Efek Antiproliferatif Pentagamavunon-0 terhadap Sel Kanker Payudara T47D ) Antiproliferative Effect of Pentagamavunon-0 on Breast Cancer Cell Line T47D
Bebas
10
0
0
Teks penuh
Gambar
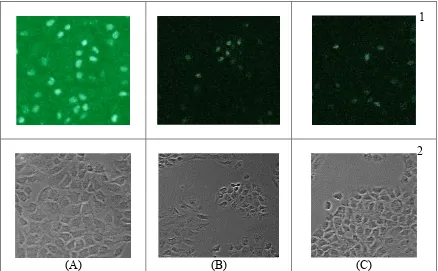
Dokumen terkait
Hal ini memperlihatkan bahwa sifat antiproliferasi ekstrak buah mahkota dewa terhadap sel THP- 1 lebih kuat daripada terhadap sel HeLa, diduga senyawa kimia yang terkandung
untuk mengetahui efek ekstrak daun dewandaru terhadap sel kanker payudara T47D dan MCF-7 yang dilakukan secara in vitro dengan metode MTT assay..
