i
POTENSI
Streptomyces
sp. PADA RIZOSFER TANAMAN
ZINGIBERACEAE DALAM MENGHAMBAT
MULTIDRUG
RESISTANT Acinetobacter baumannii
Skripsi
Sebagai tugas akhir untuk memenuhi syarat mencapai derajat Sarjana S-1 Program Studi Biologi FMIPA Universitas Udayana
Oleh
Ni Kadek Losiani 1208305003
PROGRAM STUDI BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM UNIVERSITAS UDAYANA
ii SKRIPSI
POTENSI
Streptomyces
sp. PADA RIZOSFER TANAMAN
ZINGIBERACEAE DALAM MENGHAMBAT
MULTIDRUG
RESISTANT Acinetobacter baumannii
Oleh Ni Kadek Losiani
1208305003
Telah dipertahankan di depan Tim Penguji dan telah dinyatakan lulus pada tanggal 26 Mei 2016
Menyetujui
Pembimbing I Pembimbing II
Dr. Dra. Retno Kawuri M.Phil. Dr. Drs. A. A. Ketut Darmadi, M.Si. NIP. 1961 0112 198803 2 002 NIP. 1968 0115 199303 1 001
Mengesahkan
Ketua Program Studi Biologi FMIPA Universitas Udayana
iii KATA PENGANTAR
Puji syukur penulis panjatkan ke hadirat Ida Sang Hyang Widhi Wasa/Tuhan Yang Maha Esa, karena berkat rahmat-Nya penulis dapat menyelesaikan Skripsi sebagai tugas akhir untuk memenuhi syarat mencapai derajat Sarjana S-1 Jurusan Biologi FMIPA UNUD yang berjudul “Potensi Streptomyces sp. pada Rizosfer Tanaman Zingiberaceae dalam Menghambat
Multidrug Resistant Acinetobacter baumannii
”
. Pada kesempatan ini, penulis mengucapan terima kasih kepada:1. Yth. Ibu Dr. Dra. Retno Kawuri, M.Phil. dan Dr. Drs. A. A. Ketut Darmadi, M.Si., selaku dosen pembimbing yang telah memberikan ide-ide, masukan, dan bimbingan selama penulisan skripsi ini.
2. Yth. Bapak Dr. Drs. Ida Bagus Gede Darmayasa, M.Si., Ibu Dr. Ni Luh Suriani, S.Si., M.Si., dan ibu Dr. Dra. Meitini W. Proborini, M.Sc. selaku penguji yang telah memberikan masukan, kritik, dan saran selama penulisan skripsi ini.
3. Yth. Bapak Prof. Dr. Ir. I Wayan Kasa, M.Rur.Sc., selaku dosen pembimbing akademik yang telah memberikan bimbingan dan nasehat selama studi.
4. Yth. Bapak Dr. I Ketut Ginantra S.Pd., M.Si. dan Bapak Dr. Drs. Ida Bagus Gede Darmayasa, M.Si., selaku ketua dan sekretaris jurusan biologi yang telah memberi izin dan fasilitas demi terlaksananya penelitian dan penyusunan skripsi ini.
5. Yth. Bapak Drs. Ida Bagus Made Suaskara, M.Si selaku dekan FMIPA serta seluruh panitia seminar dan staf pegawai Jurusan Biologi yang telah memberi bantuan dalam kelancaran administrasi selama penyusunan skripsi ini.
iv 7. Kayika Febryanti, Mas Suratniasih, dan Diah Purnama sebagai sahabat yang telah memberi semangat dan dukungan selama penelitian. Kepada Diah Kharismawati Djereng dan Wiwik Sinta yang sudah menemani selama penelitian di Laboratorium dan memberi saran selama penulisan skripsi ini. Kepada Gede Putra Wirantana terimakasih atas motivasi serta bantuan dalam melancarkan penelitian ini. Kepada seluruh teman-teman di Jurusan Biologi dan semua pihak yang tidak dapat penulis sebutkan satu persatu, yang telah banyak memberikan bantuan dan dukungan sehingga skripsi ini dapat terselesaikan.
Penulis menyadari bahwa skripsi ini masih banyak kekurangan baik pada teknis penulisan maupun materi, maka dari itu, saran dan kritik yang bersifat membangun sangat penulis harapkan. Akhir kata penulis mengucapkan terimakasih.
Jimbaran, 27 April 2016
v ABSTRAK
Acinetobacter baumannii merupakan salah satu patogen yang sudah mengarah ke
multidrug resistant (MDR-A baumannii). Patogen ini sering menjadi penyebab
wabah infeksi pada pasien yang mendapat perawatan di ruang rawat intensif di rumah sakit. Penelitian ini bertujuan untuk mengetahui kemampuan isolat
Streptomyces sp. yang diisolasi pada rizosfer tanaman famili Zingiberaceae dalam
menghambat pertumbuhan MDR-A. baumannii, mengetahui potensi filtrat
Streptomyces sp. dalam menghambat pertumbuhan MDR-A baumannii, dan
mengetahui Minimum Inhibitory Concentration (MIC) filtrat Streptomyces sp. dalam menghambat MDR-A baumannii. Bakteri Streptomyces diisolasi pada rizosfer tanaman temu ireng (Curcuma aeruginosa Roxb), lengkuas (Alpinia
galanga L.), jahe merah (Zingiber officinale Rosc.), dan kunyit putih (Curcuma
mangga Val.). Isolat MDR-A. baumannii diperoleh dari stock culture
Laboratorium Mikrobiologi Klinik RSUP Sanglah Denpasar. Uji antagonistik isolat Streptomyces sp. terhadap pertumbuhan MDR-A. baumannii menggunakan metode dual culture dan untuk mengetahui MIC filtrat Streptomyces sp. menggunakan metode sumur difusi. Rancangan penelitian yang digunakan adalah Rancangan Acak Lengkap (RAL) dan data diameter zona hambat uji MIC dianalisis dengan Analysis of Varian (ANOVA) kemudian dilanjutkan dengan uji
Duncan Multiple Range Test taraf signifikan 5 %. Hasil penelitian diperoleh 5
isolat Streptomyces sp. (LCM1, KCM2, KAG3, LCM4, dan LZO5). Isolat
Streptomyces sp. KCM2 menghasilkan diameter zona hambat terbesar terhadap
MDR-A. baumannii yaitu 15,59 mm sehingga dilanjutkan untuk diisolasi
filtratnya dan diuji MIC. Filtrat Streptomyces sp. KCM2 mampu menghambat dengan diameter zona hambat sebesar 23,44 mm dan MIC filtratnya adalah 4% (v/v) dengan diameter zona hambat 8,77 mm.
Kata kunci: Streoptomyces sp., MDR-Acinetobacter baumannii, filtrat
vi ABSTRACT
Acinetobacter baumannii is one of the pathogen which leads to multidrug resistant
(MDR-A baumannii). This pathogen is often causing outbreaks of infections to the patients which are receiving treatment in the intensive care unit in hospital. This study aims to find out isolate’s ability of Streptomyces sp. which is isolated on the plants Zingiberaceae rhizosphere in inhibiting the growth of MDR-A. baumannii, to find out filtrate potential of Streptomyces sp. in inhibiting the growth of
MDR-A baumannii, and to find out the Minimum Inhibitory Concentration (MIC) of
Streptomyces sp. filtrate in inhibiting the growth of MDR-A baumannii.
Streptomyces bacteria were isolated in rhizosphere of black zingiber (Curcuma
aeruginosa Roxb), galangal (Alpinia galanga L.), red ginger (Zingiber officinale
Rosc.), and white turmeric (Curcuma mango Val.). MDR-A. baumannii isolates were obtained from stock culture in Clinical Microbiology Laboratory Sanglah Hospital in Denpasar. The antagonistic test of Streptomyces sp. isolates to the growth of MDR-A. baumannii was using dual culture method, meanwhile to find out the MIC of Streptomyces sp. filtrate was using the wells diffusion method. The research design used was Completely Randomized Design (CRD) and the inhibitory zone diameter data of MIC test were analyzed by Analysis of Varian (ANOVA), and then continued by Duncan Multiple Range Test in significance level 5%. The results of this study obtained 5 isolates of Streptomyces sp. (LCM1, KCM2, KAG3, LCM4, and LZO5). Streptomyces sp. KCM2 produce the largest diameter of inhibition zone against MDR-A. baumannii which is 15.59 mm, therefore it is continued to be isolated its filtrate and MIC tested. The
Streptomyces sp. KCM2 filtrate was able inhibiting the inhibition zone diameter
of 23,44 mm and MIC of filtrate was 4% (v/v) with inhibition zone diameter of 8.77 mm.
vii
2.2. Tanaman Familia Zingiberaceae ... 8
2.3. Antibiotik ... 10
2.3.1. Mekanisme kerja antibiotik... 10
2.3.2. Mekanisme resistensi bakteri terhadap antibiotik ... 14
2.4. Bakteri Acinetobacter baumannii ... 15
viii
3.1.3. Isolasi bakteri Streptomyces ... 20
3.1.4. Pengamatan makroskopis koloni bakteri Streptomyces ... 21
3.1.5. Pengamatan mikroskopis bakteri Streptomyces ... 21
3.1.6. Identifikasi bakteri Streptomyces ... 22
3.1.7. Uji aktivitas antagonistik isolat Streptomyces sp. terhadap multidrug resistant A. baumannii secara in vitro ... 22
3.1.8. Pembuatan filtrat Streptomyces sp. KCM2 ... 23
3.1.9. Uji aktivitas filtrat Streptomyces sp. KCM2 terhadap multidrug resistant A. baumannii ... 24
3.1.10. Rancangan penelitian ... 25
3.1.11. Variabel pengamatan ... 26
3.2. Metode Pengolahan Data ... 26
IV. HASIL DAN PEMBAHASAN 4.1. Isolasi Streptomyces sp. pada Rizosfer Tanaman Zingiberaceae ... 27
4.2. Aktivitas Antagonistik Isolat Streptomyces sp. terhadap Multidrug Resistant Acinetobacter baumannii secara in Vitro ... 33
4.3. Aktivitas Filtrat Streptomyces sp. KCM2 terhadap Multidrug Resistant Acinetobacter baumannii ... 37
V. KESIMPULAN DAN SARAN 5.1. Kesimpulan ... 43
5.2. Saran ... 43
DAFTAR PUSTAKA ... 44
ix DAFTAR TABEL
Halaman Tabel 1. Rancangan acak lengkap pada uji aktivitas filtrat Streptomyces sp.
terhadap MDR-A. baumannii ... 25 Tabel 2. Isolat Streptomyces sp. yang terisolasi dari rizosfer tanaman
Zingiberaceae pada 2 Kawasan ... 27 Tabel 3. Rata-rata diameter zona hambat isolat Streptomyces sp. terhadap
multidrug resistant Acinetobacter baumannii ... 34
x Gambar 4. Biofilm A. baumannii (in vitro) yang terbentuk pada permukaan
penutup kaca setelah 24 jam inkubasi pada suhu 370C di media
Luria-bertani broth. Perbesaran 5.000 x, dengan Scanning
Electron Microscopy (SEM) ... 17 Gambar 5. Biofilm polimikroba (A. baumanii, K. pnomuniae, dan P.
aeruginosa) tumbuh dalam lumen kateter urin silikon yang
diisolasi dari pasien rumah sakit di Roma. Perbesaran 10.000 x, dengan Scanning Electron Microscopy (SEM) ... 17 Gambar 6. (A) Koloni Streptomyces sp. LCA1 usia 5 hari pada media YMA/ISP4
dan (B) Struktur mikroskopis (a. konidia dan b. hifa). Perbesaran 1.000 x dengan Mikroskop Cahaya (Yazumi)... 28
Gambar 7. (A) Koloni Streptomyces sp. KCM2 usia 5 hari pada media YMA/ISP4
dan (B) Struktur mikroskopis (a. konidia dan b. hifa). Perbesaran 1.000 x dengan Mikroskop Cahaya (Yazumi)... 29
Gambar 8. (A) Koloni Streptomyces sp. KAG3 usia 5 hari pada media YMA/ISP4
dan (B) Struktur mikroskopis (a. konidia dan b. hifa). Perbesaran 1.000 x dengan Mikroskop Cahaya (Yazumi)... 30
Gambar 9. (A) Koloni Streptomyces sp. LCA4 usia 5 hari pada media YMA/ISP4
dan (B) Struktur mikroskopis (a. konidia dan b. hifa). Perbesaran 1.000 x dengan Mikroskop Cahaya (Yazumi)... 30
Gambar 10. (A) Koloni Streptomyces sp. LZO5 usia 5 hari pada media YMA ISP4
dan (B) Struktur mikroskopis (a. konidia dan b. hifa). Perbesaran 1.000 x dengan Mikroskop Cahaya (Yazumi)... 31
Gambar 11. Daya Hambat isolat Streptomyces terhadap MDR-A. baumannii.
(1) Streptomyces sp. LCA1, (2) Streptomyces sp. KCM2, (3)
Streptomyces sp. KAG3, (4) Streptomyces sp. LCA4, (5)
Streptomyces sp. LZO5, dan (6) Kontrol negative ... 34
xi Gambar 13. Uji MIC Filtrat Streptomyces sp. KCM2 dengan konsentrasi
xii DAFTAR LAMPIRAN
Halaman
Lampiran 1. Hasil analisis Rancangan Acak Lengkap uji in vitro filtrat
Streptomyces sp. KCM2 dengan perlakuan filtrat konsentrasi
1
I. PENDAHULUAN
1.1. Latar Belakang
Antibiotik digunakan untuk membunuh bakteri yang dapat menyebabkan penyakit sehingga antibiotik dikatakan berkontribusi besar untuk kesehatan manusia (Chopra and Roberts, 2001). Antibiotik pertama kali digunakan untuk mengobati infeksi serius pada tahun 1940-an. Sejak tahun itulah antibiotik menjadi penyelamat jutaan nyawa, namun selama 70 tahun terakhir bakteri telah menunjukkan kemampuan untuk menjadi resisten terhadap setiap antibiotik yang telah dikembangkan (Saga and Yamaguchi, 2009). Bakteri resisten merupakan bakteri yang tidak dapat dikendalikan atau dibunuh dengan antibiotik karena bakteri tersebut terus menerus mengalami perubahan antigenik untuk melawan kekebalan tubuh pasien dan terapi antibiotik. Resistensi antibiotik dapat menimbulkan penyakit serius bagi pasiennya sehingga hal ini menjadi ancaman paling serius dalam dunia kesehatan (Gustawan dkk., 2014). Salah satu bakteri patogen yang resisten terhadap beberapa antibiotik adalah Acinetobacter
baumannii (Bou et al., 2000).
Bakteri A. baumannii resisten diakui sebagai salah satu bakteri aerobik golongan Gram negatif yang tahan terhadap beberapa antibiotik seperti ampisillin,
ceftazidime, meropenem, levofloxacin, amikacin, dan
trimethoprim-sulfamethoxazole (Cucunawangsih et al., 2015). Bakteri A. baumannii resisten sering menjadi penyebab wabah infeksi dan endemik. Hal tersebut didukung oleh kemampuannya yang dapat bertahan pada berbagai kondisi lingkungan dalam waktu yang lama (Fournier and Richet, 2006) dan bahkan dapat mengkolonisasi di tangan petugas kesehatan (Jawad et al., 1998). Bakteri A. Baumannii resisten ini diketahui memiliki spektrum klinis yang luas seperti bakteremia, pneumonia, meningitis, infeksi saluran kemih, infeksi kulit dan jaringan lunak, infeksi aliran darah, endokarditis, abses intra abdominal, dan infeksi luka operasi (Howard et al., 2010).
2 dan pasien yang menggunakan ventilator mekanik (Kanokorn and Peninnah, 2007). Resistensi A. baumannii terhadap beberapa antibiotik diketahui mulai mewabah pada tahun 1992 di rumah sakit Barcelona, Spanyol. Tahun 1997, muncul A. baumannii yang resisten terhadap golongan carbapenem (Corbella et al., 2000). Munoz and Weinstein (2008), mengatakan bahwa A. baumannii menjadi perhatian utama karena sudah mengarah ke Multidrug Resistant A.
baumannii (MDR-A. baumannii). Diketahui bahwa MDR-A. Baumannii ini sudah
mewabah di negara-negara Asia dan Timur Tengah (Hong et al., 2012).
Antibiotik memiliki sifat intrinsik yang tidak bergantung pada ukuran dan jumlah materinya, oleh karena itu resistensi terhadap mikroba target dapat muncul sehingga antibiotik ini tidak lagi efektif untuk pengobatan (Saga and Yamaguchi, 2009). Berkaitan dengan hal tersebut, maka perlu dilakukan penemuan-penemuan antibiotik baru baik secara sintesis kimia, biokimia, atau melalui isolat mikroba baru (Chopra et al., 2002). Isolasi antibiotik sebelumnya sudah banyak dilakukan pada galur-galur mikroba dan sudah banyak diujikan pada beberapa bakteri patogen. Antibiotik merupakan senyawa bioaktif metabolit sekunder yang dihasilkan oleh bakteri. Namun tidak semua bakteri dapat menghasilkan antibiotik yang berpotensi untuk menghambat bakteri patogen lainnya (Sahin and Ugur, 2003). Potensi besar penghasil antibiotik salah satunya dimiliki oleh bakteri
Streptomyces sp. (Demain, 1999). Dharmawan (2009) melaporkan bahwa isolat
Streptomyces sp. yang diisolasi pada kawasan hutan provinsi Bali mampu
menghambat lima strain diarrheagenic Escherichia coli yaitu EHEC, ETEC, EIEC, EPEC dan DAEC. Penelitian lain yang dilakukan oleh Raningsih (2015) mengatakan
bahwa salah satu genus dari kelas Actinomycetes yaitu isolat Nocardia sp. yang diisolasi
pada perkebunan cengkeh mampu menghambat Methicillin Resistant Staphylococcus
aureus (MRSA).
3 juga digunakan sebagai senjata bagi Streptomyces sp. untuk bersaing dengan mikroba lain atau bahkan dengan genus yang sama (Chater and Chandra, 2006). Budiyanto (2002), menegaskan bahwa tanah rizosfer memiliki peluang besar dikolonisasi oleh Streptomyces sp. karena di daerah tersebut tersedia banyak bahan organik sebagai sumber nutrisi.
Banyak peneliti yang sudah berhasil mengisolasi bakteri Streptomyces dari berbagai rizosfer tanaman. Seperti pada penelitian Djatmiko et al. (2007), yang telah berhasil mengisolasi 2 isolat Streptomyces dari rizosfer terung (Solanum
melongena). Hassanin et al. (2007), telah berhasil mengisolasi 4 isolat
Streptomyces yang teridentifikasi sebagai Streptomyces erumpens, S. purpureus,
S. aurantiacus dan S. Microflavus dari tanaman kapas (Caravonica katoen).
Ambarwati et al. (2010), berhasil mengisolasi 23 isolat Streptomyces dari rizosfer tanaman jagung (Zea mays) dan Kartika (2015), telah berhasil mengisolasi 9 isolat Streptomyces pada rizosfer tanaman berkayu (tanaman sawo, jati, flamboyan, bungur, trengguli, dan bougenvil).
Menurut Rao (1994), ketersediaan variasi senyawa organik dalam tanah dapat mempengaruhi kualitas dan kuantitas mikrobanya. Tanaman famili Zingiberaceae merupakan salah satu tanaman rimpang yang memiliki kandungan bahan organik yang cukup banyak yang dapat dilepaskan ke daerah rizosfernya (Wohlmut, 2008). Tanaman ini tersebar luas di daerah tropis dan sub tropis sehingga mudah ditemukan di Indonesia khusunya di Bali (Sirirugsa, 1998). Beberapa genus dari famili ini banyak dimanfaatkan dalam bidang farmakologi karena mengandung komponen kimia yang beragam (Kojima et al., 1998). Isolasi bakteri Streptomyces untuk diuji antagonistik dengan patogen MDR-A. baumannii sebelumnya belum pernah dilakukan pada rizosfer tanaman famili Zingiberaceae.
Berdasarkan latar belakang tersebut maka peneliti mencoba mengisolasi bakteri Streptomyces pada rizosfer beberapa jenis tanaman famili Zingiberaceae yaitu rizosfer tanaman temu ireng (Curcuma aeruginosa Roxb), lengkuas (Alpinia
galanga L.), jahe merah (Zingiber officinale Rosc.), dan kunyit putih (Curcuma
mangga Val.) yang diketahui memiliki beberapa senyawa bioaktif penting.
4 sehingga dapat diketahui potensi isolat – isolat Streptomyces yang ditemukan dalam menghambat pertumbuhan MDR-A. baumannii.
1.2. Rumusan Masalah
Berdasarkan latar belakang tersebut, maka rumusan masalah yang dapat diajukan dalam penelitian ini adalah sebagai berikut :
1. Apakah isolat Streptomyces sp. yang diisolasi pada rizosfer tanaman famili Zingiberaceae mampu menghambat pertumbuhan MDR-A. baumannii secara in vitro?
2. Apakah filtrat Streptomyces sp. mampu menghambat pertumbuhan MDR-A.
baumannii?
3. Berapakah Minimum Inhibitory Concentration (MIC) dari filtrat
Streptomyces sp. dalam menghambat MDR-A. baumannii?
1.3. Tujuan penelitian
Tujuan dari penelitian ini adalah sebagai berikut:
1. Untuk mengetahui kemampuan isolat Streptomyces sp. yang diisolasi pada rizosfer tanaman famili Zingiberaceae dalam menghambat pertumbuhan
MDR-A. baumannii.
2. Untuk mengetahui potensi filtrat Streptomyces sp. dalam menghambat pertumbuhan MDR-A. baumannii.
3. Untuk mengetahui Minimum Inhibitory Concentration (MIC) dari filtrat
Streptomyces sp. dalam menghambat MDR-A. baumannii.
1.4. Manfaat Penelitian
Manfaat dari penelitian ini yaitu dapat memberikan informasi bahwa bakteri yang diisolasi dari rizosfer tanaman Zingiberaceae berpotensi untuk menghambat
MDR-A. baumannii. Selain itu filtrat yang paling efektif dapat digunakan sebagai
5 II. TINJAUAN PUSTAKA
1.1. Bakteri Streptomyces
Streptomyces merupakan bakteri yang menyerupai jamur berfilamen yang bersifat aerobik (Hopwood, 2007) dan tergolong dalam genus bakteri Gram positif yang memiliki materi genetik guanin dan sitosin cukup tinggi (69-73% mol) bila dibandingkan dengan bakteri lain seperti Eschercia coli hanya 50% (Bentley et al., 2002). Hal ini yang mendukung bakteri Streptomyces mampu membentuk substrat percabangan luas dan miselium aerial (Wendisch and Kutzner, 1992). Kebanyakan Streptomyces hidup sebagai saprofit dalam tanah, namun bakteri ini juga berhasil menghuni berbagai relung lainnya baik darat maupun perairan (Zhang et al., 1997).
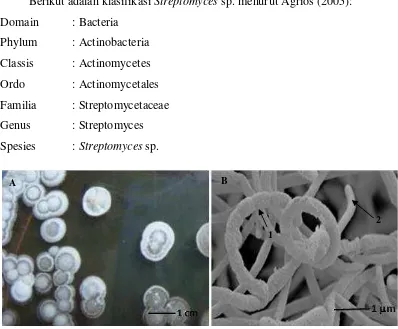
Perbedaan Streptomyces dengan bakteri lain yaitu pada media agar, koloni Streptomyces tumbuh secara perlahan yaitu koloni akan terlihat jelas pada inkubasi hari kedua atau hari ketiga. Koloni melekat erat pada permukaan media dan strukturnya kasar atau bertepung (Mathur et al., 2015)(Gambar 1), sedangkan bakteri lain tumbuh dengan cepat yaitu 24 jam inkubasi koloni sudah terlihat, serta struktur koloninya berlendir (Rao, 1994). Secara mikroskopis, hifa Streptomyces berbentuk ramping tanpa sekat dan berdiameter antara 0,5 – 2 µ m sedangkan konidianya berasal dari hifa yang terfragmentasi kemudian membentuk rantai konidia dengan diameter antara 0,3 – 1,5 µ m (Santhanam et al., 2013) (Gambar 1). Hain et al. (1997), mengatakan bahwa hifa dan konidia yang dimiliki oleh Streptomyces dapat menghasilkan pigmen serta aroma khas yang dapat dilihat pada media agar.
6 merupakan titik yang memiliki banyak fungsi. Di bagian ini, khususnya di daerah pertumbuhan apikal terjadi sekresi dan perakitan peptidoglikan serta komponen envelop sel lainnya, seperti asam teikoik, protein permukaan sel dan membran lipid.
Berikut adalah klasifikasi Streptomyces sp. menurut Agrios (2005): Domain : Bacteria
Phylum : Actinobacteria Classis : Actinomycetes Ordo : Actinomycetales Familia : Streptomycetaceae Genus : Streptomyces Spesies : Streptomyces sp.
Gambar 1. (A) Koloni Streptomyces sp. (Mathur et al., 2015) dan (B) Struktur mikroskopis (1. rantai konidia dan 2. hifa). Perbesaran 1.600 x, dengan Scanning Electron Micrographs (SEM) (Santhanam et al., 2013)
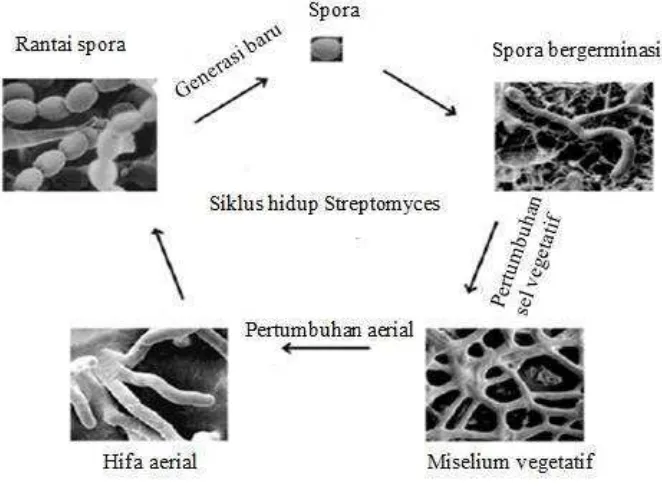
Streptomyces tumbuh sebagai percabangan hifa dengan miselium aerial multinukleat yang secara berkala membentuk septa lalu menghasilkan rantai konidia uninukleat. Ketika konidia berada pada kondisi yang menguntungkan, seperti suhu, nutrisi dan kelembaban yang cocok maka tabung bakteri dan hifa akan berkembang, selanjutnya terjadi pertumbuhan dan siklus sel (Gambar 2) (Wendisch and Kutzner, 1992). Chater and Chandra (2006), mengatakan bahwa kemungkinan sel sporogen mengandung 50 atau lebih kromosom. Urutan, posisi,
A B
7 dan pemisahan kromosom tersebut selama sporulasi terjadi secara linier yang melibatkan dua sistem yaitu ParAB dan FtsK. Sistem ini yang menyebabkan diferensiasi serta terbentuknya septa sel apikal yang membentuk rantai konidia.
Gambar 2. Siklus hidup Streptomyces sp. (Brooks et al., 2012)
Streptomyces sp. merupakan produsen antibiotik utama karena lebih dari
8 dan tingkat pertumbuhan (Bibb, 2005). Produksi antibiotik dapat terlibat pada proses penting lainnya seperti simbiosis Streptomyces sp. dengan tanaman yaitu tanaman dapat terlindungi dari serangan patogen, dan eksudat tanaman mendukung perkembangan Streptomyces sp. (Bosso et al., 2010).
1.2. Tanaman Familia Zingiberaceae
Zingiberaceae merupakan salah satu famili terbesar dari kerajaan tumbuhan. Famili tumbuhan ini memiliki 47 genera dan hampir 2.000 spesies yang tersebar diseluruh dunia. Zingiberaceae tersebar luas di daerah tropis dan sub tropis. Genera dan spesies Zingiberaceae banyak tersebar di kawasan Malesia (Indonesia, Malaysia, Singapura, Brunei, dan Papua Nugini) (Sirirugsa, 1998). Beberapa spesies Zingiberaceae diketahui memiliki senyawa bioaktif yang digunakan sebagai obat tradisional untuk berbagai penyakit (Wohlmut, 2008). Berikut adalah klasifikasi tanaman famili Zingiberaceae menurut Mabberley (1997):
Phylum : Angiospermae
Class : Monokotiledonae (Liliopsida) Subclass : Zingiberidae
Order : Zingiberales Famili : Zingiberaceae
9 Berbagai genus dari famili Zingiberaceae diketahui mengandung senyawa bioaktif yang bermanfaat seperti pada genus Curcuma yang diketahui memiliki senyawa kurkumin yang dapat bertindak sebagai antioksidan, anti kanker, dan anti inflamasi. Kurkuminoid dikenal juga dengan istilah diarilheptanoid berwarna yang terdapat pada rimpang tanaman yaitu sekitar 5% dari berat kering rimpang. Senyawa utama lainnya yaitu diferulol-metan, desmetoksi-kurkumin, bisdesmetoksi-kurkumin, dan dihidro-kurkumin sebanyak 50 – 60% yang terkandung di rimpang tanaman. Selain kurkumin, genus ini juga memiliki minyak atsiri sebanyak 5 - 6% yang terdiri dari mono dan seskuiterpen, termasuk zingiberin, kurkumin, α- dan β-turmeron (Evans, 2002). Genus Kaempferia dikenal sebagai obat untuk berbagai penyakit karena kandungan senyawa bioaktif yang dimiliki rimpang genus ini mengandung minyak atsiri dengan kandungan unsur etil-p-metoksi-e-sinamat, etil sinamat, sinamaldehid, kampen, l-Δ3-caren, borneol, p-metoksistiren dan pentadekan (Luger et al., 1996).
Genus Alpinia juga memiliki minyak atsiri yang diketahui tersebar di daun, batang, dan rimpang. Kandungan khas dari minyak esensial yang terdapat pada genus ini adalah terpenoid dan penilpropanoid, ini termasuk monoterpenoid seperti α- dan β-pinene, geraniol, borneol, sitronelol, linalool, 1,8-cineole dan kamper, seskuiterpenoid termasuk eudesmol, sesquiphellandrene β dan curcumene, dan phenylpropanoids seperti metil eugenol (Tewari et al., 1999). Aroma yang dihasilkan oleh genus ini mengandung unsur empat isomer asetoksi-sineol dan 1'-acetoksikavikol asetat yang menghasilkan sensasi pedas (Kubota et al., 1999). Genus Zingiber memiliki metabolit sekunder yang terdapat pada rimpang, senyawa ini terdiri dari senyawa volatil, nonvolatil, dan senyawa fenolik nonvolatil. Senyawa – senyawa inilah yang memiliki aktivitas farmakologi (Jolad et al., 2005).
10 tanah sekitar rimpang. Hal ini juga dapat sebagai alat untuk menarik mikroba yang dikehendaki hidup di sana maupun mengusir mikroba pengganggu (Widyati, 2013). Mikroba yang berkolonisasi di rizosfer akan berdampak pada modifikasi fisik dan kimia tanah, hal ini tentunya akan mempengaruhi tanaman dan mikroba itu sendiri (Sylvia et al., 2005).
1.3. Antibiotik
Antibiotik merupakan obat yang membunuh atau menghambat pertumbuhan bakteri. Antibiotik yang bekerja dengan membunuh bakteri disebut bakterisida, sedangkan antibiotik yang bekerja dengan mencegah bakteri bermultiplikasi disebut bakteriostatik (Chopra et al., 2002). Antibiotik pertama kali muncul sebagai terobosan terbaru dalam dunia kedokteran pada tahun 1928, yang ditemukan oleh seorang ilmuwan bernama Alexander Fleming (Taubes, 2008). Antibiotik dikatakan berspektrum luas bila berbagai macam infeksi dapat diobati dan berspektrum sempit bila hanya dapat mengobati beberapa jenis infeksi bakteri saja (Chopra et al., 2002). Antibiotik dengan jenis yang berbeda akan memberi pengaruh yang berbeda pada bakterinya dan dengan cara yang berbeda pula. Misalnya antibiotik melawan bakteri dengan menghambat kemampuan bakteri dalam mengubah glukosa menjadi energi, atau menghambat kemampuan bakteri dalam membentuk dinding selnya. Ketika hal ini terjadi maka bakteri akan mati dan tidak dapat bermultiplikasi (Kohanski et al., 2007). Tahun 1942, menjadi tahun pertama ditemukannya antibiotik yang berasal dari Streptomyces yaitu streptothricin dan dua tahun kemudian disusul dengan penemuan antibiotik kedua yang berasal dari Streptomyces yaitu streptomisin (Watve et al., 2001).
1.3.1. Mekanisme kerja antibiotik
11 A. Mekanisme antibiotik menghambat replikasi DNA
Proses sintesis DNA, mRNA transkripsi, dan pembelahan sel pada bakteri memerlukan modulasi kromosom melalui enzim topoisomerase untuk mengkatalis reaksi membukanya untai dan penggabungan untai kembali (Espeli and Marians, 2004). Reaksi ini telah dieksploitasi oleh antibiotik kelas kuinolon sintetis, termasuk fluoroquinolon. Kuinolon merupakan antibiotik turunan asam nalidiksat sebagai produk sampingan dari sintesis klorokuin. Kuinolon memiliki beberapa generasi yaitu asam oxolinik (pertama), ciprofloxacin (kedua), levofloxacin (ketiga), dan gemifloxacin (keempat). Antibiotik kelas kuinolon bekerja dengan mengganggu pemeliharaan topologi kromosom dengan cara menjebak enzim topoisomerase II dan topoisomerase IV pada tahapan pembelahan DNA dan mencegah untai bergabung kembali (Drlica et al., 2008). Drlica and Snyder (1978), mengatakan bahwa kerentanan topoisomerase II dan topoisomerase IV terhadap kuinolon bervariasi di setiap spesies bakteri walaupun keduanya memiliki kesamaan fungsi yang umum. Hasil penelitian lainnya menunjukkan bahwa pada bakteri Gram positif yang menjadi target utama kuinolon adalah topoisomerase IV (Munoz and Weinsten, 2008), sedangkan pada bakteri Gram negatif (misal: E. coli, dan Neisseria gonorrhea), yang menjadi target utama adalah topoisomerase II (Belland et al., 1994).
B. Mekanisme antibiotik menghambat sintesis RNA
Proses enzimatik sangat penting dalam pertumbuhan sel bakteri sehingga proses ini dapat menjadi target menarik bagi antibiotik. Rifampisin menghambat sintesis RNA dengan cara menghambat inisiasi transkripsi dan memblokir jalur pertumbuhan rantai ribonukleotida. Hal ini dapat terjadi karena Rifampisin menggunakan koneksi stabil dengan afinitas tinggi pada β-subunit di jalur kompleks RNA polimerase dan DNA yang menyebabkan situs aktif terpisah (Chopra et al., 2002). Rifampisin pertama kali diisolasi dari bakteri Gram positif
Amycolatopsis mediterranei yang awalnya dikenal sebagai Streptomyces
mediterranei pada tahun 1950-an. Rifampisin dengan karakter yang lebih kuat
12 bakterisida terhadap bakteri Gram positif dan bersifat bakteriostatik terhadap Gram negatif. Perbedaan tersebut telah dikaitkan dengan penyerapan antibiotik dan tidak afinitasnya antibiotik dengan β-subunit pada jalur RNA polimerase (Naryshkina et al., 2001).
C. Mekanisme antibiotik menghambat sintesis dinding sel
Dinding sel dapat memberikan kekuatan mekanik bagi bakteri itu sendiri, yaitu untuk bertahan hidup pada kondisi lingkungan yang dapat mengubah tekanan osmotiknya (Holtje, 1998). Sel bakteri dibungkus oleh lapisan peptidoglikan (murein) yang mengandung rantai asam N-acetilmuramik (MurNAc) dan residu N-asetilglukosamin (GlcNAc) yang berikatan silang melalui rantai sisi pentapeptida yang melekat pada MurNAc (Chopra et al., 2002). Gangguan sintesis atau struktur dapat menyebabkan hilangnya bentuk sel dan integritas yang diikuti oleh kematian sel (Holtje, 1998). Biosintesis peptidoglikan terjadi melalui tiga tahap. Tahap pertama melibatkan enzim yang terdapat pada sitoplasma atau dipermukaan sitoplasma untuk merakit unit monomer disakarida-peptida. Tahap kedua yaitu unit monomer ditransfer melintasi membran sitoplasma, dan tahap ketiga polimerasi unit monomer pada permukaan luar membran serta pengikatan peptidoglikan yang baru ke dinding sel yang sudah ada sebelumnya. Langkah ketiga dimanfaatkan sebagai target antibiotik melalui agen β-laktam dan glikopeptida (Park and Uehara, 2008).
β-laktam dan glikopeptida merupakan salah satu kelas antibiotik yang dapat mengganggu langkah-langkah tertentu dalam homeostasis biosintesis dinding sel. Penghambatan sintesis dinding sel ini dapat mengakibatkan perubahan bentuk sel dan ukuran, dapat menginduksi respon stres sel dan pada akhirnya sel lisis (Park and Uehara, 2008). Beberapa antibiotik yang tergolong kelas β-laktam adalah penisilin, carbapenem dan sefalosporin. Antibiotik ini dapat menghambat pembentukan ikatan peptida yang dikatalis oleh enzim PBP
(Penicillin-Binding Proteins) dengan cara memblokir ikatan silang dari unit
13 ini akan memperpanjang untaian glikan dari molekul peptidoglikan yang sudah ada dengan menambahkan pentapeptida disakarida (Park and Uehara, 2008).
D. Mekanisme antibiotik menghambat sintesis protein
Penerjemahan mRNA terjadi dalam tiga fase berurutan yaitu inisiasi, elongasi, dan terminasi yang melibatkan organel ribosom dan sitoplasma (Garrett, 2000). Organel ribosom terdiri atas dua subunit ribonukleo protein yaitu 50S dan 30S (Nissen et al., 2000). Ribosom ini dapat menjadi target antibiotik dalam menghambat sintesis protein. Antibiotik yang menghambat sintesis protein dibagi menjadi dua sub kelas yaitu 50S inhibitor dan 30S inhibitor.
Kelas antibiotik yang termasuk sebagai 50S ribosom inhibitor yaitu makrolida (misalnya, eritromisin), linkosamida (misalnya, klindamisin), streptogramin (misalnya, dalfopristin-quinupristin), amphenikol (misalnya, kloramfenikol) dan oksazolidinon (misalnya, linezolid) (Katz and Ashley, 2005). Antibiotik 50S inhibitor ini bekerja dengan menghalangi inisiasi translasi protein atau translokasi tRNA peptidil, dan hal tersebut dapat menghambat reaksi peptidiltransferase untuk memanjangkan rantai peptida yang baru. Peristiwa ini melibatkan pemblokiran akses tRNA peptidil ke ribosom untuk melakukan proses selanjutnya, kemudian pemblokiran reaksi elongasi peptidiltransferase terjadi dan akhirnya akan memicu disosiasi peptidil tRNA (Vannuffel and Cocito, 1996).
14 dari kompleks yang terbentuk antara kodon mRNA dan aminoasil tRNA di ribosom, sehingga kesalahan penerjemahan protein dapat terjadi (Pape et al., 2000).
Aminoglikosida akan menyebabkan kesalahan penerjemahan protein karena terjadi penggabungan asam amino yang tidak cocok ke dalam elongasi untaian peptida. Penerjemahan protein yang salah ini akan digabungkan ke dalam membran sitoplasma sehingga sifat permeabilitas sel menjadi lebih meningkat dan hal ini akan mempermudah akses antibiotik masuk kedalam sel. Semakin mudah antibioik masuk ke dalam sel maka penghambatan ribosom semakin meningkat dan sel bakteri akan mati (Fourmy et al., 1996).
Aminoglikosida turunan alami merupakan satu-satunya kelas antibiotik yang memiliki sifat bakterisida yang luas. Sedangkan makrolid, streptogramin, spektinomisin, tetrasiklin, kloramfenikol dan makrolida adalah antibiotik yang bersifat bakteriostatik, akan tetapi dapat bersifat bakterisida pada suatu bakteri (Fourmy et al., 1996). Hal ini didukung oleh penelitian yang dilakukan oleh Goldstein et al. (1990), bahwa kloramfenikol dan makrolida azitromisin telah menunjukan aktivitas bakterisida terhadap Haemophilus influenzae.
1.3.2. Mekanisme resistensi bakteri terhadap antibiotik
Resistensi bakteri dapat dijelaskan melalui dua cara yaitu, (1) resistensi intrinsik, dimana secara alami suatu bakteri tidak memiliki situs target untuk antibiotik sehingga antibiotik tidak berpengaruh terhadap bakteri tersebut. Bakteri dapat secara alami memiliki permeabilitas rendah terhadap antibiotik karena adanya perbedaan sifat kimia antibiotik dengan struktur membran bakteri. (2) resistensi antibiotik yang dapat diperoleh oleh bakteri, dimana mekanisme resistensi yang diperoleh ini dapat melalui berbagai cara seperti yang diungkapkan oleh Fluit et al. (2001), yaitu sebagai berikut:
a. Adanya enzim yang menginaktivasi agen antibiotik
15 c. Terjadinya mutasi pada protein target antibiotik sehingga daya berikatan
antibiotik pada bakteri rendah
d. Terjadinya modifikasi pasca transkripsi atau pasca translasi pada bakteri yang menjadi target antibiotik sehingga ini dapat mengurangi pengikatan agen antibiotik.
e. Pengurangan penyerapan zat antibiotik oleh bakteri
f. Diaktifkannya protein efluk (pompa efluk) yang akan memompa dan mencegah masuknya antibiotik ke dalam sel bakteri
g. Protein target antibiotik diproduksi berlebihan 1.4. Bakteri Acinetobacter baumannii
Acinetobacter baumannii tergolong bakteri aerobik, Gram negatif,
berbentuk batang, nonfermentasi, nonmotil, oksidase negatif, dan bakteri ini dapat tumbuh pada suhu 20 – 300C. Secara makroskopis memiliki koloni berbentuk bulat dan berwarna putih berlendir (Ajao et al., 2011) (Gambar 3). Bakteri A.
Baumannii memiliki sejarah perubahan taksonomi yaitu awalnya termasuk famili
16 Gambar 3. Isolat Acinetobacter baumannii (diameter koloni 0,5-2 mm) (Ajao et
al., 2011)
Wabah A. baumannii resisten terhadap antibiotik kelas carbapenem pertama kali terjadi di Amerika Serikat pada tahun 1991 (Go et al., 1994). Tahun 1998, Isolat A. baumannii yang resisten terhadap antibiotik carbapenem diisolasi dari pasien leukemia di rumah sakit Taiwan. Setelah diamati, ternyata isolat tersebut resisten terhadap hampir semua antibiotik (seperti; sefalosporin, aztreonam, aminoglikosida dan ciprofloxacin) (Hsueh et al., 2002). Kemampuan dalam membentuk biofilm membuat patogen ini sulit dikendalikan. Pembentukan biofilm dikendalikan oleh beberapa faktor, diantaranya adanya gen resisten antibiotik, kondisi pertumbuhan yang mendukung, dan kepadatan sel dalam mengkolonisasi (Gaddy and Actis, 2009). Seperti yang diungkapkan oleh Lee et al. (2008), bahwa kehadiran dan ekspresi salah satu gen yaitu gen blaPER-1dapat
meningkatkan kemampuan membentuk biofilm diberbagai permukaan, baik permukaan biotik maupun abiotik (Gambar 4 dan 5).
17 diperpanjang dan memungkinkan pasien memperoleh perawatan berupa makanan enteral (Mulin et al., 1995). Bahaya utama dari A. baumannii adalah kemampuannya yang dengan cepat resisten terhadap antibiotik, yang kemudian bergerak kearah multidrug resisten (Bergogne and Towner, 1996). Kejadian infeksi A. Baumannii telah menjadi masalah kesehatan di banyak negara (Landman et al., 2002).
Gambar 4. Biofilm A. baumannii (in vitro) yang terbentuk pada permukaan penutup kaca setelah 24 jam inkubasi pada suhu 370C di media
luria-bertani broth. Perbesaran 5.000 x, dengan Scanning Electron
Microscopy (SEM) (Longo et al., 2014)
Gambar 5. Biofilm polimikroba (A. baumannii, K. pnomuniae, dan P. aeruginosa) tumbuh dalam lumen kateter urin silikon yang diisolasi dari pasien rumah sakit di Roma. Perbesaran 10.000 x, dengan Scanning Electron
18 1.4.1.Mekanisme resistensi antibiotik pada Acinetobacter baumannii
Mekanisme resistensi yang umum dapat terjadi melalui resistensi yang dimediasi oleh enzim, adaptasi genetik, pengaktifan protein efluk, dan perubahan struktur komponen membran luar sel bakteri (Cloete, 2003). Enzim yang memediasi resistensi antibiotik yaitu kemampuan bakteri untuk memproduksi enzim yang dapat mengubah senyawa antibiotik menjadi senyawa yang tidak toksik atau tidak aktif bagi sel bakteri itu sendiri (Ma et al., 1998). Protein efluk befungsi untuk memompa zat antibiotik keluar dari sel, menurunkan konsentrasi antibiotik dalam sel dan mencegah antibiotik bekerja mengenai target (Nikaido, 2009). Sedangkan untuk perubahan struktur membran, ini dapat terjadi pada porin dan enzim PBP (Penicillin-Binding Proteins) (Cloete, 2003). Porin merupakan suatu protein yang diproduksi di membran luar bakteri Gram negatif, berfungsi sebagai saluran difusi nonspesifik yang memungkinkan masuknya antibiotik yang efektif (Nikaido, 2009). Perubahan struktur membran sel ini memungkinkan sel untuk meningkatkan resistensi terhadap antibiotik karena antibiotik tidak dapat menembus membran sel dan tidak mampu mencapai situs targetnya (Cloete, 2003).
19 pada A. baumannii ini merupakan target kuinolon, namun karena tidak mampunya antibiotik melekat pada situs targetnya maka A. Baumannii menjadi resisten terhadap antibiotik kuinolon. Selain itu, modifikasi lipopolisakarida dalam sel A.
baumannii juga akan menyebabkan bakteri resisten terhadap polimiksin (Perez et
al., 2007).
1.5. Minimum Inhibitory Concentration (MIC)
Minimum Inhibitory Concentration (MIC) dianggap sebagai “gold
standard” yaitu konsentrasi terendah suatu antibiotik dalam menghambat