Analisis Diversitas Genetik Aksesi Padi Warna Indonesia
Berdasarkan Marka Molekuler Fungsional
(Genetic Diversity Analysis of Indonesian Pigmented Rice Accessions
Based on Functional Molecular Markers)
Andari Risliawati1*, Yusi N. Andarini1, Rerenstradika T. Terryana1, Kristianto Nugroho1, dan Puji Lestari2 1Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumber Daya Genetik Pertanian, Jl. Tentara Pelajar 3A, Bogor 16111, Indonesia
Telp. (0251) 8337975; Faks. (0251) 8338820; *E-mail: boendar@yahoo.co.id 2Balai Besar Penelitian Tanaman Padi, Jl. Raya No. 9, Sukamandi, Subang 41256, Indonesia
Diajukan: 11 Januari 2021; Direvisi: 9 Juni 2021; Diterima: 16 Juni 2021
ABSTRACT
Pigmented rice is functional staple food that becomes popular because of its anthocyanin content which is beneficial for health. Studies on the diversity of the local variety of Indonesian pigmented rice accessions have been carried out, but are still limited to one region of germplasm origin. This study aimed to analyze the genetic diversity of local varieties of pigmented rice collections of the IAARD-ICABIOGRAD Gene Bank. A total of 93 pigmented rice accessions from 16 provinces in Indonesia were analyzed using 15 functional molecular markers of SSR, STS, and indel. The total alleles detected were 115 with an average per locus of genetic diversity value of 0.71. There were five markers with PIC values >0.75, i.e. RM167, RM223, R8M33, R10M10, and GBSS1. The accessions were divided into two main groups based on their pericarp color. It is necessary to analyze the physicochemical content of the local rice accessions to complement the existing diversity information and identify potential pigmented rice accessions with high palatability.
Keywords: Pigmented rice, genetic diversity, gene bank, accession, molecular marker. ABSTRAK
Padi warna merupakan pangan pokok fungsional yang mulai diminati masyarakat karena kandungan antosianinnya yang bermanfaat untuk kesehatan. Studi mengenai diversitas padi warna lokal Indonesia telah dilakukan, tetapi masih terbatas untuk aksesi dari satu wilayah asal plasma nutfah. Penelitian ini bertujuan menganalisis diversitas genetik satu set varietas padi warna lokal koleksi Bank Gen Pertanian Balitbangtan-BB Biogen yang berasal dari berbagai wilayah Indonesia berdasarkan marka molekuler fungsional. Sebanyak 93 aksesi padi warna yang berasal dari 16 provinsi di Indonesia dianalisis dengan menggunakan 15 marka molekuler fungsional SSR, STS, dan indel. Total 115 alel terdeteksi dengan nilai rata-rata diversitas genetik sebesar 0,71 per lokus marka. Terdapat lima marka dengan nilai PIC >0,75, yaitu RM167, RM223, R8M33, R10M10, dan GBSS1. Seluruh aksesi yang dianalisis membentuk dua kelompok utama berdasarkan warna kulit arinya. Perlu penelitian lebih lanjut mengenai kandungan fisiko-kimia beras untuk melengkapi informasi diversitas genetik yang telah ada dan mengidentifikasi aksesi padi warna lokal potensial yang memiliki karakter palatabilitas baik.
Kata kunci: Padi warna, diversitas genetik, bank gen, aksesi, marka molekuler.
PENDAHULUAN
Padi (Oryza sativa L.) merupakan komoditas utama di Indonesia dan makanan pokok sebagian besar penduduk dunia, terutama di kawasan Asia. Diversitas tanaman padi di dalam spesies padi budi daya dikenal cukup tinggi. Terdapat lima subspesies padi budi daya, yaitu padi tipe indica, japonica, aus,
aromatic, dan tropical-japonica (Garris et al. 2005).
Setiap kawasan di Asia umumnya memiliki tipe padi-nya masing-masing akibat dari pengaruh pola domestikasi tanaman dan preferensi masyarakat setempat (Khush 1997). Berdasarkan warna kulit arinya, tanaman padi mengelompok menjadi dua, yaitu padi dengan kulit ari tidak berwarna (selanjut-nya disebut padi beras putih) dan padi dengan kulit ari berwarna (selanjutnya disebut padi beras cokelat, merah, atau hitam). Sebagian besar masyarakat mengonsumsi padi beras putih. Namun, seiring dengan peningkatan pendapatan dan preferensi ke arah pola hidup yang lebih sehat, beberapa kalangan mulai beralih mengonsumsi padi warna, baik padi beras merah maupun padi beras hitam (Yuliati et al. 2012).
Munculnya berbagai warna pada kulit ari beras menunjukkan adanya pigmentasi antosianin dengan derajat yang berbeda-beda. Diversitas padi warna di tingkat global, khususnya di kawasan Asia, diketahui cukup tinggi (Gowda et al. 2012). Di Indonesia sendiri diketahui banyak varietas padi warna lokal yang ter-sebar di beberapa wilayah, seperti Sulawesi Tenggara (Suliartini et al. 2011), Daerah Istimewa Yogyakarta (Kristamtini et al. 2014), Sumatra Utara (Budi et al. 2017), dan Sumatra Selatan (Arifah 2019). Beberapa studi diversitas genetik berbasis aplikasi marka molekuler juga telah dilaporkan, tetapi sifat-nya masih terbatas untuk aksesi yang berasal dari satu provinsi, seperti padi merah dari Daerah Istimewa Yogyakarta (Kristamtini et al. 2014) dan Sumatra Selatan (Arifah 2019), yang menunjukkan hasil diversitas genetik yang sempit. Dalam rangka memperoleh informasi diversitas genetik yang lebih luas, perlu dilakukan studi diversitas genetik padi warna dari berbagai wilayah. Diversitas genetik yang luas merupakan modal dasar bagi kegiatan pemulia-an tpemulia-anampemulia-an, terutama apabila diversitas ypemulia-ang diper-oleh dari varietas lokal merupakan alel-alel yang potensial dan adaptif terhadap cekaman lingkungan tertentu (Dwivedi et al. 2016).
Marka molekuler dipilih sebagai basis analisis diversitas genetik padi warna karena dapat mende-teksi langsung diversitas pada level DNA. Selain itu, marka molekuler juga dapat memfasilitasi analisis
sampel aksesi dengan jumlah lebih banyak dan dalam waktu analisis lebih singkat dibanding dengan analisis berbasis pada karakter morfo-agronomi. Beberapa jenis marka molekuler telah diaplikasikan pada studi diversitas genetik padi warna dan juga analisis palabilitas berbagai jenis padi, seperti marka mikrosatelit atau simple sequence repeat (SSR) (Kristamtini et al. 2014), marka sequence-tagged site (STS) (Lestari et al. 2012; Lestari et al. 2015), dan marka insertion-deletion (indel) (Patel et al. 2014). Penggunaan berbagai jenis marka molekuler tersebut diharapkan dapat menghasilkan informasi yang lebih komprehensif terkait status diversitas genetik varietas padi warna lokal Indonesia yang pada akhirnya dapat mendukung program perakitan varietas padi warna nasional di masa yang akan datang. Penelitian ini bertujuan menganalisis diversitas genetik satu set varietas padi warna lokal koleksi Bank Gen Pertanian Balitbangtan-BB Biogen yang berasal dari berbagai wilayah Indonesia berdasarkan marka molekuler fungsional.
BAHAN DAN METODE
Penelitian dilaksanakan di Rumah Kaca dan Laboratorium Biologi Molekuler, Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumber Daya Genetik Pertanian (BB Biogen). Materi genetik yang digunakan pada penelitian ini terdiri atas 93 varietas padi warna lokal yang berasal dari koleksi Bank Gen Pertanian Balitbangtan-BB Biogen (Tabel 1). Sebagian besar aksesi padi warna lokal ini diakuisisi dalam kurun waktu 2012–2016 dan belum dikarakterisasi secara lengkap, baik secara morfo-agronomi maupun molekuler.
Isolasi DNA Genomik dan Uji Kualitas dan Kuantitas DNA
DNA genomik diisolasi dari daun muda yang diambil secara bulk dari 5–10 tanaman sampel dari setiap aksesi. Tanaman sampel ini ditanam pada bak semai di rumah kaca dan dipelihara sampai tanaman berumur 2–3 minggu. Isolasi DNA dilakukan dengan kit ekstraksi GeneExTM Plant Kit (GeneAll, Korea
Selatan).
Analisis kualitas DNA untuk memastikan pita DNA yang diperoleh tebal dan tidak smear dilakukan dengan teknik elektroforesis pada gel agarosa 1%. Analisis kuantitas DNA untuk mengetahui tingkat kemurnian DNA dilakukan dengan
NanoDrop®ND-100 Spectrophotometer (Thermo Fisher Scientific, AS)
menggunakan blanko bufer RE pada panjang gelom-bang A260 dan A280. Hasil kuantitas DNA yang
diper-oleh dijadikan dasar perhitungan pengenceran DNA untuk keperluan analisis PCR.
Marka Molekuler yang Digunakan untuk Analisis Diversitas Genetik Aksesi Padi Warna Indonesia
Sebanyak 15 marka molekuler digunakan untuk menganalisis diversitas genetik padi warna lokal Indonesia, yang terdiri atas 6 marka SSR, 3 marka STS, dan 6 marka indel (Tabel 2). Marka-marka molekuler tersebut dipilih berdasarkan tingkat polimorfismenya pada studi diversitas genetik padi warna yang dilakukan oleh Patel et al. (2014) dan
studi palatabilitas padi yang dilakukan oleh Lestari et al. (2009).
Analisis PCR, Elektroforesis, dan Skoring Data
Analisis PCR untuk mengamplifikasi marka target dilakukan dengan mesin PCR Thermocycler (Biometra, Jerman). Komposisi DNA dan reagen yang digunakan, yaitu 1 µl DNA (10 ng/µl), 1 µl primer F+R (10 µM), 5 µl 2× MyTaq™ HS Red Mix (Bioline, UK), dan 3 µl ddH2O steril. Program PCR yang digunakan,
yaitu predenaturasi (94°C, 4 menit), diikuti dengan 35 siklus denaturasi (94°C, 1 menit), penempelan primer
Tabel 1. Karakteristik 93 aksesi padi warna lokal Indonesia yang digunakan pada penelitian ini.
Kode No. aksesi Nama aksesi Warna
kulit ari Provinsi asal Kode No. aksesi Nama aksesi
Warna
kulit ari Provinsi asal
1Jm 2012_156 Tongbulan Merah Jawa Timur 49Jm 2012_131 Padi Biji Besar Merah Jawa Timur 2Jm 2012_157 Merah Merah Jawa Timur 50Jg 2016_304 Tjempo Merah Merah Jawa Tengah 3Jm 2012_130 Padi Merah Merah Jawa Timur 51Jg 2016_301 Abang Merah Jawa Tengah 4Jm 2012_143 Padi Merah Merah Jawa Timur 52Jg 2016_310 Merah Merah Jawa Tengah 5Jg 2012_174 Cempo Hitam Genjah Hitam Jawa Tengah 54SW 2016_506 Pulo Merah Merah Sulawesi Utara 6Jm 2012_154 Padi Merah Merah Jawa Timur 56Jb 2014_7 Marus Ekek Merah Jawa Barat 7Jm 2012_159 Renggerengan Merah Jawa Timur 57Nb 2016_330 Padi Merah Dompu Merah NTB
8Jm 2012_139 Bulu Kuning Merah Jawa Timur 58SW 2016_502 Pulo Siam Merah Sulawesi Utara 9Jm 2012_155 Pondong Merah Jawa Timur 59Jb 2013_181 Nandi Merah Jawa Barat 10Jm 2012_136 Padi Merah Merah Jawa Timur 60Jm 2012_153 Ketan Hitam Hitam Jawa Timur 11Jm 2012_152 Padi Hitam/Bulu Celang Hitam Jawa Timur 61K 2015_23 Padi Hitam Hitam Kalimantan Selatan 12Jm 2012_126 Padi Merah Merah Jawa Timur 62Jy 05020-05179 Melik Hitam DIY
13Jb 2013_158 Ketan Hitam Hitam Jawa Barat 63Jb 05020-05519 Omas Merah Jawa Barat 14Jb 2013_156 Gadog Beureum Merah Jawa Barat 64SM 05020-06323 Sawah Darek Merah Sumatra Barat 15Jb 2013_161 Pare Hitam Hitam Jawa Barat 65SM 05020-08154 Kopai Merah Aceh 16Jb 2013_155 Ketan Nangka Merah Banten 66SM 05020-08155 Muda Baru Merah Aceh
17Jb 2013_151 Ketan Hideung Hitam Banten 67SW 05020-10221 Pulut Bolong Hitam Sulawesi Selatan 18Jb 2013_144 Cere Merah Merah Banten 68SM 05020-20709 Ramos Merah Merah Sumatra Utara 19Jb 2013_182 Hangasa Merah Banten 69K 05020-21237 Pae Rabi Hitam Kalimantan Barat 20Jb 2014_2 Badigal Merah Jawa Barat 70UK 05020-21658 Segreng Merah Tidak diketahui 21SW 2016_4 Pare Tumbu’ Padang Merah Sulawesi Barat 71UK 05020-21669 Beras Hitam Hitam Tidak diketahui 22SW 2016_31 Pare Bae’ Merah Merah Sulawesi Barat 72UK 05020-21684 Padi Bintik Merah Tidak diketahui 24SW 2016_3 Pare Lesuma Merah Sulawesi Barat 73UK 05020-21685 Padi Merah A Merah Tidak diketahui 25SM 2016_192 Rotan-6 Merah Sumatra Barat 74UK 05020-21719 Bulu Merah Merah Tidak diketahui 26SM 2016_191 Guliang Merah Sumatra Barat 75UK 05020-21722 Dodokan Abang Merah Tidak diketahui 27SM 2016_193 Rotan-7 Merah Sumatra Barat 76UK 05020-21734 Loyor Merah Tidak diketahui 28SM 2016_146 Suren Merah Merah Jambi 77UK 05020-21735 Nangka Merah Tidak diketahui 29Jg 2012_169 Cempo Merah Merah Jawa Tengah 78UK 05020-21736 Janti Merah Tidak diketahui 30Jb 2013_169 Terong Merah Banten 79UK 05020-21750 Caruluk Beureum Merah Tidak diketahui 31Jb 2013_183 Carogol Merah Banten 80UK 05020-21751 Mancrit Beureum Merah Tidak diketahui 32Jb 2013_171 Loyor Merah Banten 81UK 05020-21771 Ketan Hideung Hitam Tidak diketahui 33Jb 2013_166 Badigal Merah Banten 82UK 05020-21772 Ketan Hideung Hitam Tidak diketahui 34Jb 2013_150 Ketan Hideung Hitam Banten 83UK 05020-21786 Wulung Hitam Tidak diketahui 35SM 2015_40 Ceredek Merah Merah Sumut 84N 2017_1 Woja Langko Hitam NTT
36Jg 2016_292 Kretek Merah Merah Jawa Tengah 85N 2017_2 Woja Rakot Merah NTT 37N 2016_356 Padi Hitam Hitam NTT 86N 2017_3 Laka Galung Merah NTT 38Jm 2012_129 Padi Bintik Merah Jawa Timur 87N 2017_4 Tunas Merah Merah NTT 39N 2016_355 Padi Merah Kecil Merah NTT 88N 2017_5 Lea Merah NTT 40SM 2016_45 Siangkat Merah Sumatra Utara 89N 2017_6 Woja Bogor Merah NTT 41Jy 2016_288 Pari Ireng Hitam DIY 90N 2017_7 Laka Hitam NTT 42Jy 2016_286 Melik Hitam DIY 91N 2017_8 Woja Ndua Merah NTT 43Jy 2016_274 Tangkilan Merah DIY 92N 2017_9 Kosu Ampera Merah NTT 44Jy 2016_278 Andel Hitam Hitam DIY 93N 2017_10 Kosu Mite Hitam NTT 45Jy 2016_280 Merah Mujiono Merah DIY 94N 2017_11 Kosu Toro Merah NTT 46Jy 2016_276 Mariti Merah Merah DIY 95N 2017_12 Woja Laka Rakot Hitam NTT 47Jg 2016_317 Ketan Hitam Hitam Jawa Tengah 96N 2017_13 Woja Laka Hitam NTT 48Jy 2016_272 Mandel Merah DIY
(52–62°C, 1 menit), dan pemanjangan basa (72°C, 2 menit). Pemanjangan primer akhir dilakukan pada 72°C selama 7 menit. Produk amplifikasi dielektrofo-resis pada gel poliakrilamida 6% di dalam tangki elektroforesis vertikal berisi bufer TBE 1× yang di-jalankan pada tegangan 80 volt selama 90 menit. Ukuran produk PCR diestimasi dengan mengguna-kan DNA ladder 100 bp. Hasil elektroforesis divisuali-sasi dengan menggunakan UV pada ChemiDoc™ XRS
System (Bio-Rad, AS).
Pita DNA hasil elektroforesis diberi skor dengan bantuan program GelAnalyzer (Lazar dan Lazar 2010) untuk mendapatkan ukuran alel dalam bentuk angka numerik. Ukuran alel yang diperoleh kemudian dike-lompokkan berdasarkan hasil perhitungan besaran interval basa untuk setiap kelompok yang terbentuk pada setiap marka. Alel-alel pada marka tertentu yang berada pada kelompok yang sama akan mem-peroleh nomor kode (coding) yang sama. Angka hasil pengodean ini kemudian digunakan untuk tahap analisis selanjutnya. Adapun perhitungan besaran interval basa pada setiap marka dilakukan berdasar-kan teknik distribusi frekuensi numerik sebagai berikut:
P = R = max – min
K 1 + 3,3 log n
dengan P = panjang kelas atau besaran interval basa,
R = nilai rentang ukuran alel, max = ukuran alel
tertinggi, min = ukuran alel terendah, K = jumlah kelompok, dan n = jumlah populasi.
Analisis Data
Analisis data meliputi analisis profil materi gene-tik yang digunakan, dipaparkan secara deskriptif, meliputi komposisi daerah asal dan persentase jenis padi warna yang dianalisis, dan analisis profil marka molekuler yang dilakukan dengan menggunakan program PowerMarker v.3.25 (Liu dan Muse 2005). Karakteristik marka yang diamati antara lain jumlah alel terdeteksi, diversitas gen, dan nilai polymorphism
information content (PIC). Frekuensi alel minor
di-hitung berdasarkan ukuran alel yang dihasilkan. Alel minor merupakan alel yang kemunculannya kurang dari 5% pada populasi (Panagiotou et al. 2010), yaitu antaraksesi padi warna yang diuji.
Analisis pengelompokan aksesi dilakukan dengan menggunakan program Dissimilarity Analysis
and Representation for Windows (DARWin) v.6.0.13 Tabel 2. Lima belas marka molekuler fungsional dan karakteristiknya yang digunakan pada penelitian ini.
Marka Jenis
marka Kromosom Tm (°C)
Kisaran produk
(bp) Sekuen primer forward (F) 3’ 5’ dan reverse (R) 5’ 3’ Referensi
RM166 SSR 2 61,0 155–355 F: GGTCCTGGGTCAATAATTGGGTTACC R: TTGCTGCATGATCCTAAACCGG Patel et al. (2014) RM167 SSR 11 61,3 195–415 F: GGTCCTGGGTCAATAATTGGGTTACC R: TTGCTGCATGATCCTAAACCGG Patel et al. (2014) RM206 SSR 11 54,9 115–305 F: CCCATGCGTTTAACTATTCT R: CGTTCCATCGATCCGTATGG Patel et al. (2014) RM223 SSR 8 55,0 105–210 F: GAGTGAGCTTGGGCTGAAAC R: GAAGGCAAGTCTTGGCACTG Patel et al. (2014) R8M33 Indel 8 52,6 145–225 F: CGAAAGAGGAGAGGGGTAGT R: CGAAAACGAGAAACAAATA Patel et al. (2014) R8M46 Indel 9 54,5 75–120 F: CAGCAGAGTCCAGAGAAGAT R: GCATAAGATGGCGAGTGA Patel et al. (2014) R10M10 Indel 7 55,4 110–180 F: GAATACAACCCCCTAAAAAC R: ATGGACCGTTGAGGAGAC Patel et al. (2014) R10M30 Indel 6 51,9 105–155 F: CCCTAAAAATAGAGCAACCT R: ACCCATAATACTACCAATCAAC Patel et al. (2014) Aglu3 STS 1 55,7 210–260 F: CCTCTGGAATCTTGCTATTTAGG R: ATCCGCTAGATCACTGACAAA Kobayashi et al. (2008) GBSS1 SSR 6 57,3 135–185 F: CAAATAGCCACCCACACCAC R: CTTGCAGATGTTCTTCCTGATG Lestari et al. (2009) Isa STS 8 61,1 200–295 F: CCTGTCTTGCACGTGCGGTA R: GCACGGTTCTGATGTACGAGAG He et al. (2006) Pul3 STS 4 58,9 255–290 F: GGGTTCGCTTTCACAACACAG R: GTCACGACATAAGAGAAGCTGC He et al. (2006) SBE1 SSR 6 57,5 135–160 F: ATTTCTTTGGCCACAGGCGA R: CCCAGATTCGGAACAAGAAC Bao et al. (2006)
S3cI Indel 7 53,3 125–150 F: CCACTCTCATGTCCTTGAAC R: GCCATGACATTTGGACAT
Lestari et al. (2009)
TreB Indel 7 55,1 145–170 F: CACTCCAGTTCCTGCTCAAA R: CACCTCCAAAACGAATATGG
Lestari et al. (2009)
(Perrier dan Jacquemoud-Collect 2006). Metode pengelompokan yang digunakan adalah weighted
neighbour joining dengan matriks ketidaksamaan
(dissimilarity matrix) yang dihitung melalui analisis
bootstrap 1.000 iterasi.
HASIL DAN PEMBAHASAN Profil Populasi Genotipe Uji
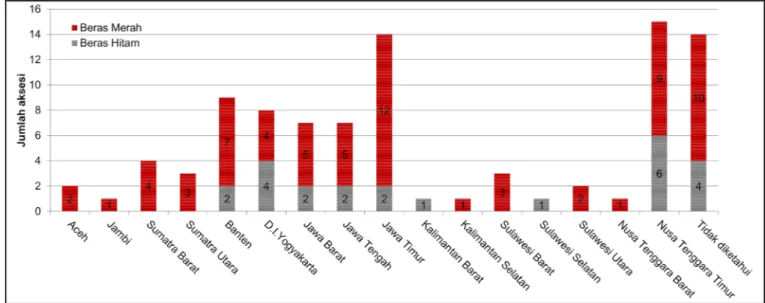
Seluruh aksesi padi warna yang digunakan pada penelitian ini merupakan varietas lokal yang berasal dari 16 provinsi di Indonesia, tersebar dari wilayah barat sampai timur, dengan lokasi asal aksesi ter-banyak adalah Jawa Timur dan Nusa Tenggara Timur (Gambar 1). Sebagian besar aksesi ini (77%) merupa-kan hasil eksplorasi intensif yang dilakumerupa-kan oleh BB Biogen dan Balai Pengkajian Teknologi Pertanian (BPTP) dalam kurun waktu 2012–2017 (Hidayatun et al. 2017). Hanya sekitar 23% aksesi yang merupakan koleksi yang telah lama dikonservasi oleh Bank Gen Pertanian Balitbangtan-BB Biogen.
Pada studi diversitas genetik, pelibatan sumber daya genetik (SDG) dalam bentuk varietas lokal yang berasal dari berbagai wilayah dapat memberikan informasi tentang hubungan kekerabatan antaraksesi dari kelompok wilayah yang berbeda serta pola domestikasi dan penyebaran tanaman (Chen dan Nelson 2005). Selain itu, analisis diversitas genetik yang menggunakan SDG yang berasal dari berbagai wilayah akan dapat menghasilkan koleksi inti (core
collection) yang lebih komprehensif dan informatif
(Mahalakshmi et al. 2007; Agrama et al. 2009; Risliawati et al. 2018).
Pada penelitian ini, padi beras merah (74,19%) lebih banyak dianalisis dibanding dengan padi beras hitam (Gambar 1). Hal ini dikarenakan jenis varietas lokal padi beras hitam telah jarang dibudidayakan oleh petani sehingga keberadaannya sulit ditemukan ketika eksplorasi SDG dilakukan (Kristamtini et al. 2014; Rohaeni dan Hastini 2015). Oleh karenanya, jumlah padi beras hitam yang dapat dikoleksi oleh Bank Gen Pertanian Balitbangtan-BB Biogen dari kegiatan eksplorasi tersebut juga tidak cukup banyak. Ekplorasi SDG sejatinya merupakan upaya untuk menyelamatkan SDG dengan cara mengonservasinya secara ex situ agar terhindar dari erosi genetik sehingga dapat terus dimanfaatkan sebagai sumber gen bagi kegiatan pemuliaan tanaman (Afza 2016).
Profil Marka Molekuler
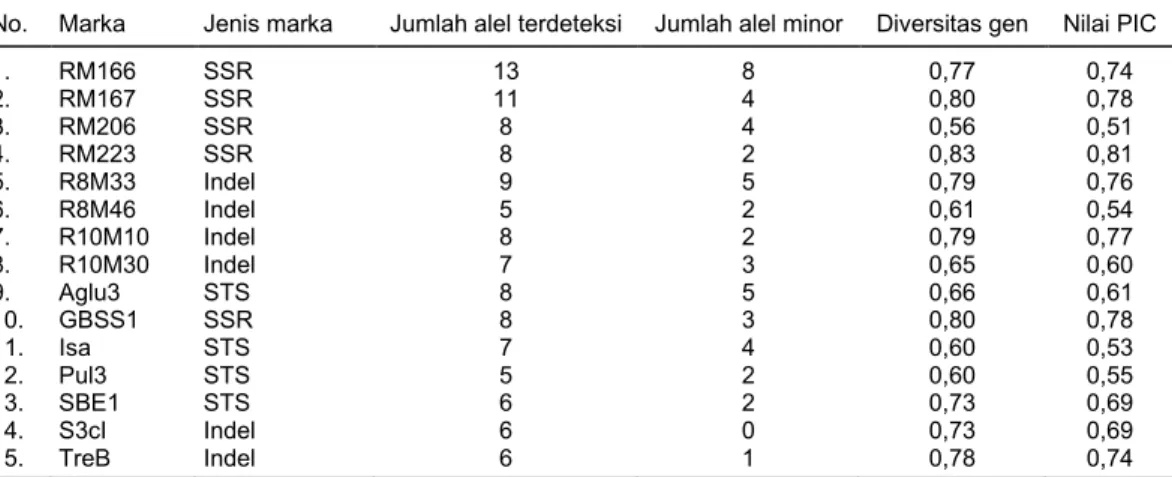
Sebanyak 115 alel terdeteksi oleh 15 marka fungsional yang digunakan, dengan kisaran 5–13 alel per lokus (Tabel 3). Di antara marka molekuler yang digunakan, marka RM166 merupakan marka mole-kuler yang menghasilkan jumlah alel terbanyak (13 alel), sedangkan marka R8M46 dan Pul3 menghasil-kan jumlah alel paling sedikit (5 alel). Jumlah alel yang diperoleh pada penelitian ini lebih banyak di-banding dengan penelitian 19 genotipe padi warna asal India yang menggunakan marka SSR yang sama (Patel et al. 2014) ataupun penelitian 11 genotipe padi warna asal Yogyakarta yang dianalisis dengan marka SSR yang berbeda (Kristamtini et al. 2014). Hal ini dapat disebabkan oleh pelibatan jumlah genotipe yang lebih banyak dan beragam daerah asalnya sehingga variasi alel yang ditemukan lebih banyak.
Selain itu, dapat juga berarti bahwa penggunaan marka SSR spesifik warna beras pada penelitian ini mampu mengindentifikasi perbedaan alel antaraksesi yang dianalisis dengan cukup baik.
Pada umumnya, marka molekuler yang dipilih untuk digunakan pada studi diversitas genetik atau studi pengembangan set marka untuk identifikasi penciri varietas adalah marka yang mempunyai nilai PIC yang tinggi, yaitu di atas 0,75 (Risliawati et al. 2015). Nilai PIC merupakan cerminan dari kemampu-an marka molekuler dalam menghasilkkemampu-an alel ykemampu-ang polimorfis (Gale et al. 2005). Dari 15 marka molekuler yang digunakan, terdapat lima marka yang memiliki nilai PIC >0,75, yaitu RM167, RM223, R8M33, R10M10, dan GBSS1. Nilai PIC marka-marka ini relatif sama dengan hasil penelitian sebelumnya (Patel et al.
2014). Marka-marka polimorfis ini menunjukkan variasi pola ukuran alel yang dihasilkannya, yaitu 3 jenis variasi alel pada marka R8M33 dan 2 jenis variasi alel pada marka GBSS1 (Gambar 2).
Nilai rata-rata diversitas genetik yang diperoleh pada studi ini adalah 0,71 yang berarti bahwa tingkat diversitas genetik SDG padi warna yang dianalisis cukup tinggi, serupa dengan penelitian Patel et al. (2014). Sementara, pada penelitian Kristamtini et al. (2014) yang hanya menganalisis padi beras warna asal Yogyakarta tidak menunjukkan hasil yang tinggi. Hal ini menandakan bahwa latar belakang genetik padi warna lokal yang berasal dari berbagai wilayah di Indonesia lebih luas. Padi warna lokal yang beradaptasi dengan baik pada suatu wilayah tertentu selama puluhan tahun umumnya telah membentuk
Tabel 3. Karakteristik marka molekuler hasil analisis menggunakan 93 aksesi padi warna lokal Indonesia.
No. Marka Jenis marka Jumlah alel terdeteksi Jumlah alel minor Diversitas gen Nilai PIC
1. RM166 SSR 13 8 0,77 0,74 2. RM167 SSR 11 4 0,80 0,78 3. RM206 SSR 8 4 0,56 0,51 4. RM223 SSR 8 2 0,83 0,81 5. R8M33 Indel 9 5 0,79 0,76 6. R8M46 Indel 5 2 0,61 0,54 7. R10M10 Indel 8 2 0,79 0,77 8. R10M30 Indel 7 3 0,65 0,60 9. Aglu3 STS 8 5 0,66 0,61 10. GBSS1 SSR 8 3 0,80 0,78 11. Isa STS 7 4 0,60 0,53 12. Pul3 STS 5 2 0,60 0,55 13. SBE1 STS 6 2 0,73 0,69 14. S3cI Indel 6 0 0,73 0,69 15. TreB Indel 6 1 0,78 0,74
SSR = simple sequence repeat, Indel = insertion-deletion, STS = sequence-tagged site, PIC = polymorphism
information content.
Gambar 2. Variasi alel beberapa aksesi padi warna lokal Indonesia hasil analisis menggunakan marka insertion-deletion (indel) dan simple sequence repeat (SSR). (A) Hasil amplifikasi menggunakan marka indel
R8M33. (B) Hasil amplifikasi menggunakan marka SSR GBSS1. Marker = DNA ladder 100 bp. 400 bp A 300 bp 200 bp 100 bp 200 bp 100 bp B
ciri khas genetiknya sendiri, dikenal baik oleh ma-syarakat setempat, dan minim intervensi perbaikan genetik (Villa et al. 2005) sehingga variasi diversitas varietas lokal antardaerah cukup tinggi.
Analisis Klaster Aksesi Padi Warna Lokal Indonesia
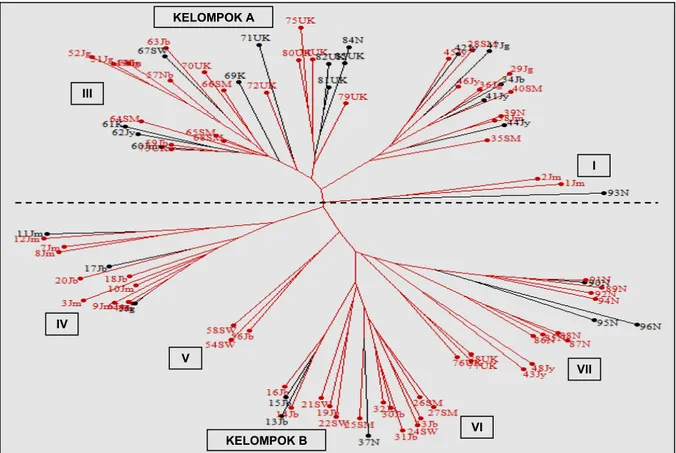
Analisis klaster 93 aksesi padi warna lokal Indonesia berdasarkan 15 marka molekuler spesifik karakter warna dan palatabilitas beras menghasilkan dua kelompok utama yang dipisahkan oleh titik per-cabangan utama, yaitu kelompok A dan kelompok B (Gambar 3). Kelompok A terdiri atas klaster I, II, dan III, sedangkan kelompok B terdiri atas klaster IV, V, VI, dan VII (Gambar 3). Pemisahan kedua kelompok utama ini cenderung berdasarkan karakter warna kulit ari beras, meskipun tidak seluruh aksesi terpisah dengan baik berdasarkan warna kulit arinya. Hal ini mungkin terjadi karena penggunaan secara bersama-sama antara marka fungsional spesifik karakter warna beras dan spesifik karakter palatabilitas.
Berdasarkan wilayah asalnya, tidak seluruh aksesi terpisah dengan baik, meskipun untuk
aksesi-aksesi yang berasal dari Nusa Tenggara Timur sebagian besar mengelompok pada klaster VII (Gambar 3). Hal ini mungkin terjadi karena marka yang digunakan adalah marka fungsional yang tidak bersifat universal atau tersebar merata di seluruh kromosom padi. Pada umumnya, studi diversitas genetik yang berhasil mengelompokkan genotipe berdasarkan daerah asalnya adalah marka yang bersifat universal seperti yang dilaporkan oleh Ali et al. (2011). Pada penelitian tersebut, diversitas padi lokal yang berasal dari 79 negara dianalisis dengan menggunakan 23 marka SSR universal yang dipilih dari 50 marka SSR standar rekomendasi oleh
Generation Challenge Program dari Consultative
Group on International Agricultural Research
(CGIAR).
Pada penelitian ini, belum dapat diidentifikasi aksesi padi warna lokal yang memiliki karakter palatabilitas baik, meskipun beberapa marka fungsional spesifik karakter telah digunakan. Oleh karenanya, perlu dilakukan analisis palatabilitas beras, seperti kandungan amilosa dan protein serta sifat fisik beras terkait viskositasnya, pada aksesi padi
Gambar 3. Dendrogram 93 aksesi padi warna lokal Indonesia berdasarkan hasil analisis menggunakan 15 marka molekuler spesifik karakter
warna dan palatabilitas beras. Garis warna merah menunjukkan aksesi dengan warna kulit ari merah dan garis warna hitam menunjukkan aksesi dengan warna kulit ari hitam. Kode huruf di belakang angka menunjukkan pulau/provinsi asal. SM = Sumatra, Jb = Banten dan Jawa Barat, Jg = Jawa Tengah, Jy = Daerah Istimewa Yogyakarta, Jm = Jawa Timur, K = Kalimantan, SW = Sulawesi, N = Nusa Tenggara Timur, Nb = Nusa Tenggara Barat.
KELOMPOK A III I IV V VI VII KELOMPOK B
warna lokal yang diuji. Dengan demikian, studi asosiasi antara data fenotipe dan genotipe terkait mutu beras dapat dilakukan, dan aksesi lokal potensial sebagai donor dalam program pemuliaan tanaman dapat diidentifikasi (Lestari et al. 2012).
Penggunaan lebih banyak marka molekuler yang polimorfis juga perlu dilakukan untuk mem-peroleh hasil pengelompokan aksesi yang lebih baik. Hal ini tidak hanya akan meningkatkan akurasi studi diversitas genetik, tetapi juga dapat menjadi dasar pengembangan set marka molekuler berbasis formula regresi dan analisis komponen utama untuk memprediksi karakter palatabilitas beras (Lestari et al. 2009; Lestari et al. 2014).
KESIMPULAN
Diperoleh tingkat diversitas genetik yang tinggi (0,71) di antara aksesi-aksesi padi warna lokal yang berasal dari berbagai provinsi di Indonesia hasil ana-lisis menggunakan 15 marka molekuler fungsional mutu beras. Analisis klaster membagi 93 aksesi padi warna lokal Indonesia menjadi dua kelompok utama berdasarkan warna kulit arinya. Setiap kelompok ter-diri atas tiga atau empat klaster. Perlu dilakukan analisis lanjutan untuk karakter fisiko-kimia beras sebagai pelengkap hasil penelitian ini. Dengan demikian, dapat diidentifikasi varietas padi warna lokal yang memiliki karakter palatabilitas baik seba-gai basis program pemuliaan padi warna Indonesia ke depan.
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Dr. Ir. Sutoro, M.S. atas ide dan bimbingannya dalam penulisan naskah ini dan kepada Meddy Saputra atas bantuan teknis laboratorium. Penelitian ini didanai melalui dana penelitian kompetitif KP4S, Balitbangtan, Kementan, 2017 dengan nomor kontrak 90.32/HM.230/H.1/05/2017.K.
KONTRIBUTOR PENULISAN
AR: kontributor utama, mengumpulkan data, menganalisis data, dan menulis naskah. YNA: kontributor anggota, menyiapkan materi genetik. RTT: kontributor anggota, menganalisis data. KN: kontributor anggota, mengumpulkan data. PL: kontributor anggota, ide penelitian, dan pemilihan marka molekuler.
DAFTAR PUSTAKA
Afza, H. (2016) Peran konservasi dan karakterisasi plasma nutfah padi beras merah dalam pemuliaan tanaman.
Jurnal Litbang Pertanian. [Online] 35 (3), 127–131.
Tersedia pada: https://doi.org/10.21082/ jp3.v35n3.2016.p143-153 [Diakses 15 Oktober 2020]. Agrama, H.A., Yan, W., Lee, F., Fjellstrom, R., Chen, M.H.,
Jia, M. & McClung, A. (2009) Genetic assessment of a mini-core subset developed from the USDA rice genebank. Crop Science. [Online] 49 (4), 1336–1346. Tersedia pada: https://doi.org/10.2135/ cropsci2008.06.0551 [Diakses 15 Oktober 2020]. Ali, M.L., McClung, A.M., Jia, M.H., Kimball, J.A., McCouch,
S.R. & Elizenga, G.C. (2011) A rice diversity panel evaluated for genetic and agro-morphological diversity between subpopulations and its geographic distribution. Crop Science. [Online] 51, 2021–2035. Tersedia pada: https://doi.org/10.2135/ cropsci2010.11.0641 [Diakses 15 Oktober 2020]. Arifah, N. (2019) Keragaman genetik padi beras merah lokal
Sumatera Selatan menggunakan marka molekuler pengkode protein pigmen warna proanthocyanidin.
Skripsi S1, Universitas Sriwijaya.
Bao, J.S., Corke, H. & Sun, M. (2006) Microsatellites, a single nucleotide polymorphisms and a sequence tagged site in starch-synthesizing genes in relation to starch physicochemical properties in non-waxy rice (Oryza sativa L.). Theoretical Applied Genetics, 113, 1185–1196.
Budi, R.S., Suliansyah, I., Yusniwati & Sobrizal (2019) Perbaikan genetik padi gogo beras merah Sumatera Utara melalui pemuliaan mutasi. Jurnal Ilmiah Aplikasi
Isotop dan Radiasi, 15 (1), 45–56.
Chen, Y. & Nelson, R.L. (2005) Relationship between origin and genetic diversity in Chinese soybean germplasm.
Crop Science. [Online] 45 (4), 1645–1652. Tersedia
pada: https://doi.org.10.2135/cropsci2004.0071 [Diakses 15 Oktober 2020].
Dwivedi, S.L., Ceccarelli, S., Blair, M.W., Upadhyaya, H.D., Are, A.K. & Ortiz, R. (2016) Landrace germplasm for improving yield and abiotic stress adaptation. Trends in
Plant Science. [Online] 21 (1), 31–41. Tersedia pada:
https://doi.org.10.1016/j.tplants.2015.10.012 [Diakses 24 Mei 2021].
Gale, K., Jiang, H. & Westcott, M. (2005) An optimization method for the identification of minimal sets of discriminating gene markers: Application to cultivar identification in wheat. Journal of Bioinformatics and
Computational Biology, 3 (2), 1–11.
Garris, A.J., Tai, T.H., Coburn, J., Kresovich, S. & McCouch, S. (2005) Genetic structure and diversity in Oryza
sativa L. Genetics. [Online] 169 (3), 1631–1638.
Tersedia pada: https://doi.org.10.1534/ genetics.104.035642 [Diakses 7 Januari 2021].
Gowda, S.J.M., Randhawa, G.J., Bisht, I.S., Firke, P.K., Singh, A.K., Abraham, Z. & Dhillon, B.S. (2012) Morpho-agronomic and simple sequence repeat-based diversity in colored rice (Oryza sativa L.) germplasm from peninsular India. Genetic Resources and Crop
Evolution. [Online] 59 (2), 179–189. Tersedia pada:
https://doi.org.10.1007/s10722-011-9674-9 [Diakses 8 Januari 2021].
He, Y., Han, Y.P., Jiang, L., Xu, C.W., Lu, J.F. & Xu, M.L. (2006) Functional analysis of starch-synthesis genes in determining rice eating and cooking qualities.
Molecular Breeding, 18, 277–290.
Hidayatun, N., Risliawati, A., Andarini Y.N. & Minantyorini (2017) Akuisisi sumber daya genetik di Bank Gen BB Biogen. Dalam: Prosiding Seminar Nasional Sumber
Daya Genetik. hlm. 66–75.
Khush, G.S. (1997) Origin, dispersal, cultivation and variation of rice. Plant Molecular Biology, 35, 25–34. Kobayashi, A., Tomita, K., Yu, F., Takeuchi, Y. & Yano, M.
(2008) Verification of quantitative trait locus for stickiness of cooked rice and amylose content by developing near-isogenic lines. Breeding Science, 58, 235–242.
Kristamtini, K., Taryono, Basunanda, P. & Murti, R.H. (2014) Keragaman genetik kultivar padi beras hitam lokal berdasarkan penanda mikrosatelit. Jurnal AgroBiogen. [Online] 10 (2), 69. Tersedia pada: https:// doi.org.10.21082/jbio.v10n2.2014.p69-76 [Diakses 15 Oktober 2020].
Lazar, I. Jr. & Lazar, I. Jr. (2010) GelAnalyzer 19.1. [Online] Tersedia pada: www.gelanalyzer.com [Diakses 15 Oktober 2020].
Lestari, P., Ham, T.H., Lee, H.H., Woo, M.O., Jiang, W., Chu, S.H., Kwon, S.W., Ma, K., Lee, J.H.I., Cho, Y.C. et al. (2009) PCR marker-based evaluation of the eating quality of japonica rice (Oryza sativa L.). Journal
of Agricultural and Food Chemistry. [Online] 57 (7),
2754–2762. Tersedia pada: https://doi.org.10.1021/ jf803804k [Diakses 5 Oktober 2020].
Lestari, P. Jiang, W., Chu, S.H., Reflinur, Sutrisno, Kusbiantoro, B., Kim, B., Piao, R., Cho, Y.C., Luo, Z. et al. (2015) DNA markers for eating quality of indica rice in Indonesia. Plant Breeding. [Online] 134 (1), 40–48. Tersedia pada: https://doi.org.10.1111/pbr.12234 [Diakses 5 Oktober 2020].
Lestari, P., Reflinur & Koh, H. (2014) Prediction of physicochemical properties of Indonesian indica rice using molecular markers. HAYATI Journal of Biosciences. [Online] 21 (2), 76–86. Tersedia pada:
https://doi.org.10.4308/hjb.21.2.76 [Diakses 5 Oktober 2020].
Lestari, P., Risliawati, A. & Koh, H.J. (2012) Identifikasi dan aplikasi marka berbasis PCR untuk identifikasi varietas padi dengan palatabilitas tinggi. Jurnal AgroBiogen, 8 (2), 69–77.
Liu, K. & Muse, S.V. (2005) PowerMarker: An integrated analysis environment for genetic marker analysis.
Bioinformatics. [Online] 21 (9), 2128–2129. Tersedia
pada: https://doi.org.10.1093/bioinformatics/bti282 [Diakses 14 Agustus 2020].
Mahalakshmi, V., Ng, Q., Lawson, M. & Ortiz, R. (2007) Cowpea (Vigna unguiculata [L.] Walp.) core collection defined by geographical, agronomical and botanical descriptors. Plant Genetic Resources: Characterisation
and Utilisation. [Online] 5 (3), 113–119. Tersedia pada:
https://doi.org.10.1017/S1479262107837166 [Diakses 15 Oktober 2020].
Panagiotou, O.A., Evangelou, E. & Ioannidis J.P.A. (2010) Genome-wide significant associations for variants with minor allele frequency of 5% or less-an overview: A huge review. American Journal of Epidemiology. [Online] 172 (8), 869–889. Tersedia pada: https:// doi.org.10.1093/aje/kwq234 [Diakses 15 Oktober 2020].
Patel, S., Ravikiran, R., Chakraborty, S., Macwana, S., Sasidharan, N., Trivedi, R. & Aher, B. (2014) Genetic diversity analysis of colored and white rice genotypes using microsatellite (SSR) and insertion-deletion (INDEL) markers. Emirates Journal of Food and
Agriculture [Online] 26 (6), 497–507. Tersedia pada:
https://doi.org.10.9755/ejfa.v26i6.15722 [Diakses 15 Oktober 2020].
Perrier, X. & Jacquemoud-Collect, J. (2006) DARWin
software. [Online] Tersedia pada: http://darwin.cirad.fr
[Diakses 15 Oktober 2020].
Risliawati, A., Kurniawan, H., Sohan, M. & Hidayatun, N. (2018) Development of sorghum core collection based on phenotypic traits in ICABIOGRAD genebank. Dalam: Wahyu, Y., Wirnas, D., Trikoesoemaningtyas, Ritonga, A.W. & Marwiyah, S. (editor) Proceeding of
PERIPI-2017 International Seminar: Sustainable Use of Genetic Resources for Quality and Productivity Improvement. Bogor, Indonesia October 2nd, 2017.
hlm. 70–77.
Risliawati, A., Riyanti, E.I., Lestari, P., Utami, D.W. & Silitonga, T.S. (2015) Development of SSR marker set to identify fourty two Indonesian soybean varieties.
Jurnal AgroBiogen, 11 (2), 49–58.
Rohaeni, W. & Hastini, T. (2015) Inventarisasi padi lokal di Kawasan Ciater, Subang, Provinsi Jawa Barat. Dalam: Setyawan, A., Sugiyarto, Pitoyo, A., Hernawan, U., Widiastuti, A., Sutarno, Saharjo, B., Supriatna, J., Jamil, A. & Djufri (editor) Prosiding Seminar Nasional
Masyarakat Biodiversitas Indonesia. Depok, 20 Desember 2014. Vol. 1. hlm. 189–193.
Suliartini, N.W.S., Sudimantara, G.R., Wijayanto, T. & Muhidin (2011) Pengujian kadar antosianin padi gogo beras merah hasil koleksi plasma nutfah Sulawesi Tenggara. Crop Agro, 4 (2), 43–48.
Villa, T.C.C., Maxted, N., Scholten, M. & Ford-Lloyd, B. (2005) Defining and identifying crop landraces. Plant
Genetic Resources, 3 (3), 373–384.
Yuliati, L.N., Retnaningsih, R. & Aprilia, D. (2012) Pengaruh kelompok acuan terhadap kesadaran dan konsumsi beras merah (Oryza nivara). Jurnal Ilmu Keluarga dan
Konsumen. [Online] 5 (2), 166–174. Tersedia pada:
http://doi.org.10.24156/jikk.2012.5.2.166 [Diakses 8 Januari 2021].