DINDING SEL EUKARIOTIK
(FUNGI DAN TUMBUHAN)
Makalah
Disusun untuk Memenuhi Tugas Mata Kuliah Metabolisme Sel Dosen Pengampu : Prof. Dr. Priyantini Widyaningrum. M. Si
Oleh
Ristiana Nugrahani 0402514041
Kelas Khusus IPA (Konsentrasi Biologi)
PROGRAM STUDI PENDIDIKAN IPA KONSENTRASI
BIOLOGI PROGRAM PASCASARJANA
UNIVERSITAS NEGERI SEMARANG
2015
BAB I PENDAHULUAN
A. Latar Belakang
Sel terdiri dari organel-organel sel yang masing-masing memiliki tugas khusus. Salah satu organel sel yang memiliki tugas yang tak kalah penting tersebut adalah Dinding sel. Dinding sel merupakan sebuah membran yang terbentuk pada bagian luar dari membran sel yang berperan sangat penting dalam membentuk struktur sel yang kaku, memberi kekuatan dan perlindungan kepada sel terhadap tekanan mekanik.
Dinding sel adalah struktur di luar membran plasma yang membatasi ruang bagi sel untuk membesar. Dinding sel merupakan ciri khas yang dimiliki tumbuhan, bakteri, fungi (jamur), dan alga, meskipun struktur penyusun dan kelengkapannya berbeda. Hewan dan protista kebanyakan tidak memiliki dinding sel.
Pada dinding sel ada bagian yang tidak menebal, yaitu bagian yang disebut noktah. Melalui noktah ini terjadi hubungan antara sitoplasma satu dengan yang lain yang disebut plasmodesmata. Plasmodesmata berupa juluran plasma, yang berfungsi menjadi pintu keluar masuknya zat.
Sesuai dengan arti dari Metabolisme Sel itu sendiri, bahwa kajian darimatakuliah ini adalah mengenai aktivitas metabolisme yang terjadi dalam sel. Pembahasan tersebut berkenaan dengan struktur, komponen penyusun dari organel tersebut, fungsi, dan mekanisme kerja, khusunya pada Dinding sel, maka makalah ini disusun dengan tujuan meningkatkan pemahanan mahasiswa tentang Metabolisme yang terjadi di Dinding Sel.
B. Rumusan Masalah
Adapun rumusan masalah dalam pembahasan mengenai kompleks golgi ini adalah sebagai berikut:
1. Bagaimana Fungsi Dinding Sel Tumbuhan? 2. Bagaimana Struktur Dinding Sel Tumbuhan?
3. Apa Saja Kkomponen penyusun dinding sel Tumbuhan? 4. Bagaimana Teori Pembentukan Dinding Sel Tumbuhan? 5. Bagaimana Struktur Plasmodesmata?
6. Bagaimana perpindahan material melewati Plasmodesmata? 7. Bagaimana Struktur Dinding Sel Fungi?
8. Apa Saja Komponen Penyusun Dinding Sel Fungi? 9. Apa Fungsi Dinding Sel Fungi?
C. Tujuan Penulisan
Penulisan makalah ini ditujukan untuk:
1. Memahami Fungsi Dinding Sel Tumbuhan? 2. Memahami Struktur Dinding Sel Tumbuhan?
3. Memahami Kkomponen penyusun dinding sel Tumbuhan? 4. Memahami Teori Pembentukan Dinding Sel Tumbuhan? 5. Memahami Struktur Plasmodesmata?
6. Memahami perpindahan material melewati Plasmodesmata? 7. Memahami Struktur Dinding Sel Fungi?
8. Memahami Komponen Penyusun Dinding Sel Fungi? 9. Memahami Fungsi Dinding Sel Fungi?
BAB II PEMBAHASAN
A. DINDING SEL TUMBUHAN
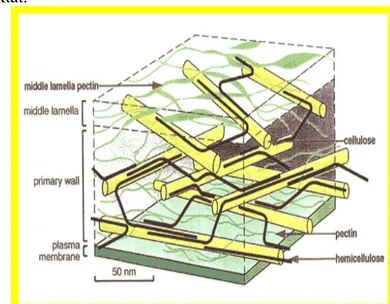
Salah satu ciri yang membedakan antara sel hewan dan sel tumbuhan adalah adanya dinding sel. Seperti dikemukakan sebelumnya bahwa dinding sel terdiri atas dinding primer dan dinding sekunder, diantara dinding primer dari suatu sel dengan dinding primer dari sel tetangganya, terdapat lamella tengah. Lamella tengah merupakan perekat yang mengikat sel-sel secara bersama-sama untuk membentuk jaringan dan oleh sebab itu dijumpai diantara dinding sel-sel primer yang berdekatan.
1. Fungsi Dinding Sel
Dinding sel memiliki berbagai fungsi diantaranya a. Mempertahankan/menentukan bentuk sel.
b. Mendukung dan memberi kekuatan secara mekanik (memungkinkan tanaman untuk tumbuh tinggi, dll).
c. Mencegah membran sel pecah dalam medium hipotonik atau tekanan turgor.
d. Mengontrol laju dan arah pertumbuhan sel serta mengatur isi sel. e. Bertanggung jawab pada struktur bentuk tumbuhan dan mengontrol
morfogenesis tanaman. f. Memiliki peran metabolisme.
g. Memberi perlindungan fisik dari serangan serangga dan pathogen. h. Penyimpanan karbohidrat yang merupakan salah satu komponen
dinding selyang dapat digunakan kembali dalam proses metabolisme lainnya. Jadi, di satu sisi dinding sel berfungsi sebagai gudang penyimpanan untuk karbohidrat.
i. Sebagai produk ekonomis, dimana dinding sel sangat penting untuk menghasilkan produk-produk seperti kertas, serat, serat kayu,sumber energy, papan dan bahkan sebagai bahan dasar dalam produk makanan diet kita.
j. Tabung untuk trasportasi jarak jauh.
k. Kegiatan fisiologis dan biokimia pada dinding sel berperan untuk komunikasi antar sel-sel.
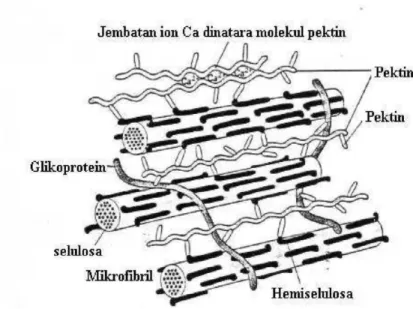
Komponen utama dinding sel tumbuhan adalah polisakarida yang terdiri atas tiga tipe utama yaitu Selulosa, Hemiselulosa, Polisakarida pektat.
Gambar 1. Struktur Dinding Sel Tumbuhan
a. Selulosa
Selulosa pada kayu kurang lebih 45% dari berat keringnya, sedangkan pada kapas kurang lebih 98%. Selulosa adalah polimer lurus yang terdiri atas unit-unit glukosa yang membentuk rantai yang saling berhubungan melalui ikatan glikosida (3 1-4. Satu molekul selulosa terdiri atas 8.000-15.000 unit glikosa. Dalam satu mikrofibril, setiap rantai glukosa membentuk ikatan hidrogen dengan rantai glukosa yang ada didekatnya sehingga secara struktural mikrofibril menjadi lebih stabil.
CH2OH CH2OH CH2OH CH2OH CH2OH
b. Hemiselulosa
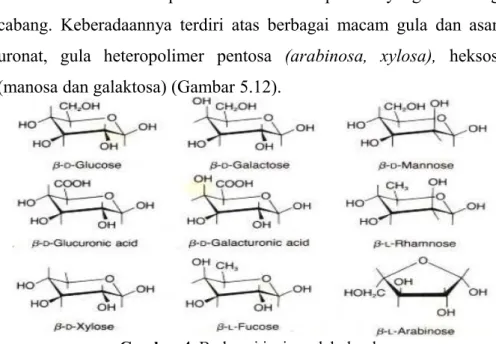
Hemiselulosa merupakan molekul heteropolimer yang bercabang-cabang. Keberadaannya terdiri atas berbagai macam gula dan asam uronat, gula heteropolimer pentosa (arabinosa, xylosa), heksosa (manosa dan galaktosa) (Gambar 5.12).
Gambar 4. Berbagai jenis molekul gula
Berbagai jenis hemiselulosa mempunyai dua ciri struktur yang sama, yaitu :
1) Satu tulang punggung yang terdiri atas ikatan b 1-4 2) Rantai samping yang pendek, misalnya glukomannan.
Gambar 3. Ikatan hidrogen yang menghubungkan molekul selulosa dengan molekul selulosa yang lain (Thorpe, 1984)
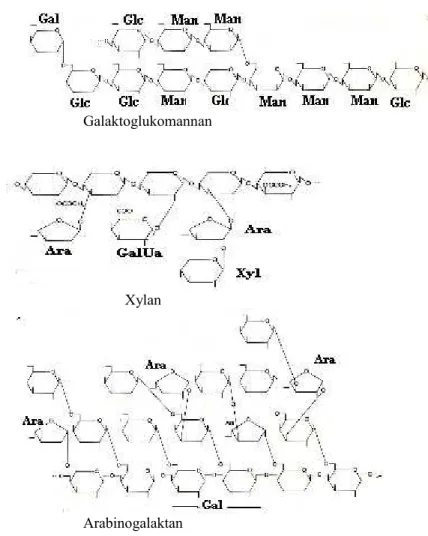
Beberapa hemiselulosa diberi nama xylan, arabinok xylan, glukomannan, galaktomannan, dan xyloglukan. Dari nama tersebut, mencerminkan strukturnya (Gambar 4.15).
Galaktoglukomannan
Xylan
Arabinogalaktan
Gambar 5. Struktur berbagai molekul hemiselulosa (Thorpe, 1984)
c. Pektin
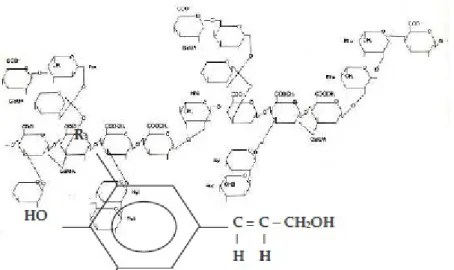
Pektin adalah suatu famili dari polisakarida dan memiliki struktur yang sangat bervariasi. Satu ciri utama yang dimilikinya adalah adanya gugus asam yang disebabkan oleh adanya residu asam glukoronat dan galaktoronat. Hemiselulosa berfungsi melapisi mikrofibril sekaligus sebagai perekat. Pektin mempunyai struktur yang sangat bervariasi dengan ciri utama adanya gugus asam yang merupakan residu asam glukoronat dan galaktoronat (Gambar 6).
Gambar 6. Struktur Pektin, Thorpe 1984.
d. Protein Non Struktural
Dinding sel tumbuhan juga mengandung komponen-komponen non polisakarida, yaitu berupa protein-protein struktural yang kaya dengan hidroksi prolin yaitu sekitar 25%. Diduga bahwa, fungsi dari protein tersebut adalah dalam pengorganisasian dinding sel.
e. Lignin dan Kutin
Selain itu, terdapat plastik biologi, yaitu lignin dan kutin. Lignin biasanya mengisi dinding sekunder dan menyebabkan dinding menjadi kaku. Lignin dibentuk dari hasil polimerisasi prekuersor lignin. Lignin berfungsi sebagai bahan pengisi dinding sel. Ada tiga tipe prekuersor lignin, yaitu (i) Coumaril alkohol (R1=H dan R2=H); (ii) Cineferil alkohol =H dan R2=OCHs); dan (iii) Synapyl alkohol (R1=OCH3 dan R2=OCH3).
Kutin biasanya terdapat pada permukaan dinding sel dan berfungsi agar permukaan sel resisten terhadap dehidrasi dan juga sebagai proteksi sel terhadap luka. Struktur kitin belum jelas, namun ia mengandung asam lemak hidroksi (C16-C18) yang terikat secara kovalen satu dengan yang lain melalui ikatan ester.
Dinding primer tersusun atas selulosa, yaitu suatu polimer glukosa dengan ikatan p 1-4. Kurang lebih 8.00015.000 gugus p-glukosa secara bersama-sama membentuk satu rantai selulosa. Kurang lebih 40-70 tantai molekul selulosa terdapat dalam kelompok-kelompok yang sejajar membentuk mikrofibril. Mikrofibril-mikrofibril saling berkelompok membentuk Mikrofibril-mikrofibril dengan diameter ± 0,5 ¡1 dan tampak dengan mikroskop cahaya (Thorpe, 1984). Di dalam dinding sel, mikrofibril dilapisi oleh hemiselulosa yang selanjutnya dihubungkan ke hemiselulosa lain oleh pektin dan polisakarida lain (Albert et al., 1983).
Pada dinding primer, mikrofibril-mikrofibril tersusun erat dan letaknya tersebar (Gambar 4.20). Ruang-ruang di antara mikrofibril diisi oleh air, protein dan bahan dinding sel lain, yaitu hemiselulosa dan pektin. Mikrofibril-mikrofibril tersebut bersifat lentur dan dapat memanjang. Kandungan hemiselulosa tinggi dan selulosa rendah. Dinding primer adalah struktur yang pertama dibentuk dan diletakkan pada lamella tengah.
Dinding sekunder terutama terdiri atas selulosa. Pada beberapa sel terdapat pektin. Lapisan terluar dinding sel sekunder terdapat lignin,
Gambar 7. Ikatan Antara Mikrofibril pada Dinding Sel (Albert et al, 1983)
kutin, dan suberin. Lignin terdiri atas tiga macam yaitu koniferin alkohol, senafil alkohol, dan kumoril alkohol. Kutin merupakan rentai karbohidrat dengan jumlah atom C antara 21-35. Lilin adalah ester asam lemak dengan alkohol.
(a) (b)
Gambar 8. Dinding sel primer (a), Dinding sel sekunder (b)
Pada dinding primer, mikrofibril tersebar dalam suatu matriks , bersifat lentur, dan memanjang bersama-sama dengan pemanjangan protoplasma, kadar hemiselulosa tinggi dan hemiselulosa relatif rendah. Dinding primer merupakan struktur yang pertama kali diletakkan pada lamella tengah. Pada dinding sekunder, mikrofibrilnya tersusun sejajar, kaku dan tidak dapat memanjang, kadar hemiselulosa relatif rendah dan selulosanya lebih banyak. Dinding sekunder dibentuk setelah sel mencapai ukuran yang maksimum. Dinding sekunder merupakan suatu struktur multilamella yang terdiri atas tiga lapisan yang disebut S1, S2, dan S3. Mikrofibril pada lapisan ini terle-tak sejajar tetapi menurut arah yang berbeda pada lapisan yang berbeda.
3. Pertumbuhan Dinding Sel
Pada pertumbuhan dinding sel, ada dua proses yang terlibat, yaitu pembelahan sel dan pemanjangan sel. Pembelahan sel berlangsung pada jaringan meristematis. Sel-sel anak yang dihasilkan pada jaringan meristematis mempunyai ukuran yang lebih kecil dari pada sel-sel dewasa. Setelah sel anak terbentuk, maka selanjutnya ter-jadi pemanjangan sel. Ada
dua teori yang berkenaan dengan pemanjangan dinding sel, yaitu teori multinet, dan teori orientasi aktif.
a. Teori Multinet
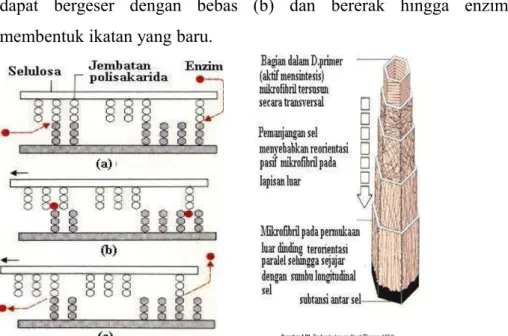
Menurut teori multinet , mikrofibril diletakkan pada permukaan bagian dalam dinding sel menurut arah melintang terhadap panjang sel. Pada waktu dinding sel memanjang, mikrofibril-mikrofibril mengalami reorientasi ulang ke arah sumbu longitudinal sel hingga mikrofibril sejajar dengan sumbu. Dengan demikian orientasi mikrofibril menurut teori multinet berlangsung secara pasif mengikuti perentangan dinding sel selama berlangsungnya pertumbuhan (gambar 9).
Selama pemanjangan dinding sel, mikrofibril bergerak satu terhadap yang lain. Pada gambar 9 ditunjukkan mekanisme pemanjangan dinding sel. Dalam hal ini terdapat enzim-enzim yang memutuskan ikatan antara dua polisakarida dinding sel (a) dan tetap melekat pada salah satu titik pemotongan, kemudian polisakarida dapat bergeser dengan bebas (b) dan bererak hingga enzim membentuk ikatan yang baru.
Gambar 9. Mekanisme Pemanjangan Dinding Sel (Thorpe, 1984)
b. Teori Orientasi Aktif
Menurut teori orientasi aktif, mengemukakan bahwa terbentuknya lapisan mikrofibril yang sejajar pada dinding sel tumbuhan yang tidak
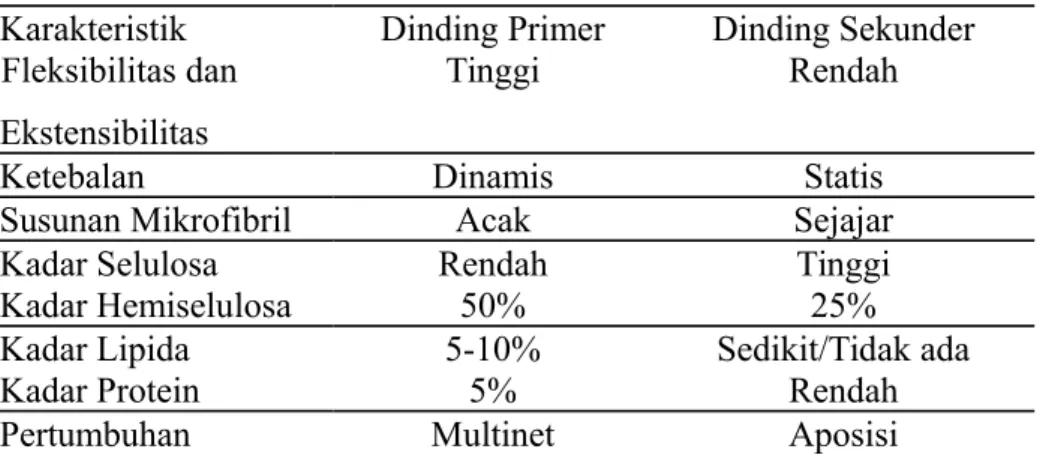
tumbuh lagi berlangsung secara siklosis, (mengalirnya bahan-bahan sitoplasma di dalam sel tumbuhan) pada bagian dalam sel. Aliran siklosis ini orientasi mikrofibril pada bagian luar sel. Pada tabel 1, ditunjukkan perbedaan dinding primer dengan dinding sekunder.
Tabel 1. Perbedaan antara dinding primer dengan dinding sekunder. Karakteristik Dinding Primer Dinding Sekunder Fleksibilitas dan
Ekstensibilitas
Tinggi Rendah
Ketebalan Dinamis Statis
Susunan Mikrofibril Acak Sejajar
Kadar Selulosa Rendah Tinggi
Kadar Hemiselulosa 50% 25%
Kadar Lipida 5-10% Sedikit/Tidak ada
Kadar Protein 5% Rendah
Pertumbuhan Multinet Aposisi
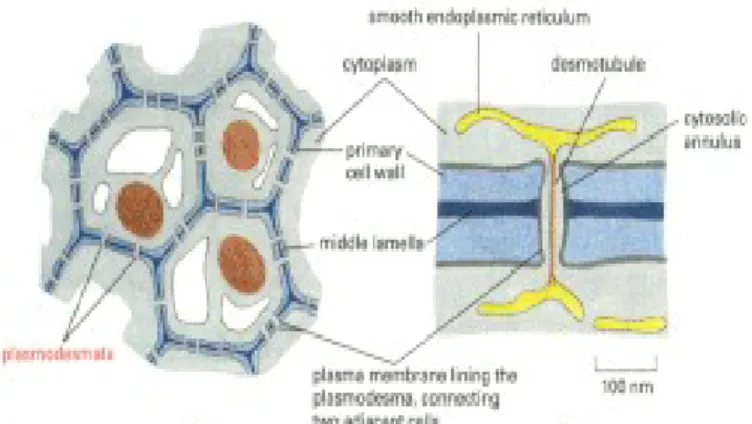
4. Plasodesmata
Plasmodesma (desmos:mengikat,jamak: plasmodesmata) merupakan saluran penghubung antar sel tumbuhan atau dapat dikatakan sebagai media komunikasi pada sel tumbuhan. Plasmodesmata terletak di antara dua dinding sel tumbuhan dan berhubungan langsung dengan kedua RE halus masing-masing sel. Transportasi antar melalui plasmodesmata mengontrol keberlangsungan sel tanaman, dan merupakan hal penting mendasar dalam gerakan virus,ketahanan penyakit, dan penyebaran sinyal RNAi. Meskipun plasmodesmata tampak unik untuk sel tanaman, mereka mungkin memiliki kesamaan struktural dan fungsional dengan tunneling nanotube yang baru ditemukan yang menghubungkan sel-sel hewan. Baru-baru ini, protein yang membatasi plasmodesmata telah diidentifikasi, dan protein mikrotubulus terkait ditemukan negatif mengatur pengangkutan pergerakan protein virus. Kemajuan lain telah memberikan wawasan baru ke dalam fungsi dan sifat molekul plasmodesmata dan telah menunjukkan bahwa lalu lintas protein melalui plasmodesmata telah diatur, membuka kemungkinan bahwa kontrol genetik dari fungsi plasmodesmal akan segera dipahami.
melalui PD. Protein endogen pertama ditemukan untuk melintasi sel-sel melalui PD adalah jagung protein homeodomain perkembangan KNOTTED1 (KN1). Segera setelah itu, floem protein ditemukan untuk meningkatkan PD SEL dan lalu lintas sel-sel. Signifikansi fungsional dari berbagai protein seluler floem masih dijelaskan.
Virus tanaman juga mengambil keuntungan dari PD untuk menyebarkan genom mereka dari sel ke sel. Protein bergerak (MPs) adalah protein virus khusus yang meningkatkan SEL dan mengizinkan transport genom virus. MPs berhubungan dengan sitoskeleton dan ER. Penemuan faktor endogen tanaman yang terlibat dalam fungsi MP berakibat lebih meningkatkan pemahaman kita tentang molekul-molekul yang sulit dipahami.
Dalam ulasan ini, kita akan membahas kemajuan terbaru dalam pemahaman kita tentang mekanisme transportasi melalui PD, bagaimana virus menggunakan mesin PD untuk lalu lintas sel-sel, dan penemuan pemain molekul menarik dalam fungsi PD. Kami juga akan membahas percobaan pilihan terbaru yang menunjukkan peran besar PD dalam perkembangan tumbuhan.
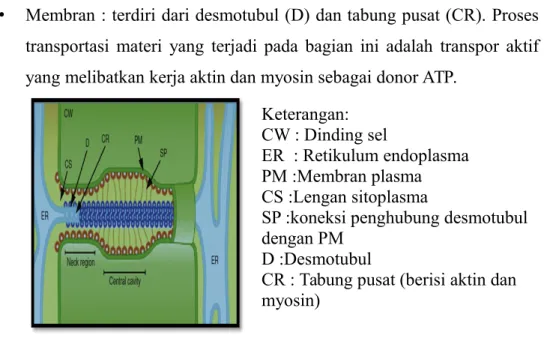
Gambar 10. Plasmodesmata
a. Struktur Plasmodesmata
Plasmodesmata terdiri dari 2 bagian, yaitu:
• Pori: terdiri dari lengan sitoplasma (CS) dan koneksi penghubung (SP).Materi yang melewati pori adalah materi yang ditransfer ke sel lain tanpa membutuhkan energi karena mengikuti gradien potensial (transpor pasif)
• Membran : terdiri dari desmotubul (D) dan tabung pusat (CR). Proses transportasi materi yang terjadi pada bagian ini adalah transpor aktif yang melibatkan kerja aktin dan myosin sebagai donor ATP.
Gambar 11. Struktur anatomi plasmodesmata
b. Mekanisme Zat Melintasi Plasmodesmata
1) Transportasi Pasif
PD mengizinkan transportasi pasif makromolekul jika mereka tersedia secara bebas dalam sitoplasma dan jika ukuran di bawah SEL tersebut. Percobaan yang melibatkan baik pemuatan atau mikroinjeksi pemeriksaan flourecent menunjukkan bahwa benda kecil berwarna (~ 1 kDa) bisa bergerak dengan difusi (Ulasan di [24]). Dalam beberapa kasus, SEL dapat lebih besar dari 1 kDa; misalnya protein fluorescent hijau (GFP) dapat melintasi bebas di jaringan seperti kelopak, ujung akar dan daun roset muda [25]. Meskipun transportasi pasif melalui PD mungkin menjadi hasil dari difusi, mungkin selektif seperti halnya dengan gap junction hewan [26].
2) Transpor aktif
Pergerakan MPs virus dan protein endogen seperti KN1 mungkin terjadi melalui mekanisme yang ditargetkan (Ulasan di [27]). Hipotesis mekanisme aktif, gerakan yang ditargetkan melalui PD masih dalam penyelidikan. Termasuk mekanisme PD-reseptor-mekanisme dimediasi dimana protein non-sel-otonom (setiap protein yang melintasi sel-sel melalui PD) memiliki sinyal target PD , dan mekanisme klasik exo- dan endo-cytosis [27] . Kami mengusulkan
Keterangan: CW : Dinding sel
ER : Retikulum endoplasma PM :Membran plasma CS :Lengan sitoplasma
SP :koneksi penghubung desmotubul dengan PM
D :Desmotubul
CR : Tabung pusat (berisi aktin dan myosin)
hipotesis baru untuk transportasi melalui PD dengan analogi modus baru transportasi sel ke sel yang baru-baru ini ditemukan pada hewan (yang akan dibahas kemudian dalam bagian ini). Hipotesis mekanisme ini berbeda, tapi mungkin tidak saling terpisah.
3) Exo-endositosis
Baluska et al. [28] menggunakan pelabelan berwarna fluorescent untuk menunjukkan bahwa daur cairan endositosis terjadi di PD dari apeks akar jagung. Kompartemen Tubulo-vesikular menonjol ke darah dalam dari membran plasma di daerah biji yang diperkaya aktin dan meosin pada maisng-masing PD. Pembentukan kompartemen ini diblokir oleh latrunculin B, menunjukkan mekanisme bergantung-aktin, sedangkan gangguan mikrotubulus tidak berpengaruh. Kehadiran vesikel endocytic di PD menunjukkan bahwa PD berperan dalam lalu lintas endo-membran atau internalilasi vesikel, mungkin di lokasi pemberhentian vesikel, dan fusi membran. Dinding sel (ECM) jauh lebih tipis di dearah dimana PD terkumpul, yang mungkin karena kebetulan menjadi daerah yang baik untuk exo- dan endositosis. Jika ditambah dengan eksositosis, mekanisme tersebut bisa mengakibatkan translokasi protein antara sel-sel tanaman, mirip dengan mekanisme yang diusulkan untuk transportasi ENGRAILED homeodomain antara sel hewan [29].
4) Tunneling nanotube
Penemuan jenis baru dari saluran antar sel hewan, yang disebut 'tunneling nanotube' (TNT), dapat memberikan bukti pertama bahwa mode fungsional serupa komunikasi antar ada di tanaman dan hewan. Independen dari endo klasik dan eksositosis, TNTs mengizinkan lalulintas vesikel endomembran antara sel-sel, seperti yang ditunjukkan oleh adanya synaptophysin, penanda untuk endosomes awal dan sinaptik seperti mikrovesicles endosome diturunkan di TNTs. TNTs menyerupai PD dalam beberapa aspek mekanistik. Misalnya, TNTs sensitif terhadap latrunculin B, dan karena itu dimungkinkan menggunakan mekanisme transportasi
F-dependent-aktin, dan transportasi virus melalui PD juga sensitif terhadap inhibitor ini. Paralel lain adalah bahwa sitoskeleton mikrotubulus tampaknya tidak menjadi penting untuk transportasi TNT atau PD. Namun, perbedaan utama adalah bahwa TNTs bersifat sementara dan variabel dalam lokasi, sedangkan di sel tanaman PD dianggap stabil. Salah satu kemungkinan adalah bahwa PD bisa memberikan tempat untuk pembentukan nano-tube antara sel-sel tumbuhan. Penemuan TNTs ini menarik karena menunjukkan hipotesis baru dan hipotesis dapat diuji mengenai lalu lintas makromolekul yang melibatkan vesikel melalui PD.
Meskipun TNTs secara struktural berbeda dari PD ditemukan pada tumbuhan yang lebih tinggi, TNTs lebih mirip dengan struktur yang PD dari Chara corallina, ganggang characean dianggap sebagai spesies transisi antara ganggang dan tanaman yang lebih tinggi [33] (Gambar 1). PD pada Chara setelah sitokinesis dan, seperti TNTs, kurang memiliki desmotubule ER. Sebuah ide menarik adalah bahwa PD primitif di Chara adalah prekursor dari PD tanaman yang lebih tinggi dan juga berbagi beberapa kesamaan fungsionaldengan TNTs. Baru-baru ini diidentifikasi protein PD melokalisir protein mendukung hipotesis dari lalu lintas vesikel PD melalui exo-endositosis atau melalui TNTs.
Faktor transkripsi tanaman dapat bergerak melalui plasmodes-mata oleh salah satu mekanisme di atas, dan ada bukti untuk kedua transportasi non-selektif pasif [37] dan transportasi selektif [10,38 **]. Gagasan bahwa gerakan faktor transkripsi melalui PD terjadi oleh mekanisme reseptor-mediated adalah menarik, karena jenis mekanisme transportasi selektif diamati di pori nuklir. Namun, tidak ada PD menargetkan sinyal faktor transkripsi atau reseptor PD untuk faktor transkripsi telah definitif diidentifikasi. Ada kemungkinan bahwa perdagangan beberapa faktor transkripsi tanaman dapat terjadi setelah pengemasan di Golgi melalui exo-dan endositosis klasik, atau melalui gerakan mikrovesicles sepanjang TNT-seperti struktur di PD.
5) Gerakan Virus melalui Plasmodesmata
Meskipun mikrotubulus pernah dianggap penting bagi lalulintas MP virus, data baru mepermasalahkan hipotesis ini. Dengan menggunakan DNA shuffling, Gillespie et al. menemukan bahwa mutan MP TMV (MPR3) dengan afinitas terbatas mikrotubulus benar-benar menunjukkan peningkatan lalulintasnya , dan mereka menunjukkan bahwa mutan ini peran mikrotubulus dalam degradasi MP, tidak hanya menargetkan PD. Temuan konsisten digambarkan oleh Kragler et al., Yang menggunakan layar dua-hybrid sitoplasma untuk mengidentifikasi MPB2C, protein mikrotubulus terkait yang berinteraksi dengan TMV MP. Studi elegan rekan lokal MPB2C, mikrotubulus dan MPs in vivo, dan tes fungsional menunjukkan bahwa co-ekspresi MPB2C sebenarnya menghambat sel-sel untuk gerakan TMV MP, sekali lagi menunjukkan hubungan negatif antara lokalisasi MP untuk mikrotubulus dan gerakan dari sel ke sel. Selanjutnya, partikel utuh replikasi TMV virus melintasi sel-sel tanpa keterlibatan mikrotubulus.
Komponen molekul dari plasmodesmata Komponen molekul PD dan jalur perdagangan yang terkait sampai saat ini, telah sulit dipahami. Sangat mungkin bahwa sitoskeleton aktin memainkan peran yang berbeda, termasuk mengatur SEL, lalulintas muatan dari dan ke PD, dan daur ulang muatan. Centrin, protein sitoskeleton terikat kalsium, dan calreticulin, sebuah protein mengisolir kalsium, juga melokalisasi ke PD dan mungkin memodulasi fungsi mereka dalam menanggapi kalsium sinyal. Memang, sinyal kalsium cepat dan secara sementara mengatur permeabilitas PD, meskipun mekanisme yang mereka gunakan untuk mempengaruhi ukuran pori plasmodesmal belum ditemukan. Sebuah mekanisme potensial berasal dari penelitian terhadap elemen saringan, sel-sel konduktif dari floem, yang reversibel terpasang oleh protein kontraktil kalsium-sensitif yang bertindak sebagai 'stopcocks seluler'.
B. DINDING SEL FUNGI
Fungi merupakan organisme yang tidak mempunyai klorofil dan bereproduksi dengan spora (Carris dan Lori, 2009). Fungi bersifat khemoorganotrof dan memperoleh nutrisinya secara absorpsi dengan bantuan enzim ekstraseluler untuk memecah biomolekul kompleks seperti karbohidrat, protein, dan lemak menjadi monomernya yang akan diasimilasi menjadi sumber karbon dan energi (Madigan et al., 2012). Bahan makanan ini akan diurai dengan bantuan enzim yang diproduksi oleh hifa menjadi senyawa yang dapat diserap dan digunakan untuk tumbuh dan berkembang (Sinaga, 2000). Penyerapan makanan dilakukan oleh hifa yang terdapat pada permukaan tubuh fungi (Lockwood, 2011). Dinding sel fungi berbeda dengan tumbuhan yang memiliki komponen utama selulosa, kitin dan glukan memiliki peran penting sebagai komponen penyusun dinding sel fungi.
Gambar 12. Sruktur Dinding Sel Fungi
1. Komponen Dinding Sel Fungi
Sebagian besar dinding sel fungi mengandung khitin dan glukan. Kitin yang merupakan polimer glukosa derivatif dari N-acetylglucosamine. Khitin tersusun pada dinding sel dalam bentuk ikatan mikrofibrillar yang dapat memperkuat dan mempertebal dinding sel. Beberapa polisakarida lainnya, seperti manann, galaktosan, maupun selulosa dapat menggantikan khitin pada dinding sel fungi. Komponen lain penyusun dinding sel fungi yaitu glukan yang strukturnya mirip selulosa pada dinding sel tumbuhan tingkat tinggi. Beta-glukan merupakan polimer D-glukosa dengan ikatan
Gambar 13. Struktur Kimia Kitin dan Glukan
Selain kitin dan glukan, penyusun dinding sel fungi juga terdiri dari 80-90% polisakarida, protein, lemak, polifosfat, dan ion anorganik yang dapat mempererat ikatan antar matriks pada dinding sel (Madigan et al., 2012). Perbandingan komposisi dinding sel pada tiap-tiap divisi fungi dijelaskan dalam Gambar 14.
Gambar 14. Tabel perbandingan komposisi dinding sel Fungi
2. Fungsi Dinding Sel Fungi
Dinding sel fungi berfungsi untuk melindungi protoplasma dan organel-organel dari lingkungan eksternal. Struktur dinding sel tersebut dapat memberikan bentuk, kekuatan seluler dan sifat interaktif membran plasma. Selain khitin, dinding sel fungi juga tersusun oleh fosfolipid bilayer yang mengandung protein globular. Lapisan tersebut berfungsi sebagai tempat masuknya nutrisi, tempat keluarnya senyawa metabolit
sel, dan sebagai penghalang selektif pada proses translokasi. Komponen lain yang menyusun dinding sel fungi adalah antigenik glikoprotein dan aglutinan, senyawa melanins berwarna coklat berfungsi sebagai pigmen hitam. Pigmen tersebut bersifat resisten terhadap enzim lisis, memberikan kekuatan mekanik dan melindungi sel dari sinar UV, radiasi matahari dan pengeringan) (Kavanagh, 2011).
BAB III PENUTUP Kesimpulan
Dinding sel sebagai bagian terluar dari sel memiliki peran dan fungsi khusus terutama berkaitan dengan transportasi zat masuk dan keluar sel, memberikan bentuk pada sel, melindungi keseluruhan bagian dalam sel. Dinding sel Tumbuhan memiliki komponen penyusun yang utama berupa selulosa, hemiselulosa, lignin dan kutin, serta protein non struktural lainnya. Transportasi antar sel tumbuhan sangat di bantu dengan adanya plasmodesmata. Dinding sel jamur memiliki komponen utama kitin dan glukan yang keduanya merupakan polimer dari β-1,3-D-Glyco piranosil yang terikat pada unit β-1,6-β-1,3-D-Glyco piranosil dan β-1,4-N-Acetil-glucosamin.
DAFTAR PUSTAKA
Adnan. 2008. Dinding Sel. Available online at: https://www.scribd.com/doc/20535810/DINDING-SEL-Adnan-UNM. Universitas Negeri Makassar.
Campbell,dkk.2008.Biologi Jilid 1. Erlangga: Jakarta
Cilia, Michelle Lynn and David Jackson. 2004. Plasmodesmata Form and Function. International Journal of Science Direct. Available online at http://jacksonlab.cshl.edu/publications/Cilia2004.pdf. Elsevier Ltd. All rights reserved.