Summary Both drought and root pruning (RP) increased the number of cones induced when black spruce (Picea mariana (Mill.) B.S.P.) grafts were injected with gibberellins A4/7 (GA), but their effects on predawn shoot water potential and current-year needle development differed. Drought decreased predawn shoot water potential (Ψpd), but only during the period when irrigation was withheld, and it had no effect on the growth or gas exchange properties of current-year needles. Conversely, root pruning had little effect on Ψpd, but it resulted in trees with smaller current-year needles that had lower nitrogen and chlo-rophyll concentrations and reduced rates of gas exchange up to the later stages of shoot elongation compared with needles of control trees. These findings are discussed in relation to poten-tial effects on the development of induced cones in the follow-ing growth cycle.
Keywords: drought, GA4/ 7, photosynthesis, pollen-cone
pro-duction, root pruning, seed-cone propro-duction, shoot water po-tential, stomatal conductance.
Introduction
Applying gibberellins A4 and A7 (GA), often in combination with root pruning, drought, heat, and stem or branch girdling, is commonly done to induce cone production in the Pinaceae to expedite breeding (Ho 1988, 1991a, Greenwood et al. 1993), and for operational seed production (see reviews in Bonnet-Masimbert 1987, Pharis et al. 1987, Philipson 1990). The success of miniaturized or containerized orchards (Ross et al. 1986, Ross 1989, Smith and Powell 1992, Sweet et al. 1992) depends on a regular supply of high quality seed from GA-in-duced cones.
When properly formulated and applied, such induction treatments are generally effective (Ross and Pharis 1981, Ross and Bower 1989). However, needle loss following GA and other induction treatments may occur (Ho 1991b, Ross 1991a, Greenwood et al. 1993). Furthermore, although the GA-in-duced cones usually develop normally (Ross et al. 1980, Bon-net-Masimbert 1987, BonBon-net-Masimbert and Zaerr 1987), they are sometimes smaller than noninduced cones and have lower seed yields (Puritch et al. 1979, Ross et al. 1980, Philipson 1987, Ross 1988, Philipson et al. 1990). The cause(s) of the
smaller cones has not been determined; however, a negative correlation between number of cones produced and seed yield per cone has been reported most often when trees produce large cone crops.
Developing cones act as strong sinks for photosynthate and nutrients (Dickmann and Kozlowski 1968, 1970). Competition for nutrients may be a factor leading to the abortion of polli-nated ovules and premature drop of conelets in pine (see review by Owens 1991), but results for black spruce (Picea mariana (Mill.) B.S.P.) are lacking. Similarly, the effects of cone-induction treatments on needle development and gas exchange and, concomitantly, the availability of photosynthate to developing cones in black spruce, have not been quantified. The objectives of this study were to: (1) quantify the effects of injecting GA4/7 in combination with drought and root-prun-ing treatments on current-year needle morphology and nitro-gen content, photosynthetic capacity, and gas-exchange properties; and (2) determine whether these effects are caus-ally related to successful cone induction.
Material and methods
Tree culture
Seven-year-old grafts from seven black spruce clones were grown in 25-liter containers by J.D. Irving, Ltd. in their breed-ing hall in Sussex, N.B., Canada, then shipped to Fredericton in January 1993. The phenological stages of developing cur-rent-year shoots were used to time the application of treat-ments. These stages were vegetative bud burst in the upper one-third of the crown (VBB); early slow growth phase of shoot elongation (ESE); rapid or exponential shoot elongation phase (RSE); late slow growth phase of shoot elongation (LSE); cessation of shoot elongation (CSE) (shoots visibly lignified for approximately one-third to one-half their length and all but a few needles at the apex reflexed and almost perpendicular to the shoot axis); and completion of shoot lignification (CSL) (Ho 1991a, Smith and Greenwood 1995). The length of one lateral shoot on a 3-year-old branch was measured twice weekly throughout the study periods in both 1993 and 1994.
Effects of cone-induction treatments on black spruce (
Picea mariana
)
current-year needle development and gas exchange properties
RONALD F. SMITH
1and MICHAEL S. GREENWOOD
2 1Canadian Forest Service--Atlantic Forestry Centre, P.O. Box 4000, Fredericton, N.B. E3B 5P7, Canada 2 Department of Forest Ecosystem Science, University of Maine, Orono, ME 04469, USA
Received December 20, 1995
In 1993, trees received 3.6 l of water and fertilizer daily using a drip irrigation system. The trees received starter N,P,K (11,41,8) at 50 ppm nitrogen (N) up to VBB, grower N,P,K (20,8,20) at 100 ppm N during ESE and then at 200 ppm N until CSL, and hardener N,P,K (8,20,30) at 35 ppm N, thereaf-ter. In 1994, two changes were made: (1) trees were watered every second day, and (2) N,P,K (20,8,20) was applied at 200 ppm N during ESE. Trees were grown in a greenhouse in a 16-h photoperiod provided by Sylvania Lumalux GTE high pressure sodium vapor lamps. The means (range) of tempera-tures were 22 (18--25) °C day and 18 (17--22) °C night up to LSE and 30 (28--33) °C day and 20 (19--22) °C night, thereaf-ter.
Treatments
In 1993, four ramets from each of seven clones received one of four treatments: (1) control (C), (2) drought for 7 days from March 19 to 26, 1993 (D), (3) stem injection of 10 mg GA4/7 (Zeneca Agro, Stoney Creek, Ontario) in 0.1 ml of 95% ethyl alcohol on March 23, 1993 (GA), and (4) GA + D. The drought was applied during the early slow growth phase of shoot elongation (lateral shoots in the upper one third of the crown were between 1 and 2 cm in length, except for Clone 3, in which shoots averaged 4 to 5 cm in length). Treated grafts were grown from March to May, hardened-off, and placed outside in a shade area on June 15. All trees were moved to a green-house to thaw at 5 °C on January 19, 1994. After the plants had thawed, the greenhouse day/night temperature was increased to 15/10 °C. Conelet receptivity was completed by February 20, 1994.
On February 28, 1994 (before vegetative bud burst), two ramets from each clone, the GA + D and D 1993 treatments, were root pruned (RP) by making three vertical cuts plus removing approximately 10 cm of roots from the bottom of the rootball. On March 15, 1994, cones that had been induced in 1993 were counted and the root-pruned trees were given a stem injection of 10 mg GA4/7. Ramets in the C and GA 1993 treatments were retained as controls in 1994. On June 29, 1994, after shoots had lignified, trees were placed in a cooler at 5 ± 2 °C until September 6 when they were returned to the greenhouse, flushed, and the cones induced in 1994 counted.
Needle morphology and gas exchange
Morphology and rates of gas exchange of current-year needles were assessed for second-order lateral shoots from 3-year-old branches from shortly after vegetative bud burst to the time when elongating shoots became lignified. Gas exchange was measured between 0800 and 1200 h with an LI-6200 portable photosynthesis system (Li-Cor Inc., Lincoln, NE) equipped with a 250-ml cuvette and calibrated before each set of meas-urements against a certified CO2 source (500 ppm). Before commencing measurements, shoots were inserted in the cu-vette and acclimated for 30 to 45 s to a Sylvania 90-W capsylite halogen light providing a mean (± SE) photosynthetically active radiation (PAR) of 1092 ± 40 and 1079 ± 20 µmol m−2 s−1, for 1993 and 1994, respectively. Ambient CO2 was
467 ± 3 and 412 ± 3 ppm in 1993 and 1994, respectively. Two shoots per ramet were measured on each sampling date. Meas-urements were made successively on shoots in comparable positions to ensure that starting CO2 concentration, chamber temperature, relative humidity, and light did not change sig-nificantly between measurements. Except for respiring shoots that had elongated less than 2 cm, PAR, relative humidity, and temperature within the cuvette did not vary by more than 20% between successive measurements for a given ramet and were not significantly different at α = 0.05 in paired t-tests (Snedecor and Cochrane 1980). Net photosynthesis (Pn) and stomatal conductance (gs) were expressed per unit of needle surface area as µmol m−2 s−1 and mol m−2 s−1, respectively. Total shoot length, numbers of needles, and their fresh and dry (48 h at 65 °C) weights were determined for one of the shoots per ramet used for gas-exchange measurements. One-sided projected surface area (SA) of a random 50-needle sample was measured with a Delta-T area meter (Delta-T Devices Ltd., Cambridge, England). The leaf dry weight/leaf area ratio (W, gDW mm−2) was determined as described by Oren et al. (1986). Total nitrogen concentration (mg gDW−1) was determined by a modified micro-Kjeldahl method (Technicon Industrial Method No. 334-74W/B+) (Technicon Instruments Corp., Tar-rytown, NY) using a Technicon BD40 acid block digester.
Chlorophyll of needles from the second shoot per ramet used for gas-exchange measurements was extracted by a modi-fied dimethylsulfoxide (DMSO) method (Hiscox and Israel-stam 1979, Greenwood et al. 1989) and absorbance was measured with a Technicon AA II autoanalyzer equipped with wavelength specific filters of 666 and 648 nm. Absorbancy values were converted to concentrations (mg gDW−1) based on the extinction coefficients of chlorophylls a and b from spinach (Sigma Chemical Co., St. Louis, MO). Chlorophyll estimates were checked with a Spectronic 1201 spectrophotometer (Mil-ton Roy Co., Rochester, NY) in 1993 and a Phillips PU 8800 UV/VIS spectrophotometer (Pye Unicam Ltd., Cambridge, England) in 1994. The R2 values were 0.863 (P < 0.001) and 0.926 (P < 0.001) for 1993 and 1994, respectively (Smith 1995).
Water status measurements
Predawn shoot water potential (Ψpd, MPa) was measured with a pressure chamber (PMS Instrument Co., Corvallis, OR). The first reading was taken on 1-year-old shoots because the cur-rent-year shoots had not developed sufficiently for a reliable reading. Subsequent readings were taken on current-year shoots. The volumetric field capacity (FC) of the containers was determined as the total pore space less the drainable pore space (see Whitcomb 1988) measured with a TRASE unit (Soil Moisture Corp., Santa Barbara, CA).
Data analyses
Results
Cone production
The GA + D-treated trees produced significantly (P < 0.01) more seed cones than the C, D-, and GA-treated trees (Table 1). All of the 162 seed cones induced on the GA + D-treated trees proliferated (see Caron and Powell 1991). In 1994, the GA + RP-treated trees produced more seed and pollen cones than the control trees, but the differences were only significant (P < 0.01) for seed cones (Table 1). Four of 13 control trees produced seed cones, but only one ramet produced pollen cones. In contrast to the 1993 experiment, only four of the cones initiated during the 1994 treatments proliferated, and all four cones were on one tree.
Clonal differences in cone production responses to the treat-ments were evident within a year, but the clones that failed to produce cones in response to GA + D in 1993 were not the clones that exhibited a poor response to GA + RP in 1994. The number of cones produced in response to the 1994 treatments was not affected by the GA treatment in March 1993. The ortets ranged from 17 to 64 years of age, but neither respon-siveness to treatment nor degree of cone proliferation in re-sponse to the 1993 treatments was related to ortet age.
Tree development and shoot elongation
In both 1993 and 1994, the treatments did not significantly affect the timing of shoot development or final shoot length. Similarly, the 1993 treatments had no effect on growth in 1994. Although asynchronous with natural vegetative bud develop-ment, the sequence of vegetative bud development appeared normal in both years (Smith 1995).
Correlations between cone production and shoot water potential and needle water content
In 1993, Ψpd values at the end of the drought period were generally lower in all trees, including the trees that had not
been subjected to drought, compared with pre-drought values. This difference was probably due in part to the difference in
Ψpd between current-year and 1-year-old shoots. Predawn shoot water potentials were significantly (P < 0.001) lower in drought-treated trees than in control trees. Although trees ex-hibiting the lowest Ψpd produced the most cones, only trees receiving both GA and drought produced cones. Shoot water potentials of drought-treated trees recovered to control values within 24 h of resuming irrigation. There were no significant differences in needle water content between treatments. The dry weight/fresh weight ratio of developing needles generally increased up to CSE (Smith 1995), but neither needle fresh weight nor needle dry weight was correlated with Ψpd or successful cone induction in either year.
In 1993, mean (± SE) field capacity for all pots was 44.6 ± 2.0% and ranged from 35.8 to 56.7%. In 1993, container water contents generally remained above 95% of field capacity (Smith 1995), except during the drought period. By the end of the drought period, container water contents in the D and GA + D treatments were reduced to 58.6 ± 10.2 and 60.3 ± 9.2% of field capacity, respectively, and there were significant (P < 0.001) differences among clones. In 1994, container water content, measured 1 hour before irrigation, decreased from 88% of field capacity at the time of vegetative bud burst to 62% of field capacity at the time when shoot elongation stopped, but remained constant thereafter.
Correlations between container water content and Ψpd of trees subjected to 3 and 7 days of drought were R = −0.6204 (P < 0.0179) and R = −0.7304 (P < 0.0030), respectively. The corresponding correlations for irrigated trees were also nega-tive, but they were not significant: R = −0.3269 (P = 0.2540) and R = −0.3405 (P = 0.2236). Gibberellin injections had no effect on the rate of water use. Container water content was higher for root-pruned trees than for control trees, but the differences were not significant.
Needle development
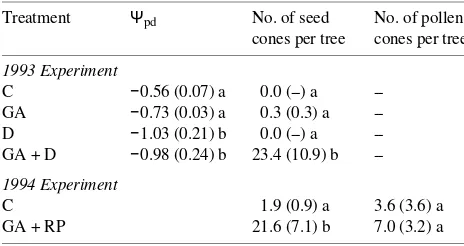
In both 1993 and 1994, the mean surface area of developing current-year needles increased little prior to the late phase of slow shoot elongation, but steadily thereafter, whereas needle dry weight increased gradually throughout the sampling pe-riod (Figure 1, Smith 1995). None of the treatments had a significant effect on needle characteristics in 1993. Needle weight, surface area, leaf dry weight/leaf area ratio, nitrogen content and chlorophyll concentration were positively corre-lated with each other in both years (Smith 1995). In 1994, increases in needle dry weight and surface area with successive collections were less for GA + RP-treated trees than for control trees, resulting in smaller needles with a higher leaf dry weight/leaf area ratio in the treated trees (Figure 1).
In both years, total N content per needle and total chloro-phyll concentration (Figure 2B, C, Smith 1995) increased from ESE to CSE, whereas percent N (Figure 2A) and the N/chlo-rophyll ratio decreased (Table 2). In 1994, total N content and total chlorophyll concentration of needles were lower in GA + RP trees than in C trees (Figures 2B and 2C). The
N/chloro-Table 1. Mean (SE) number of seed cones and pollen cones per treated tree and mean (SE) predawn shoot water potential (Ψpd, MPa) of treated trees. The 1993 treatments were: C = control, GA = stem injection with 10 mg GA4/7, D = drought (irrigation withheld for
7 days), and GA + D; and the 1994 treatments were: C = control and GA + RP = stem injection with 10 mg GA4/7 plus root pruning (RP).
Within a year, means followed by different letters are significant at
P < 0.05.
Treatment Ψpd No. of seed No. of pollen cones per tree cones per tree
Figure 1. Means (± 1 SE) of (A) needle dry weight, (B) needle surface area (SA), and (C) needle dry weight/needle area ratio (W) for control trees (d); and trees receiving GA4/7 plus root pruning (j). ESE = early
phase of slow shoot elongation, e-RSE = early rapid phase of shoot elongation, m-RSE = mid rapid phase of shoot elongation, LSE = late slow phase of shoot elongation, and CSE = cessation of shoot elongation.
Figure 2. Means (± 1 SE) of (A) needle nitrogen concentration (per-cent dry weight), (B) needle nitrogen content (µg per needle), and (C) total chlorophyll concentration (mg gDW−1) for control trees (d) and
trees receiving GA4/7 plus root pruning (j). See caption to Figure 1
for definition of abbreviations.
Table 2. Comparison of means (SE) of nitrogen/chlorophyll ratios for cone-bearing (F) and non-cone-bearing (NF) grafts in 1993 and 1994, and between drought-treated (D) and control (C), and between root-pruned (GA + RP) and control (C) trees at different stages of shoot development. Treatment means within collections followed by different letters are significant at P < 0.05. ESE = early phase of slow shoot elongation, RSE = rapid shoot elongation phase, LSE = late phase of slow shoot elongation, CSE = cessation of shoot elongation, and CSL = completion of shoot lignification.
Stage of shoot development
1993 Experiment ESE RSE CSE CSL
F 40.2(4.7) a 11.2(0.5) a 5.4(0.3) a 6.1(0.5) a NF 36.4(2.0) a 10.7 4.7(0.3) a 5.7(0.5) a C 36.4(1.9) a 9.9(0.6) a 5.0(0.2) a 6.7(0.5) b D 34.0(2.2) a 9.0(0.4) a 4.7(0.2) a 5.3(0.2) a
1994 Experiment ESE early-RSE mid-RSE LSE CSE
phyll ratio was also lower in GA + RP trees than in C trees but the differences were only significant (P < 0.05) for collections made during early RSE and LSE (Table 2). Needle growth
parameters did not differ between cone-producing and non-cone-producing trees in either year.
Gas exchange measurements
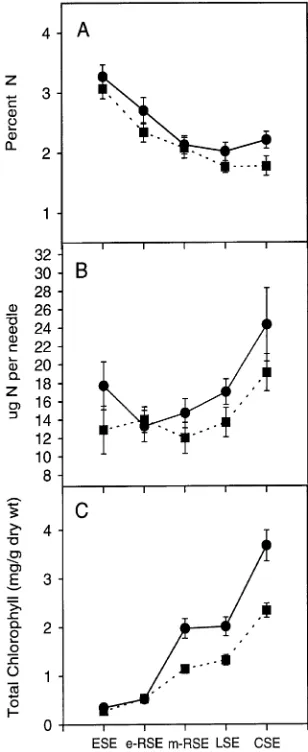
Net photosynthesis was lower for GA + RP trees than for C trees from ESE to mid-RSE (Figure 3). The standard errors of the gas exchange measurements were large during the phase of slow shoot elongation. Net photosynthesis and stomatal con-ductance were lower for drought-treated trees than for control trees during ESE and were negative during the later stages of the drought period; however gas exchange returned to control values within 24 h of watering the drought-treated trees.
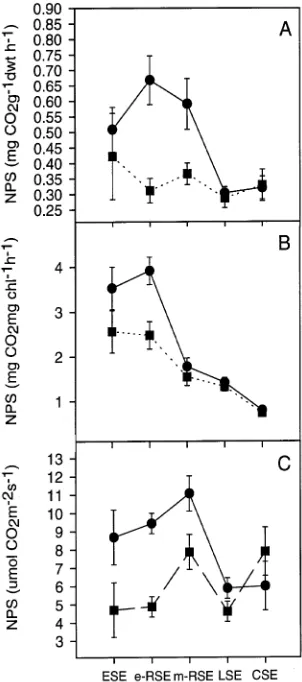
The GA + RP trees exhibited significantly lower net photo-synthesis (Figure 3) and stomatal conductance (Figure 4) than control trees for all clones. In both 1993 and 1994, gas ex-change measurements differed significantly among shoot de-velopment stages (Table 3).
Figure 3. Means (± 1 SE) of (A) net photosynthesis per gram dry weight, (B) net photosynthesis per mg total chlorophyll, and (C) net photosynthesis on a surface area basis for control trees (d); and trees
receiving GA4/7 plus root pruning (j). See caption to Figure 1 for
definition of abbreviations.
Figure 4. Means (± 1 SE) of stomatal conductance for control (d) trees
and trees receiving GA4/7 (j) plus root pruning. See caption to Figure
1 for definition of abbreviations.
Table 3. Summary of mean squares and probabilities (P) for gas exchange measurements: Pn,DW = net photosynthesis on a dry weight basis, Pn,area = net photosynthesis on a surface area basis, Pn,chl = net photosynthesis on a chlorophyll basis, and gs = stomatal conductance, CD =
collection date, ns = not significant. Values represent analyses on log transformed data.
Treatment df Pn,DW Pn,area Pn,chl gs 1993 Experiment
CD (C) 3 19.71 (0.001) 35.25 (0.001) 83.27 (0.001) 9.22 (0.001) Treatment (T) 3 0.68 ns 0.9 ns 0.54 ns 0.04 ns C × T 9 0.25 ns 0.09 ns 0.28 ns 0.51 ns
Error 138 0.34 0.64 0.38 0.34
1994 Experiment
CD (C) 4 1.89 (0.002) 1.84 (0.003) 12.78 (0.001) 5.38 (0.001) Treatment (T) 1 5.06 (0.001) 4.99 (0.001) 2.93 (0.005) 8.55 (0.001) C × T 4 0.64 ns 0.96 (0.073) 0.39 ns 0.69 (0.082)
Discussion
In 1993, the drought treatment had no persistent effect on needle development or gas exchange properties. However, only trees receiving both the GA and drought treatments pro-duced cones. Drought may reduce oxidative metabolism of exogenously applied GA, thereby increasing the less polar or florigenic forms (Chalupka et al. 1982, Dunberg et al. 1983).
Although proliferated cones comprise only a small percent-age of the total numbers of cones produced under natural conditions (Caron and Powell 1991), all of the cones induced in 1993 proliferated. Increased rates of cone proliferation in response to stresses imposed during cone induction is common (Ross and Pharis 1987); however, this is the first report of 100% of induced cones being proliferated. Although seed-cone development in response to the GA + D treatment in 1993 was abnormal, the elongating vegetative shoots and differenti-ating vegetative primordia developed normally (Smith 1995). Unfortunately, the GA and root pruning treatments were not tested separately in 1994; however, their combination resulted in a significant increase in cone production compared with the untreated controls. Several modes of action by which root pruning increases the efficacy of exogenously applied GAs in inducing cone production have been proposed: (1) bud devel-opment may be delayed to a time when warm and dry weather conditions prevail and thus favor buds differentiating repro-ductively; (2) the numbers of buds that are receptive to GA may be increased (Owens 1987, Owens et al. 1992); or (3) cytokinin export from the roots may be reduced, thereby either slowing cell division within developing buds and increasing their period of receptivity to both endogenous and exo-genously applied GA4/7, or relieving the bud of the presence of cytokinins, if inhibitory to cone production (Smith and Green-wood 1995).
Successful cone induction was not correlated with any measurable effects on shoot elongation. The failure of the D and RP treatments to reduce shoot elongation is contrary to the results of Webber et al. (1985), but similar to the findings of Hall (1988), Ross (1991b), and Smith and Greenwood (1995). Cone induction treatments may have a greater effect on the elongation of leaders and vigorous terminal shoots than the less vigorous laterals measured in this study (Philipson et al. 1990).
In 1993, although drought increased Ψpd, the effects did not persist when the trees were rewatered (cf. Ross 1991a). Root pruning had no significant effect on Ψpd. This observation is contrary to findings in other studies (Ross et al. 1985, Webber et al. 1985), but supports the hypothesis that root pruning does not exert its effects by causing plant water stress, i.e., it does not simulate drought (Ross 1991a) even though both root pruning and drought treatments increase the cone-inducing effectiveness of GA.
In both 1993 and 1994, total N content and total chlorophyll concentration of needles were positively correlated and in-creased until lateral shoot elongation ceased (cf. Oku et al. 1977, Kozlowski et al. 1991). Drought had no effect on needle chlorophyll concentration; a similar finding has been reported for Picea abies (L.) Karst. (Cornic 1987). In 1994, current-year
needles from GA + RP trees had a lower concentration of total chlorophyll and a lower total N content than control trees (Figure 2). Similarly, the total chlorophyll concentration of current-year needles of field-grown black spruce was reduced following six foliar applications of GA4/7 (R. Smith, unpub-lished observations). Although synthesis of the GA precursor ent-kaurene (Graebe and Ropers 1978, Moore 1989) has been correlated with chlorophyll concentration (Moore and Cool-baugh 1991), we conclude that the total chlorophyll concentra-tion of current-year needles cannot be used as a general response indicator of successful cone induction in black spruce.
The reduction in N content of needles following the D and GA + RP treatments contrasts with many studies showing that N contents of shoots, needles, and buds are increased follow-ing drought (Cyr et al. 1990, Beyers et al. 1992) or successful cone induction, or both (Barnes and Bengston 1968, Ebell and McMullin 1970, Daoudi et al. 1994). The decrease in percent total N (dry weight basis) of developing needles during the period of shoot elongation was similar to that found for Picea sitchensis (Bong.) Carrière (Lewandowska and Jarvis 1977) and may have confounded the effects of treatments on needle N content.
Current-year needles of trees receiving the GA + RP treat-ment in 1994 were smaller than those of control trees. These smaller needles may have a reduced capacity to provide pho-tosynthate to developing cones in the following growth cycle, which would reduce the size of the previously initiated cone scales and would also explain why cones produced in response to GA plus an adjunct stress treatment are sometimes smaller than natural cones. Part of the reduction in tree growth follow-ing a good cone-bearfollow-ing year may be the result of lower needle areas in response to ‘‘stress,’’ i.e., dry weather, rather than the presence of female cones per se (Dick et al. 1990).
Conclusions
Both drought and root pruning, when applied within 10 days of vegetative bud burst, acted synergistically with GA to in-crease cone production, but these treatments affect current-year needle development and gas exchange properties differently. Drought-treated trees exhibited more negative pre-dawn pressure potentials and reduced rates of gas exchange than control trees, but these effects did not persist after trees were rewatered. There were no significant differences in need-le size, weight or needneed-le N or chlorophyll content between drought-treated and control trees. Conversely, when compared to control trees, current-year needles of GA + RP trees were smaller with lower N and chlorophyll contents, and exhibited lower net photosynthesis up to the latter stages of shoot elon-gation. We postulate that these smaller needles have a reduced capacity to provide photosynthate to the cones that mature in the following year.
Acknowledgments
assis-tance of Cindy Henderson in developing protocols and technical ex-pertise in using the Technicon for chlorophyll analyses is greatly appreciated. We also thank S.I. Cameron, C.H.A. Little, Y.S. Park and G. Powell for reviews of an earlier draft of this manuscript, and D. Perry for word processing. Finally, we wish to acknowledge Laurie Yeates for exemplary technical assistance throughout the project.
References
Barnes, R.L. and G.W. Bengston. 1968. Effects of fertilization, irriga-tion, and cover cropping on flowering and on nitrogen and soluble sugar composition of slash pine. For. Sci. 14:172--180.
Beyers, J.L., G.H. Reichers and P.J. Temple. 1992. Effects of long-term ozone exposure and drought on the photosynthetic capacity of ponderosa pine (Pinus ponderosa Laws.). New Phytol. 122:81--90. Bonnet-Masimbert, M. 1987. Floral induction in conifers: a review of
available techniques. For. Ecol. Manage. 19:135--146.
Bonnet-Masimbert, M. and J.B. Zaerr. 1987. The role of plant growth regulators in promotion of flowering. Plant Growth Regul. 6:13--35. Caron, G.E. and G.R. Powell. 1991. Proliferated seed cones and pollen
cones in young black spruce. Trees 5:65--74.
Chalupka, W., M. Giertych and J. Kopcewicz. 1982. Effects of poly-ethylene covers, a flower inducing treatment on the content of endogenous gibberellin-like substances in grafts of Norway spruce. Physiol. Plant. 54:79--82.
Cornic, G. 1987. Interaction between sublethal pollution by sulphur dioxide and drought stress: The effect on photosynthetic capacity. Physiol. Plant. 71:115--119.
Cyr, D.R., G.F. Buxton, D.P. Webb and E.B. Dumbroff. 1990. Accu-mulation of free amino acids in the shoots and roots of three northern conifers during drought. Tree Physiol. 6:293--303. Daoudi, E.H., P. Doumas and M. Bonnet-Masimbert. 1994. Changes
in amino acids and polyamines in shoots and buds of Douglas-fir trees induced to flower by nitrogen and gibberellin treatments. Can. J. For. Res. 24:1854--1863.
Dick, J. McP., R.B.B. Leakey and P.G. Jarvis. 1990. Influence of female cones on the vegetative growth of Pinus contorta trees. Tree Physiol. 6:151--163.
Dickmann, D.I. and T.T. Kozlowski. 1968. Mobilization by Pinus resinosa cones and shoots of 14C-photosynthate from needles of different ages. Am. J. Bot. 55:900--906.
Dickmann, D.I. and T.T. Kozlowski. 1970. Mobilization and incorpo-ration of photo-assimilated 14C by growing vegetative and repro-ductive tissues of adult Pinus resinosa Ait. trees. Plant Physiol. 45:284--288.
Dunberg, A., G. Malmberg, T. Sassa and R.P. Pharis. 1983. Metabo-lism of tritiated gibberellins A4, and A9 in Norway spruce, Picea abies (L.) Karst. Plant Physiol. 71:257--262.
Ebell, L.F. and E.E. McMullin. 1970. Nitrogenous substances associ-ated with differential cone production responses of Douglas-fir to ammonium nitrate fertilization. Can. J. Bot. 48:2169--2177. Graebe, J.E. and H.J. Ropers. 1978. Gibberellins. In Phytohormones
and Related Compounds----A Comprehensive Treatise, Vol. 1. Eds. D.S. Letham, P.B. Goodwin and T.J.V. Higgins. Elsevier, Holland, Chapter 3, 641 p.
Greenwood, M.S., C.A. Hopper and K.W. Hutchison. 1989. Matura-tion in larch. I. Effects of age on shoot growth, foliar characteristics, and DNA methylation. Plant Physiol. 90:406--412.
Greenwood, M.S., G. Adams and S. Kempton. 1993. Using recent advances in the control of conifer reproductive development to increase genetic gain. In Proc. 24th Mtg. Can. Tree Improv. Assoc., Part 2. Ed. J. Lavereau. Fredericton, N.B., pp 19--26.
Hall, P.J. 1988. Promotion of flowering in black spruce using gibberel-lins. Silvae Genet. 37: 3--4.
Hiscox, J.D. and G.F. Israelstam. 1979. A method for the extraction of chlorophyll from leaf tissue without maceration. Can. J. Bot. 57:1332--1334.
Ho, R.H. 1988. Gibberellin A4/7 enhances seed-cone production in
field-grown black spruce. Can J. For. Res. 18:139--142.
Ho, R.H. 1991a. Timing of gibberellin A4/7 application for cone
production in potted black spruce grafts. Can. J. For. Res. 21:1137--1140.
Ho, R.H. 1991b. Promotion of cone production in potted black-spruce grafts, using gibberellins, heat-treatment and root-pruning. For. Ecol. Manage. 40:243--260.
Kozlowski, T.T., P.J. Kramer and S.G. Pallardy. 1991. The physiologi-cal ecology of woody plants. Academic Press, New York, 657 p. Lewandowska, M. and P.G. Jarvis. 1977. Changes in chlorophyll and
carotenoid content, specific leaf area and dry weight fraction in sitka spruce, in response to shading and season. New Phytol. 79:247--256.
Moore, T.C. 1989. Biochemistry and physiology of plant hormones. Springer Verlag, New York, 330 p.
Moore, T.C. and R.C. Coolbaugh. 1991. Correlations between appar-ent rates of ent-kaurene biosynthesis and parameters of growth and development in Pisum sativum. In Gibberellins. Eds. N. Takahashi, B.O. Phinney and J. Macmillan. Springer-Verlag, New York, pp 188--198.
Oku, T., S. Furuta, K. Shimado, Y. Kobayashi, T. Ogawa, Y. Indie and K. Shibata. 1977. Development of the photosynthetic apparatus in spruce leaves. In Photosynthetic Organelles: Structure and Func-tion. Jap. Plant Physiol. Centre, Acad. Publ., Nakanishi Print Co., Kyoto, Japan, pp 437--444.
Oren, R., E.-D. Schulze, R. Maryssek and R. Zimmermann. 1986. Estimating photosynthetic rate and annual carbon gain in conifers from specific leaf weight and leaf biomass. Oecologia 70:187--193. Owens, J.N. 1987. Development of Douglas-fir apices under natural
and cone-inducing conditions. For. Ecol. Manage. 19:85--97. Owens, J.N. 1991. Measuring growth and development of
reproduc-tive structures. In Techniques and Approaches in Forest Tree Eco-physiology. Eds. J.P. Lassoie and T.M. Hinckley. CRC Press, Boca Raton, FL, pp 423--452.
Owens, J.N., J.J. Philipson and D.L.S. Harrison. 1992. The effects of the duration and timing of drought plus heat plus gibberellin A4/7 on
apical meristem development and coning in Sitka spruce (Picea sitchensis (Bong.) Carr.). New Phytol. 122:515--528.
Pharis, R.P., J.E. Webber and S.D. Ross. 1987. The promotion of flowering in forest trees by gibberellin A4/7 treatments: a review of
the possible mechanisms. For. Ecol. Manage. 19:65--84.
Philipson, J.J. 1987. Promotion of cone and seed production by gib-berellin A4/7 and distribution of pollen and seed cones on Sitka
spruce grafts in a clone bank. For. Ecol. Manage. 19:147--154. Philipson, J.J. 1990. Prospects for enhancing flowering of conifers and
broadleaves of potential silvicultural importance in Britain. For-estry 63:223--240.
Philipson, J.J., J.N. Owens and M.A. O’Donnell. 1990. Production and development of seed and pollen in grafts of Picea sitchensis
(Bong.) Carr. treated with gibberellin A4/7 to induce coning and the
effect of forcing treatments. New Phytol. 116:695--705.
Puritch, G.S., E.E. McMullin, M.D. Meager and C.S. Simmons. 1979. Hormonal enhancement of cone production in Douglas-fir grafts and seedlings. Can. J. For. Res. 9:193--200.
Ross, S.D. 1988. Pre- and post-pollination polyhouse environment effects on pollen and seed development in potted Picea engelmannii
Ross, S.D. 1989. Long term cone production and growth responses to crown management and gibberellin A4/7 treatment in a young
west-ern hemlock seed orchard. New For. 3:235--245.
Ross, S.D. 1991a. Effect of heat sums and of heat applied separately to shoots and roots on flowering in potted Picea glauca grafts. Can. J. For. Res. 21:672--679.
Ross, S.D. 1991b. Promotion of flowering in potted white spruce grafts by root pruning: its relationship to drought and shoot elonga-tion. Can. J. For. Res. 21:680--685.
Ross, S.D. and R.P. Pharis. 1981. Recent developments in enhance-ment of seed production in conifers. In Seed Orchards and Strate-gies for Tree Improvement, Part 2. Eds. D.F.W. Pollard, D.G. Edwards and C.W. Yeatman. Proc. 18th Mtg. Can. Tree Improv. Assoc., Duncan, B.C. pp 26--38.
Ross, S.D. and R.P. Pharis. 1987. Control of sex expression in conifers. Plant Growth Regul. 6:37--60.
Ross, S.D. and R.C. Bower. 1989. Cost-effective promotion of flow-ering in a Douglas-fir seed orchard by girdling and pulsed stem injection of gibberellin A4/7. Silvae Genet. 38:189--194.
Ross, S.D., R.P. Pharis and J.C. Heaman. 1980. Promotion of cone and seed production in grafted and seedling Douglas-fir seed orchards by application of gibberellin A4/7 mixture. Can. J. For. Res.
10:464--469.
Ross, S.D., J.E. Webber, R.P. Pharis and J.N. Owens. 1985. Interaction between gibberellin A4/7 and root-pruning on the reproductive and
vegetative process in Douglas-fir. I. Effects on flowering. Can. J. For. Res. 15:341--347.
Ross, S.D., A.M. Eastham and R.C. Bower. 1986. Potential for indoor-potted seed orchards. In Proc. Conifer Tree Seed in the Inland Mountain West Symp., Missoula, Montana. Ed. R. Shearer. USDA For. Serv. Intermountain Res. Stn., Gen Tech. Rep. INT-203, pp 180--186.
Smith, R.F. 1995. Effects of cone induction treatments on seed- and pollen-cone production, bud anatomy, and needle development in black spruce (Picea mariana (Mill.) B.S.P.). Ph.D. Thesis, Univ. Maine, Orono ME, 89 p.
Smith, R.F. and G.R. Powell. 1992. Is there a place for alternatives to conventional seed orchards in the Maritimes? Can. For. Serv., At-lantic R & D Tech. Note No. 4, 3 p.
Smith, R.F. and M.S. Greenwood. 1995. Effects of gibberellin A4/7,
root-pruning, and cytokinins on seed- and pollen-cone production in black spruce (Picea mariana (Mill.) B.S.P.). Tree Physiol. 15:457--466.
Snedecor, G.W. and W.G. Cochrane. 1980. Statistical methods. Iowa State University Press, Ames IA, 507 p.
Sweet, G.B., R.L. Dickson, B.D. Donaldson and H. Litchwark. 1992. Controlled pollination without isolation----a new approach to the management of radiata pine seed orchards. Silvae Genet. 41:95--99. Webber, J.E., S.D. Ross, R.P. Pharis and J.N. Owens. 1985. Interaction between gibberellin A4/7 and root-pruning on the reproductive and
vegetative process in Douglas-fir. II. Effects on shoot elongation and its relationship to flowering. Can. J. For. Res. 15:348--353. Whitcomb, C.E. 1988. Plant production in containers. Lacebark