HASIL DAN PEMBAHASAN
Keadaan Lokasi Penelitian
Desa Sukagalih merupakan salah satu desa penghasil sayuran di Kecamatan Mega Mendung. Desa Sukagalih terletak pada ketinggian ± 900-1500 meter dpl dengan topografi berbukit-bukit, curah hujan diatas 150 mm/bulan, tanah latosol coklat dan bertekstur gembur sesuai untuk budi daya sayuran. Keadaan ini dimanfaatkan oleh masyarakat, khususnya petani untuk mengusahakan berbagai tanaman sayuran. Jenis tanaman sayuran yang banyak diusahakan diantaranya adalah kubis, tomat, wortel, cabai, caisin, kacang panjang, ketimun dan buncis. Selain tanaman sayuran, juga ditanam tanaman pangan antara lain padi, jagung, ubi kayu dan ubi jalar. Tanaman buah-buahan yang cukup luas dibudidayakan adalah pepaya. Hasil pertanian tersebut umumnya dijual kepada pedagang pengumpul di desa, namun terdapat juga yang dijual langsung ke konsumen.
Sistem budi daya tanaman sayuran umumnya masih dilakukan secara konvensional baik secara monokultur maupun tumpangsari. Di Desa Sukagalih terdapat beberapa petani yang telah melaksanakan sistem budi daya sayuran organik. Luas lahan budi daya sayuran organik masih relatif sempit.
Hama yang sering ditemukan menyerang tanaman kubis di Desa Sukagalih diantaranya adalah P. xylostella, C. pavonana dan P. vittata, sedangkan
penyakitnya adalah yang disebabkan oleh X. campestris, P. brassicae, E. carotovora dan Alternaria spp. Praktik pengendalian hama dan penyakit yang
dilakukan oleh petani sayuran organik diantaranya adalah menanam secara tumpangsari, pergiliran tanaman, mengambil kelompok telur dan larva hama secara langsung, meramu insektisida botani dan menanam tanaman berbunga di sekitar lahan. Petani konvensional dalam mengendalikan hama dan penyakit menggunakan berbagai merek dagang pestisida yang tersedia pada kios pertanian di desa. Tutu (2002) melaporkan bahwa 68% petani di Kecamatan Cisarua dan Mega Mendung menyemprot secara berjadwal, 18% petani menyemprot apabila ada kerusakan dan 14% petani menyemprot apabila ada imago di pertanaman.
Hama Kubis
Hama utama yang ditemukan pada pertanaman kubis yaitu P. xylostella dan C. pavonana. Hama lainnya yang juga ditemukan yaitu Gryllotalpa sp., Agrotis ipsilon Hufn. (Lepidoptera: Noctuidae), P. vittata dan Spodoptera spp. (Lepidoptera: Noctuidae).
Plutella xylostella
Populasi larva P. xylostella pada MK tampak di atas AE pada umur 14 hst pada perlakuan OM (0,9 larva /tanaman), OT (1,8 larva/tanaman), KM (1,2 larva/tanaman) dan KT (1,7 larva /tanaman). Pada perlakuan input rendah populasi larva di bawah AE yaitu LM (0,2 larva/tanaman) dan LT (0,1 larva /tanaman) (Gambar 3 dan La mpiran 1). Hal ini disebabkan sejak ditanam sampai 14 hst belum dilakukan pengendalian. Setelah dilakukan pengendalian secara mekanik pada perlakuan organik, populasi larva P. xylostella tampak menurun hingga di bawah AE. Ini menunjukkan bahwa pengendalian secara mekanik dapat memberikan hasil pengendalian yang cukup efektif. Pengendalian secara mekanik selain dapat menekan populasi hama juga berpengaruh baik terhadap lingkungan dan produksi kubis karena tidak terjadi pencemaran. Sementara itu padaperlakua n konvensional yang menggunakan pestisida sintetik, populasi larva P. xylostella
0,0 0,2 0,4 0,6 0,8 1,0 1,2 1,4 1,6 1,8 2,0 14 21 28 35 42 49 56 63 70 OM OT LM LT KM KT
Gambar 3 Rata-rata populasi larva P. xylostella pada tanaman kubis pada MK. Umur tanaman (hst)
masih di atas AE yakni pada perlakuan KM (0,6 larva/tanaman) pada umur 21 hst dan KT (0,7 larva /tanaman) pada umur 42 hst. Hal ini kemungkinan disebabkan oleh larva P. xylostella tersebut resisten terhadap insektisida yang digunakan. Sastrosiswojo (1992) menyatakan bahwa awal tahun 90-an hama P. xylostella menunjukkan resistensi terhadap berbagai jenis insektisida yang sering diaplikasikan untuk pengendalian hama tersebut. Kemungkinan hal lain yang dapat terjadi akibat dari penggunaan insektisida berjadwal adalah matinya musuh alami. Pengaruh buruk penggunaan pestisida diantaranya adalah resistensi dan matinya musuh alami (Flint dan van den Bosch 1981; Untung 1992; Oka 1995). Populasi larva P. xylostella pada MH di atas AE terjadi pada perlakuan OM (1,2 larva /tanaman) dan KT (0,8 larva /tanaman) pada umur 40 hst, sedangkan pada perlakuan input rendah populasi larva masih di bawah AE (Gambar 4 dan Lampiran 2). Pada perlakuan organik selain pengendalian secara mekanik juga dikendalikan dengan insektisida botani (mengandung ekstrak S. mahogani dan A. odorata). Tindakan ini dapat membatasi perkembangan larva.
0,0 0,2 0,4 0,6 0,8 1,0 1,2 1,4 13 19 26 33 40 47 61 68 74 OM OT LM LT KM KT
Populasi P. xylostella pada perlakuan konvensional di atas AE walaupun dilakukan aplikasi insektisida secara berjadwal baik pada MH maupun MK. Hal tersebut menunjukkan bahwa penggunaan insektisida kurang dapat menekan perkembangan larva P. xylostella secara permanen. Hal ini kemungkinan karena larva P. xylostella telah resisten terhadap insektisida yang digunakan yakni
Umur tanaman (hst)
Populasi (larva/tanaman)
berbahan aktif deltametrin dan profenofos. Terjadinya resistensi ini kemungkinan disebabkan aplikasi insektisida tersebut oleh petani kubis di lokasi penelitian kurang bijaksana (dari musim ke musim dengan dosis dan frekwensi tinggi). Merujuk hasil survei Tutu (2002) bahwa 68% petani kubis di Kecamatan Cisarua dan Mega Mendung, melakukan penyemprotan berjadwal. Kasus resistensi larva P. xylostella terhadap bahan aktif ya ng sama dan bahan aktif yang lainnya telah banyak dilaporkan di beberapa tempat seperti: bahan aktif sipermetrin, fenvalerat dan deltametrin di India (Saxena et al. 1989); bahan aktif fenvalerat dan deltametrin di Korea (Kim et al. 1990). Di Indonesia tela h dilaporkan, adanya kasus resistensi P. xylostella terhadap DDT (Ankersmit 1953 dalam Oka 1995); P. xylostella strain Lembang, Pacet, Kopeng dan Tawangmangu tesisten terhadap deltametrin, sipermetrin dan fenvalerat (Adiputra 1984); P. xylostella strain Lembang resisten terhadap deltametrin dan asefat (Sastrosiswojo 1992).
Pada perlakuan input rendah, selama perkembangan tanaman kubis baik pada MK maupun MH, populasi larva P. xylostella tidak melebihi AE, sehingga tidak perlu aplikasi insektisida untuk pengendalian. Akan tetapi aplikasi insektisida terpaksa dilakukan untuk mengendalikan larva C. pavonana pada MK, saat tanaman berumur 35 hst dan pada MH saat tanaman berumur 61 dan 68 hst. Dengan mengurangi aplikasi insektisida terlihat bahwa populasi larva P. xylostella dapat dipertahankan pada batas tidak merugikan. Hal ini kemungkinan juga
disebabkan oleh peran musuh alami dalam menekan perkembangan larva P. xylostella. Di Jawa Barat D. semiclausum dan Apanteles sp. dapat menekan
populasi larva P. xylostella dengan tingkat parasitisasi 79%-88% (Kartosuwondo 1994).
Secara umum, populasi larva P. xylostella pada perlakuan LT lebih rendah dibandingkan dengan perlakuan LM baik pada MK maupun MH. Hal ini menunjukkan adanya pengaruh tanaman tomat sebagai repellent (penolak) dari bahan kimia yang dihasilkan untuk menghambat imago P. xylostella meletakkan telur pada tanaman kubis (Sastrosiswojo et al. 2000).
Crocidolomia pavonana
Hama perusak krop C. pavonana ditemukan pada MK sejak tanaman berumur 35 hst dan pada MH sejak tanaman berumur 26 hst. Populasi C. pavonana pada MK tertinggi terjadi pada perlakuan KT (3,7 larva/tanaman), KM (2,3 larva/tanaman), LM (1,8 larva/tanaman) dan LT (1,3 larva /tanaman), sedangkan pada perlakuan OM dan OT 0,1 larva /tanaman (Gambar 5 dan Lampiran 3). Hal ini menunjukkan aplikasi insektisida berjadwal pada perlakuan konvensional kurang efektif. Keadaan larva C. pavonana yang berada di bawah daun dan selanjutnya masuk ke dalam krop, menyulitkan insektisida kontak untuk mengenai sasaran. Hal ini kemungkinan membuat populasi larva C. pavonana dapat berkembang lebih tinggi pada perlakuan konvensional dibandingkan dengan perlakuan organik. Pada perlakuan organik dengan pengendalian secara mekanik, mencari larva C. pavonana relatif lebih mudah walaupun berada di bawah daun dan cara tersebut lebih aman bagi lingkungan.
0 0,5 1 1,5 2 2,5 3 3,5 4 35 42 49 56 63 70 OM OT LM LT KM KT
Populasi larva C. pavonana tertinggi pada setiap perlakuan pada MH berturut-turut adalah sebagai berikut LM (6,4 larva /tanaman), KM (3,8 larva/tanaman), LT (3,2 larva /tanaman), OM (3,1 larva/tanaman), OT (3 larva/tanaman) dan KT 1,4 larva /tanaman (Gambar 6 dan Lampiran 4). Secara umum populasi larva C. pavonana lebih tinggi pada perlakuan konvensional dan input rendah dengan aplikasi insektisida dibandingkan dengan perlakuan organik.
Gambar 5 Rata-rata populasi C. pavonana pada tanaman kubis pada MK.
Populasi (la
rva/tanaman)
0 1 2 3 4 5 6 7 26 33 40 47 61 68 74 OM OT LM LT KM KT
Keadaan ini menunjukkan, penggunan insektisida untuk pengendalian larva C. pavonana kurang efektif. Hal ini kemungkinan disebabkan larva C. pavonana resisten terhadap insektisida yang digunakan. Menurut Untung (1992) salah satu penyebab terjadinya resistensi pada serangga adalah penurunan laju penetrasi insektisida melalui kulit. Keberadaan telur dan larva C. pavonana yang tersembunyi sulit bagi insektisida dan musuh alami untuk mencapainya hal ini dapat diatasi dengan pengendalian mekanik.
Gryllotalpa sp.
Hama perusak akar Gryllotalpa sp. ditemukan pada MK dan MH (Tabel 3). Gejala kerusakan yang ditimbulkan adalah akar muda terpotong hingga tanaman bertahan hidup pada akar tua, tanaman layu dan krop susah terbentuk (Gambar 7). Gryllotalpa sp. merupakan serangga penghuni tanah yang lembab, mempunyai tungkai depan yang lebar berbentuk sekop (Borror et al. 1996). Tungkai depan
Tabel 3 Intensitas kerusakan tanaman kubis oleh Gryllotalpa sp. Intensitas (%)
Pertanaman
OM OT LM LT KM KT
MK 20,87 28,75 4,82 14,82 11,43 15,35
MH 4,72 4,45 4,89 3,90 5,42 5,09
Gambar 6 Rata-rata populasi C. pavonana pada tanaman kubis pada MH.
Populasi (larva/tanaman)
Gambar 7 Gejala kerusakan akar kubis oleh serangan Gryllotalpa sp.
berfungsi sebagai pencabik akar-akar muda untuk dimakan, kerusakan ini akan mengakibatkan tanaman tidak bisa membentuk krop. Kerusakan pada setiap perlakuan bervariasi. Pada MK intensitas kerusakan pada perlakuan organik lebih tinggi dibandingkan perlakuan konvensional dan input rendah sedangkan pada MH hampir tidak berbeda. Pada MK intensitas kerusakan oleh Gryllotalpa sp. pada pertanaman tumpangsari lebih tinggi dibandingkan dengan pertanaman monokultur. Hal ini kemungkinan disebabkan pada pertanaman tumpangsari terdapat inang utama yakni tanaman tomat dan tanah lebih gembur. CABI (2003) menyebutkan bahwa G. hexadactyla Perty merupakan hama perusak akar pada tanaman tomat, jagung dan kedele.
Parasitisasi
Parasitoid yang muncul dari larva P. xylostella pada MK adalah D. semiclausum sedangkan pada MH adalah D. semiclausum dan Apanteles sp.
Persentase parasitisasi sangat bervariasi (Tabel 4). Secara umum, rata-rata parasitisasi pada perlakuan tumpangsari lebih tinggi dari pada monokultur. Hal ini kemungkinan karena imago parasitoid lebih menyukai tanaman yang mempunyai bunga (nektar tanaman tomat). Parasitoid D. semiclausum mengisap nektar pada tumbuhan berbunga sebagai sumber makanannya (Tooker dan Hanks 2000). Idris dan Grafius (1995) melaporkan bahwa nektar bunga dapat meningkatkan lama hidup, keperidian dan persentase parasitisasi serangga parasitoid Hymenoptera.
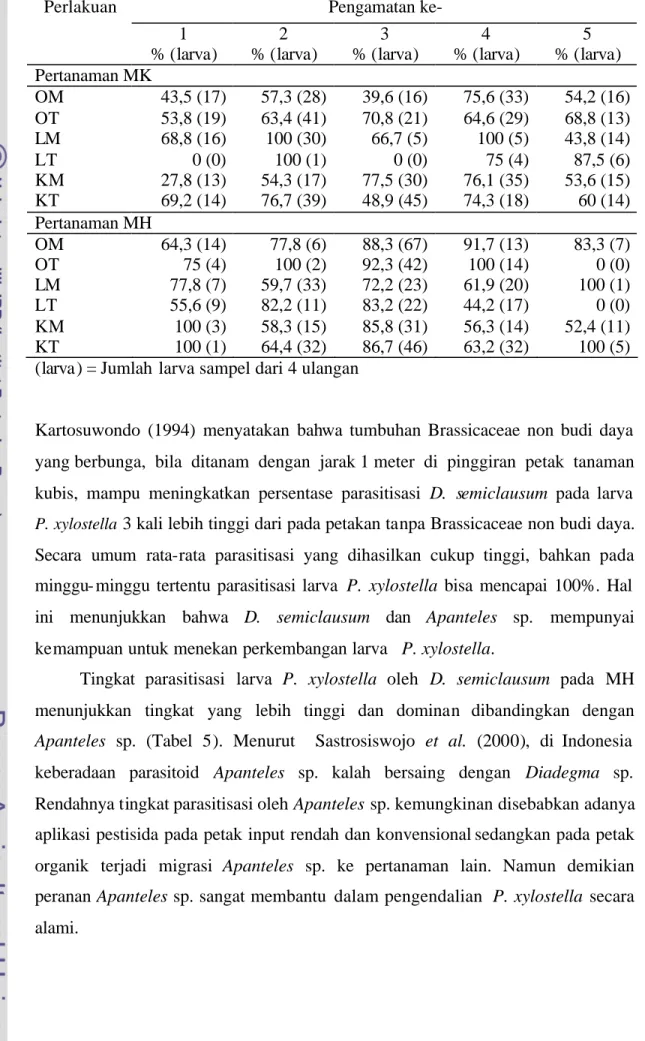
Tabel 4 Rata-rata tingkat parasitisasi larva P. xylostella Pengamatan ke- Perlakuan 1 % (larva) 2 % (larva) 3 % (larva) 4 % (larva) 5 % (larva) Pertanaman MK OM OT LM LT KM KT 43,5 (17) 53,8 (19) 68,8 (16) 0 (0) 27,8 (13) 69,2 (14) 57,3 (28) 63,4 (41) 100 (30) 100 (1) 54,3 (17) 76,7 (39) 39,6 (16) 70,8 (21) 66,7 (5) 0 (0) 77,5 (30) 48,9 (45) 75,6 (33) 64,6 (29) 100 (5) 75 (4) 76,1 (35) 74,3 (18) 54,2 (16) 68,8 (13) 43,8 (14) 87,5 (6) 53,6 (15) 60 (14) Pertanaman MH OM OT 64,3 (14) 75 (4) 77,8 (6) 100 (2) 88,3 (67) 92,3 (42) 91,7 (13) 100 (14) 83,3 (7) 0 (0) LM LT KM KT 77,8 (7) 55,6 (9) 100 (3) 100 (1) 59,7 (33) 82,2 (11) 58,3 (15) 64,4 (32) 72,2 (23) 83,2 (22) 85,8 (31) 86,7 (46) 61,9 (20) 44,2 (17) 56,3 (14) 63,2 (32) 100 (1) 0 (0) 52,4 (11) 100 (5) (larva) = Jumlah larva sampel dari 4 ulangan
Kartosuwondo (1994) menyatakan bahwa tumbuhan Brassicaceae non budi daya yang berbunga, bila ditanam dengan jarak 1 meter di pinggiran petak tanaman kubis, mampu meningkatkan persentase parasitisasi D. semiclausum pada larva
P. xylostella 3 kali lebih tinggi dari pada petakan tanpa Brassicaceae non budi daya. Secara umum rata-rata parasitisasi yang dihasilkan cukup tinggi, bahkan pada minggu- minggu tertentu parasitisasi larva P. xylostella bisa mencapai 100%. Hal ini menunjukkan bahwa D. semiclausum dan Apanteles sp. mempunyai kemampuan untuk menekan perkembangan larva P. xylostella.
Tingkat parasitisasi larva P. xylostella oleh D. semiclausum pada MH menunjukkan tingkat yang lebih tinggi dan dominan dibandingkan dengan Apanteles sp. (Tabel 5). Menurut Sastrosiswojo et al. (2000), di Indonesia keberadaan parasitoid Apanteles sp. kalah bersaing dengan Diadegma sp. Rendahnya tingkat parasitisasi oleh Apanteles sp. kemungkinan disebabkan adanya aplikasi pestisida pada petak input rendah dan konvensional sedangkan pada petak organik terjadi migrasi Apanteles sp. ke pertanaman lain. Namun demikian peranan Apanteles sp. sangat membantu dalam pengendalian P. xylostella secara alami.
Tabel 5 Rata-rata tingkat parasitisasi larva P. xylostella oleh D. semiclausum dan Apanteles sp. pada MH Pengamatan ke- Perlakuan 1 (%) 2 (%) 3 (%) 4 (%) 5 (%) D. s A. sp D. s A. sp D. s A. sp D. s A. sp D. s A. sp OM 24,4 39,9 77,8 0 84,6 3,7 91,7 0 83,3 0 OT 50,0 25,0 100 0 84,8 7,5 100 0 0 0 LM 33,3 44,4 59,7 0 68,5 3,7 61,9 0 100 0 LT 55,5 0 75,5 6,7 69,9 13,3 44,2 0 0 0 KM 100 0 58,3 0 76,0 9,8 46,2 0 52,4 0 KT 0 100 56,1 8,3 81,0 5,6 63,2 0 100 0 D. s = D. semiclausum, A. sp = Apanteles sp.
Selama penelitian, tidak diperoleh parasitoid dari larva C. pavonana yang dipelihara. Hal ini kemungkinan disebabkan musuh alami dari C. pavonana seperti Eriborus argenteopilosus Cemeron (Hymenoptera: Ichneumonidae) dan Sturmia inconspicuoides Bar. (Diptera: Tachinidae) tidak ada di lahan penelitian. Kemungkinan lain karena parasitoid sulit menemukan larva yang berada di bawah daun atau di dalam krop. Menurut Othman (1982), kedua parasitoid tersebut tingkat parasitisasinya rendah.
Penyakit Kubis
Busuk Hitam
Penyakit busuk hitam yang disebabkan oleh X. campestris pv. campestris ditemukan berkembang selama pertumbuhan kubis baik pada MK maupun MH. Patogen menyerang tanaman kubis pada semua sistem pertanaman dengan pola yang sama yakni terjadi peningkatan dan penurunan intensitas serangan dengan kisaran yang tidak jauh berbeda (Gambar 8a, 8b, Lampiran 5 dan 6). Hal ini kemungkinan disebabkan sifat X. campestris pv. campestris yang mudah menyebar dari tanah ke daun melalui percikan air hujan (Semangun 2001) dan penyiraman yang sama pada semua perlakuan. X. campestris pv. campestris dapat bertahan di dalam tanah dan sisa tanaman sakit (Semangun 2000). Penurunan intensitas pada MK maupun MH pada semua perlakuan disebabkan adanya tindakan sanitasi, yakni membuang bagian tanaman yang sakit, hingga penilaian pada waktu pengamatan menur un.
0 5 10 15 20 25 13 19 26 33 40 47 61 68 74 OM OT LM LT KM KT
Secara umum intensitas serangan X. campestris pv. campestris pada perlakuan monokultur lebih tinggi dari perlakuan tumpangsari. Hal ini kemungkinan karena kondisi iklim mikro pada perlakuan monokultur lebih hangat. Tanaman tomat pada perlakuan tumpangsari membuat iklim mikro lebih sejuk dibandingkan dengan perlakuan monokultur. Selain itu tanaman tomat juga sebagai penghalang gesekan antara daun kubis dan penghalang percikan air langsung ke permukaan tanah atau ke tanaman kubis. Penyakit busuk hitam yang disebabkan oleh X. campestris pv. campestris sering berjangkit pada tanaman kubis dengan kondisi lingkungan hangat dan kelengasan udara tinggi (Parmadi dan Sastrosiswojo 1993). 0 5 10 15 20 25 30 35 40 45 50 14 21 28 35 42 49 56 63 70 OM OT LM LT KM KT
Gambar 8 Rata-rata intensitas serangan X. campestris pv. campestris pada tanaman kubis pada MK (a) dan MH (b).
. (b) (a) Umur tanaman (hst) Umur tanaman (hst) Intensitas serangan (% ) Intensitas serangan (%)
Akar Gada
Kejadian penyakit akar gada yang disebabkan oleh P. brassicae ditemukan tertinggi pada pertanaman kubis yang dibudidayakan secara organik. Gejala yang terlihat adalah tanaman kubis layu pada waktu siang hari bila panas terik dan kembali normal pada pagi hari, akar yang dihasilkan sedikit dan membengkak (Gambar 9a dan 9b). Bila akar dicabut terlihat membengkak, berumbi menyerupai gada (Direktorat Perlindungan Hortikultura 2002). Semangun (2000) menyatakan bahwa akar-akar yang terinfeksi P. brassicae sel-selnya membelah dan membesar hingga terbentuk seperti gada. Selanjutnya dinyatakan bahwa rusaknya susunan jaringan akar menyebabkan rusaknya jaringan pengangkut air dan hara.
(a) (b)
Gambar 9 Gejala penyakit akar gada (a) layu (b) gada pada akar
Timbulnya serangan diduga karena pupuk kandang yang digunakan telah mengandung inokulum P. brassicae. Pupuk kandang yang telah terinvestasi P. brassicae dapat menyebarkan penyakit akar gada (Suryaningsih 1981; Parmadi dan Sastrosiswojo 1993). Kejadian penyakit akar gada yang disebabkan oleh P. brassicae berkorelasi positif dengan jumlah pupuk kandang yang diaplikasikan. Semakin banyak jumlah pupuk kandang yang diaplikasikan semakin tinggi kejadian penyakit akar gada pada tanaman kubis (Tabel 6 dan Tabel 1).
Tabel 6 Rata-rata kejadian penyakit akar gada pada tanaman kubis Rata-rata perlakuan (%)
Pertanaman
OM OT LM LT KM KT
MK 21,8 23,2 9,3 14,8 6,8 7,5
Kejadian penyakit akar gada pada MK (menggunakan pupuk kandang) lebih tinggi dibandingkan dengan MH (menggunakan kompos). Hal ini kemungkinan kompos yang digunakan pada MH mengandung mikroorganisme yang dapat menekan perkembangan P. brassicae. Kompos mengandung berbagai macam mikroorganisme dari golongan bakteri, aktinomisetes dan cendawan (Wibisono 2004). Kemungkinan hal lain yang menyebabkan tingginya kejadian penyakit akar gada pada MK dibandingkan dengan MH adalah faktor cuaca.
Hama Tomat
Selama penelitian, hama yang ditemukan pada pertanaman tomat adalah Epilachna varivestis Muls. (Coleoptera: Coccinellidae), Liriomyza bryoniae Kalt. (Diptera: Agromyzidae), Bemisia tabaci Genn. (Hemiptera: Aleyrodidae) dan Helicoverpa armigera Hubn. (Lepidoptea: Noctuidae). Kerusakan yang ditimbulkan pada MK oleh masing- masing hama yang ditemukan sangat kecil. Kelimpahan populasi B. tabaci menjadi pengamatan khusus sehubungan dengan kamampuannya sebagai vektor geminivirus. Meningkatnya populasi B. tabaci pada MK seiring dengan bertambahnya umur tanaman sedangkan jumlah populasi pada ketiga perlakuan tidak terlalu berbeda (Gambar 10 dan Lampiran 7).
Hama E. varivestis dan L. bryoniae pada MH menyerang pada awal pertanaman dan tingkat kerusakannya sangat rendah. Perkembangan populasi
0 1 2 3 4 5 6 7 8 9 10 21 28 35 42 49 56 63 Organik Input Rendah Konvensional Populasi ( individu /tanaman) Umur tanaman (hst)
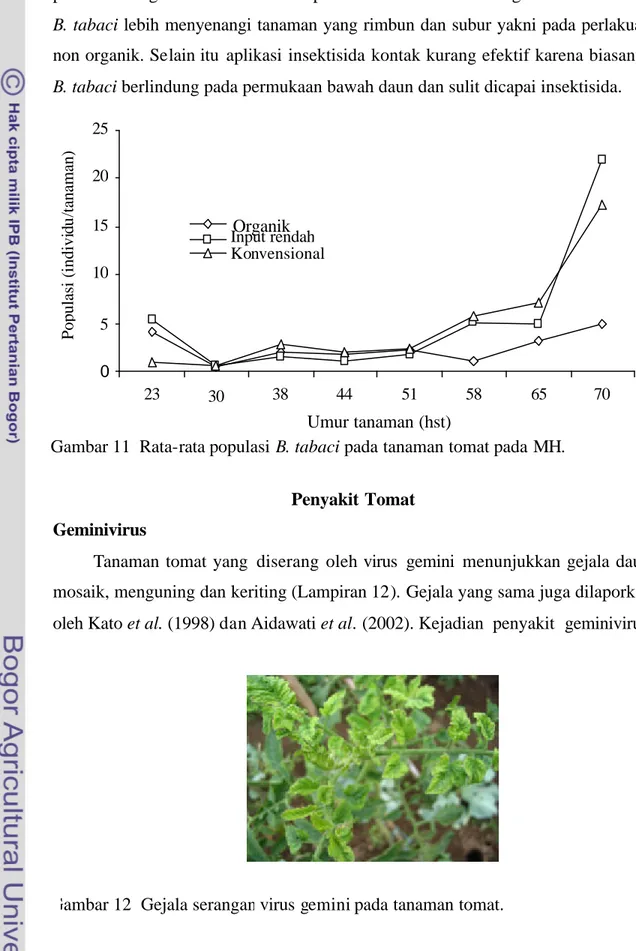
B. tabaci terlihat lebih stabil pada MH (Gambar 11). Hal ini kemungkinan karena seringnya turun hujan yang memperlambat perkembangan dan penyebaran B. tabaci. Populasi pada perlakuan non organik lebih tinggi dibandingkan dengan perlakuan organik baik MK maupun MH. Hal ini kemungkinan disebabkan B. tabaci lebih menyenangi tanaman yang rimbun dan subur yakni pada perlakuan non organik. Selain itu aplikasi insektisida kontak kurang efektif karena biasanya B. tabaci berlindung pada permukaan bawah daun dan sulit dicapai insektisida.
Penyakit Tomat Geminivirus
Tanaman tomat yang diserang oleh virus gemini menunjukkan gejala daun mosaik, menguning dan keriting (Lampiran 12). Gejala yang sama juga dilaporkan oleh Kato et al. (1998) dan Aidawati et al. (2002). Kejadian penyakit geminivirus
0 5 10 15 20 25 23 30 38 44 51 58 65 70 Organik Input rendah Konvensional Umur tanaman (hst)
Gambar 11 Rata-rata populasi B. tabaci pada tanaman tomat pada MH.
Populasi (
indivi
du
/tanaman)
pada tanaman tomat pada setiap perlakuan tidak terlalu berbeda. Pada MK umur 49 hst, kejadian penyakit geminivirus hampir mencapai 100% pada semua perlakuan (Gambar 13a dan Lampiran 8). Pada MH umur 70 hst, kejadian penyakit geminivirus tidak jauh berbeda yakni pada perlakuan input rendah 62,5%, organik 58% dan konvensional 53,2% (Gambar 13b dan Lampiran 8).
Kejadian penyakit geminivirus yang tidak terlalu jauh berbeda pada ketiga perlakuan, kemungkinan karena sumber inokulum virus gemini dan B. tabaci yang infektif telah berada pada semua perlakuan. Untuk me nyebarkan virus gemini hanya memerlukan jumlah vektor yang sedikit. Satu individu B. tabaci yang infektif sudah dapat menularkan virus (Mehta et al. 1994) dan penularan semakin efektif seiring dengan meningkatnya jumlah B. tabaci (Aidawati et al. 2002).
0 20 40 60 80 100 120 14 21 28 35 42 49 Umur tanaman (hst) Organik Input rendah Konvensional 0 20 40 60 80 23 30 38 44 51 58 65 70 Umur tanaman (hst) Organik Input rendah Konvensional Gambar 13 Rata-rata luas serangan penyakit geminivirus pada tanaman tomat pada
MK (a) dan MH (b). (a) (b) Kej adian penyakit (%) Kejadian penyakit (%)
Aplikasi insektisida pada perlakuan konvensional dan seringnya turun hujan pada MH hanya mampu memperlambat perkembangan dan penyebaran populasi B. tabaci. Pada lahan non organik yang lebih rimbun, peluang kejadian penyakit akan lebih besar karena aktifitas hidup B. tabaci cenderung pada kondisi lahan tersebut. Sudiono (2001) menyatakan bahwa penularan penyakit geminivirus pada tanaman inang melalui serangga vektor (B. tabaci) menunjukkan masa inkubasi antara 15-29 hari setelah inokulasi dan kejadian penyakit antara 20-100%.
Bercak Daun
Penyakit bercak daun pada tanaman tomat yang disebabkan oleh A. solani menunjukkan gejala bercak bulat atau bersudut, coklat tua sampai hitam dan terdapat jalur klorotik (Gambar 14). Pada MK ditemukan menyerang tanaman tomat hanya pada awal pertumbuhan dengan intensitas serangan rendah (< 5%), namun pada MH intensitas serangan jauh lebih tinggi. Hal ini kemungkinan disebabkan oleh pengaruh cuaca pada MH yang cocok untuk perkembangan A. solani. Semangun (2000) menyatakan bahwa A. solani membentuk banyak
Gambar 14 Gejala serangan A. solani pada tanaman tomat.
konidium bila cuaca lembab. Selanjutnya konidia menyebar ke bagian tanaman yang lain dengan bantuan air hujan dan angin. Serangan A. solani pada MH sudah terlihat sejak tanaman tomat berumur 23 hst, intensitas serangan meningkat sampai umur 51 hst kemudian turun pada umur 58 hst pada semua perlakuan (Gambar 15 dan Lampiran 9). Agrios (1997) menyatakan bahwa tanaman tomat pada awal pertumbuhan rentan, kemudian agak tahan pada fase pertumbuhan dan kembali
0 5 10 15 20 25 30 35 40 45 50 23 30 38 44 51 58 65 70 77 Umur tanaman (hst) Intensitas serangan (%) Organik Input rendah Konvensional
rentan pada fase pemasakan, terhadap bercak daun (early blight) dan busuk daun (late blight). Intensitas serangan pada ketiga perlakuan sampai umur tanaman 58 hst tidak terlalu jauh berbeda. Intensitas serangan menurun setelah umur tanaman 51 hst disebabkan adanya tindakan sanitasi yakni membuang bagian tanaman yang sakit pada semua perlakuan. Bila dibandingkan dengan perlakuan lain, pada perlakuan organik intensitas serangan cenderung menurun terus setelah tanaman berumur 51 hst. Hal ini disebabkan oleh bagian tanaman (relung) yang sebelumnya menjadi tempat berkembang A. solani didominasi oleh perkembangan P. infestans. Pada perlakuan non organik, setelah menurun pada umur 58 hst intensitas serangan meningkat sampai umur 70 hst yang kemudian menurun kembali. Hal ini menunjukkan aplikasi fungisida kurang mempengaruhi penekanan serangan A. solani. Dengan kondisi cuaca dan inang yang sama terlihat dominasi relung oleh P. infestans lebih cepat dibandingkan dengan A. solani.
Busuk Daun
Penyakit busuk daun pada tanaman tomat yang disebabkan oleh P. infestans menunjukkan gejala pada daun hawar hitam kecoklatan (Gambar 16a), buah hawar coklat tua, keras dan berkerut (Gambar 16b). Serangan P. infestans pada MK mulai terlihat sejak tanaman berumur 14 hst kemudian intensitas serangan Gambar 15 Rata-rata intensitas serangan A. solani pada tanaman tomat pada MH.
(a) (b)
Gambar 16 Gejala serangan P. infestans pada (a) daun dan (b) buah tomat.
meningkat seiring dengan bertambahnya umur tanaman (Gambar 17 dan Lampiran 10). Intensitas serangan P. infestans pada MK tidak terlalu berbeda antara perlakuan. Intensitas tertinggi pada setiap perlakuan sebagai berikut organik (82,1%), input rendah (76,5%) dan konvensional (75,7%). Pada kondisi ini, tanaman tomat masih dapat berproduksi karena P. infestans menyerang sebagian besar pada daun sedangkan serangan pada buah masih sedikit.
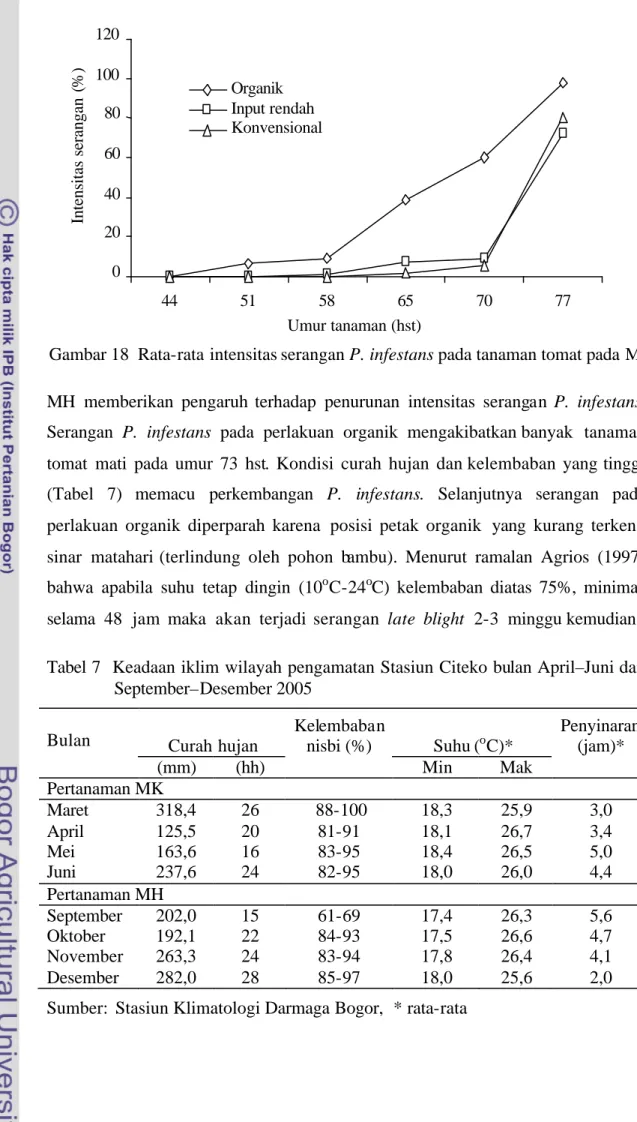
Serangan P. infestans pada MH mulai tampak pada umur 51 hst dan meningkat seiring dengan bertambahnya umur tanaman (Gambar 18 dan Lampiran 10). Pada umur 70 hst, intensitas serangan P. infestans pada perlakuan organik (60,2%) sangat tinggi dibandingkan dengan perlakuan input rendah (9%) dan konvensional (5,4%). Hal ini menunjukkan bahwa penggunaan fungisida pada
0 10 20 30 40 50 60 70 80 90 14 21 28 35 42 49 56 63 70 Organik Input Rendah Komvensional Umur tanaman (hst)
Gambar 17 Rata-rata intensitas serangan P. infestans pada tanaman tomat pada MK.
Intensitas serangan
(
%
MH memberikan pengaruh terhadap penurunan intensitas serangan P. infestans. Serangan P. infestans pada perlakuan organik mengakibatkan banyak tanaman tomat mati pada umur 73 hst. Kondisi curah hujan dan kelembaban yang tinggi (Tabel 7) memacu perkembangan P. infestans. Selanjutnya serangan pada perlakuan organik diperparah karena posisi petak organik yang kurang terkena sinar matahari (terlindung oleh pohon bambu). Menurut ramalan Agrios (1997) bahwa apabila suhu tetap dingin (10oC-24oC) kelembaban diatas 75%, minimal selama 48 jam maka akan terjadi serangan late blight 2-3 minggu kemudian. Tabel 7 Keadaan iklim wilayah pengamatan Stasiun Citeko bulan April–Juni dan
September–Desember 2005
Bulan Curah hujan Kelembaban nisbi (%) Suhu (o C)* Penyinaran (jam)* (mm) (hh) Min Mak Pertanaman MK Maret 318,4 26 88-100 18,3 25,9 3,0 April 125,5 20 81-91 18,1 26,7 3,4 Mei 163,6 16 83-95 18,4 26,5 5,0 Juni 237,6 24 82-95 18,0 26,0 4,4 Pertanaman MH September 202,0 15 61-69 17,4 26,3 5,6 Oktober 192,1 22 84-93 17,5 26,6 4,7 November 263,3 24 83-94 17,8 26,4 4,1 Desember 282,0 28 85-97 18,0 25,6 2,0
Sumber: Stasiun Klimatologi Darmaga Bogor, * rata-rata 0 20 40 60 80 100 120 44 51 58 65 70 77 Organik Input rendah Konvensional Intensitas serangan ( % ) Umur tanaman (hst)
Selanjutnya dinyatakan jika dalam periode tersebut terjadi hujan, embun dan kelembaban nisbi mendekati jenuh selama beberapa jam maka akan terjadi epidemi late blight. Crosier (1934) dalam Semangun (2000) menyatakan bahwa cendawan hanya membentuk sporangium bila kelembaban udara lebih dari 91% dan paling baik pada 100% dengan suhu 18o–22oC. Lebih lanjut dinyatakan bahwa busuk daun tomat merupakan masalah berat di dataran tinggi hanya pada musim hujan karena perkembangan P. infestans memerlukan kelembaban nisbi tinggi dan suhu rendah.
Tinggi Tanaman Tomat
Rata-rata tinggi tanaman tomat antara perlakuan baik pada MK maupun MH tidak terlalu jauh berbeda. Pada MK tanaman tertinggi terdapat pada perlakuan konvensional yaitu 136,6 cm, organik 132,7 cm dan input rendah 126,6 cm (Gambar 19 dan Lampiran 11). Hal ini tampaknya disebabkan pada semua perlakuan tanaman tomat terserang penyakit (geminivirus dan busuk daun) yang dapat mengha mbat pertumbuhan tanaman.
0 20 40 60 80 100 120 140 160 14 21 28 35 42 49 56 63 Organik Input rendah Konvensional
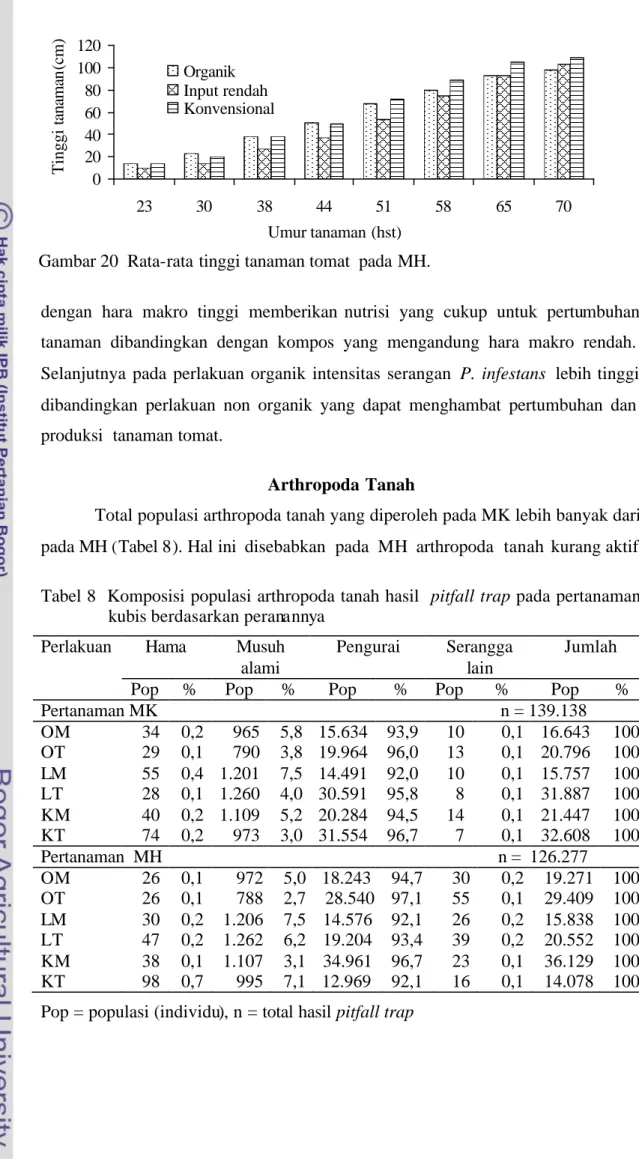
Rata-rata tinggi tanaman tomat pada MH sejak awal pertumbuhan sampai dengan umur 58 hst, pada perlakuan organik dan konvensional lebih tinggi dibandingkan dengan perlakuan input rendah (Gambar 20 dan Lampiran 11). Pada umur 70 hst, tinggi tanaman tomat non organik lebih tinggi dibandingkan organik yakni pada perlakuan konvensional 109,4 cm dan input rendah 102,9 cm sementara organik hanya 98,2 cm. Hal ini disebabkan aplikasi pupuk anorganik Gambar 19 Rata-rata tinggi tanaman tomat pada MK.
Umur tanaman (hst)
Tinggi tanaman
(cm
0 20 40 60 80 100 120 23 30 38 44 51 58 65 70 Organik Input rendah Konvensional
dengan hara makro tinggi memberikan nutrisi yang cukup untuk pertumbuhan tanaman dibandingkan dengan kompos yang mengandung hara makro rendah. Selanjutnya pada perlakuan organik intensitas serangan P. infestans lebih tinggi dibandingkan perlakuan non organik yang dapat menghambat pertumbuhan dan produksi tanaman tomat.
Arthropoda Tanah
Total populasi arthropoda tanah yang diperoleh pada MK lebih banyak dari pada MH (Tabel 8). Hal ini disebabkan pada MH arthropoda tanah kurang aktif Tabel 8 Komposisi populasi arthropoda tanah hasil pitfall trap pada pertanaman
kubis berdasarkan peranannya Hama Musuh alami Pengurai Serangga lain Jumlah Perlakuan
Pop % Pop % Pop % Pop % Pop %
Pertanaman MK n = 139.138 OM 34 0,2 965 5,8 15.634 93,9 10 0,1 16.643 100 OT 29 0,1 790 3,8 19.964 96,0 13 0,1 20.796 100 LM 55 0,4 1.201 7,5 14.491 92,0 10 0,1 15.757 100 LT 28 0,1 1.260 4,0 30.591 95,8 8 0,1 31.887 100 KM 40 0,2 1.109 5,2 20.284 94,5 14 0,1 21.447 100 KT 74 0,2 973 3,0 31.554 96,7 7 0,1 32.608 100 Pertanaman MH n = 126.277 OM 26 0,1 972 5,0 18.243 94,7 30 0,2 19.271 100 OT 26 0,1 788 2,7 28.540 97,1 55 0,1 29.409 100 LM 30 0,2 1.206 7,5 14.576 92,1 26 0,2 15.838 100 LT 47 0,2 1.262 6,2 19.204 93,4 39 0,2 20.552 100 KM 38 0,1 1.107 3,1 34.961 96,7 23 0,1 36.129 100 KT 98 0,7 995 7,1 12.969 92,1 16 0,1 14.078 100 Pop = populasi (individu), n = total hasil pitfall trap
Gambar 20 Rata-rata tinggi tanaman tomat pada MH.
Tinggi
tanaman
(cm)
dan banyak yang mati. Aplikasi bahan organik menjadikan tanah lebih gembur dan lembab. Kondisi tersebut dapat meningkatkankan populasi arthropoda tanah yang penting seperti ordo Collembola. Komposisi arthropoda tanah pada semua perlakuan dominan dihuni oleh ordo Collembola (5 famili) yang berperan sebagai pengurai dan ordo Hymenoptera (famili Formicidae) yang berperan sebagai musuh alami (Lampiran 12). Famili Formicidae (kelompok semut) sering digunakan sebagai bioindikator kesehatan ekosistem pertanian (Samways 1995).
Pada MK jumlah ordo dan famili arthropoda tanah lebih beragam pada pertanaman tumpangsari dibandingkan dengan pertanaman monokultur. Hal ini menunjukkan bahwa arthropoda tanah lebih menyenangi kondisi lahan dengan beragam tanaman dan mempunyai kelembaban cukup.
Secara umum total populasi arthropoda tanah pada perlakuan konvensional lebih tinggi dibandingkan dengan perlakuan input rendah dan organik baik pada MK maupun MH (Tabel 9). Keadaan populasi arthropoda tanah kemungkinan
Tabel 9 Jumlah ordo, famili dan populasi arthropoda tanah hasil pitfall trap pada berbagai perlakuan pada tanaman kubis
Perlakuan Penggolongan OM OT LM LT KM KT Pertanam MK Ordo 12 12 12 13 10 10 Famili 34 34 32 35 31 31 Populasi * 16.643 20.796 15.757 31.887 21.447 32.608 Pertanam MK Ordo 9 12 10 12 10 13 Famili 33 34 30 32 36 32 Populasi * 19.271 20.409 15.838 20.552 36.129 14.078 * = individu
dipengaruhi oleh aplikasi bahan organik berupa pupuk kandang atau kompos. Hal ini ditunjukkan dengan tingginya komposisi ordo Collembola dan famili Formicidae pada semua perlakuan. Hal lain kemungkinan yang mempengaruhi keadaan populasi arthropoda tanah adalah faktor lingkungan disekitar lahan penelitian. Posisi petak perlakuan konvensional yang berdekatan dengan budi daya tanaman lain yang memungkinkan terjadinya perpindahan serangga, dibandingkan dengan petak organik yang berdekatan dengan pohon bambu dan jalan. Samways
(1995) menyatakan bahwa koridor perpindahan dapat berfungsi sebagai penghubung yang membantu perpindahan atau pemencaran serangga dari suatu lahan ke lahan yang lainnya. Jalan merupakan koridor penghambat perpindahan dan pergerakan spesies tertentu dalam melintas lanskap (Forman dan Godron 1986).
Mikroorganisme
Mikroorganisme yang berhasil diisolasi dari tanah perakaran kubis dan tomat serta daun tomat disajikan pada Tabel 10.
Tabel 10 Mikroorganisme hasil isolasi dari tanah perakaran kubis dan tomat serta daun tomat
Kode Jenis koloni A B C D E F G H I J K L M N O P Q R S T U V
Pseudomonas kelompok fluorescens Warna putih, pinggir bergelombang Warna put ih, pinggir berambut Warna putih, bulat kecil
Warna putih, bentuk tidak teratur Warna putih, transparan, kecil
Warna putih, bentuk tidak teratur, tepi koloni berambut Warna kuning muda, bulat
Warna putih, bulat besar
Warna kuning, bentuk tidak teratur
Warna kuning, permukaan koloni berkerut-kerut Warna putih, berambut panjang
Warna putih, transparan besar Warna putih, mengkerut
Warna putih, transparan konsentris Cendawan, putih mengkerut ke atas Warna merah, permukaan atas kuning Cendawan, warna merah mengkerut ke atas Cendawan, warna cokelat mengkerut ke atas Warna merah, bulat, pinggir berwarna putih Warna merah, bulat, atas berwarna putih
Cendawan, warna hijau, seperti Aspergillus sp.
Jenis koloni mikroorganisme yang diisolasi dari tanah pada perlakuan organik dan input rendah sama yakni 15 jenis, sedangkan pada konvensional hanya 11 jenis (Tabel 11). Hal ini menunjukkan bahwa mikroorganisme tanah pada perlakuan organik dan input rendah lebih beragam dibandingkan dengan perlakuan
Tabel 11 Total koloni dan jumlah jenis mikroorganisme (MO) yang berhasil diisolasi dari tanah perakaran kubis dan tomat pada tiga media tumbuh
Media tumbuh
King’s B Triptic soy agar Martin agar Perlakuan (Jenis MO) Jumlah jenis MO Rata-rata total koloni Jumlah jenis MO Rata-rata total koloni Jumlah jenis MO Rata-rata total koloni Organik (A,B,C,D,F,G,H,I, K,L,M,N,O,Q,R) 9 3,10x 104 11 2,87x104 3 0,27x104 Input rendah (A,B,C,D,E,F,G, H,I,J,K,N,P,Q,R) 11 8,30x104 7 1,70x104 3 0,33x104 Konvensional (B,C,D,F,H,I,K, L,M,N,P) 8 4,10x104 7 2,13x104 1 0,03x104
konvensional. Mikroorganisme yang diisolasi tersebut diantaranya ada yang dapat tumbuh pada dua media (king’s B dan triptic soy agar). Jumlah koloni mikroorganisme yang tumbuh dari ketiga media sangat bervariasi. Pseudomonas kelompok fluorescens (A) merupakan bakteri yang bermanfaat sebagai agens biokontrol, ditemukan hanya pada perakaran tanah perlakuan organik dan input rendah. Hal ini menguntungkan sekali dalam upaya pengendalian ha yati. Beberapa isolat Pseudomonas kelompok fluorescens sudah banyak diketahui potensinya sebagai agens biokontrol (Lindow et al. 1996)
Jumlah jenis mikroorganisme yang diisolasi dari daun tomat pada ketiga perlakuan hampir tidak berbeda, pada perlakuan konvensional 12 jenis, perlakuan organik dan input rendah sama yakni 11 jenis (Tabel 12). Rata-rata koloni mikroorganisme yang tumbuh dari ketiga media sangat bervariasi.
Tabel 12 Total koloni dan jumlah jenis mikroorganisme (MO) yang berhasil diisolasi dari daun tomat pada tiga media tumbuh
Media tumbuh
King’s B Triptic soy agar Martin agar Perlakuan (Jenis MO) Jumlah jenis MO Rata-rata total koloni Jumlah jenis MO Rata-rata total koloni Jumlah jenis MO Rata-rata total koloni Organik (B,H,I,G,K,N, P,S,T,U,V) 3 17,5x104 3 0,005x104 5 0,185 x104 Input rendah (B,F,H,I,J,M, P,Q,S,T,U) 6 32,5 x104 1 0,005x104 5 0,135 x104 Konvensional (B,H,I,J,K,M, P,Q,S,T,U,V) 6 33,0 x104 1 0,005x104 6 0,310 x104 Produksi Kubis
Produksi kubis pada perlakuan non organik lebih tinggi dari pada perlakuan organik, baik secara monokultur maupun tumpangsari (Gambar 21). Hal ini kemungkinan disebabkan kurangnya ketersediaan pupuk baik kuantitas maupun kualitas pada perlakuan organik. Pada perlakuan non organik, aplikasi pupuk sintetik dilakukan secara bertahap sebagai penyedia hara makro tinggi yang siap
0 20 40 60 80 100 120 140 160 180 OM OT LM LT KM KT MK MH
Gambar 21 Rata-rata produksi tanaman kubis (ha) pada MK dan MH. Perlakuan P roduksi (ku /ha )
diserap sepanjang pertumbuhan tanaman. Pada perlakuan organik, kandungan hara makro pada pupuk kandang rendah (Tabel 13) dan pemupukan dilakukan hanya sekali. Dibandingkan dengan pupuk sintetik, pupuk kandang merupakan pupuk dengan reaksi yang lambat, akan tetapi tanah yang diberi pupuk kandang dalam jangka panjang akan memberikan hasil yang lebih baik (Simatupang 1992). Selama perkembangan tanaman, pupuk kandang mengalami dekomposisi dan mungkin terjadi pula percucian hara, sehingga pada saat tanaman membutuhkan hara ditanah kurang tersedia. Mahimairaja et al. (1995) menyatakan bahwa kehilangan N terbesar pada pupuk kandang adalah melalui denitrifikasi dan penguapan amonia. Produksi kubis pada perlakuan organik lebih rendah dibandingkan dengan non organik, juga disebabkan tingginya intensitas penyakit akar gada dan hama Gryllotalpa sp.
Tabel 13 Kandungan unsur hara pada pupuk kandang dan pupuk sintetik Unsur hara (%)
Jenis pupuk
N P2O5 K2O C organik
Ayam & kambing* 0,74 0,75 2,20 7,44
Kuda (kompos)* Urea ** TSP ** KCl ** NPK ** 0,49 45 0 0 15 0,36 0 46 0 15 0,20 0 0 50 15 9,33 0 0 0 0 Sumber: * Laboratorium Kimia, Balai Penelitian Tanah Bogor
** Program Nasional PHT (1999)
Produksi kubis pada perlakuan input rendah lebih tinggi dari perlakuan konvensional baik pada MK maupun MH. Hal ini menunjukkan bahwa tanaman pada perlakuan input rendah dengan pupuk setengah dosis dari perlakuan konvensional serta pengendalian kimia memperhatikan AE mampu membuat tanaman kubis berproduksi dengan baik.
Produksi kubis pada pertanaman tumpangsari lebih rendah dibandingkan dengan pertanaman monokultur baik pada MK maupun MH. Hal ini kemungkinan disebabkan adanya kompetisi penggunaan hara, sinar matahari dan air antara kubis dan tomat. Menururt Baharjah et al. (1993), selain keuntungan sistem pertanaman
tumpang sari juga mempunyai kelemahan antara lain terjadinya kompetisi penggunaan hara, air dan cahaya.
Tomat
Produksi tomat pada MK, perlakuan organik (26,3 ku/ha) hampir sama dengan perlakuan input rendah (27,5 ku/ha) tetapi lebih lebih rendah dibandingkan dengan produksi tomat perlakuan konvensional (51,4 ku/ha) (Gambar 22). Produksi tomat pada MH, perlakuan konvensional (76,5 ku/ha) lebih rendah dari perlakuan input rendah (87,7 ku/ha) tetapi jauh lebih tinggi dibandingkan dengan produksi perlakuan organik (3 ku/ha). Hal ini disebabkan tanaman tomat pada perlakuan organik terserang P. infestans dengan intensitas tinggi bahkan patogen tersebut menyebabkan banyak kematian tanaman tomat pada umur 73 hst.
0 20 40 60 80 100 OT LT KT Perlakuan MK MH
Analisis Usaha Tani
Hasil perhitungan analisis usaha tani yakni perbandingan antara pendapatan dan biaya (R/C rasio) pada MK menunjukkan bahwa perlakuan OM, OT dan LM lebih efisien dari ketiga perlakuan lainnya (Tabel 14). Pada MH analisis menunjukkan bahwa perlakuan LT saja yang menguntungkan karena produksi kubis dan tomat cukup berimbang dibandingkan dengan kelima perlakuan lainnya. Perlakuan konvensional pada MK dan MH mengalami kerugian, sedangkan perlakuan organik pada MK walaupun produksinya rendah tetapi menguntungkan karena harga jual produk 3 kali lebih tinggi dari produk non organik. Pertanian organik juga me mberikan dampak yang positif terhadap lingkungan dan kesehatan manusia.
Gambar 22 Rata-rata produksi tanaman tomat (ha) pada MK dan MH
P
roduksi (ku/ha)
Tabel 14 Analisis usaha tani tanaman kubis monokultur dan tumpangsari dengan tanaman tomat (ha) pada MK dan MH
Produksi Perlakuan Biaya produksi (Rp) Kubis (Rp) Tomat (Rp) Nilai (Rp) Untung/ rugi (Rp) R/C rasio Pertanaman MK OM 14.920.000 31.572.000 - 31.572.000 16.652.000 2,12 OT 18.004.000 10.206.000 9.208.500 19.414.500 1.410.500 1,08 LM 12.755.500 12.912.000 - 12.912.000 156.500 1,01 LT 16.233.500 6.962.400 2.750.000 16.233.500 -6.521.100 0,60 KM 12.205.000 12.205.000 - 12.205.000 -1.030.600 0,92 KT 16.649.000 2.702.400 5.140.000 16.649.000 -8.806.600 0,47 Pertanaman MH OM 15.168.000 8.040.000 - 8.040.000 -7.128.000 0,53 OT 17.856.000 1.176.000 1.032.500 2.208.500 -5.647.500 0,12 LM 14.128.000 10.524.000 - 10.524.000 -3.604.000 0,74 LT 17.726.000 6.897.000 13.155.000 20.052.000 2.326.000 1,13 KM 14.950.000 10.071.600 - 10.071.600 -4.878.400 0,67 KT 19.394.000 3.377.000 11.470.500 14.847.500 -4.456.500 0,77
Untuk menekan biaya sarana produksi (pupuk kandang dan kompos) pada pertanian organik, petani dapat membuat atau mengusahakan sendiri sarana tersebut. Selain itu petani dapat mengupayakan bahan yang dipergunakan untuk membuat pupuk organik tersebut lebih terjamin kualitasnya. Pupuk organik yang mempunyai hara standar (Sutanto 2002) dan aman dari patogen berbahaya merupakan langkah awal dalam pengendalian hama dan penyakit.
Sebagai salah satu bentuk usaha tani, sistem budi daya pertanian kubis input rendah dan organik baik secara monokultur dan tumpangsari layak untuk diusahakan dengan mengatur waktu tanam, mempelajari dan mempertimbangkan kesesuaian iklim.