dari
dua bersaudara.
Penulis menyelesaikan sekolah menengah atas di SMA Negeri I Cibadak
Kabupaten Sukabumi pada tahun 2006. Pada tahun yang sama, penulis diterima di
Institut Pertanian Bogor melalui jalur Undangan Seleksi Masuk IPB (USMI).
Tahun 2007 penulis diterima sebagai mahasiswa Departemen Biokimia, Fakultas
Matematika dan Ilmu Pengetahuan Alam.
Selama mengikuti perkuliahan, penulis pernah menjadi asisten praktikum mata
kuliah Biokimia Umum dan mata kuliah Struktur dan Fungsi Biomolekul pada
tahun yang 2009/2010. Penulis melakukan praktek kerja lapang di Laboratorium
Biologi Sel dan Jaringan Hewan, Pusat Penelitian Bioteknologi LIPI dari bulan
Juli sampai Agustus 2009 dengan judul laporan praktek lapang, yaitu Pengaruh
Penambahan Fetal Bovine Serum (FBS) Pada Media Dulbecco’s Modified of
Eagle Medium (DMEM) Terhadap Pertumbuhan Sel Cumulus Mencit (Mus
musculus). Pada tahun 2008/2009 dan 2009/2010 penulis aktif di Himpunan
Profesi (Himpro) CREBs (Community Research and Education Biochemistry
DAFTAR ISI
Halaman
DAFTAR GAMBAR ... ii
DAFTAR TABEL ... ii
DAFTAR LAMPIRAN ... ii
PENDAHULUAN ... 1
TINJAUAN PUSTAKA
Kapang Endofit ... 2
Virus Hepatitis C ... 2
RNA Helikase ... 3
SDS-PAGE (Sodium Dedosil Sulfat Poliakrilamid Gel Elektroforesis) ... 3
Kromatografi Gel Filtrasi ... 4
BAHAN DAN METODE PERCOBAAN
Bahan dan Alat ... 4
Metode ... 5
HASIL DAN PEMBAHASAN
Hasil Isolasi RNA Helikase HCV ... 7
Hasil Pemurnian RNA Helikase ... 7
Waktu Pemanenan Protein Inhibitor dari CgKTM 5 F ... 8
Isolat Protein Inhibitor dari Kapang Endofit CgKTm 5 F ... 8
Hasil Pemurnian Protein Inhibitor dari Kapang Endofit CgKTm 5 F ... 9
Bobot Molekul Protein Inhibitor RNA helikase HCV ... 10
Konsentrasi Protein Inhibitor RNA Helikase HCV ... 10
SIMPULAN DAN SARAN
Simpulan ... 11
Saran ... 11
DAFTAR PUSTAKA ... 11
DAFTAR GAMBAR
Halaman
1 Kapang endofit CgKTm 5 F ... 2
2 Struktur virus hepaitis C ... 3
3 Mekanisme RNA heilkase ... 3
4 Kromatografi gel filtrasi ... 4
5 Elektroforegram SDS-PAGE RNA helikase hepatitis C. ... 8
6 Aktivitas inhibisi RNA helikase HCV terhadap waktu inkubasi ... 8
7 Aktivitas inhibisi RNA helikase RNA helikase HCV pada beberapa
tingkatan pengendapan amonium sulfat ... 9
8 Aktivitas inhibisi RNA helikase HCV fraski hasil kromatografi gel filtasi .. 10
9 Elektroforegram SDS-PAGE protein CgKTm 5 F ... 10
DAFTAR TABEL
Halaman
1 Variasi konsentrasi gel berdasarkan bobot protein ... 4
2 Ukuran protein minimum yang dapat dipisahkan oleh matriks gel ... 4
3 Kadar dan aktivitas spesifik protein inhibitor RNA helikase HCV dari
kapang CgKTm 5 F ... 11
DAFTAR LAMPIRAN
Halaman
1 Diagram alir penelitian ... 15
2 . Bobot molekul relatif enzim RNA helikase HCV ... 17
3 Bobot molekul relatif protein kapang CgKTm 5 F ... 18
4 Kurva standar Bovine Serum Albumin (BSA) ... 19
5 Kurva standard fosfat (Uji ATPase) ... 20
6 Nilai inihibisi RNA helikase HCV optimasi pemanenan kultur kapang
pengendapan amonium sulfat ... 21
7 Nilai inihibisi RNA helikase HCV dari optimasi pengendapan amonium
sulfat ... 22
8 Nilai inhibisi RNA helikase HCV hasil kromatografi kolom Shepadex-50 . 23
9 Perhitungan aktivitas protein inhibitor ... 25
PENDAHULUAN
Hepatitis C merupakan penyakit yang disebabkan oleh virus hepatitis C (Hepatitis C Virus atau HCV). Virus hepatitis C memiliki satu untai positif RNA, berpelindung, dan berbentuk bola dengan diameter 50-60 nm (Borowski et al. 2008). Virus ini memiliki tingkat virulensi yang tinggi. Sebanyak 180 juta jiwa penderita di seluruh dunia mengidap penyakit ini dan bertambah setiap tahunnya. Penularan penyakit ini umumnya terjadi melalui transfusi darah atau jarum suntik yang tercemar darah yang mengandung HCV (Kandil et al. 2009).
Saat ini pengobatan yang diberikan kepada penderita penyakit hepatitis C yaitu terapi interferon-alfa, ribavirin, dan kombinasi keduanya. Namun, pengobatan ini memiliki beberapa kelemahan, yaitu mahal dan memiliki efek samping. Efek samping yang dapat terjadi akibat penggunaan ribavirin yaitu dapat menyebabkan anemia yang dapat terjadi secara tiba-tiba dan kecacatan pada janin. Terapi kombinasi interferon alfa dengan ribavirin belum dapat mengobati dengan efek yang luas, karena hanya dapat menghilangkan virus sampai 50% dari HCV genotipe 1 dan 80% dari genotipe 2 dan 3 (Yayasan Spiritia 2005 dalam Megawati 2008).
Terapi untuk hepatitis C diharapkan memiliki efektifitas yang tinggi, dapat digunakan secara oral, memiliki efek samping yang sedikit, murah, dan cocok untuk kebanyakan pasien (McHutchison & Patel 2002). Salah satu pendekatan pengobatan yang sedang berkembang adalah terapi target molekuler yang bertujuan menghambat siklus hidup HCV. Salah satunya yaitu dengan cara menghambat kerja RNA helikase .
Enzim RNA heilkase HCV memiliki peranan penting dalam tahapan replikasi virus ini yaitu membuka ikatan dupleks RNA virus agar dapat ditranslasikan (Borowski et al. 2008). RNA helikase memiliki tiga macam aktivitas, yaitu aktivitas pengikatan RNA, pengikatan adenosine triphosphate (ATP), dan pembukaan rantai RNA (unwinding). Apabila proses pembukaan ikatan dupleks RNA virus sebagai pustaka genetik tidak dapat dilakukan, maka proses translasi informasi genetik tidak dapat berjalan sehingga siklus hidup HCV terhenti. Oleh karena itu, enzim ini dapat dijadikan target obat yang potensial untuk pengembangan dan penemuan obat anti HCV yang baru (Utama et al. 2000).
Kinerja dari RNA helikase dapat dihambat oleh suatu inhibitor. Inhibitor enzim dapat diperoleh dari hasil metabolit sekunder, misalnya metabolit dari kapang endofit. Kapang endofit adalah kapang yang hidup berkoloni di dalam jaringan tanaman dan tidak menyebabkan efek yang negatif bagi tanaman inang (Maksum 2005). Endofit ini merupakan sumber dari metabolit sekunder yang belum diketahui potensinya di bidang medis, pertanian, dan pemanfaatan industri. Oleh karena itu, pemanfaatan kapang endofit perlu untuk dilakukan.
Pemanfaatan kapang endofit sebagai sumber obat baru saat ini sedang banyak
dilakukan. Hal ini dikarenakan
mikroorganisme ini mampu menghasilkan senyawa bioaktif yang sama dengan tanaman inangnya. Satu tanaman dapat memiliki satu atau lebih mikroorganisme endofit yang hidup di dalamnya, sementara di dunia terdapat
30000 jenis tanaman, maka dapat
dibayangkan kekayaan biodiversitas mikroorganisme ini sangatlah besar (Strobel & Daisy 2003). Biodiversitas yang besar inilah yang dapat digunakan pemanfaatannya dalam mencari inhibitor RNA helikase HCV.
Megawati (2008) menyebutkan bahwa protein metabolit sekunder dari isolat endofit CgKTm 5 F yang diisolasi dari tanaman temu
putih gombyok memiliki kemampuan
menghambat RNA helikase HCV, sehingga dapat dimanfaatkan sebagai obat bagi pasien yang terkena penyakit hepatitis C. Namun, perlu dilakukan isolasi dan pemurnian terhadap protein inhibitor ini sehingga memiliki efektifitas yang maksimal dalam menghambat kinerja RNA helikase.
Penelitian ini bertujuan melakukan isolasi dan memurnikan protein inhibitor RNA helikase virus hepatitis C dari isolat CgKTm 5 F. Belum ditemukannya obat atau vaksin yang dapat mengobati infeksi virus hepatitis C secara efektif sampai saat ini memberikan inspirasi untuk mencoba mencari senyawa antivirus yang berasal dari bahan yang berasal dari alam bukan sintetik.
Hipotesis penelitian ini adalah protein metabolit sekunder yang dihasilkan oleh
kapang endofit CgKTm 5 F dapat
menghambat aktivitas RNA helikase virus hepatitiis, sehingga dapat digunakan sebagai kandidat obat hepatitis C. Penelitian ini diharapkan dapat memberikan informasi mengenai protein metabolit sekunder yang dihasilkan oleh kapang endofit CgKTm 5 F sebagai inhibitor RNA helikase virus hepatisis C.
TINJAUAN PUSTAKA
Kapang EndofitEndofit adalah mikroorganisme yang hidup di dalam tanaman tingkat tinggi. Bacon dan White (2000) menyebutkan bahwa endofit merupakan mikroorganisme yang hidup berkoloni di dalam jaringan tumbuhan tanpa menyebabkan efek negatif terhadap tumbuhan tersebut. Mikroorganisme yang hidup pada jaringan tumbuhan ini memilki hubungan simbiosis. Mikroorganisme yang banyak ditemukan hidup sebagai endofit adalah bakteri dan kapang.
Endofit diperkirakan menghasilkan metabolit yang karakteristiknya mirip dengan tumbuhan inangnya. Schuzt et al. (2002) menjelaskan bahwa metabolit yang dihasilkan dipengaruhi oleh lingkungan dan tingkatan organisme tertentu. Pemilihan tumbuhan yang
akan diisolasi endofitnya harus
mempertimbangkan beberapa hal. Pertama, tumbuhan tersebut berasal dari lingkungan yang unik dan mempunyai kemampuan untuk bertahan hidup. Kedua, tumbuhan tersebut memiliki sejarah etnobiologi yang digunakan sebagai obat dari penyakit tertentu. Ketiga, ketersediaan tumbuhan bersifat endemik karena dikaitkan dengan lingkungan tempat tumbuhnya sehingga diduga menghasilkan metabolit aktif untuk mempertahankan kelangsungan hidup. Keempat, tumbuhan yang hidup di dalam lingkungan dengan keragaman tinggi sehingga kemungkinan memiliki endofit dengan keragaman yang tinggi pula (Strobel & Daisy 2003).
Mikroorganisme yang hidup sebagai endofit salah satunya adalah kapang. Isolat CgKTm 5 F merupakan kapang endofit yang diisolasi dari tanaman temu putih. Endofit ini termasuk ke dalam kelompok kapang dengan morfologi bulat (Gambar 1). Temu putih (Curcuma zedoaria) dapat dijadikan sebagai obat antivirus (Chungsamarnyart et al. 2007).
Gambar 1 Kapang endofit CgKTm 5 F.
Suhu optimal yang dibutuhkan untuk pertumbuhan kapang berkiksar 25-30oC. Pada umumnya kapang dapat tumbuh pada kisaran pH 2-8.5, akan tetapi pertumbuhannya akan lebih baik pada kondisi pH rendah. Nilai pH optimum untuk pertumbuhan kapang berkisar antara 6-7 (Fardiaz 1992).
Virus Hepatitis C
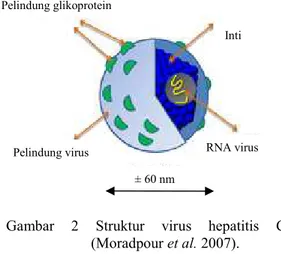
Virus hepatitis C (HCV) ditemukan pertama kali pada tahun 1989 dengan nama virus hepatitis non-A dan non-B. Virus ini termasuk kedalam genus Hepacivirus dan famili Flaviviridae. Genom HCV terdiri atas utas tunggal RNA sense positif yang berukuran sekitar 9.6 kilobasa (kb). Virus ini berbentuk bulat, berpelindung, dan berdiameter sekitar 50-60 nm (Gambar 2) (Tellinghuisen 2007).
Genom HCV terdiri dari open reading frame (ORF) tunggal yang mengkodekan poliprotein tunggal. Poliprotein tersebut merupakan prekusor bagi 3000 jenis asam amino. Poliprotein ini akan diubah menjadi sekitar 10 jenis protein yang berbeda dan terbagi dalam dua kelompok besar protein virus, yaitu protein struktural (protein inti, E1, E2, dan p7) dan protein nonstruktural (NS) (NS2, NS3, NS4A, NS4B, NS5A, dan NS5B) (Lauer &Walker 2001).
Protein struktural dari HCV terletak pada daerah N terminal. Protein inti diperlukan untuk perakitan partikel virus baru (virion) dalam bentuk nukleokapsid. Protein inti berperan sebagai modulator metabolisme lipid dan hepatokarsinogenesis. Protein pelindung envelope 1 (E1) mempunyai bobot molekul sekitar 30-35 kDa, sedangkan protein pelindung envelope 2 (E2) sekitar 70-75 kDa. Pelindung tersebut banyak mengandung glikoprotein. Kedua pelindung ini mempunyai tingkat mutasi yang sangat tinggi dan bersifat sangat spesifik terhadap antibodi. Pada E2 terdapat region yang berfungsi unuk mengikat pada cluster of diffrerentiation 81 (CD81), yaitu reseptor untuk HCV. Protein p7 berperan dalam pengaturan kanal ion (Tellinghuisen 2007).
Protein nonstruktural (NS) terdiri dari NS2, NS3, NS4A, NS4B, NS5A, dan NS5B. Protein nonstruktural berperan dalam replikasi virus. Protein NS2 mempunyai aktivitas protease. Protein NS3 mempunyai dua aktivitas utama, yaitu serin protease dan NTPase atau helikase. Protein NS4A berperan sebagai kofaktor serin protease NS3, sedangkan NS4B belum diketahui fungsinya secara jelas. NS5A merupakan fosfoprotein Kapang
endofit CgKTm 5 F
yang fungsinya belum diketahui secara jelas. Protein ini bersifat hidrofilik dan sangat sensitif terhadap interferon. NS5B mempunyai peranan dalam aktivitas
RNA-dependent RNA polimerase (RdRp)
(Tellinghuisen 2007).
Virus hepatitis C menyerang sel hati atau limfosit B. Virus ini menyebabkan penyakit hepatitis C yang dalam jangka panjang mengakibatkan peradangan hati, sirosis, dan kanker hati. Penyakit ini sulit dideteksi karena gejala yang ditimbulkan mirip dengan penyakit yang lain, seperti mual, nafsu makan berkurang, mudah lelah, timbul kekuningan (mata, kulit), dan urin berwarna gelap. Umumnya penyakit ini terdeteksi apabila sudah mencapai tingkat akut, sekitar 30-80% infeksi (Sy & Jamal 2006).
Gambar 2 Struktur virus hepatitis C (Moradpour et al. 2007).
RNA Helikase
Helikase adalah enzim yang berperan dalam membuka untai ganda nukleotida (DNA atau RNA) menjadi untai tunggal. RNA helikase merupakan enzim yang membuka ikatan dupleks RNA sense positif dengan antisense negatifnya menjadi untai tunggal. Enzim ini pertama kali ditemukan pada Escherichia coli. Enzim ini bekerja secara katalitik dengan memutus ikatan hidrogen yang terjadi antara kedua untai tersebut (Kadare & Haenni 1997).
RNA helikase yang terdapat pada virus hepatitis C (HCV) dikodekan oleh protein NS3 RNA helikase (Ceng et al. 2007). Enzim ini juga memiliki aktivitas ATPase dan pengikatan terhadap untai RNA. Mekanisme kerja dari RNA helikase pertama-tama adalah mengikat untai RNA pada ujung 3’. ATP akan terikat pada sisi aktif enzim tersebut dan dihidrolisis oleh RNA helikase menjadi ADP dan fosfat anorganik. Energi yang dilepaskan digunakan oleh RNA helikase untuk
membuka ikatan hidrogen pada dupleks RNA. Enzim akan bergerak sepanjang arah 3’-5’ dalam memisahkan kedua untai RNA dan berperan dalam proses translasi, pembentukan poliprotein, dan memutus interaksi RNA dengan protein (Gambar 3) (Utama et al. 2000).
RNA helikase berperan penting dalam replikasi virus dapat dijadikan target dalam pencarian obat HCV. Target pencarian obat dapat dilakukan dengan menghambat salah satu aktivitas dari RNA helikase tersebut. Penghambatan dilakukan dengan mencari inhibitor RNA helikase sehingga proses replikasi virus menjadi terhambat (Megawati 2008). Aktivitas ATPase dari RNA helikase lebih mudah digunakan sebagai target pencarian obat antivirus. Hal tersebut dikarenakan substrat yang digunakan, yaitu ATP, bersifat lebih stabil (Tellinghuisen 2007).
Gambar 3 Mekanisme RNA helikase (Utama et al. 2000).
Sodium Dodecylsulphate Polyacrylamide Gel Electrophoresis (SDS PAGE)
Elekroforesis adalah teknik pemisahan yang memisahkan analit berdasarkan kemampuannya bergerak dalam medium konduksi yang biasanya berupa larutan bufer dan akan memberikan respons setelah ditambahkan medan listrik. Suatu zat yang bermuatan jika diberi potensial, maka zat tersebut akan berpindah sepanjang medium yang kontinu ke arah katode atau anode sesuai dengan muatan yang dibawanya (Harvey 2000).
Elektroforesis SDS-PAGE termasuk ke dalam kelompok elektroforesis zona/wilayah, yaitu kelompok elektroforesis yang dibedakan
berdasarkan medium penyangganya.
Elektroforesis SDS-PAGE menggunakan gel
± 60 nm RNA virus Pelindung virus Inti Pelindung glikoprotein RNA helikase 3’ 5’ ADP Pi 5’ 3’ Pembukaan ikatan dupleks RNA Pengikatan RNA 5’ 3’ 3’ 5’ RNA RNA helikase Hidrolisis ATP ATP 5’ 3’ 3’ 5’ RNA helikase
buatan sebagai medium penyangga. Gel yang digunakan terbentuk dari polimerisasi akrilamida dengan N, N’-metilena bis akrilamida sehingga terbentuk ikatan silang karena polimerisasi akrilamida sendiri hanya menghasilkan ikatan linear yang tidak membentuk gel kaku (Girindra 1993). Polimerisasi dapat terjadi dengan cepat pada suhu kamar dengan adanya katalis dan inisiator. Katalis dan inisiator yang umum
digunakan ialah N, N’, N’,
N’-tetrametilenadiamina (TEMED) dan amonium persulfat (APS) sebagai sumber radikal bebas yang akan menginisiasi pembentukan polimer (Caprette 2005). Pada metode ini, digunakan natrium dodesil sulfat (SDS) dan β-merkaptoetanol. SDS merupakan detergen
anionik yang bersama dengan
β-merkaptoetanol dan pemanasan menyebabkan rusaknya struktur tiga dimensi protein menjadi konfigurasi acak. Hal ini disebabkan oleh pecahnya ikatan disulfida yang selanjutnya tereduksi menjadi gugus-gugus sulfidril. Umumnya analisis dengan elektroforesis protein menggunakan gel poliakrilamida dengan konsentrasi yang sesuai dengan bobot proteinnya (Tabel 1).
Pergerakan partikel di dalam medium bergantung pada ukuran partikel dan ukuran medium penunjang. Ukuran pori dari gel akan
ditentukan oleh konsentrasi gel
poliakrilamida. Protein yang besar mempunyai mobilitas yang lebih lambat dibandingkan dengan kompleks protein yang lebih kecil. Bobot molekul protein dapat ditentukan dengan kalibrasi menggunakan standar protein yang sudah diketahui bobot molekulnya (Rybicki et al. 1996). Teknik elektroforesis gel banyak digunakan baik di bidang kimia maupun biokimia, karena teknik ini memiliki banyak keuntungan, diantaranya memiliki daya resolusi tinggi, sederhana, dan mudah dibawa (Girindra 1993).
Tabel 1 Variasi konsentrasi gel berdasarkan bobot protein
% gel Bobot protein (kDa)
7 50 – 500
10 20 – 300
12 10 – 200
15 3 – 100
Sumber: Laemmli (1970)
Kromatografi Gel Filtrasi Kromatografi adalah metode pemisahan yang dapat digunakan untuk memisahkan suatu komponen dari komponen lainnya atau memisahkan komponen dari sekumpulan
komponen lainnya. Metode ini merupakan teknik yang efektif dan dapat digunakan untuk memisahkan komponen yang sulit dipisahkan dengan metode lain (Wilson & Walker 1994).
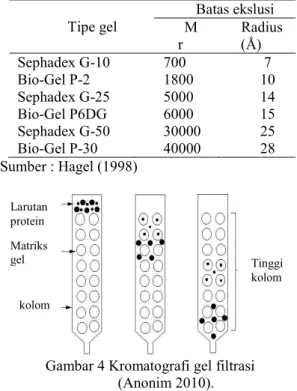
Kromatografi gel filtrasi merupakan teknik pemisahan yang berdasarkan pada ukuran dan atau bentuk dari partikel analit. Pemisahan tersebut dilakukan menggunakan matriks yang berpori. Masing-masing molekul memiliki tingkatan yang berbeda untuk melewati pori tersebut (molekul yang lebih kecil memilki kemampuan lebih tinggi untuk melewati pori tesebut dibandingkan dengan molekul yang lebih besar), sehingga menyebabkan pemisahan analit. Batasan pemisahan dari sebuah ukuran yang dipisahkan merupakan indikasi bobot molekul, umumnya untuk tipe polimer, dari analit tersebut (Hagel 1998). Ukuran protein yang dapat dipisahkan oleh beberapa matriks gel dapat dilihat pada Tabel 2.
Pemisahan molekul yang terjadi yaitu molekul yang memiliki ukuran besar akan terelusi oleh fase gerak dari kolom kromatografi dan akan keluar paling awal melalui ruang antar matriks dengan laju alir yang tinggi. Molekul yang berukuran kecil akan terelusi ke dalam fase diam oleh fase gerak dengan laju alir yang rendah, sehingga akan keluar dari kolom paling akhir (Wilson & Walker 1994). Ilustrasi pemisahan protein yang terjadi pada kromatografi kolom ditunjukkan pada Gambar 4.
Tabel 2 Ukuran protein minimum yang dapat dipisahkan oleh matrik gel
Sumber : Hagel (1998)
Gambar 4 Kromatografi gel filtrasi (Anonim 2010). Tipe gel Batas ekslusi M r Radius (Å) Sephadex G-10 Bio-Gel P-2 Sephadex G-25 Bio-Gel P6DG Sephadex G-50 Bio-Gel P-30 700 1800 5000 6000 30000 40000 7 10 14 15 25 28 Tinggi kolom Matriks gel Larutan protein kolom
BAHAN DAN METODE
Bahan dan AlatBahan-bahan yang digunakan untuk isolasi enzim RNA Helikase meliputi bakteri E.coli pET BL21 (DE3) pLysS yang membawa gen NS3 RNA helikase virus hepatitis C dalam plasmid pET 21b (koleksi Andi Utama), media Luria Bertani broth (LB), akuades, ampisilin, bufer B (10 mM Tris HCl pH 8.5, 0.3 M isopropil β-D-thiogalaktopiranosida (IPTG), resin Talon, dan bufer elusi (400 mM imidazola dalam bufer B).
Bahan-bahan yang digunakan untuk isolasi dan pemurnian kapang endofit CgKTm 5 F adalah isolat kapang endofit CgKTm 5 F (koleksi Titik K Prana, Laboratorium Biopolimer Puslit Bioteknologi LIPI), media potato dextrose broth (PDB), potato dextrose brothi (PDA), 0.1 mM ATP (Adenosin trifosfat), 1 mM MgCl2, larutan hijau malakit,
0.1 mM MOPS (4-Morpholinopropane
sulphonic acid), 2.3% polivinil alkohol, amonium molibdat, natrium sitrat, gel Sephadex G-50, metanol, akuabides, amonium sulfat, amonium persulfat, N, N’, N’, N’-tetrametilenadiamina (TEMED), Tris HCl pH 7.4, Tris pH 6.8 mengandung 0.4% SDS, Tris pH 8.8, sukrosa, akuades, dan pewarnaan perak.
Alat-alat yang digunakan untuk isolasi RNA helikase adalah sonikator (Labsonic), ultrasentrifugasi Sorvall RC-26 plus, tabung sentrifus, Erlenmeyer 2000 mL, inkubator berpenggoyang, spin down, dan rotator (N-Biotec). Peralatan untuk pemurnian protein inhibitor kapang endofit CgKTm 5 F meliputi 96-well microplate (Nalge Nunc), pipet mikro, neraca analitik, peralatan gelas, microplate reader (Multiscan EX Thermo), chamber SDS PAGE, laminar, oven, kolom gel filtrasi, tabung falcon, dan vial.
Metode
Isolasi dan Pemurnian RNA Helikase HCV (Utama et al. 2000)
Isolasi RNA helikase HCV. Sebanyak 10 mL kultur E.coli pET BL21(DE3) pLysS yang membawa gen NS3 RNA helikase HCV diinokulasikan ke dalam 400 mL medium LB cair yang mengandung ampisilin. Selanjutnya, kultur tersebut diinkubasi di dalam inkubator berpenggoyang pada suhu 37oC dengan kecepatan 200 rpm selama 30 menit. Setelah 30 menit kultur tersebut dihitung OD600. Apabila telah mencapai ± 0.3, maka
ditambahkan 0.3 M IPTG ke dalam kultur tersebut. Kultur E.coli selanjutnya diinkubasi di dalam inkubator berpenggoyang dengan suhu 37oC dengan kecepatan 200 rpm selama 3 jam atau nilai OD600 mencapai ± 1, kemudian kultur tersebut disentrifugasi pada suhu 4oC dengan kecepatan 7000 g selama 10 menit. Pelet yang diperoleh kemudian diresuspensikan kembali menggunakan 5 mL medium LB cair. Selanjutnya, hasil resuspensi pelet tersebut disentrifugasi kembali pada suhu 4⁰C dengan kecepatan 7000 g selama 10 menit. Pelet yang dihasilkan pada proses sebelumnya diberi perlakuan pengering bekuan (freeze & thawing) sebanyak 3 kali. Hasil freeze thawing tersebut kemudian disonikasi (amplitudo 40; siklus 0.5; waktu 3 x 15 detik; interval waktu 1 menit). Suspensi sel disentrifus dengan kecepatan 12000 g, selama 10 menit, pada suhu 4oC. Pelet kemudian dibuan sedangkan supernatan dikoleksi.
Pemurnian Enzim RNA Helikase Virus Hepatitis C. Supernatan hasil sentrifugasi pada tahap isolasi kemudian dicampur dengan resin TALON yang telah diseimbangkan dengan buffer B. Resin TALON yang sudah diseimbangkan kemudian dicampur dengan supernatan menggunakan pemutar selama 3 jam dalam ruang pendingin (4oC). Setelah itu, disentrifus selama 10 menit dengan kecepaatan 5000 g pada suhu 4oC. Supernatan dibuang kemudian ditambahkan buffer B sebanyak 15 ml, dan disentrifus kembali pada kecepatan 5000 g selama 7 menit. Pelet kemudian ditambahkan bufer elusi dan diinkubasi semalam pada rotator yang ditempatkan pada suhu 4oC. Setelah itu, supernatan diambil dan dipisahkan dari pelet dengan sentrifugasi dengan kecepatan 5000 g selama 3 menit pada suhu 4oC. Supernatan merupakan enzim RNA helikase yang telah dipurifikasi.
Uji aktivitas ATPase RNA Helikase HCV (Utama et al. 2000)
Pengujian aktivitas RNA helikase dilakukan dengan uji ATPase secara kolorimetri (Utama et al. 2000). Pengujian ini dimaksudkan untuk mengukur jumlah fosfat yang dilepaskan dari hidrolisis senyawa ATP menjadi ADP. Sebanyak 50 µL campuran reaksi yang dimasukkan ke dalam satu sumur mengandung 5 µL 10 mM bufer MOPS (pH 6.5), 1 mM ATP, 0.5 µL 1 mM MgCl, 38.5 µL H2O, dan 5 µL RNA helikase HCV. Campuran reaksi tersebut diinkubasikan di dalam 96-well microplate, pada suhu ruang
selama 45 menit. Setelah itu reaksi tersebut divisualisasikan menggunakan pewarnaan dengan cara penambahan 100 µL larutan pewarna pada masing-masing sumur. Larutan
pewarna yang digunakan merupakan
campuran 0.081% hijau malakit, H20, 5.7% amonium molibdat dalam 6 M HCl, dan 2.3% polivinil alkohol dengan perbandingan 2:2:1:1. Pewarnaan tersebut diinkubasi selama 5 menit. Setelah masa inkubasi dilakukan penghentian reaksi pewarnaan dengan menambahkan 25 µL natrium sitrat. Hasil reaksi diukur pada panjang gelombang 620 nm dengan referensi 405 nm.
Isolasi Protein Inhibitor RNA Helikase HCV (Hairany 2010)
Optimasi masa inkubasi isolat CgKTm 5 F. Isolat CgKTm 5 F diinokulasikan sebanyak
satu ose ke dalam Erlenmeyer yang berisi medium PDB 25 mL sebagai media prekultur. Selanjutnya, media tersebut diinkubasi pada suhu ruang selama 3 hari dengan bantuan alat berpenggoyang. Setelah 3 hari media tersebut dipindahkan ke dalam media PDB 250 mL sebanyak 2.5 mL. Media produksi tersebut diinkubasi pada suhu ruang selama 10 hari dengan bantuan alat berpenggoyang. Pada rentang masa inkubasi dilakukan pengambilan sampel setiap hari untuk dihitung aktivitas inhibisi terhadap RNA helikase HCV dan produksi protein inhibitor. Sebanyak 5 mL biakan diambil dalam setiap pengambilan sampel. Sampel tersebut selanjutnya disentrifugasi dengan kecepatan 12.000 g selama 10 menit. Filtrat yang diperoleh dipisahkan dari endapan dan digunakan sebagai ekstrak kasar protein inhibitor. Selanjutnya diuji aktivitas inhibitornya terhadap aktivitas ATPase dan kadar proteinnya.
Optimasi pengendapan protein. Supernatan yang merupakan ekstrak kasar
dioptimasi fraksinasi proteinnya
menggunakan amonium sulfat berdasarkan metode Scope (1987) dengan berbagai tingkatan pengendapan. Supernatan tersebut diendapkan menggunakan amonium sulfat 60%, 70%, 80%, dan 90% (w/v). Pelarutan amonium sulfat dibantu dengan pengaduk magnetic stirrer dengan kecapatan 20 rpm pada suhu 4⁰C. Hasil pengendapan tersebut disimpan selama 12 jam pada suhu 4⁰C. Selanjutnya dilakukan sentrifugasi terhadap hasil pengendapan menggunakan amonium sulfat tersebut dengan kecepatan 12.000 g selama 30 menit pada 4⁰C. Pelet yang didapatkan kemudian diresuspensikan
kembali menggunakan bufer Tris-HCl pH 7.4. Selanjutnya diuji aktivitasnya menggunakan uji ATPase.
Produksi protein inhibitor dari isolat CgKTm 5 . Isolat CgKTm5 F diinokulasikan
sebanyak satu ose ke dalam Erlenmeyer yang berisi media PDB 25 mL sebagai media pre-kultur. Selanjutnya, media tersebut diinkubasi pada suhu ruang selama 3 hari dengan bantuan alat berpenggoyang. Setelah 3 hari, media tersebut dipindahkan ke dalam media produksi PDB 250 mL sebanyak 2.5 mL. Media produksi tersebut diinkubasi pada suhu ruang sesuai dengan hasil optimasi dengan bantuan shaker.
Setelah mencapai masa inkubasi optimum, kultur kapang endofit CgKTm 5 F disentrifugasi dengan kecepatan 12.000 g selama 30 menit pada suhu 4⁰C. Supernatan dikoleksi, sedangkan peletnya dibuang. Supernatan yang merupakan ekstrak kasar,
kemudian proteinnya diendapkan
menggunakan amonium sulfat. Sebelum dilakukan pengendapan diuji aktivitas inhibisinya menggunakan uji ATPase.
Pengendapan protein inhibitor. Tahapan ini dilakukan sesuai dengan hasil optimasi pengendapan protein yang dilakukan sebelumnya Selanjutnya diuji aktivitas inhibisinya menggunakan uji ATPase.
Pemurnian Protein Inhibitor RNA Helikase HCV (Hairany 2010)
Protein yang telah diendapkan
mengunakan amonium sulfat selanjutnya dimurnikan menggunakan kromatografi gel filtrasi. Fase diam yang digunakan adalah Sephadex G-50.
Kolom kromatografi yang sudah
dipadatkan dicuci menggunakan ddH2O sebanyak 1.5 kali volume kolom dengan laju alir 0.5 mL/menit. Setelah itu, sebanyak 1 mL sampel protein inhibitor diinjeksikan ke dalam kolom. Eluen yang digunakan adalah larutan H2O:MeOH (6:4) dengan laju alir sebesar 1 mL/menit. Hasil elusi ditampung dalam tabung vial dengan volume masing-masing 1 mL. Keseluruhan tahapan ini dilakukan di suhu 4oC. masing-masing fraksi diuji aktivitas inhibisinya dengan uji ATPase dan dikarakterisasi proteinnya.
Pengukuran Kadar Protein Inhibitor RNA Helikase HCV (PIERCE 2003)
Kadar protein diukur dengan Bichincionic Acid (BCA) Protein Assay kit. Sebanyak 0.1 mL sampel ditambahkan 2 mL working reagent. Campuran tersebut kemudian
diinkubasi pada suhu 37ºC selama 30 menit. Absorbansi diukur menggunakan panjang gelombang 562 nm. Standar protein yang digunakan adalah albumin serum sapi (Bovine Serum Albumin (BSA)) pada kisaran 0.05 – 0.5 mg/mL.
Analisis Bobot Molekul Protein Inhibitor RNA Helikase HCV (Hairany 2010 termodifikasi)
Analisis bobot molekul protein inhibitor dari berbagai tahap dilakukan dengan metode sodium dedosil sulfate polyacrilamide gel electrophoresis (SDS-PAGE). Komposisi separating gel yang digunakan sebesar 18% akrilamid. Gel kemudian dielektroforesis pada 30 mA selama 60 menit dalam bufer elektroforesis (24% gliserol, 8% SDS, 100 mM Tris). Setelah itu, gel tersebut diwarnai dengan pewarnaan perak.
HASIL DAN PEMBAHASAN
Hasil Isolasi RNA Helikase HCV Bakteri E.coli BL21 pET 21b (D3E) pLysS yang telah disisipi oleh gen NS3 RNA helikase HCV dikulturkan pada media LB. Kultur tersebut diinkubasi pada suhu 37oC dan digojok dengan kecepatan 150 rpm. Kultur yang telah memasuki fase logaritmik yaitu setelah mencapai nilai optical density (OD600) sebesar 0.3 diinduksi menggunakan IPTG. Penambahan IPTG bertujuan menginduksi pengekspresian gen NS3 yang telah disisipkan, sehingga akan terjadi ekspresi berlebih pada gen tersebut. Tujuan dari penginduksian tersebut yaitu meningkatkan produksi RNA helikase yang ditranslasikan karena ekspresi berlebih terjadi pada gen NS3. Pemanenan kultur dilakukan ketika memasuki fase stasioner dengan nilai OD600 sebesar 1. RNA helikase terekspresi secara intraseluler, sehingga pengisolasiannya dilakukan pengkoleksian E. coli terlebih dulu menggunakan teknik sentrifugasi. Sentrifugasi akan memisahkan sel (pelet) dengan medianya (supernatan). Pelet yang telah dikoleksi selanjutnya akan memasuki tahap pemecahan sel untuk mengeluarkan enzim RNA helikase yang terdapat didalam sel.Pemecahan sel dilakukan sebanyak dua kali, yang pertama menggunakan teknik pengeringbekuan (freeze & thawing) dan sonikasi. Teknik pengeringbekuan pada sel E.coli menyebabkan pembekuan yang cepat pada cairan intraselular dan ekstraselular,
sehingga akan membentuk kristal es pada intraselular dan ekstraselular yang akan menyebabkan kerusakan terhadap sel (Scawen & Meling 1985). Setelah tahap pengeringbekuan, disonikasi terhadap sel tersebut dengan tujuan semua sel tersebut dapat dipecah dan dikoleksi enzimnya.
Pada waktu sonikasi, sel tersebut ditambahkan bufer B (Tween 20, Tris HCl,
dan NaCl) yang berfungsi untuk
mempertahankan aktivitas RNA helikase HCV. Tris HCl digunakan untuk menjaga keseimbangan pH larutan, agar RNA helikase HCV tidak mengalami denaturasi dan menurun aktivitasnya. Penambahan NaCl pada larutan bufer ini berfungsi menghilangkan asam nukleat dan kontaminan lainnya yang berikatan tidak spesifik dengan RNA helikase HCV dengan cara interaksi ionik (Vanz et al. 2008). Tween 20 digunakan untuk menghancurkan lipid bipolar pada membran sel, sehingga bagian hidrofobik dari enzim RNA helikase yang terikat pada membran sel dapat terdisosiasi. Detergen ini juga dapat melarutkan enzim tersebut di dalam kondisi tidak terdenaturasi dan menjaga kelangsungan aktivitas biologis. Selanjutnya dilakukan sentrifugasi dengan tujuan memisahkan semua bagian sel yang terlarut pada air termasuk RNA helikase dengan pecahan-pecahan sel (Moradpur et al. 2007).
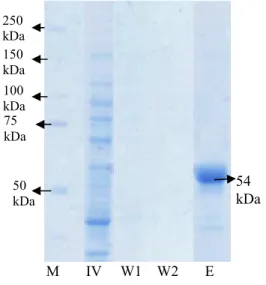
Hasil Pemurnian RNA Helikase HCV Semua tahapan isolasi dan pemurnian enzim RNA helikase di analisis menggunakan SDS-PAGE untuk menentukan kemurnian
dari enzim tersebut. Gambar 5
memperlihatkan enzim RNA helikase yang telah termurnikan memiliki ukuran sebesar 54 kDA (E) (Lampiran 2). Ukuran RNA helikase ini sesuai dengan pemurnian yang telah dilakukan Utama et al. (2000). Lajur inner volume (IV) terdapat banyak pita protein, karena merupakan supernatan hasil sonikasi yang mengandung metabolit intraseluler yang belum dimurnikan. Lajur washing 1 (W1) dan washing 2 (W2) merupakan bufer hasil pencucian enzim. Kedua lajur tidak terdapat pita protein.
Kromatografi afinitas digunakan dalam tahapan pemurnian RNA helikase HCV. Teknik ini dapat mengikat enzim RNA helikase HCV secara spesifik yang terdapat pada supernatan hasil sonikasi. Resin yang digunakan pada tahapan ini yaitu TALON logam afinitas yang secara spesifik dapat mengikat RNA helikase yang memiliki penanda 6xHis-Tag. Pengikatan residu His
dilakukan oleh logam Co2+ yang terdapat dalam resin TALON. Penandaan yang terdapat pada enzim RNA helikase yaitu ujung His dilakukan pada saat konstruksi gen NS3 yang disisipkan pada E.coli. Setelah enzim tersebut terikat pada resin TALON, kemudian dipisahkan dengan protein lainnya dengan cara sentrifugasi. RNA helikase yang terikat pada resin akan terdapat pada bagian pelet, sedangkan protein lainnya terpisahkan pada bagian supernatan. Tahapan selanjutnya yaitu memutus ikatan antara resin TALON dan ujung His.
Pemutusan ini dilakukan dengan cara menambahkan bufer elusi pada resin yang telah mengikat enzim. Bufer elusi mengandung imidazol dan bufer B. Imidazol berfungsi sebagai analog pengganti residu His enzim yang diikat oleh logam Co2+, sehingga resin tersebut akan memutus ikatannya dengan enzim RNA helikse dan mengikat imidazol sebagai penggantinya. Teknik sentrifugasi digunakan untuk memisahkan resin yang mengikat imidazol dan enzim yang telah murni. Sentrifugasi pada kecepatan rendah bertujuan mengurangi kerusakan yang terjadi pada enzim.
Gambar 5 Elektroforegram SDS-PAGE isolasi dan pemurnian RNA helikase HCV; (M) Marker; (IV) inner volume; (W1) Hasil washing 1; (W2) Hasil washing 2; (E) Enzim. Waktu Pemanenan Protein Inhibitor dari
Kapang Endofit CgKTm 5 F Isolat kapang CgKTm 5 F merupakan kapang endofit yang berasal dari gembyok (temu putih atau Curcuma zedoaria). Isolat ini ditumbuhkan pada media Potato Dextrose Broth (PDB). Pertumbuhan kapang ini pada media PDB dapat terlihat pada kekeruhan
yang terjadi pada media tersebut yang tadinya berwarna kuning jernih menjadi kuning keruh. Media PDB merupakan media yang telah memenuhi syarat minimum pertumbuhan kapang, karena memiliki sumber karbon yang berasal dari dekstrosa dan pati kentang serta sumber nitrogen yang berasal dari asam amino yang terdapat pada kentang (Hadioetomo 1993).
Pertumbuhan dari kapang CgKTm 5 F juga dipengaruhi oleh masa inkubasi dan agitasi. Masa inkubasi yang dibutuhkan oleh isolat ini untuk menghasilkan protein inhibitor RNA helikase HCV optimum pada 7 hari. Hal ini ditentukan berdasarkan perhitungan inhibisi protein tersebut terhadap RNA helikase HCV setiap hari selama masa inkubasi. Masa inkubasi 7 hari merupakan masa inkubasi yang paling optimum yang dibutuhkan oleh kapang isolat ini untuk memproduksi protein tersebut dengan nilai inhibisi sebesar 85.86% (Gambar 6). Agitasi bertujuan meningkatkan aerasi dan distribusi nutrisi sehingga sel kapang dapat mencapai kondisi yang optimum (Rahman 2009).
Pemanenan protein inhibitor RNA helikase HCV yang dihasilkan oleh kapang CgKTm 5 F dengan teknik sentrifugasi untuk memisahkan protein tersebut dengan sel kapangnya. Supernatan selanjutnya digunakan untuk proses isolasi protein yang mempunyai aktivitas inhibisi terhadap RNA helikase HCV.
Gambar 6 Aktivitas inhibisi RNA helikase HCV pada berbagai waktu inkubasi kapang endofit CgKTm 5 F.
Isolat Protein Inhibitor RNA Helikase HCV
Tahap isolasi protein dari fraksi
supernatan diendapkan menggunakan
amonium sulfat. Pengendapan protein M IV W1 W2 E 250 kDa 150 kDa 75 kDa 100 kDa 50 kDa 54 kDa
menggunakan amonium sulfat merupakan teknik yang dapat memenuhi dua tujuan sekaligus yaitu pemurnian dan pengendapan protein yang spesifik. Penggunaan amonium sulfat umum digunakan dalam proses pengendapan ini karena memiliki beberapa kelebihan yaitu memiliki kelarutan yang tinggi, tingkat toksisitas yang rendah untuk sebagian besar protein, murah, dan pada beberapa kondisi memiliki efek penstabil pada protein. Protein akan terendapkan karena molekul air yang berikatan dengan ion-ion garam semakin banyak. Hal ini menyebabkan terjadinya penarikan selubung air yang mengelilingi permukaan protein sehingga mengakibatkan protein saling berinteraksi dan beragregasi (Scawen & Melling 1985).
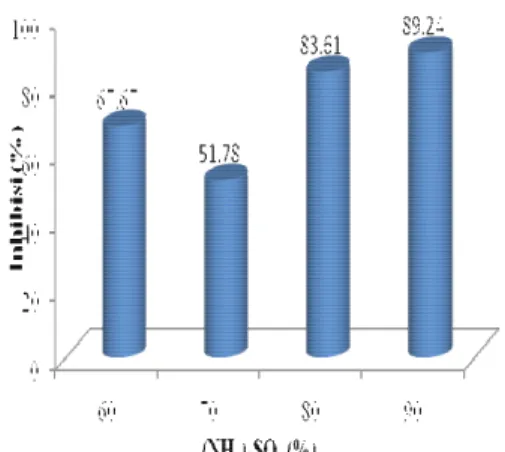
Pengendapan protein target yang terdapat pada fraksi supernatan dilakukan pada konsentrasi amonium sulfat 90% (w/v). Hasil pengendapan pada konsentrasi ini memilki aktivitas inhibisi terhadap RNA helikase HCV paling tinggi dibandingkan dengan hasil pengendapan amonium sulfat dengan konsentrasi yang lainnya yaitu sebesar 89.24% (Gambar 7). Perbedaan aktivitas inhibisi yang diperlihatkan oleh beberapa hasil pengendapan protein dimungkinkan karena protein yang terendapkan pada berbagai konsentrasi pengendapan tersebut berbeda. Karena protein yang terendapkan akan mengalami fase pengendapan yang berbeda-beda sesuai dengan titik isolistriknya. Titik isolistrik ini tercapai karena kekuatan ionik yang terdapat pada garam dan garam dengan nilai valensi yang tinggi memiliki kemampuan yang lebih besar dalam mengendapkan dibandingkan dengan garam yang memiliki nilai valensi yang rendah (Scawen & Meling 1985).
Gambar 7 Aktivitas inhibisi RNA helikase HCV pada beberapa tingkatan pengendapan amonium sulfat.
Protein yang terendapkan menggunakan amonium sulfat dilarutkan kembali menggunakan larutan bufer Tris HCl pH 7.4 (Hairany 2010). Penggunaan bufer fosfat tidak dapat dilakukan karena dapat mempengaruhi hasil perhitungan aktivitas inhibisi RNA helikase HCV, karena menggunakan uji ATPase yang menghitung fosfat anorganik yang bebas.
Hasil Pemurnian Protein Inhibitor dari Kapang CgKTm 5 F
Pemurnian protein inhibitor RNA helikase HCV dari kapang endofit CgKTm 5 F dilakukan dengan kromatografi gel filtrasi. Kromatografi gel filtrasi merupakan teknik pemisahan campuran senyawa berdasarkan bobot molekulnya. Ukuran yang dapat dipisahkan fase diam tergantung pada poros matriksnya. Fase diam yang digunakan yaitu
Sephadex G-50 dengan kemampuan
memisahkan molekul protein sampai 30 kDa. Fase gerak yang digunakan yaitu metanol 40%. Hasil dari pemisahan protein menggunakan kolom kromatografi gel filtrasi didapatkan fraksi sebanyak 28 fraksi. Semua fraksi yang didapatkan selanjutnya diuji aktivitas inhibisinya dengan uji ATPase.
Uji ATPase merupakan metode yang digunakan untuk menghitung pelepasan fosfat anorganik yang berasal dari ATP dengan bantuan enzim ATPase. Penggunaan uji ATPase pada penentuan aktivitas dari enzim RNA helikase HCV karena enzim ini memiliki aktivitas yang distimulasi oleh ATP. Larutan campuran dan larutan pewarna digunakan dalam uji ATPase. Larutan
campuran terdiri atas asam
4-morfolinopropanafosfat sulfonat (MOPS), ATP, dan MgCl2. MOPS berfungsi sebagai bufer dalam larutan campuran. ATP berfungsi sebagai substrat yang akan dihidrolisis oleh enzim RNA helikase HCV menjadi ADP dan fosfat anorganik bebas (Pi). Selanjutnya yang terakhir yaitu Mg2+ berfungsi sebagai kofaktor dari RNA helikase HCV. Kuo et al. (1997) menyebutkan bahwa RNA helikase dari golongan flavivirus memerlukan Mg2+ atau Mn2+ untuk mengoptimalkan aktivitasnya dan diinbihisi oleh keberadaan K+.
Larutan pewarna terdiri atas hijau malakit, amonium molibdat, polivinil alkohol, dan akuades. Pereaksi hijau malakit dan amonium molibdat berfungsi sebagai pembentuk warna hijau kebiruan. Warna yang terbentuk merupakan hasil reaksi antara kedua pereaksi tersebut dengan Pi menjadi kompleks fosfomolibdat. Polivinil alkohol berfungsi
sebagai pencegah terbentuknya endapan akibat reaksi protein dengan kompleks fosfomolibdat (Chan et al. 1986).
Penghentian reaksi warna dengan penambahan Na-sitrat terhadap campuran. Hal ini dilakukan karena sitrat dapat berikatan dengan molibdat yang bebas, sehingga mencegah proses pembentukan warna yang berlebihan. Molibdat bebas tersebut dapat berikatan dengan ATP labil yang dapat terhidrolisis selama proses uji berlangsung.
Penambahan sitrat tersebut dapat
mempertahankan warna secara stabil sampai enam jam (Gawronski dan Benson 2004).
Berdasarkan hasil uji ATPase didapatkan bahwa fraksi ke-8 mempunyai aktivitas inhibisi RNA helikase HCV tertinggi sebesar 64.11% (Gambar 8). Kenaikan aktivitas inhibisi terhadap RNA helikase HCV terjadi pada fraksi-fraksi awal. Hal ini menunjukkan bahwa protein yang memiliki bobot molekul tinggi yang memiliki aktivitas inhibisi terhadap RNA helikase HCV. Protein inhibitor tersebut diperkirakan menghambat RNA helikase secara alosterik. Inhibitor menempel pada enzim selain di situs katalitiknya sehingga merubah konfromasi enzim. Perubahan tersebut menyebabkan interaksi enzim-substrat berkurang sehingga tidak ada produk yang dihasilkan (Boroswki et al. 2008).
Gambar 8 Aktivitas inhibisi RNA helikase HCV fraksi hasil kromatografi gel filtrasi.
Bobot Molekul Protein Inhibitor RNA Helikase HCV
Penentuan bobot molekul protein penghambat RNA helikase HCV yang berasal dari kapang endofit CgKTm 5 F dilakukan dengan metode SDS-PAGE. Konsentrasi media penyangga yang digunakan bergantung pada bobot molekul campuran protein yang akan dipisahkan. Penggunaan konsentrasi gel sebesar 20% pada penelitian ini dikarenakan
dugaan protein yang akan dipisahkan memiliki kisaran yang lebih rendah dari 3 kDa.
Fraksi terbaik hasil kolom kromatografi ini yaitu fraksi ke- 8 setelah dianalisis menggunakan teknik SDS-PAGE memilki empat pita protein (Gambar 9). Semua pita tersebut menunjukkan bahwa molekul protein yang terdapat pada fraksi ke-8 memiliki bobot diatas 17 kDa. Hal ini dimungkinkan karena pada fraksi-fraksi awal yang terpisahkan merupakan molekul yang memiliki bobot molekul besar. Bobot molekul relatif (Lampiran 3) dari keempat pita protein berturut turut yaitu 47.2 kDa, 31.9 kDa, 25.4 kDa, dan 20.2 kDa untuk pita protein 1, 2, 3, dan 4.
Hasil analisis pada fraksi ke- 8 memiliki kesamaan pita pada analisis hasil pengendapan amonium sulfat. Jumlah pita protein yang lebih sedikit dibandingkan dengan hasil pengendapan amonium sulfat. Hal ini menunjukkan bahwa kromatografi gel filtrasi memperkecil jumlah molekul protein dari hasil pengendapan amonium sulfat. Pewarna perak memiliki sensitivitas yang lebih baik dibandingkan dengan pewarna commasie blue (Rozaida et al. 2003). Konsentrasi protein yang kemungkinan
rendah setelah proses pemurnian
menggunakan kromatogarafi kolom, dengan metode pewarnaan ini masih bisa dilihat secara jelas.
Gambar 9 Elektroforegram protein CgKTm 5 F; (M) Marker; (Cr) pengendapan amonium sulfat; (F8) Fraksi ke-8 hasil pemurnian; (1, 2, 3, dan 4) pita-pita protein pada fraksi ke-8 kolom kromatografi. 1 42 kDa 26 kDa 17 kDa 10 kDa 4.6 kDa 2 3 4
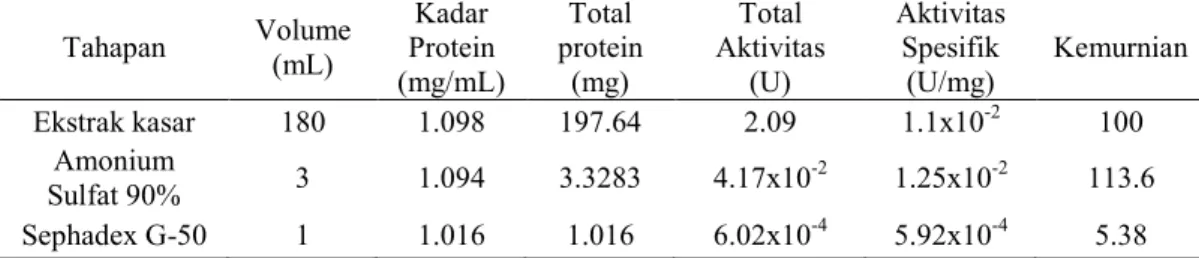
Tabel 3 Kadar dan aktivitas spesifik protein inhibitor RNA helikase HCV dari kapang endofit CgKTm 5 F Tahapan Volume (mL) Kadar Protein (mg/mL) Total protein (mg) Total Aktivitas (U) Aktivitas Spesifik (U/mg) Kemurnian Ekstrak kasar 180 1.098 197.64 2.09 1.1x10-2 100 Amonium Sulfat 90% 3 1.094 3.3283 4.17x10 -2 1.25x10-2 113.6 Sephadex G-50 1 1.016 1.016 6.02x10-4 5.92x10-4 5.38
Konsentrasi Protein Inhibitor RNA Helikase HCV.
Konsentrasi protein tiap tahapan pada penelitian ini ditentukan dengan uji Bichincionic Acid (BCA) protein kit assay. Prinsip uji ini yaitu pereduksian Cu2+menjadi Cu+ oleh protein pada media alkalin (reaksi biuret). Kation cupro yang terbentuk akan bereaksi dengan asam bicinchoninat membentuk warna ungu yang dapat dideteksi pada panjang gelombang 562 nm. Uji ini dapat mendeteksi konsentrasi protein antara 20-2000 µg/mL (PIERCE 2003).
Tabel 3 memperlihatkan aktivitas spesifik dari protein inhibitor pada beberapa tahapan isolasi dan pemurnian yang dilakukan. Unit pada aktivitas protein inhobitor didefinisikan sebagai jumlah aktivitas protein yang dibutuhkan untuk menghambat 1 mol substrat menjadi produk. Protein yang telah diendapkan dengan amonium sulfat 90% memiliki aktivitas yang tidak berbeda jauh dalam menghambat aktivitas RNA helikase HCV dibandingkan dengan ekstrak kasarnya yaitu sebesar 1.25x10-2 U/mg dengan tingkat kemurnian sebesar 113.6. Hal ini dikarenakan amonium sulfat tidak hanya mengendapkan protein yang memiliki aktivitas penghambatan RNA helikase HCV melainkan juga beberapa protein lainnya yang terdapat pada ekstrak kasar.
Aktivitas spesifik dari protein inhibitor hasil pemurnian menggunakan kromatografi gel filtrasi Sephadex G-50 lebih kecil dibandingkan dengan aktivitas ekstrak kasarnya sebesar 5.92x10-4 U/mg dan memilki kemurnian sebesar 5.38. Nilai aktivitas dan kemurnian yang kecil dari hasil kromatografi kolom dimungkinkan protein yang didapatkan dalam konsentrasi yang kecil, sebab sebelum dimasukkan pada kolom kromatografi proteinnya tidak dipekatkan terlebih dahulu (Udin et al. 1996). Selain itu, diperkirakan terjadi autolisis pada protein saat proses pemurnian terjadi (Scopes 1987). Proses autolisis yang terjadi disebabkan oleh suhu saat pemurnian dilakukan tidak stabil.
SIMPULAN DAN SARAN
SimpulanRNA helikase HCV yang telah dimurnikan memilki ukuran 54 kDa. Protein inhibitor yang telah dimurnikan menggunakna kromatografi gel filtrasi dari kapang CgKTm 5 F dapat menginhibisi RNA helikase dengan aktivitas inhibisi sebesar 64.11%. Protein tersebut dipanen setelah masa inkubasi 7 dan diisolasi dengan ammonium sulfat 90% w/v. Protein inhibitor tersebut mempunyai memiliki empat pita, dengan bobot 47.2 kDa, 31.9 kDa, 25.4 kDa, dan 20.2 kDa. Aktivitas protein sebagai inhibitor dari setiap tahapan yaitu ekstrak kasar, hasil pengendapan amonium sulfat, dan hasil pemurnian berturut-turut sebesar 1.1x10-2 U/mg, 1.25x10-2 U/mg, dan 5.92x10-4 U/mg. Kemurnian setiap tahapan berturut-turut sebesar 100, 113.6, dan 5.38.
Saran
Pemurnian lanjutan terhadap protein inhibitor RNA helikase yang dihasilkan oleh kapang endofit CgKTm 5 F perlu dilakukan. Pemurnian lanjutan tersebut bisa dengan teknik KCKT preparatif dan kromatografi pertukaran ion. Karakterisasi dan optimasi terhadap protein inhibitor perlu juga dilakukan. Karakterisasi meliputi pengaruh pH, suhu, dan suhu penyimpanan protein terhadap aktivitas inhibisinya. Optimasi juga perlu dilakukan untuk meningkatkan aktivitas inhibisi dari protein inhibitor tersebut.
DAFTAR PUSTAKA
Anonim. 2010. Column Chromatography.[terhubung berkala]. Http://www. Newarkbioweb. Rutgers. Edu/bio301s/ Lab4-molwt-column chromatography.htm. [26 Desember 2010].
Bacon CW, White JW. 2000. Microbial Endhophytes. New York: Marcel Dekker Inc.
Borowski P et al. 2008. Viral NS3 helicase activity is inhibited by peptides reproducing the Arg-rich conserved motif of the enzyme (motif VI). Biochemical Pharmacology 76: 28–38.
Caprette DR. 2005. Preparing SDS-Gels. Experimental Biosciences. Introductory Laboratory-Bios 211. Texas: Rice University.
Chan Kwok Ming, Delfert D, Junger KD. 1986. A direct colorimetricc assay for Ca2+-stimulated ATPase activity. Anal Biochem 157: 375-380.
Cheng et al. 2007. NS3 helicase actively separates RNA strands and senses sequence barriers ahead of the opening fork. PNAS 104: 13954–13959.
Chungsamarnyart N, Sirinarumitr T, chumsing W, Wajjawalku W. 2007. in vitro study of antiviral activity of plant crude extract againts the foot and mouth disesease virus. Kasetsart Journal (National Science) 41: 97-103.
Fardiaz S. 1992. Mikrobiologi pangan I. Jakarta: Gramedia Pustaka Utama.
Gawronski JD, Benson DR. 2004. Microtiter assay for glutamin synthetase biosynthetic activity using inorganic phosphate detection. Anal Biochem 327: 114-118. Girindra A. 1993. Immunokimia. Bogor:
PAU-IPB.
Hagel L. 1998. Currenct Proocols in Protein Science. New York: John Willey & Sons Inc.
Hadioetomo RS. 1993. Mikrobiologi Dasar Dalam Praktek. Jakarta: Gramedia Pustaka Utama.
Hairany A. 2010. Pemurnian dan karakterisasi protein inhibitor RNA helikase virus hepatitis C dari Streptomyces chartreusis 5-095 [tesis]. Bogor: Pascasarjana IPB. Harvey D. 2000. Modern Analytical
Chemistry. Ed ke-1. New York: McGraw Hill.
Kadare G, Haenni A. 1997. Virus encoded RNA helicases. J of Virology 71: 2583-2590.
Kandil S et al. 2009. Discovery of a novel HCV helicase inhibitor by a de novo drug design approach. Bioorganic & Medicinal Chemistry Letters 19: 2935–2937
Kuo et al. 1997. Characterization of the NTPase activity of japanese enchephalitis virus NS3 protein. J General Virologi. 77: 2077-2084.
Laemmli UK. 1970. Cleavage of stuctural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685. Lauer GM, Walker BD. 2001. Review article: hepatitis c virus infection. N Engl J Med 1: 41-50.
Maksum R. 2005. Peranan bioteknologi dan mikroba endofit dalam pengembangan obat herbal. J Farmasi UI 2: 113-126. McHutchison JG, Patel K. 2002. Future
therapy of hepatisis C. Hepatology 36: 245-252.
Megawati. 2008. Penapisan senyawa inhibitor RNA helikase virus hepatitis C dari isolat kapang endofit Curcuma spp. [skripsi]. Jakarta: Fakultas Framasi, Universitas Pancasila.
Moradpour D, Penin F, Rice CM. 2007. Replication of hepatitis C virus. Nat Rev Microbiol 5:453–463.
PIERCE. 2003. Instruction BCATM Protein Assay Kit. Rockford: Pierce Biotechnology Inc.
Promega. 2003. Nucleic Acid Purification. Wisconsin: VWR International Inc. Qiagen. 2006. DNeasy Palnt Handbook.
Shanghai: F. Hoffman-La Roche Ltd. Rahman MN. 2009. Aktivitas senyawa
antibakteri hasil biotransformasi kurkumin oleh mikrob endofit asal kunyit [skripsi]. Bogor: FMIPA IPB.
Rozaida et al. 2003. An attempt to produce Brugia Malayi L3 Antigen Fractions for studies on antibody responses in JIRD (Meriones unguiculatus). Di dalam : seminar penyelidikan jangka pendek 2003; Kuala Lumpur , 12-13 Maret 2003. Kuala Lumpur: 1-5.
Rybicki EP et al. 1996. Molecular Biology Techniques Manual. Ed ke-3. Rondebosch: University of Capetown.
Sambrook J, Russel DW. 2001. Molecular cloning: laboratory manual Vol 3. Ed ke-3. Cold Spring Harbour: Spring Laboratory Pr.
Scawen MD, Melling J. 1985. Handbook of Enzyme Biotechnology. Alan Wiseman, editor. West Sussex: Ellis Horword Ltd. Schutz et al. 2002. Endophytic fungi: a source
of novel biologically active secondary metabolites. J Mycol. Res. 106: 996-1004. Scopes RK. 1987. Protein purification,
principles, and practices. Ed ke-2.. New York: Springer Verlag.
Strobel G, Daisy B. 2003. Bioprospecting for microbial endophytes and their natural products. Microbiology and Moleculer Biology Reviews 67: 491-502.
Suzuki et al. 2007. Review: molecular biology of hepatitis C virus. J Gastroenterol 42: 411-423.
Sy T, Jamal MM. 2006. Reviews:
Epidemiology of hepatitis C virus (HCV) Infection. Int. J. Med. Sci 3:41-46.
Tellinghuisen TL et al. 2001. Sudying hepatitis C virus: making the best of a bad virus. J Virology 81: 8853-8867.
Udin et al. 1996. Isolasi dan karakterisasi glukosa isomerase Aspergillus oryzae. Buletin IPT 4:36-40.
Utama A et al. 2000. Identifcation and characterization of the RNA helicase activity of japanese enchepalitis virus NS3 protein. FEBS Letters 456: 74-78.
Vanz et al. 2008. Human granulocyte colony stimulating factor (hG-CSF): Cloning, overexpression, purification, and characterization. Microbial Cell Factories. 7:13.
Wilson K, Walker JM. 1994. Principles and Techniques of Practical Biochemistry. Ed 4. Cambridge: Cambridge University Pr.
Lampiran 1 Diagram alir penelitian
Isolasi dan Pemurnian RNA Helikase HCV
Kultur E. coli BL 21 (D3E)
pLysS yang membawa gen
NS3 (RNA Helikase)
Isolasi RNA Helikase
HCV
Purifikasi RNA Helikase
HCVmenggunakan kromatografi
afinitas dengan resin Ni-NTA
RNA Helikase HCV
Lanjutan Lampiran 1
Isolasi dan Pemurnian Protein Inhibitor RNA Helikase HCV
Kultur kapang endofit
CgKtm 5 F
Isolasi Protein
Inhibitor RNA
Helikase HCV
Fraksi terbaik yang
menginhibisi RNA
helikase HCV
Purifikasi Protein inhibitor
menggunakan kromatografi
gel filtrasi
Uji ATPase,
SDS-PAGE, dan penentuan
konsentrasi protein
Uji ATPase,
SDS-PAGE, dan penentuan
Lampiran 2 Bobot molekul relatif enzim RNA helikase HCV
Bobot molekul relatif RNA helikase :
Rf
=
4
.
7
8
.
4
= 0.65
Y
= -1.091x + 2.44
Y
= -1.091(0.65) + 2.44
Y
= 1.73
Log BM = 1.73
BM
= 54 kDa
Bobot
molekul
(BM) marker
Log BM
Rf
250
150
100
75
50
2.39
2.17
2
1.87
1.69
0.12
0.22
0.37
0.48
0.74
Lampiran 3 Bobot molekul relatif protein kapang CgKTm 5 F
Bobot
molekul
(BM) marker
Log BM
Rf
42
26
17
10
4.6
1.62
1.41
1.23
1
0.66
0.12
0.21
0.39
0.58
0.75
Perhitungan bobot molekul relatif protein inhibitor kapang CgKTm 5 F pita nomor
3 :
Rf
=
4
.
7
8
.
1
Rf
= 0.25
y
= -1.42x + 1.76
y
= -1.42(0.25) + 1.76
y
= 1.41
Log BM = 1.41
BM
= 25. 4 kDa
Lampiran 4 Kurva standar Bovine Serum Albumin (BSA)
[BSA] (mg/mL)
Absorbansi 562 nm
0
25
125
250
500
750
1000
0
0.078
0.2495
0.383
0.649
0.911
1.179
Perhitungan kadar protein untuk ekstrak kasar protein CgKTm 5 F :
y
= 0.0011x + 0.0623
0.0635 = 0.0011x + 0.0623
x
=
0011
.
0
0623
.
0
0635
.
0
−
= 1.098 mg/mL
Lampiran 5 Kurva standard fosfat (Uji ATPase)
Konsentrasi K
2HPO
4(mM)
Absorbasi 620 nm
dengan referensi 405 nm
0.0
0.1
0.2
0.4
0.6
0.8
1.0
0.000
0.102
0.239
0.417
0.622
0.834
1.022
Perhitungan konsentrasi enzim RNA helikase HCV saat uji ATPase hasil
pemurnian menggunakan kromatografi gel filtrasi :
y
= 1.0207x + 0.0103
0.873 = 1.0207x + 0.0103
x
=
0207
.
1
0103
.
0
873
.
0
−
= 0.845 mM
y = 1.0207x + 0.0103
R
2= 0.9989
21
Nama Sampel
Blanko
Enzim (10x)
Masa inkubasi hari ke-1
Masa inkubasi hari ke-2
Masa inkubasi hari ke-3
Masa inkubasi hari ke-4
Masa inkubasi hari ke-5
Masa inkubasi hari ke-6
Masa inkubasi hari ke-7
Masa inkubusi hari ke-8
Masa inkubasi hari ke-9
Masa inkubasi hari ke-10
Masa inkubasi hari ke-11
Pengendapan amoniun sulfat
90%
Persentase (%)2.578389
32.42953
65.20465
63.18445
81.41943
78.52204
85.85856
79.77137
78.5752
75.99678
58.16053
103.6151
(Abs
enzim-Abs
enzimdginhibitor
)/Abs
enzim0.025784
0.324295
0.652047
0.631844
0.814194
0.78522
0.858586
0.797714
0.785752
0.759968
0.581605
1.036151
Abs -
blanko
1.254
1.221667
0.847334
0.436334
0.461667
0.233
0.269334
0.177334
0.253667
0.268667
0.301
0.524667
-0.04533
Selisih
λ
620-λ
4050.598333
1.852333
1.82
1.445667
1.034667
1.06
0.831333
0.867667
0.775667
0.852
0.867
0.899333
1.123
0.553
Rata-rata
1.027333
2.883333
2.782
2.230667
1.740333
1.797333
1.578
1.605667
1.450333
1.536667
1.535667
1.591333
1.935
1.332667
Ulangan
3
1.004
2.898
2.74
2.049
1.677
1.658
1.534
1.558
1.412
1.623
1.542
1.508
1.864
1.26
2
0.997
2.898
2.787
2.24
1.765
1.784
1.656
1.773
1.415
1.487
1.484
1.619
1.934
1.345
1
1.081
2.854
2.819
2.403
1.779
1.95
1.544
1.486
1.524
1.5
1.581
1.647
2.007
1.393
22
Lampiran 7 Nilai inihibisi RNA helikase HCV dari optimasi pengendapan amonium sulfat
Nama Sampel
Blanko
Enzim (10x)
Pengendapan 60%
Pengendapan 70%
Pengendapan 80%
Pengendapan 90%
Persentase (%)67.67121
51.7808
83.60075
89.23676
(Abs
enzim-Abs
enzimdginhibitor