Other uses, including reproduction and distribution, or selling or
licensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of the
article (e.g. in Word or Tex form) to their personal website or
institutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies are
encouraged to visit:
Responses of the re
fl
ectance indices PRI and NDVI to experimental warming and
drought in European shrublands along a north
–
south climatic gradient
Pille Mänd
a,⁎
, Lea Hallik
a,b, Josep Peñuelas
c, Tiit Nilson
d, Pierpaolo Duce
e, Bridget A. Emmett
f,
Claus Beier
g, Marc Estiarte
c, János Garadnai
h, Tibor Kalapos
i, Inger Kappel Schmidt
j, Edit Kovács-Láng
h,
Patricia Prieto
c, Albert Tietema
k, Joke W. Westerveld
k, Olevi Kull
aaDepartment of Botany, Institute of Ecology and Earth Sciences, University of Tartu, Lai, 40, Tartu 51005, Estonia
bDepartment of Plant Physiology, Institute of Agricultural and Environmental Sciences, Estonian University of Life Sciences, Kreutzaldi 1, Tartu 51014, Estonia cEcophysiology and Global Change Unit CSIC
–CEAB–CREAF, CREAF (Center for Ecological Research and Forestry Applications), Edifici C,
Universitat Autònoma de Barcelona, 08193 Bellaterra, Barcelona, Spain
dTartu Observatory, Tõravere, Estonia
eInstitute of Biometeorology, Consiglio Nazionale delle Ricerche, CNR
–IBIMET, Via Funtana di lu colbu 4/a, 07100, Sassari, Italy
fCentre for Ecology and Hydrology
—Bangor, Environment Centre Wales, Deiniol Rd., Bangor, Gwynedd LL57 2UW, United Kingdom
gRISØ National Laboratory, P.O. Box 49, DK-4000 Roskilde, Denmark
hInstitute of Ecology and Botany, Hungarian Academy of Sciences, H-2163 Vácrátót, Alkotmány u. 2-4, Hungary
iDepartment of Plant Taxonomy and Ecology, Institute of Biology, Loránd Eötvös University, H-1117 Budapest, Pázmány P.s. 1/c, Hungary jForest & Landscape Denmark, Copenhagen University, Hørsholm Kongevej 11, DK-2970 Hørsholm, Denmark
kInstitute for Biodiversity and Ecosystem Dynamics (IBED), University of Amsterdam, Nieuwe Achtergracht 166, 1018 WV Amsterdam, The Netherlands
a b s t r a c t
a r t i c l e
i n f o
Article history:
Received 14 February 2008
Received in revised form 5 November 2009 Accepted 15 November 2009
Keywords:
Chlorophyllfluorescence Climate change Drought Green LAI NDVI PRI
Reflectance indices Remote sensing Soil reflectance VULCAN Warming
The aim of this study was to evaluate the use of ground-based canopy reflectance measurements to detect changes in physiology and structure of vegetation in response to experimental warming and drought treatment at six European shrublands located along a North–South climatic gradient. We measured canopy reflectance, effective green leaf area index (green LAIe) and chlorophyllfluorescence of dominant species. The treatment effects on green LAIe varied among sites. We calculated three reflectance indices: photochemical reflectance index PRI [531 nm; 570 nm], normalized difference vegetation index NDVI680[780 nm; 680 nm]
using red spectral region, and NDVI570[780 nm; 570 nm] using the same green spectral region as PRI. All three
reflectance indices were significantly related to green LAIe and were able to detect changes in shrubland vegetation among treatments. In general warming treatment increased PRI and drought treatment reduced NDVI values. The significant treatment effect on photochemical efficiency of plants detected with PRI could not be detected byfluorescence measurements. However, we found canopy level measured PRI to be very sensitive to soil reflectance properties especially in vegetation areas with low green LAIe. As both soil reflectance and LAI varied between northern and southern sites it is problematic to draw universal conclusions of climate-derived changes in all vegetation types based merely on PRI measurements. We propose that canopy level PRI measurements can be more useful in areas of dense vegetation and dark soils.
© 2009 Elsevier Inc. All rights reserved.
1. Introduction
The climate is changing and further increases in temperature and changes in precipitation are projected for the future (IPCC, 2007). These changes dictate the need to predict possible changes in ecosystem functioning.Harte et al. (1995) and Dunne et al. (2004)
have proposed that a combination of manipulative experiments and natural gradient investigations are required in order to distinguish among ecosystem responses those that are generally uniform, those expressed at different temporal or spatial scales, and those that are highly context dependent. A European shrubland vulnerability study consisting of a series of replicated climate changefield experiments in
–
Abbreviations:green LAI, green Leaf Area Index; green LAIe, effective projected green Leaf Area Index; Fv/Fm, maximal photosynthetic efficiency of leaves; LHCII, Light-Harvesting Complex of photosystem II; NDVI, Normalized Difference Vegetation Index; NDVI680, Normalized Difference Vegetation Index calculated from spectral regions at
780 nm and 680 nm; NDVI570, Normalized Difference Vegetation Index calculated from
spectral regions at 780 nm and 570 nm; NPQ, non-photochemical quenching; PAM, Pulse Amplitude Modulation; PRI, Photochemical Reflectance Index; PSII, Photosystem II; qN, non-photochemical quenching; ΦII, photosystem II quantum yield; VI,
Vegetation Index.
⁎Corresponding author.
E-mail address:[email protected](P. Mänd).
0034-4257/$–see front matter © 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.rse.2009.11.003
Contents lists available atScienceDirect
Remote Sensing of Environment
six shrubland ecosystems across Europe involving gradients of mean annual temperature and precipitation is an example of this type of combined approach (Beier et al., 2004).
This combined approach faces several challenges including the need to detect changes in plant growth or functioning in heteroge-neous, natural shrubland canopies in response to realistic climate change treatments; to integrate responses at the canopy level; and to develop practical tools that can be used to monitor future changes at the landscape scale. Optical reflectance provides a tool that can detect physiological change at both the leaf and canopy scale. Remote sensing technology from airborne and satellite platforms also allows rapid evaluation of vegetation properties and their response to global change over large areas (Kerr and Ostrovsky, 2003; Running et al., 1999). Therefore, this technology has great potential in monitoring changes at a range of scales and to evaluate in vegetation the extent of changes driven by climate change.
Remote sensing studies widely use a reflectance index NDVI (Normalized Difference Vegetation Index) as an estimator of the changes in green LAI and green biomass (Gamon et al., 1995), although the relationship between NDVI and green LAI is in practice largely empirical and needs to be verified using ground measurements (Chen & Cihlar, 1996). In some cases, NDVI has been shown to correlate also with net primary production (Prince, 1991) and photosynthetic rates (Gamon et al., 1995). However, NDVI manages to detect long-term changes in photosynthesis primarily if LAI is also affected. Although minor changes in NDVI have been detected also due to changes in leaf angles (Dobrowski et al., 2005) and pigment pool sizes (Gamon & Surfus, 1999; Gitelson & Merzlyak, 1997), NDVI is shown to be relatively insensitive to initial declines in photosynthesis (Grace et al., 2007; Stylinski et al., 2002). Thus, because ecosystem variables estimated by NDVI change slowly, early diagnosis of initiated changes requires indices that focus on specific, short-term stress responses.
Several stress factors induced by changing climate, such as drought and supraoptimal or suboptimal temperatures, often lead to decreased photosynthetic yield in plants. Under such stress, light intensity is greater than needed for PS II photochemistry. Several photoprotective pathways rechannel light energy to protect plant photosynthetic systems from the damaging effect of excess irradiance. One means to detect vegetation stress–response to changing environments is to track changes in photoprotective mechanisms of plants (Maxwell & Johnson, 2000). Measuring chlorophyll afluorescence is a way to monitor changes in the efficiency of photochemistry and photoprotection, as absorbed light energy is either used in photosynthesis, re-emitted asfluorescence or dissipated through photoprotective pathways as heat (Bilger et al., 1995; Maxwell & Johnson, 2000, Walters & Horton, 1991). However, several unsolved problems remain in measuringfluorescence by remote sensing tools (Moya et al., 2004). Although the exact mechanisms of heat dissipation are still under debate, one of the best understood photo-protective mechanisms of photosynthetic systems—which has been shown to vary in response to temperature and water stress—is de-epoxidation of xanthophylls (Demmig-Adams & Adams, 1992), as zeaxanthin induces changes in the conformational state of LHCII (Moya et al., 2001). The de-epoxidation-state of xanthophylls and aggregation state of PSII antennas are reflected accordingly by absorbance changes near 505 and 535 nm (Bilger et al., 1989). A specific Photochemical Reflectance Index (PRI) (Gamon et al., 1992; Peñuelas et al., 1995) is calculated by using a reflectance band at 531 nm. The reflectance signal at 531 nm contains information on changes in both the xanthophyll cycle and the aggregation state of PSII antennas, and thus PRI is a good approximation of their combined effect (detailed explanation in
Peñuelas et al. (1995) and Gamon et al. (1997). The 570 nm reference waveband remains unaffected by the de-epoxidation reaction. Indeed, stress-induced changes in photosynthetic efficiency have been success-fully measured using this specific Photochemical Reflectance Index (PRI) (Evain et al., 2004; Gamon et al., 1992; Guo and Trotter, 2006; Peñuelas et al., 1995; Stylinski et al., 2002).Filella et al. (2009)
dem-onstrated that PRI reveals both short-term changes in de-epoxidation state of xanthophylls, but it also follows long-term changes in carotenoids/ chlorophyll. Indeed, in studies where larger timescales or several species are involved, direct measurements of plant pigment pools (such as carotenoid/chlorophyll ratio), changes in which are often related to long-term stress–response of plants, have been found to correlate with PRI (across species: Filella et al., 2009; Guo & Trotter, 2006; Martin et al., 2007; Sims & Gamon, 2002; across seasons:Stylinski et al., 2002). The usefulness of NDVI and PRI indices as tools to track plant physiological changes has been demonstrated recently in shrubland communities (Filella et al., 2004; Sims et al., 2006). However, reflectance indices derived from spectral reflectance in visible/near infrared wavebands are also sensitive to changes in biomass, foliar chemistry, canopy structure, water content etc. (Grace et al., 2007; Peñuelas & Filella, 1998; Ruimy et al., 1994; Sims & Gamon, 2002; Ustin et al., 1991). As a result, the relationship between PRI and photosynthetic perfor-mance is also dependent on canopy structure, including leaf area index and leaf angle distribution (Barton & North, 2001; Grace et al., 2007). Furthermore, leaf and canopy properties determine the proportion of bare soil, which contributes to remotely measured canopy reflectance. The relationship between canopy PRI and photosynthesis of plants depends also on soil reflectance, as soil PRI tends to be considerably more negative than that of dense canopies (Barton & North, 2001).
The aim of our study was to evaluate the performance of chlo-rophyllfluorescence measurements and ground-based canopy refl ec-tance measurements in visible/near-infra-red wavebands to detect changes in canopy physiology and structure in response to experi-mental warming and drought treatments in shrubland ecosystems across Europe. Specific attention was given to evaluate the perfor-mance of canopy level measured PRI as an estimator offluorescence parameters qN and Fv/Fm among sites with different LAI and soil brightness. The study was conducted as part of a European-wide manipulative-gradient study on shrublands in six different locations (Beier et al., 2004; Peñuelas et al., 2007).
2. Materials and methods
2.1. Sites and manipulations
Manipulations were carried out on shrublands at 6 sites (Wales-UK, Denmark, Netherlands, Hungary, Sardinia-Italy, and Catalonia-Spain) across a European gradient in temperature and precipitation (Beier et al., 2004). Site parameters of location, temperature, precipitation, nutrient availability, soil type, dominant species and plant coverage are given inTable 1. In each site, we conducted plot-scale night-time warming and repeated drought treatments and the response to the treatments was compared to control plots. Each type of manipulation was performed in three replicate plots per treatment at each site (see details inBeier et al., 2004).
2.1.1. Warming treatment
Night-time warming was performed by covering the vegetation with reflective curtains at night (Beier et al., 2004). Solar energy is normally accumulated in the ecosystem during the day and a fraction of the energy is radiated back to the atmosphere at night as long wave infrared-radiation. Covering of the plots with reflective aluminium foil curtains (ILS ALU, AB Ludvig Svensson, Sweden) reduced the loss of infrared-radiation. The curtains reflected 97% of the direct and 96% of the diffuse radiation. The warmed plots were 20 m2(5 × 4 m) covered by a light-weight scaffolding that supported the reflective aluminium curtain. The curtains of the study plots were activated automatically according to preset light (b200 lux), rain and wind (b10 m s−1) conditions (Beier et al., 2004). The curtains reduced the night-time heat loss by 64%, from 33 W m−2to 12 W m−2, and increased the temperature of the soil and plants by 0.5–1.5 °C (DK), 0–2 °C (UK), 0–1 °C (NL), 1–2 °C (SP), 0.3–1.3 (HU), and 0.2–0.6 °C (IT). In order
to reduce the effect on the hydrological cycle, sensors automatically removed the covers during rain events. The warming treatment had been applied since spring 1999 in the UK, NL, DK and SP and spring 2001 in HU and IT. These moderate temperature elevations led to an increase in the average annual growth potential (Growing Degree Days) by 9–16% at the non-Mediterranean sites and a clear reduction (19–44%) of the number of frost days (Beier et al., 2004).
2.1.2. Drought treatment
The drought treatment was performed for 2-month periods in the spring/summer growing seasons since 1999 (at the Spain and Italian site an additional drought period was established in the autumn growing season;Beier et al., 2004) by covering the vegetation with waterproof, transparent covers. Likewise, treatments started later in HU and IT, in spring 2001. The drought plots were constructed similarly to the warming plots except that the curtain material was a transparent plastic and that the withdrawal of the curtains was governed only by rain and wind. During the drought periods rain sensors activated the curtain to cover the plots during rain and remove the curtains when the rain stopped. The curtains were removed automatically if the wind speed exceeded 10 m s−1. For the part of the year without drought treatment, the drought plots were run parallel to the control plots (Beier et al., 2004). The amount of rainfall reduced by drought treatment in 2003 was 349 mm (UK), 334 mm (SP), 149 mm (IT), 142 mm (DK), 48 mm (NL) and 39 mm (HU). For the rest of the year, the drought treatment was inactive allowing rewetting of the soil. Three untreated control plots with a similar light scaffolding as for the warming and drought treatments but without any curtain were installed for comparison.
2.2. Reflectance measurements
Canopy reflectance was measured in summer 2003 at the same time with plant stress measurements (chlorophyllfluorescence). At each site, the measurements were made during summer growing-period (de-tailed dates atTable 1), where drought treatment had been going on at least for two weeks (see drought treatment description atBeier et al., 2004). Canopy reflectance was measured with a ground-based S2000-FL spectrometer (Ocean Optics Inc., Dunedin, FL, USA)—using a
laptop-computer and a 5 m long, 50μm diameterfiber-optics—on the same subplots where vegetation cover was estimated by pin-point measure-ments (Peñuelas et al., 2007). Reflectance data of 1637 approximately evenly spaced spectral bands (FWHM∼3 nm) in the spectral region between 400 nm and 950 nm was collected from nadir on sunny days during time period between 2 h before and 2 h after local solar noon at 1-m height above the canopy with afield of view angle of 12°. For spatial averaging, thefiber optic was placed over the canopy and the scans were taken in this position. At least 90 reflectance spectra per plot were recorded. In addition to plot reflectance spectra, separate reflectance measurements were made above each plant measured forfluorescence. In order to determine spectral properties of the canopy background, bare (unvegetated) soil outside the plots were also scanned. A calibrated grey standard was used as a reference surface. The vegetation indices were calculated as follows:
NDVI680=ðR780−R680Þ=ðR780 + R680Þ ð1Þ
NDVI570=ðR780−R570Þ=ðR780 + R570Þ ð2Þ
PRI =ðR531−R570Þ=ðR570 + R531Þ ð3Þ
where Rxis the reflectance atxnm. R780 was calculated as the mean of five wavebands centred on 779.82 nm, 780.14 nm, 780.47 nm, 780.79 nm, and 781.12 nm respectively. R680 was calculated as average of three wavebands centred on 680.16 nm, 680.5 nm, and 680.84 nm. R570 was calculated as average of three wavebands centred on 570.13 nm, 570.48 nm, and 570.83 nm. R531 was calculated as average of three wavebands centred on 531.13 nm, 531.49 nm, and 531.84 nm. Due to variable LAI among studied sites and in order to assure that differences between the performance of NDVI680and PRI were not merely the result of saturation in the red band, we calculated a second NDVI using the green band at 570 nm. As the correlation between NDVI680and NDVI570was strong across all countries (0.93brb0.99, pb0.0001) we henceforth use NDVI without subscript to refer to the relationships that apply to both NDVI680and NDVI570.
Table 1
Characteristics of the six study sites at Wales-UK (UK), Denmark (DK), Netherlands (NL), Hungary (HU), Catalonia-Spain (SP) and Sardinia-Italy (IT). Mean values are depicted.amarks
shrublike species,bis for graminoids,cis for other types of plants. For further details seeBeier et al. (2004).
UK DK NL HU SP IT
Altitude (m) 490 58 25 130 210 30
Air temperature (Co)
Year (2003) 7.6 8.7 8.7 11.4 15.7 17.0
July 13.9 19.7 15.4 23.2 26.3 25.6
January 2.0 0.1 2.3 −1.5 6.7 10.1
Precip. (Study period mm/yr)
1741 734 1093 411 582 513
Nitrogen deposition
Peaty podzol Sandy podzol Sandy podzol Calcaric arenosols Petrocalcic
2.3. Leaf area index estimation (pin-point measurements)
The pin-point method (Jonasson, 1988) was used to measure plant frequency, and indirectly to estimate effective projected leaf area index. These pin-point measurements were conducted within two weeks of reflectance measurements. A sharpened pin was lowered through the vegetation at a number of points. Each plant contacted by the pin was counted and the plant part and species (or species group) was recorded. At least 300 such measurements points were made in each experimental plot. These points were arranged at 5 cm intervals alongfive 3-m long transect lines (in Spain, Italy and Hungary) or four 4-m long transect
lines (in the Netherlands) or in subplots (in the UK and Denmark). We used the total number of hits for each plant part per transect, and also as a proportion of all pin hits per transect. Effective projected leaf area index (green LAIe) was calculated as average number of contacts with green parts (leaves and green branches) of the plants per pin.
2.4. Fluorescence measurements
Plant photochemical efficiency was assessed in summer 2003 from in situ measurements of electron transport capacity using modulated chlorophyllfluorescence equipment PAM-2000 (Walz GmbH, Effeltrich, Germany). These measurements were usually made on two dominant species (except in Netherlands where only one species was dominant and in Hungary where three species had similar abundances) at each site in conjunction (measuring time as close as possible) with reflectance measurements (except for predawn measurements). At each treatment plot (3*3 plots), 6 leaves were measured byfluorescence method. To be able to compare changes in reflectance andfluorescence parameters, thefluorescence measurements were performed only at the topmost layer of the canopy. Canopy level reflectance was measured (at each plot) above the same plants where leaf-levelfluorescence measurements were performed. Maximal photosynthetic efficiency (Fv/ Fm;Genty et al., 1989) was monitored in dark-adapted leaf samples during the measurement period around midday and predawn. For measuring daytime Fv/Fm plants were covered with a dark cloth for a 10-min period. Electron transport capacity of PSII (ΦII;Genty et al., 1989) was measured following to daytime Fv/Fm measurements, after plant was kept at saturating light intensity for 5 min around midday (between 2 h before and 2 h after solar noon) using, when necessary, an additional light source. In order to choose the appropriate intensity for saturating light, we measured a light response curve at every site on a single leaf. We found, that at every site we could use the light intensity of 800 µmol m−2s−1without causing photoinhibition and reaching still on the plateau in a light response curve. Fractions of absorbed light that were used in photochemistry and dissipated thermally were assessed using thefluorescence quenching analysis technique, during which a fluorescence parameter qN (non-photochemical quenching;Schreiber et al., 1995) was measured.
2.5. Statistical analyses
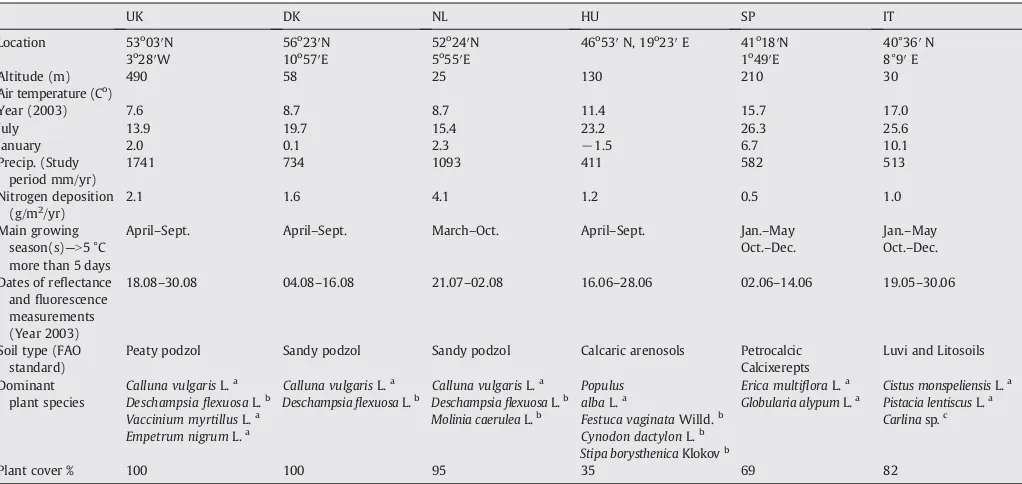
STATISTICA 7.1, StatSoft, Inc., software was used for statistical analysis. We generated general linear models to test the effects of cross-sites climatic variables as continuous factors on green LAIe, fluorescence parameters and reflectance variables NDVI and PRI. A separate model tested the effect of green LAIe on NDVI and PRI. In an additional model, the effects of differentfluorescence parameters on PRI were tested using PRI data measured on the same plants asfluorescence measurements. Treatment was included as a categorical factor in all models and also the interactions of treatments and continuous predictors were included to test the dependence of different relationships on manipulations. The normality and homoscedasticity of data were also Fig. 1.Relationship between effective projected green leaf area index (green LAIe) and
annual precipitation (A) and mean temperature of previous year (B). The drought and warming treatment effects are expressed as precipitation and temperature values. Mean values ± standard errors (N= 13) from six sites are shown: Wales-UK (UK), Denmark (DK), Netherlands (NL), Hungary (HU), Catalonia-Spain (SP) and Sardinia-Italy (IT).
Table 2
The effect of experimental treatments (Treat.—experimental warming and drought treatments) and site-specific differences in precipitation and temperature (Precip.—mean annual precipitation at every site; Temp.—mean temperature of 12 month previous to measurements at every site ) on measured vegetation properties (green LAIe—effective projected green Leaf Area Index; Fv/Fm—maximal photosynthetic efficiency of leaves;ΦII—photosystem II quantum yield; qN—non-photochemical quenching) and reflectance indices (NDVI680—
Normalized Difference Vegetation Index calculated from spectral regions at 780 nm and 680 nm; NDVI570—Normalized Difference Vegetation Index calculated from spectral regions at 780 nm and 570 nm; PRI—Photochemical Reflectance Index.). Table shows the results of across-site GLM test:N= 13;pvalues for significant differences are shown, ns—pN0.05.
Green LAIe Predawn Fv/Fm Midday Fv/Fm ΦII qN NDVI680 NDVI570 PRI
Treat. 0.02 ns ns ns ns ns ns ns
Precip. b0.001 ns 0.002 ns ns 0.03 ns b0.001
Treat.*Precip. b0.001 ns ns ns ns ns ns ns
Temp. b0.001 b0.001 ns b0.001 b0.001 b0.001 b0.001 b0.001
Treat.*Temp. ns ns ns ns ns ns ns ns
Temp.2
b0.001 b0.001 b0.001 b0.001 b0.001 b0.001 b0.001 b0.001
Treat.*Temp.2 ns ns ns ns ns ns ns ns
tested. The squared component of a continuous factor was included in the models if the tested variable was nonlinearly related to a given factor. As the effect of plot was found insignificant, this factor was not included in thefinal models. The effect of country as a factor was omitted from the models.
A different model was constructed to test the effect of treatment and country and their interaction on NDVI and PRI. In addition, we tested the warming and drought treatment effects on green LAI, NDVI and PRI at all six sites separately using Fisher LSD post-hoc test. We used Fisher LSD post-hoc test also to analyse the differences in soil reflectance parameters among sites.
Pearson's correlation coefficients were calculated between refl ec-tance indices: PRI, NDVI measured above plants, and vegetation properties: photosynthetic efficiency (Fv/Fm), photosystem II quan-tum yield (ФII), and green LAIe. Multiple regression analysis was performed to estimate midday Fv/Fm and non-photochemical quenching (qN) from reflectance indices measured above plants
(PRI, NDVI570) and above bare soil (PRIsoil). Correlation and regression analysis were performed for pooled data from all six countries and separately for the three northern sites (UK, DK, NL) and the three southern sites (HU, IT, SP) in order to distinguish countries with very different canopy structure (dense versus sparse).
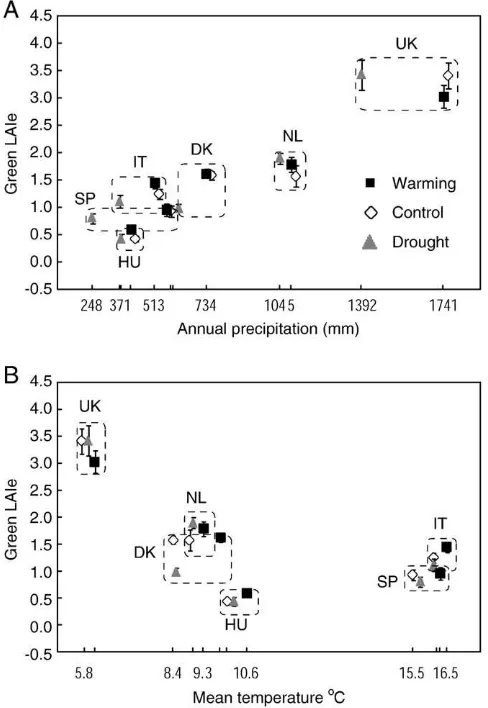
Fig. 2.Changes in green LAIe, NDVI680and PRI determined by warming (A) and drought
(B) treatment at different sites. Changes are expressed as a percentage relative to the control treatment. * indicates significant differences (pb0.05) compared to control according
to Fisher LSD test.
Table 3
The effect of experimental treatments (Treat.—experimental warming and drought treatments) and sites on reflectance indices (NDVI680—Normalized Difference Vegeta-tion Index calculated from spectral regions at 780 nm and 680 nm; NDVI570—
Normalized Difference Vegetation Index calculated from spectral regions at 780 nm and 570 nm; PRI—Photochemical Reflectance Index.). Table shows the results of across-site GLM test:N= 13;pvalues for significant differences are shown, ns—pN0.05.
NDVI680 NDVI570 PRI
Treat. 0.003 0.002 0.02
Country b0.001 b0.001 b0.001
Treat.*Country 0.01 0.02 ns
Fig. 3.Comparison of the treatment effects on NDVI680and PRI (data from Wales
excluded). Effects of both drought and warming treatments are included.
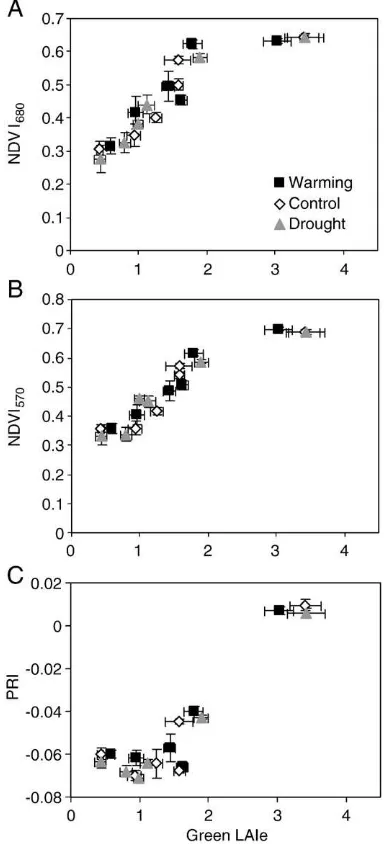
Fig. 4.Relationships between reflectance indices NDVI680(A), NDVI570(B) and PRI
(C)versuseffective projected green leaf area index (green LAIe). Data from the three treatments in the six sites are included. Bars indicate standard error (N= 13).
3. Results
3.1. The effects of temperature and precipitation
Green LAIe increased significantly with increasing annual precipita-tion at different sites (Fig. 1A,Table 2). The relationship between green LAIe and precipitation slightly changed as a result of different treatments (Table 2). Differences in mean annual temperature did not account for changes in green LAIe as clearly as precipitation, and the relationship was significantly non-linear (Fig. 1B,Table 2). Although green LAIe started to decrease with increasing mean annual tempera-ture at northern sites, no such trend was evident at southern sites—Italy and Spain. Different treatments did not affect the relationship between green LAIe and temperature (Table 2). In general, reflectance indices followed similar patterns as green LAIe when plotted against different climatic parameters. However, no treatment effect was found for reflectance indicesversusannual rainfall (Table 2). Fluorescence-based estimations for plant photochemical efficiency did not follow changes along the large-scale precipitation gradient as clearly as green LAIe. Only maximal photochemical efficiency (Fv/Fm), measured at leaf level after 10 min of darkening at midday, was significantly influenced by differences in precipitation (Table 2). Unlike midday depression of Fv/ Fm, the predawn regeneration of Fv/Fm was influenced by temperature, not by precipitation (Table 2). All measuredfluorescence parameters showed significant non-linear relationships with mean annual temper-ature (Table 2); Fv/Fm (predawn and midday) andΦIIwere lowest and qN highest at mid-ranged temperatures. Relationship betweenfl uores-cence values and climatic variables remained unchanged with different treatments (Table 2).
In general, differences in reflectance parameters and green LAIe among the different sites were significantly larger than differences determined by treatments. Higher values of PRI in warming treatment plots and lower values of NDVI in drought treatment plots were recorded in cross-sites comparison (mean effect atFig. 2). However, the effect of treatments was significant only when the effect of country was included in the model (Table 3). Treatments also significantly affected green LAIe (Table 2). When we analysed each
experimental site separately, warming treatment significantly in-creased NDVI in Italy and PRI in Spain, and drought treatment significantly decreased NDVI and green LAIe in Denmark (Fig. 2). Generally, the treatment-induced changes in green LAIe, NDVI and PRI were related, despite the saturation of NDVI. When excluding data from Wales, the treatment effects on NDVI680and PRI values were strongly positively correlated (Fig. 3). At the Welsh site, reflectance was influenced somewhat by heavyflowering ofCalluna, asflowering increased reflectance in blue and red spectral regions.
3.2. Relationships between reflectance indices,fluorescence parameters and green LAIe
Comparison of NDVI680(using red band) values with green LAIe showed a significant relationship with strong saturation at medium to high LAIe (Fig. 4A). As this reflectance index, using red spectral region (chlorophyll absorbance maximum), might saturate already at moderate LAI conditions, we constructed an additional index, which uses the green spectral region (NDVI570). However, the similar saturation occurred if NDVI570(using green band) was plotted against green LAIe (Fig. 4B). No significant systematic changes were found in the NDVIversusgreen LAIe relationship when different treatments were considered (Table 4). Photochemical reflectance index (PRI) was also significantly related to green LAIe (Fig. 4C; Table 4), but no saturation of PRI was evident. However, the relationship of PRIversus green LAIe showed a significant non-linear component and the relationship depended on the treatment (Table 4).
As the relationships between canopy level PRI and fluorescence parameters are affected by LAI, we investigated correlations of the green LAIe andfluorescence parameters (of topmost leaves of canopy) versus both NDVIs (NDVI680and NDVI570) and PRI. For the pooled data of all six sites,fluorescence parameters were correlated significantly to canopy level PRI (Table 5, qNversusPRI atFig. 5), suggesting that besides the effect of canopy structure on PRI, also a physiological component is detectable in PRI. Fluorescence parameters were also significantly correlated to canopy level NDVIs (Table 5). Similarly, green LAIe was correlated to both NDVIs and PRI (Table 5). When we investigated the three northern sites, which are characterised by denser vegetation, fluorescence parameters were related stronger to PRI than to NDVI, but the correlation between PRI and green LAIe was stronger than that between fluorescence parameters and PRI (Table 5). In the three southern sites, where the vegetation was sparse and bare soils more abundant, PRI correlated only weakly with green LAIe, and the relationships between fluorescence parameters and PRI were non-significant (Table 5). Whereas most fluorescence parameters were weakly but significantly correlated to NDVI in the southern sites, it was not for qN (Table 5). Green LAIe was strongly correlated to NDVIs in the southern sites as well (Table 5). In general, the relationships with vegetation indices using the green band (NDVI570 and PRI) were stronger in the northern sites, whereas they were stronger with indices using the red band (NDVI680) in the southern sites (Table 5).
Table 5
Pearson's correlation coefficients between reflectance indices measured above the plants (NDVI680—Normalized Difference Vegetation Index calculated from spectral regions at 780 nm and 680 nm; NDVI570—Normalized Difference Vegetation Index calculated from spectral regions at 780 nm and 570 nm; PRI—Photochemical Reflectance Index.) and vegetation properties: photochemical efficiency parameters (Fv/Fm—maximal photosynthetic efficiency;ФII—photosystem II quantum yield; qN—non-photochemical quenching) and effective projected green Leaf Area Index (green LAIe). Correlation coefficients were calculated for pooled data of all six countries and separately for three northern sites (UK, DK, NL) and three southern sites (HU, IT, SP). Statistical significance is shown as:pb0.001***,pb0.01**,pb0.05* andpN0.05 ns.
All six countries 3 northern sites 3 southern sites
PRI NDVI570 NDVI680 PRI NDVI570 NDVI680 PRI NDVI570 NDVI680
Fv/Fm midday 0.52*** 0.52*** 0.53*** 0.77*** 0.62*** 0.48*** 0.15 ns 0.40 ** 0.45 **
Fv/Fm predawn 0.29 ** 0.34 ** 0.36*** 0.58*** 0.45 ** 0.34 * 0.28 ns 0.30 * 0.32 *
YieldФII 0.43*** 0.58*** 0.54*** 0.63*** 0.46 ** 0.33 * 0.12 ns 0.43 ** 0.43 **
qN −0.65*** −0.56*** −0.45*** −0.72*** −0.50*** −0.34 * −0.17 ns −0.23 ns −0.19 ns
green LAIe 0.81*** 0.78*** 0.74*** 0.82*** 0.77*** 0.66*** 0.36 ** 0.65*** 0.71***
Table 4
The effect of green leaf area index (green LAIe) and experimental treatments (Treat—
experimental warming and drought treatments) on reflectance indices (NDVI680—
Normalized Difference Vegetation Index calculated from spectral regions at 780 nm and 680 nm; NDVI570—Normalized Difference Vegetation Index calculated from spectral regions at 780 nm and 570 nm; PRI—Photochemical Reflectance Index). Table shows the results of across-site GLM test:N= 13; pvalues for significant differences are shown, ns—pN0.05.
NDVI680 NDVI570 PRI
Treat. ns ns 0.03
green LAIe b0.001 b0.001 ns
Treat*LAIe ns ns 0.006
green LAIe2
b0.001 b0.001 b0.001
Treat*LAIe2 ns ns 0.002
As all the reflectance indices used in this study are known to be sensitive to background spectral properties, reflectance was also measured above the bare soil. In general, the soil was brighter in southern sites (Table 6). The reflectance indices measured above the bare soil differed significantly among the sites (Table 6). PRI values measured above bare soils were significantly more negative in Italy and Spain than in the other sites. The northern sites did not differ significantly (Table 6). Both NDVI680and NDVI570of bare soils were lower in southern sites than in northern sites (data not shown). As the empirical relationship betweenfluorescence parameters and PRI can be influenced by the background spectral properties and the amount of vegetation, multiple regression analyses were conducted (Table 7). To control for the simple effect of biomass, NDVI570, which utilizes the same reflectance band as PRI, was included in the regression model. In addition to reflectance measured above the vegetation, soil PRI were also included to control for the effect background properties. Midday Fv/Fm and qN were used as dependent variables, because these fluorescence parameters had the strongest correlations with PRI measured above the plants (Table 5). For pooled data of all six sites, the
regression model explained 47% of the variability of qN and 48% of the variability of midday Fv/Fm (Table 7). Similarly to pair-wise correla-tions, the regression analysis showed that canopy level PRI measure-ments could be used to estimate qN and midday Fv/Fm in northern sites, particularly if soil reflectance measurements are included, but in the southern sites the relationships between the fluorescence parameters and PRI are dominated by soil reflectance (Table 7).
4. Discussion
Our results show that spectral reflectance is able to distinguish differences in canopy structure and physiology due to site and treatment effects, although many of these differences are confounded by multiple, interacting factors. According to our measurements, differences in effective projected green leaf area index (green LAIe) were in accordance with changes in reflectance indices NDVI and PRI. This is consistent with earlier studies, in which NDVI has proved to be a good indicator of green LAI and green biomass, whereas the relationship with total LAI and total canopy biomass is usually less adequate due to large variability in the abundance of non-green standing biomass (Gamon et al., 1995). However, differences in reflectance indices among treatments in this study were considerably smaller than differences among sites, which concur with ourfindings on green LAIe. Similarly,
Peñuelas et al. (2007)reported moderate changes in plant productivity in response to treatments. Nevertheless, NDVI values in general (when data from all sites was pooled) decreased significantly due to drought treatment in agreement with changes in total aboveground biomass that was detected in another study of the same experiment (Peñuelas et al., 2004, 2007). A similar sensitivity of remote-sensed NDVI to climate-derivedfluctuations in vegetation was evident in a study by
Barbosa et al. (2006). Fluctuations in NDVI in that study were consistent with spatial and temporal changes in precipitation during a 20-year period at several Brazilian ecosystem types (including shrublands) despite the large degree of noise always present in natural systems. Differences in NDVI as a result of large-scale changes in temperature as well as precipitation have been demonstrated in investigations covering
Table 6
Mean values ± standard errors of reflectance (R531—reflectance at 531 nm; R570—reflectance at 570 nm; R680—reflectance at 680 nm; R780—reflectance at 780 nm; PRI—
Photochemical Reflectance Index) of bare soil at study sites in Wales-UK (UK), Denmark (DK), Netherlands (NL), Hungary (HU), Catalonia-Spain (SP) and Sardinia-Italy (IT). Means with the same letter are not significantly different.
Soil R531 Soil R570 Soil R680 Soil R780 Soil PRI
UK 0.044a± 0.004 0.051a± 0.004 0.089a± 0.008 0.143a± 0.014
−0.079a± 0.008
DK 0.048a± 0.003 0.057a± 0.004 0.087a± 0.005 0.128a± 0.008
−0.078a± 0.004
NL 0.048a± 0.007 0.057a± 0.008 0.095a± 0.010 0.156a± 0.016
−0.086ab± 0.005
HU 0.191b± 0.021 0.225b± 0.026 0.268b± 0.028 0.295b± 0.026
−0.082ab± 0.005
SP 0.250c± 0.016 0.306c± 0.019 0.368c± 0.027 0.434c± 0.026
−0.101b± 0.004
IT 0.131d± 0.006 0.168d± 0.010 0.227b± 0.013 0.284b± 0.010
−0.124c± 0.011
Table 7
Results of multiple regression analyses estimating midday photosynthetic efficiency (Fv/Fm) and non-photochemical quenching (qN) from reflectance indices measured above plant (PRIplant, NDVI570 plant) and above bare soil (PRIsoil).ndenotes sample size.
Dependent variable
n Intercept PRIplant Independent variables
NDVI570plant PRIsoil MultipleR2
All six countries qN 88 0.01 −0.48 −0.30 0.19 0.47
p= 0.89 pb0.0001 pb0.005 p= 0.06
Fv/Fm midday 90 0.06 0.46 0.28 −0.37 0.48
p= 0.40 pb0.0005 pb0.005 pb0.0005
3 northern countries qN 42 −0.05 −0.63 −0.14 −0.32 0.54
p= 0.66 pb0.001 p= 0.44 pb0.05
Fv/Fm midday 43 0.05 0.67 0.17 0.14 0.62
p= 0.60 pb0.0001 p= 0.31 p= 0.22
3 southern countries qN 46 0.06 −0.02 0.08 0.66 0.38
p= 0.57 p= 0.83 p= 0.52 pb0.0001
Fv/Fm midday 47 0.01 0.24 0.21 −0.40 0.35
p= 0.90 pb0.05 p= 0.12 pb0.05
Fig. 5.Relationship between reflectance index PRI and non-photochemical quenching (qN). No significant treatment effect was found.
several regions and vegetation types (Gong & Shi, 2003). However, when comparing sites on very different locations (as in our study) with each other, it is possible that differences in sun angle also contribute with adding moderate variance into reflectance data (Goodin et al., 2004; Sims et al., 2006), especially as the effect of sun angle on reflectance values depends on canopy structure (Middleton, 1991), which in our study differed among northern (dense canopies) and southern sites (sparse canopies; see percentages of plant cover at
Table 1).
Despite the positive relationship between NDVI and green LAIe, our results showed that the NDVI—green LAIe relationship tends to saturate at medium to high green LAIe, such as Calluna-type vegetation in northern sites. Our results support the findings of other similar studies (e.g.Gamon et al., 1995; Pontailler et al., 2003) which showed that NDVI is relatively insensitive to changes in canopy structure when LAI is large. Thus, indices in addition to NDVI should be considered to indicate climate-induced changes in vegetation properties at different scales of biomass and productivity, e.g. the photochemical reflectance index (PRI). PRI showed no saturation at medium to high green LAIe, in contrast to NDVIs, which saturated even though we had constructed NDVI570 using the green band (570 nm) to account for the effect that in the red spectral region, which is near to the absorbance maximum of chlorophyll, saturation occurs at lower LAI than in the green band, where chlorophyll absorbance is less. Indeed, we found that the relationships between green LAIe,fluorescence parameters and vegetation indices using the green band (NDVI570 and PRI) were stronger in the northern sites, whereas the relationships with VI using the red band (NDVI680) were stronger in the in southern sites.
PRI is calculated from wavelength regions at 531 nm to detect changes in plant physiological properties, and a reference waveband at 570 nm, which remains unaffected by the de-epoxidation reaction, yet remains influenced by changes in canopy structure and biomass. While PRI responds to changes both in physiological and structural properties of a canopy, NDVI570 should be sensitive mainly to the influence of canopy structure and biomass. Indeed, it has been shown that PRI changes with photosynthetic rate, while NDVI may remain more stable for a longer time (Grace et al., 2007; Stylinski et al., 2002). Classical leaf-level studies (Gamon et al., 1992, 1997; Peñuelas et al., 1995, 1997) have shown strong relationships between PRI and leaf photochemical efficiency. Additional backing for the applicability of PRI as an efficient remote sensing tool for shrublands also comes from earlier canopy or community-level studies (Evain et al., 2004; Filella et al., 1996; Guo & Trotter, 2006; Stylinski et al., 2002). Although classical photochemical efficiency measurements are conducted at leaf level and it is thus problematic to correlate this data to canopy level PRI measurements, earlier studies have proved a strong relationship between leaf-level and canopy level PRI (Gamon & Qiu, 1999; Stylinski et al., 2002), especially if top-canopy leaves are sampled and if the canopy is relatively dense with only little soil background showing. However, interpretation of changes in PRI depends largely on sampling protocol. For instance, PRI is expected to be a good indicator of changes in photosynthesis related to photoprotective mechanisms (Bilger et al., 1989; Peñuelas et al., 1995), which is why PRI has been shown to be best correlated with changes influorescence parameters qN and NPQ (non-photochemical quenching) (Evain et al., 2004; Peñuelas et al., 1997), but also with xanthophyll concentration (Evain et al., 2004) and carotenoid/chlorophyll ratio (Filella et al., 2009; Guo & Trotter, 2006; Martin et al., 2007; Sims & Gamon, 2002; Stylinski et al., 2002). Therefore, as PRI can simultaneously detect changes in both fast and long-term photoprotective mechanisms (Filella et al., 2009), time-scale of sampling and species properties must be considered to distinguish whether PRI detects changes in the xanthophyll cycle per se or in pigment pool sizes (e.g. the relative levels of carotenoids to chlorophylls). It is probable, that in our study, where changes in PRI were followed over several species and study sites, PRI might also
reflect changes in pigment pools, not only short-term photoprotective processes. This is supported by several multi-species studies, which show good correlation between PRI and pigment pool size (Guo & Trotter, 2006; Martin et al., 2007; Sims & Gamon, 2002). In species with naturally fewer photoprotective pigments, the total change in the concentration of xanthophyll pigments in the de-epoxidised state due to stress–response is inevitably small (Guo & Trotter, 2006), and thus the relation between PRI and photosystem II quantum yield may appear species-specific (Peñuelas et al., 1995). In this study we concentrated on short-term photoprotective processes, however, to distinguish also long-term changes in pigment pools as another component affecting PRI, further studies should integrate both measurements of fast processes in xanthophyll cycle and also of pigment pool sizes.
In the present study, our pooled data from all sites indicate a strong relationship between PRI (measured above the plant) and changes in qN (fluorescence parameters were determined on upper leaves of the same plant), and data from 3 northern sites show good agreement between PRI and quantum yield at both fully relaxed and light-saturated state. As northern sites had relatively dense plant canopies, this strong relationship between leaf-levelfluorescence and canopy level reflectance parameters was in accordance with above-mentionedfindings ofGamon and Qiu (1999), as dense canopies have been shown to function much like single leaves when it comes to interpreting the data of PRI (Filella et al., 2004; Stylinski et al., 2002). Moreover, our data indicated that PRI was more strongly related to maximum potential photosynthetic efficiency (Fv/Fm) measured after 10 min of darkening at midday, rather than with predawn Fv/Fm. Probably 10 min of darkening resulted in a status by which only faster relaxation processes of electron transport system had taken place and, since PRI was measured during midday, a stronger relationship with midday Fv/Fm was expected.
Ourfluorescence measurements revealed that treatment related changes in photosynthetic parameters were overshadowed by plant-to-plant or within-plant variability and therefore no statistically significant treatment effects onfluorescence parameters were found at any site. No warming effect on photochemistry was either found at any site during the earlier years of the experiment (Llorens et al., 2004). As a decline in photosynthetic efficiency of plants should be one of the earliest warning indicators of stress caused by changes in temperature or water availability (Larcher, 1995), our results show, either that temperature increase was well within the temperature tolerance of local species, and the heat treatments were not severe enough to cause any changes in photochemistry, or that by alleviating night-time low temperature stress, the night-time heat increases might actually benefit PSII efficiency by enhancing the relaxation of daytime drop in Fv/Fm. Indeed, if we looked at ourfluorescence parameters along the north–south climate gradient, we found that midday depression in Fv/Fm appeared to be more dependent on the precipitation regime, whereas temperature seems to control the overnight regeneration of maximum photochemical efficiency (pre-dawn Fv/Fm). This mechanism of night-time relaxation of photo-chemistry could be even more evident during the extremely dry summer 2003 (Ciais et al., 2005) of present study, when all over Europe control plots were also suffering from drought (Peñuelas et al., 2007) and the amount of rain excluded from drought treated plots was relatively small. This severe drought may be also one important reason behind the relative insensitivity of plant photochemistry to drought treatment that we found during the year of sampling. Another reason behind this lack of response to drought may be demonstrated in an earlier phase of this experiment, whenLlorens et al. (2004)found that drought treatment increased midday leaf potential photochemical efficiency (Fv/Fm) of photosystem II, butΦII remained unchanged, indicating that water stress plants had higher photorespiration rates than control plants, thus protecting PSII from photodamage (Epron, 1997).
We didfind, instead, some evidence of warming treatment effect on plants from reflectance data. Warming treatment generally increased PRI whereas the general effect of warming treatment on NDVI was insignificant. The effect of drought treatment was less evident on PRI, although it was significant on NDVI. The warming effect on PRI in our study appeared to differ among sites, with stronger effects on more energy-limited ecosystems as expected. But as fluorescence data does not confirm the treatment changes in photochemistry, the question remains, which of the parameters affecting PRI changed due to warming treatment. Some changes in plant physiology can still be suggested as we found no treatment effect on the relationship of PRI versusfluorescence parameters, but slight statistical evidence for differences in PRI versus green LAIe relation-ship, and at the same time the relationship between NDVI and green LAIe was unaffected by treatments. Fluorescence measurements may not be sensitive enough merely because of the study-protocol of present study. The number offluorescence measurements was limited and as we dealt with very minimal (intentionally realistic) changes in temperature and precipitation, the sample size might have been too small to detect any differences. A complication with allfluorescence measurements, which may reduce its effectiveness in detecting treatment effects, is thatfluorescence infield conditions depends on several factors, such as light conditions, temperature, time of day etc., increasing uncertainty at different times and locations. Although we made our measurements during growth period and sunny days, close to solar noon, inevitably different study sites had different field conditions and also different life histories of plant species. As such, the interpretation of measuredfluorescence parameters when comparing different study sites depends on those factors (Rascher et al., 2000). Although similarly to fluorescence measurements, PRI data is also strongly affected by different site parameters (as discussed below), the finding of significant effect of warming on PRI suggests, that PRI may be more helpful in detecting small differences in vegetation of very variable natural ecosystems, than limited number of leaf-level fluorescence measurements.
However, exactly which canopy parameter is causing the detected changes in PRI is largely a matter of certain context and often very difficult to determine. Barton and North (2001) used a one-dimensional ray tracing radiative transfer model to investigate the correlation between PRI and canopy light use efficiency (based on gas analysis measurements) and found that the relationship is a function of LAI in sparse vegetation. This is because the proportion of bare soil area with relatively low PRI increases. In their modelBarton and North (2001)showed that soil reflectance has a significant influence on the canopy level measured PRI when LAIb3, and soil brightness increases the effects of soil reflectance. Our northern sites were characterised by denser vegetation and darker soils, whereas the vegetation in the southern sites was sparse and the soils much brighter. Thus, according toBarton and North (2001), we should expect large interference by soil reflectance in southern sites. Indeed, we found a strong relationship between PRI and photochemical efficiency parameters for the northern sites, but no significant correlation between PRI and chlorophyll fluorescence parameters for the southern sites. These findings illustrate the modelling results ofBarton and North (2001)
that predict soil is a confounding variable for PRI in sparse canopies and stands. Furthermore, the pooled data of all six sites showed that the relationship betweenfluorescence parameters (qN and midday Fv/ Fm) and PRI was stronger when the effect of soil and biomass were included in the model. In fact, the low soil PRI in southern countries dominated the canopy scale PRI measurements so strongly, that canopy PRI was only slightly (midday Fv/Fm) or unrelated (qN) to fluorescence parameters, whereas in dense vegetation of northern countries biomass and soil properties added little, if anything, to improve the relationship between PRI andfluorescence. Despite the lack of significant relationships between PRI and fluorescence parameters in southern countries,Filella et al. (2004), studying our
Spanish site, found that PRI followed the seasonal variations in photosynthetic rates in one dominant species,Erica multiflora, and detected decreased photosynthetic rates with drought treatment. However, PRI did not follow the photosynthetic rates of the other dominant species,Globularia alypumplants, whose LAI is lower than E. multiflora, which is consistent with the confounding effect of soil on PRI shown here. Differently from present study,Filella et al. (2004)
measured reflectance at closer distance to leaves of a single species leaving less effect of soil background into their reflectance data. Thus we suggest that future canopy level PRI studies should include also leaf-level reflectance in order to distinguish better between the physiological and the structural components affecting canopy level PRI.
Because reflectance techniques allow for abundant point-measure-ments, PRI measurements may allow detection of small responses on the ecosystem or plant community level. Ourfindings of significant treatment effect on PRI, but no effect onfluorescence values, may reflect the higher number of PRI measurements. On the other hand, changed PRI may only mean changed LAI (or any other canopy structure or soil properties). PRI readings in a study of this scale, where very different locations and species are involved, may also be affected by differences in sun angle (Sims et al., 2006) and even changes in leaf angle distribution (Barton & North, 2001). Thus, in the absence of this information, when interpreting PRI, the question remains how much of the variation in PRI is due to changes in canopy structure (e.g. LAI), how much is due to changing pigment ratios (e.g. carotenoid: chlorophyll levels), and how much stems from changes in photosyn-thetic efficiency due to the xanthophyll cycle. Our data also revealed that the correlation between PRI and green LAIe was stronger than the correlation between PRI and any of thefluorescence parameters. In any case, the treatment effects on NDVI, (reflecting changes in LAI) and the treatment effects on PRI, (reflecting changes in LAI and variation in photochemical efficiency) were generally related to each other. Because biomass accumulation depends on efficiency of photosynthe-sis, the significant correlations we found between photochemical efficiency parameters and NDVI can be expected and even interpreted as causal to some extent. However, as canopy level PRI is also strongly dependent on soil reflectance, particularly in the case of sparse vegetation and bright soils such as in the southern sites in our study, the influence of soil reflectance on the relationship between canopy level PRI and leaf-level photochemical efficiency parameters also needs further research, considering that changes in soil properties may influence both. For further practices it would also be useful to add other remote sensing methods (e.g. canopy radiative transfer modelling or spectral mixture analysis) into experiments which try to distinguish between structural and physiological effects on PRI.
5. Conclusions
This study demonstrates the strength, and many of the challenges, associated with understanding global change effects on plant physiol-ogy with remote sensing. Canopy level reflectance indices were able to track small changes in shrubland vegetation, however the interpre-tation of those changes depends largely on different environmental, ecosystem-specific and species-specific factors. Reflectance indices differed considerably among sites and much less among treatments. Even so, general trend (for pooled data of all six sites) showed that drought treatment tended to reduce NDVI values and warming treatment enhanced PRI. Our study revealed that changes in NDVI can largely be interpreted as changes in effective projected leaf area index (green LAIe) with some limitations in areas with dense vegetation. PRI, on the other hand, had no tendency to saturate at high green LAIe conditions, making PRI a more valuable tool for assessing vegetation status in such conditions. PRI was in good agreement with measured plant-level photochemical efficiencies in dense canopies of northern sites, where we even found higher PRI values in warmed plots, whereas
we found no treatment effect on leaf-levelfluorescence parameters. However, interpretation of changes in PRI needs background informa-tion on pigment pool sizes, canopy structure and soil properties, especially in sparse vegetation areas with bright soils, where soil reflectance strongly influences the recordings of canopy PRI.
Acknowledgements
This research was funded by EU projects CLIMOOR (Contract ENV4-CT97-0694) and VULCAN (Contract EVK2-CT-2000-00094). We also receivedfinancial help from the Estonian Sciences Foundation (Grant 6969), targeted project of the Estonian Ministry of Education and Research (SF0182732s06..11), the Spanish Government (Grant CGL2006-04025/BOS and Consolider Montes CSD2008-00040) and the Catalan Government (Grant SGR2009-458). The research groups from UK, Denmark, Netherlands, Hungary, Spain and Italy are gratefully acknowledged for their work in maintaining the VULCAN sites and collecting climatologic and pin-point data. Our greatest gratitude goes to our colleague and supervisor, Prof. Olevi Kull (deceased 2007), who led the plant stress measurement workgroup in the VULCAN project, who started the writing of this paper, and to whom this paper is dedicated.
References
Barbosa, H. A., Huete, A. R., & Baethgen, W. E. (2006). A 20-year study of NDVI variability over the Northeast region of Brazil.Journal of Arid Environments,67, 288−307.
Barton, C. V. M., & North, P. R. J. (2001). Remote sensing of canopy light use efficiency using the photochemical reflectance index: Model and sensitivity analysis.Remote Sensing of Environment,78, 264−273.
Beier, C., Emmett, B., Gundersen, P., Tietema, A., Peñuelas, J., Estiarte, M., et al. (2004). Novel approaches to study climate change effects on terrestrial ecosystems in the field: Drought and passive nighttime warming.Ecosystems,7, 583−597.
Bilger, W., Bjorkman, O., & Thayer, S. S. (1989). Light-induced spectral absorbance changes in relation to photosynthesis and the epoxidation state of xanthophyll cycle components in cotton leaves.Plant Physiology,91, 542−551.
Bilger, W., Schreiber, U., & Bock, M. (1995). Determination of the quantum efficiency of photosystem II and of non-photochemical quenching of chlorophyllfluorescence in thefield.Oecologia,102, 425−432.
Chen, J. M., & Cihlar, J. (1996). Retrieving leaf area index of boreal conifer forests using Landsat TM images.Remote Sensing of Environment,55, 153−162.
Ciais, P., Reichstein, M., Viovy, N., Granier, A., Ogée, J., Allard, V., et al. (2005). Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature,437, 529−533.
Demmig-Adams, B., & Adams, W. W., III (1992). Photoprotection and other responses of plants to high light stress.Annual Review of Plant Physiology and Plant Molecular Biology,43, 599−626.
Dobrowski, S. Z., Pushnik, J. C., Zarco-Tejada, P. J., & Ustin, S. L. (2005). Simple reflectance indices track heat and water stress-induced changes in steady-state chlorophyllfluorescence at the canopy scale.Remote Sensing of Environment,97, 403−414.
Dunne, J. A., Saleska, S. R., Fischer, M. L., & Harte, J. (2004). Integrating experimental and gradient methods in ecological climate change research.Ecology,85, 904−916.
Epron, D. (1997). Effects of drought on photosynthesis and on the thermotolerance of photosystem II in seedlings of cedar (Cedrus atlanticaandC. libani).Journal of Experimental Botany,48, 1835−1841.
Evain, S., Flexas, J., & Moya, I. (2004). A new instrument for passive remote sensing: 2. Measurement of leaf and canopy reflectance changes at 531 nm and their relationship with photosynthesis and chlorophyllfluorescence.Remote Sensing of Environment,91, 175−185.
Filella, I., Amaro, T., Araus, J. L., & Peñuelas, J. (1996). Relationship between photosynthetic radiation-use efficiency of barley canopies and the photochemical reflectance index (PRI).Physiologia Plantarum,96, 211−216.
Filella, I., Peñuelas, J., Llorens, L., & Estiarte, M. (2004). Reflectance assessment of seasonal and annual changes in biomass and CO2 uptake of Mediterranean
shrubland submitted to experimental warming and drought.Remote Sensing of Environment,90, 308−318.
Filella, I., Porcar-Castell, A., Munné-Bosch, S., Bäck, J., & Peñuelas, J. (2009). PRI assessment of long-term changes in carotenoids/chlorophyll ratio and short-term changes in de-epoxidation state of the xanthophyll cycle.International Journal of Remote Sensing,30, 4443−4455.
Gamon, J. A., & Qiu, H. (1999). Ecological applications of remote sensing at multiple scales. In F. I. Pugnaire, & F. Valladares (Eds.),Handbook of functional plant ecology (pp. 805−846). New York: Marcel Dekker.
Gamon, J. A., & Surfus, J. S. (1999). Assessing leaf pigment content and activity with a reflectometer.New Phytologist,143, 105−117.
Gamon, J. A., Peñuelas, J., & Field, C. B. (1992). A narrow-waveband spectral index that tracks diurnal changes in photosynthetic efficiency.Remote Sensing of Environment, 41, 35−44.
Gamon, J. A., Field, C. B., Goulden, M. L., Griffin, K. L., Hartley, A. E., Joel, G., et al. (1995). Relationship between NDVI, canopy structure and photosynthesis in three Californian vegetation types.Ecological Applications,5, 28−41.
Gamon, J. A., Serrano, L., & Surfus, J. S. (1997). The photochemical reflectance index: an optical indicator of photosynthetic radiation use efficiency across species, functional types, and nutrient levels.Oecologia,112, 492−501.
Genty, B., Briantais, J. M., & Baker, N. R. (1989). The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyllfl uores-cence.Biochimica et Biophysica Acta,990, 87−92.
Gitelson, A. A., & Merzlyak, M. N. (1997). Remote estimation of chlorophyll content in higher plant leaves.International Journal of Remote Sensing,18, 2691−2697.
Gong, D. -Y., & Shi, P. -J. (2003). Northern hemispheric NDVI variations associated with large-scale climate indices in spring.International Journal of Remote Sensing,24, 2559−2566.
Goodin, D. G., Gao, J. C., & Henebry, G. M. (2004). The effect of solar illumination angle and sensor view angle on observed patterns of spatial structure in tallgrass prairie. IEEE Transactions on Geoscience and Remote Sensing,42, 154−165.
Grace, J., Nichol, C., Disney, M., Lewis, P., Quaife, T., & Bowyer, P. (2007). Can we measure terrestrial photosynthesis from space directly, using spectral reflectance andfluorescence?Global Change Biology,13, 1484−1497.
Guo, J. M., & Trotter, C. M. (2006). Estimating photosynthetic light-use efficiency using the photochemical reflectance index: The effects of short-term exposure to elevated CO2and low temperature.International Journal of Remote Sensing,20, 4677−4684.
Harte, J., Torn, M. S., Chang, F. R., Feifarek, B., Kinzig, A. P., Shaw, R., et al. (1995). Global warming and soil microclimate: Results from a meadow-warming experiment. Ecological Applications,5, 132−150.
IPCC (2007). Climate change 2007: The physical science basis. Contribution of working group I. In S. Solomon, D. Qin, M. Manning, Z. Chen, M. Marquis, K. B. Averyt, M. Tignor, & H. L. Miller (Eds.),Fourth Assessment Report of the Intergovernmental Panel on Climate ChangeCambridge, UK: Cambridge University Press.
Jonasson, S. (1988). Evaluation of the point intercept method for the estimation of plant biomass.Oikos,52, 101−106.
Kerr, J. T., & Ostrovsky, M. (2003). From space to species: Ecological applications for remote sensing.Trends in Ecology & Evolution,18, 299−305.
Larcher, W. (1995). Photosynthesis as a tool for indicating temperature stress events. In E. D. Schulze, & M. M. Caldwell (Eds.),Ecophysiology of photosynthesis(pp. 261−277).
Berlin: Springer.
Llorens, L., Peñuelas, J., Beier, C., Emmett, B., Estiarte, M., & Tietema, A. (2004). Effects of an experimental increase of temperature and drought on the photosynthetic performance of two ericaceous shrub species along a north–south European gradient.Ecosystems,7, 613−624.
Martin, R. E., Asner, G. P., & Sack, L. (2007). Genetic variation in leaf pigment, optical and photosynthetic function among diverse phenotypes ofMetrosideros polymorpha grown in a common garden.Oecologia,151, 387−400.
Maxwell, K., & Johnson, G. N. (2000). Chlorophyllfluorescence—A practical guide. Journal of Experimental Botany,51, 659−668.
Middleton, E. M. (1991). Solar zenith angle effects on vegetation indexes in tallgrass prairie.Remote Sensing of Environment,38, 45−62.
Moya, I., Silvestri, M., Vallon, O., Cinque, G., & Bassi, R. (2001). Time-resolved fluorescence analysis of the photosystem II antenna proteins in detergent micelles and liposomes.Biochemistry,40, 12552−12561.
Moya, I., Camenen, L., Evain, S., Goulas, Y. Z., Cerovic, G., Latouche, G., et al. (2004). A new instrument for passive remote sensing: 1. Measurements of sunlight-induced chlorophyllfluorescence.Remote Sensing of Environment,91, 186−197.
Peñuelas, J., & Filella, I. (1998). Visible and near-infrared reflectance techniques for diagnosing plant physiological status.Trends in Plant Science,3, 151−156.
Peñuelas, J., Filella, I., & Gamon, J. A. (1995). Assessment of photosynthetic radiation-use efficiency with spectral reflectance.New Phytologist,131, 291−296.
Peñuelas, J., Llusia, J., Piñol, J., & Filella, I. (1997). Photochemical reflectance index and leaf photosynthetic radiation-use-efficiency assessment in Mediterranean trees. International Journal of Remote Sensing,18, 2863−2868.
Peñuelas, J., Gordon, C., Llorens, L., Nielsen, T., Tietema, A., Beier, C., et al. (2004). Nonintrusivefield experiments show different plant responses to warming and drought among sites, seasons, and species in a north–south European gradient. Ecosystems,7, 598−612.
Peñuelas, J., Prieto, P., Beier, C., Cesaraccio, C., de Angelis, P., de Dato, G., et al. (2007). Response of plant species richness and primary productivity in shrublands along a north–south gradient in Europe to seven years of experimental warming and drought. Reductions in primary productivity in the heat and drought year of 2003. Global Change Biology,13, 2563−2581.
Pontailler, J. Y., Hymus, G. J., & Drake, B. G. (2003). Estimation of leaf area index using ground-based remote sensed NDVI measurements: Validation and comparison with two indirect techniques.Canadian Journal of Remote Sensing,29, 381−387.
Prince, S. D. (1991). A model of regional primary production for use with coarse resolution satellite data.International Journal of Remote Sensing,12, 1313−1330.
Rascher, U., Liebig, M., & Lüttge, U. (2000). Evaluation of instant light-response curves of chlorophyll fluorescence parameters obtained with a portable chlorophyll fluorometer on site in thefield.Plant, Cell and Environment,23, 1397−1405.
Ruimy, A., Saugier, B., & Dedieu, G. (1994). Methodology for the estimation of terrestrial net primary production from remotely sensed data.Journal of Geophysical Research, 99, 5263−5284.
Running, S. W., Baldocchi, D. D., Turner, D. P., Gower, S. T., Bakwin, P. S., & Hibbard, K. A. (1999). A global terrestrial monitoring network integrating towerfluxes,flask
sampling, ecosystem modelling and EOS satellite data. Remote Sensing of Environment,70, 108−127.
Schreiber, U., Bilger, W., & Neubauer, C. (1995). Chlorophyll fluorescence as a nonintrusive indicator for rapid assessment of in vivo photosynthesis. In E. -D. Schulze, & M. M. Caldwell (Eds.),Ecophysiology of photosynthesis(pp. 49−70).
Berlin-Heidelberg: Springer-Verlag.
Sims, D. A., & Gamon, J. A. (2002). Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and develop-mental stages.Remote Sensing of Environment,81, 337−354.
Sims, D. A., Luo, H., Hastings, S., Oechel, W. C., Rahman, A. F., & Gamon, J. A. (2006). Parallel adjustments in vegetation greenness and ecosystem CO2exchange in
response to drought in a Southern California chaparral ecosystem.Remote Sensing of Environment,103, 289−303.
Stylinski, C. D., Gamon, J. A., & Oechel, W. C. (2002). Seasonal patterns of reflectance indices, carotenoid pigments and photosynthesis of evergreen chaparral species. Oecologia,131, 366−374.
Ustin, S. L., Wessman, C. A., Curtiss, B., Kasischke, E., Way, J. -B., & Vanderbilt, V. C. (1991). Opportunities for using the EOS imaging spectrometers and synthetic aperture radar in ecological models.Ecology,72, 1934−1945.
Walters, R. G., & Horton, P. (1991). Resolution of components of non-photochemical chlorophyllfluorescence quenching in barley leaves.Photosynthesis Research,27, 121−133.