11. TINJAUAN PUSTAKA 1. Koksidiosis p a d a ayam
Coccidia merupakan parasit intraseluler dengan beberapa perkecualian merupakan parasit pada epitel usus. Parasit ini mempunyai stadium penular yang memiliki ketahanan tinggi yaitu ookista, dan biasanya tidak membunuh induk semang (Long, 1978). Kegagalan memasuki tubuh induk semang vang sesuai merupakan satu-satunya pembatas bagi penyebaran Eimeria (Macpherson, 1978).
Mungkin semua coccidia mempunyai potensi untuk dapat menyebabkan kerusakan epitel organ induk semangnya. Berbagai akibat oleh infeksi coccidia tergantung pada dosis ookista bersporulasi yang tertelan d a n kecepatan bereproduksi di dalam induk semang (Long, 1990). Meskipun 3 (tiga) genus, Eimeria, Isospura d a n Cryptosporidia, ikut berperanan dalam menimbulkan penyakit pada hewan, tetapi tampaknya koksidiosis pada ayam berhubungan dengan genus Eimeria (Urquhart, Armour, Duncan, Dunn d a n Jennings, 1987).
Jumlah ookista yang dibutuhkan untuk menimbulkan efek patologik beragam untuk masing-masing jenis. Umur d a n kondisi dari induk semang juga berpengaruh, serta pada keadaan induk semang pernah terinfeksi atau belum (Long, 1978). Menurut Gregory (1990), koksidiosis umumnya t e j a d i hanya apabila hewan terkena infeksi yang berat dan daya tahan tubuhnya rendah.
Daya tahan tubuh berkaitan dengan umur, status kekebalan, nufxisi d a n stres. Masing-masing jenis yang berhasil diisolasi mempunyai karakter yang spesifik (Long, 1978).
Secara klinis koksidiosis pada ayam disebabkan oleh E. tenella, E. netmtrix, E . bruneffi, E . maxima a f a u E . acervulina, berdasarkan diagnostik yang diambil dari lokasi lesio (Joyner, 1978). Menurut Urquhart e f al. (1987), diagnosis yang paling baik adalah melalui pemeriksaan pasca mati, karena kesalahan dapat terjadi bila dilakukan dengan melalui pemeriksaan keberadaan ookista saja. Kesalahan disebabkan 1) efek patogenik muncul sebelum produksi ookista dan 2) tergantung dari jenisnya, jumlah ookista pada tinja tidak berkorelasi dengan derajat perubahan patologi pada usus. Pemeriksaan pasca mati dapat merupakan petunjuk yang baik yang kemudian dapat dikonfirmasikan dengan pemeriksaan ookista pada tinja. Misalnya E. necahix merupakan jenis yang "miskin" dalam memproduksi ookista, tetapi dibutuhkan hanya sedikit ookista untuk d a p a t menimbulkan koksidiosis, sednngkan E. acervulina memproduksi ookista dalam jumlah besar, tetapi untuk menimbulkan gejala klinis dibutuhkan pula jumlah ookista yang besar (Long, 1978).
Tanda klinis dari koksidiosis secara umum adalah diare dengan ataupun tanpa kehilangan darah tergantung jenis yang menginfeksi (Gregory, 1990). Dengan mempelajari patogenesis d a n patofisiologi infeksi oleh genus Elmeria,
diharapkan dapat terlihat mengapa ha1 ini dapat te j a d i , meskipun lokasi usus yang diinfeksi secara primer berbeda.
Pada ayam, koksidiosis dapat dibedakan atas koksidiosis sekum dan koksidiosis usus halus (Urquhart e t al., 1987).
1.1. Koksidiosis sekum
E . tenella merupakan jenis yang berperanan terhadap koksidiosis sekurn, meskipun stadium gametogoni dari E. necatrix dapat ditemukan dalam sekum d a n juga beberapa stadium dari E. brunetti.
Koksidiosis yang disebabkan oleh E tenella umumnya menyerang ayam berumur tiga sampai tujuh minggu. Skizon generasi ke satu berkembang pada mukosa kelenjar. Skizon generasi ke dua berkembang d a n meninggalkan mukosa, bermigrasi ke lamina propria d a n s u b mukosa. Tujuh puluh d u a jam seteIah ookista tertelan, skizon ini matang dan ruptur, kemudian timbul perdarahan, permukaan mukosa menebal d a n tanda klinis mulai muncul. Masa
prepaten adalah tujuh hari dan ookista bersporulasi d u a sampai tiga hari dalam kondisi normal (Urquhart et a1 ., 1987). Sedangkan menurut Levine (1985), rnasa prepaten adalah enam hari d a n waktu sporulasi 18 jam sampai 2 hari. Gejala klinis muncul apabila ookista tertelan dalam jumlah besar d a n dalam waktu relatif singkat, ditandai dengan tinja lembek d a n sering beserta darah. Ayam menjadi lesu disertai dengan rontoknya bulu. Pada keadaan subklinis timbul
penurunan laju pertambahan berat badan dengan keefisienan pakan yang buruk (Urquhart et al., 1987). Tanda-tanda pertama muncul pada saat merozoit generasi ke d u a membesar dan menimbulkan perdarahan pada sekum (Levine, 1985).
1.2. Koksidiosis usus halus
Pada usus halus terdapat beberapa jenis Eimeria. E. necatrix merupakan jenis yang paling patogen. Meskipun demikian prevalensi koksidiosis akibat E. necatrix mengalami penurunan sebagai akibat penggunaan obat anti coccidia secara intensif, sehingga jenis lain d i usus halus menunjukkan angka prevalensi yang meningkat, termasuk E. brunetti. Yang lebih umum adalah E. a c m u l i n a , E. maxima, d a n E . mitis serta kadang-kadang E. praecox yang tidak begitu patogen. Mass prepaten bervariasi, dari empat sampai tujuh hari. Tanda klinis muncul tiga hari setelah ookista terteIan. Umumnya ayam-ayam yang lebih tua terinfeksi oleh jenis-jenis yang terdapat dalam usus halus, dengan gejala klinis mirip dengan koksidiosis sekum, dengan perkecualian bahwa hanya jenis tertentu seperti E. necatrix dan E . brunetti yang menyebabkan kerusakan dan penyebab keluarnya darah dalam tinja. Pada pemeriksaan pascamati, tempat d a n lesio bervariasi tergantung jenisnya (Urquhart e f al., 1987). Gejala klinis yang timbul bervariasi mulai dari penurunan laju kenaikan berat badan pada
infeksi ringan, sampai pada hemoragi dan kematian, tergantung dari jenis Eimeria, derajat infeksi d a n status kekebalan induk semang (Long, 1990).
2. Genus Eimeria
Karakteristik yang terutama dari genus ini adaIah 1) komposisi dari ookista bersporulasi yang selalu mengandung empat sporokista, yang masing-masing mengandung dua sporozoit di dalamnya, 2) ditandai dengan induk semang yang spesifik, 3) adanya spesifisitas dari reaksi induk semang, dimana kekebalan terhadap satu jenis tidak melindungi terhadap jenis lain dan 4) adanya lokasi spesifik dalam menginfeksi induk semang.
2.1. Morfologi
Ookista dapat diidentifikasi berdasarkan bentuk d a n ukuran. Umumnva berbentuk bulat, oval atau elips. Berukuran antara 15
-
50 pm (Urquhart e t al., 1987). Dindingnya terdiri dari satu atau dua lapis, dan dapat dilapisi oleh suatu selaput (Levine, 1985). Dapat mempunyai mikropil d a n topi mikropil (Levine, 1985; Urquhart et al., 1987). Ookista yang belum bersporulasi, berisi massa inti dari protoplasma yang dilindungi oleh dinding yang resisten (Urquhart et al., 1987). Dalam kondisi yang sesuai, seperti keberadaan Oksigen (0,), kelembaban dan suhu, inti membelah menjadi empat sporoblas, dimana kadang-kadang sisa protoplasma membentuk badan sisa ookista. Masing-masing sporoblasmenghasilkan sporokista, dimana d i dalamnya terbentuk sporozoit (Levine, 1985; Urquhart et al., 1987). Dalam genus ini dijumpai empat sporokista dengan masing-masing berisi d u a sporozoit. Bentuk ini disebut juga sebagai ookista yang bersporulasi, dan merupakan bentuk yang infektif (Urquhart ef al., 1987). Struktur ookista dari E. tenella disajikan pada Gambar 1.
Sporokista dapat memiliki benda Stieda. Sporozoit umumnya berbentuk seperti pisang, berisi bulatan-bulatan kecil yang terang d a n bersifat seperti protein (Levine, 1985).
Struktur merozoit harnpir sama dengan sporozoit, yang membedakannya hanya adanya badan refraktil pada sporozoit, yang biasanya satu terdapat di anterior d a n satu d i posterior. Akan tetapi ada beberapa perkecualian dimana badan refraktil juga dapat dijumpai pada merozoit misalnya pada biakan jaringan (Scholtyseck et al., 1981 dalam Long, 1990). Merozoit mempunvai permukaan yang halus dengan pembukaan mikrospora tunggal dan dilapisi oleh satu selaput luar (selaput plasma). Di bawahnya terdapat selaput dalam yang terdiri dari unit selaput yang melengkapi garis plasmalemma, kecuali pada mikropor dan pada cincin polar anterior d a n posterior. Kadang-kadang terdapat satu atau d u a cincin pada polar anterior. Cincin polar anterior mengelilingi conoid silindris berisi enam atau tujuh, bahkan kadang-kadang 18-20 sub unit diagonal (Long, 1990). Dari cincin anterior, 22 atau lebih mikrotubul subpelikuler berjalan longitudinal sepanjang seluruh tubuh parasit. Di anterior
d u a atau lebih rhoptri muncul dengan muara melalui conoid dan mirrotlema. Terdapat pula butir-butir polisakarida, mitokondria, vesikula lemak, badan golgi, sisterna dari retikulo endoplasma, ribosom dan nukleus (Long, 1990).
Merozoit ini, terutama dari coccidia, sudah banyak diisolasi dan dipurifikasi oleh beberapa peneliti ( Witlock dan Danforth, 1982; Stobsh dan Wang, 1975; Fernando, Al-Attar, dan Bowles, 7984; Xie, Gilbert, Fuller d a n Mc Dougald, 1990). Struktur merozoit disajikan pada Gambar 2.
1. tutup mikropil; 2. mikropil; 3. granula kutub; 4. badan stieda; 5. geIembung refraktil kecil d i dalam sporozoit; 6. gelembung refraktil besar di dalam sporozoit; 7. sporokista; 8. bahan sisa ookista; 9. bahan sisa sporokista; 10. inti sporozoit; 11. sporozoit; 12. lapisan dalam dinding ookista; 13. lapisan luar dinding ookista
G a m b a r 1. Morfologi ookista E. tenella yang telah bersporulasi (Levine, 1985)
dndn M u r , anterior
G a m b a r 2. MorfoIogi ultrastruktur merozoit generasi I1 E. tenella (Smyth, 1981)
Stotish dan Wang (1975), menyatakan bahwa merozoit relatif tahan terhadap enzim hialuronidase dan dapat diperoleh dalam jumlah besar dari jaringan sekum yang terinfeksi atau selaput chorio alantoic dari telur berembrio dengan pemberian enzim.
2.2. Daur Hidup
Perkembangan hidup Eirneria terdiri dari sporogoni, skizogoni dan gametogoni (Lillehoj dan Trout, 1993; Watkins, 1997). Sporogoni te rjadi di luar tubuh induk semang dan menghasilkan stadium infektif, sementara skizogoni yang merupakan stadium aseksual dan gametogoni yang merupakan stadium seksual, terjadi di dalam sel induk semang yang spesifik.
Sporogoni dimulai dengan tertelannya ookista bersporulasi, yang merupakan stadium infektif, oleh ayarn yang peka. Sporozoit akan terekskistasi di dalam usus karena adanya rangsangan biokimia (Reid d a n Johnson, 1970), seperti adanya garam-garam empedu, tripsin dan CO, (Long, 1990).
Gerakan dari gizzard juga membantu dalam memecahkan dinding ookista. Sporozoit masuk ke dalam induk semang melalui penetrasi villi atau permukaan sel epitel dari mukosa usus (Long, 1990). Dalam waktu 12 - 48 jam, sporozoit berubah menjadi stadium tropozoit. Tropozoit mulai membesar d a n inti membelah yang diikuti dengan diferensiasi sitoplasma melalui proses reproduksi aseksual yang disebut skizogoni atau merogoni (Conway dan Mc
Kenzie, 1991; Urquhart ef al., 1987; Levine, 1985; Long, 1990) d a n stadium paraslt saat ini disebut skizon atau rneron. Skizon pecah ketika matang dalam waktu tiga hari, d a n melepaskan rnerozoit untuk kemudian berkembang dan rnenginvasi sel lain, sehingga te rjadi pengulangan proses perkembangan melalui stadium tropozoit d a n skizon.
Gametogoni atau reproduksi seksual, mengikuti daur skizon yang terakhir. Pada tahap ini berlangsung transfer sifat toleransi terhadap obat-obatan dan patogenitas (Watkins, 1997). Merozoit, hasil dari daur skizon ke dua kembali melakukan penetrasi sel epitel induk semang. Beberapa atau semuanya masuk ke dalam daur skizon ke tiga, tergantung dari jenisnya, sebelum membentuk gametosit jantan (mikrogametosit) atau gametosit betina (makrogametosit).
Gametosit jantan menjadi matang dan pecah, melepaskan sejumlah besar mikrogamet, melalui suatu proses pembelahan mirip pada skizogoni, sementara rnakrogametosit tidak membelah, tetapi masing-masing menjadi makrogamet. Masing-masing makrogamet mengandung inti, mitokondria d a n dua atau tiga flagela (Chobotar dan Scholtysek, 1982). Mikrogamet meninggalkan sel dan mengawini makrogamet ketika makrogamet masih di dalam sel induk semang. Bagaimana mikrogamet dapat mengenal makrogamet yang matang, sampai saat ini belum diketahui. Kemungkinan makrogamet memproduksi sendiri atau merangsang sel induk semang untuk memproduksi molekul-molekul reseptor di permukaan sel terinfeksi. Setelah te rjadi fertilisasi makrogarnet oleh
mikrogamet, dinding yang menebal terbentuk d i sekeliling zigot. Stadium ini merupakan ookista muda atau ookista yang belum matang (Conway dan M c Kenzie, 1991). Skema daur hidup dari genus Eimeria disajikan pada Gambar 3.
A. Ookista bersporulasi termakan unggas; B. ekskistasi;
I.
sporozoit menginvasi sel epitel usus; 2. sporozoit berkembang menjadi tropozoit selanjutnya berkembang menjadi skizon generasi 1 yang berisi merozoit; 3. skizon pecah, merozoit generasi 1 keluar; 4. merozoit generasi I menginvasi sel epitel vang baru; 5. merozoit I berkembang menjadi skizon generasi 2; 6. skizon generasi IJberkembang menjadi merozoit generasi 11; 7.merozoit generasi I1 berkem bang menjadi mikrogamet dan 8. makrogamet ; 9. mikrogamet aktif memasuki makrogamet; 10. fertilisasi; 11. terbentuk zigot intraseiuler dengan dinding permeabel berkembang menjadi ookista muda; 12. ookista muda keluar dari tubuh unggas melalui tinja, sporulasi di luar tubuh induk semang;
3. Patogenesis
Dalam mempelajari patogenesis dari koksidiosis terlibat dua faktor yang mengalami kerusakan yaitu parasit itu sendiri d a n induk semang yang memberi reaksi pertahanan tubuh.
Reaksi pertahanan induk semang yang hebat menyebabkan induk semang lebih mengalami kerusakan dibanding parasitnya sendiri, dan keduanya menyebabkan perubahan struktural yang berlanjut pada perubahan fisiologis pada induk semang.
Perubahan ultrastruktur pada sel yang bukan terinfeksi secara primer, diduga bukan disebabkan Iangsung oleh parasit, tetapi oleh beberapa produk hasil interaksi parasit d a n induk semang. Penemuan ini memberi dugaan bahwa infeksi coccidia menyebabkan penurunan absorpsi nutrien, dan Iebih lanjut malabsorpsi disebabkan oleh karena hilangnya permukaan mukosa yang bersifat absorptif (Allen dan Danforth, 1984). Berbagai efek fisik, fisiologis dan metabolik berinteraksi memproduksi efek akhir dari infeksi coccidia (Ruff dan Allen, 1990). Sebagai contoh, kematian akibat E. tenella mayoritas akibat adanya empat macam stres fisiologi yaitu 1) hipotermia, 2) berkurangnya cadangan karbohidrat, 3) asidosis metabolis dan 4) disfungsi tubuli ginjal (Witlock dan Danforth, 1982). Berbagai perubahan patoIogi selama koksidiosis tidak selalu muncul d a n pada galur penyebab koksidiosis sekum yang memproduksi lesio yang hebat sebenarnya kehilangan darah lebih cepat t e j a d i pada ayam yang
terinfeksi oleh galur patogen dan karena darah tidak terbuang bersama tinja (Ruff d a n Allen, 1990).
Koksidiosis juga menyebabkan peningkatan keasaman d i dalam alat pencernaan pada tempat yang terinfeksi. Perubahan konsentrasi ion hidrogen dalam alat pencernaan selama fase hemoragi adalah karena destruksi dari sel-sel usus halus yang rnenghasilkan sekretin. Hormon ini terbentuk pada dinding usus haIus d a n rnenunjukkan jumlah cairan pankreas yang tersekresi (Sturkie, 1965 dalam Stephens, Borst d a n Barnett, 1974). Konsentrasi ion hidrogen dalam alat pencernaan tergantung dari jumlah HCl yang disekresikan ke dalam proventrikulus, d a n cairan pankreas bereaksi untuk menetralisasi asam ini. Oleh karena itu apabila produksi sekretin berkurang, maka cairan pankreas yang disekresi juga berkurang, sehingga situasi dalam alat pencernaan menjadi lebih asam. Adanya perubahan p H d a n mikroflora usus akan menggangu absorbsi zat-zat nutrisi seperti lemak, pigmen karotinoid, xantofil, vitamin, mineral d a n asam amino (Van d e r Sluis, 1993; Mc Dougald d a n Reid, 1997). Menurut Sturkie (1965) dalam Stephens et al. (1974), penurunan konsentrasi ion hidrogen dalam alat pencernaan akibat infeksi E. necatrix adalah karena tidak adanya makanan d i dalamnya, degradasi jaringan dalam usus atau ketiadaan efek bufer pada makanan.
Keadaan yang cukup menarik adalah, apabila d u a jenis coccidia menginfeksi bagian usus yang sama, misalnya E. brunetti d a n E . rnitis, mereka
akan berkompetisi, dan apabila mereka bekeqa sama, efek yang ditimbulkan tidak lebih dari efek yang terjadi apabila mereka berdiri sendiri-sendiri. Sedangkan apabila d u a jenis coccidia menginfeksi di lokasi yang berbeda, kerjasama mereka akan dapat memberikan efek melebihi efek dua dosis dari infeksi tiap jenis.
Telah terungkap bahwa masing-masing jenis mempunyai karakteristik sendiri-sendiri dalam menimbulkan koksidiosis beserta lokasi yang diminatinya, yang berpengaruh pada pejalanan penyakit. Usus halus merupakan lokasi yang populer bagi coccidia. Tampaknya coccidia yang menginfeksi usus besar lebih patogen dari pada yang menginfeksi usus halus. Keadaan ini dikarenakan laju pergantian sel lebih rendah, d a n karena apabiln usus besar mengalami kerusakan tidak ada lagi bagian usus yang bersifat absorptif yang terletak lebih posterior lagi, dan juga karena d i dalam usus besar cenderung Iebih banyak organisme yang bersifat oportunis yang siap untuk merusak mukosa. Pada beberapa jenis (E. a c m u l i n a , E. necatrix dan E . brunetti), tempat tejadinya parasitisme tergantung pada bagian usus mana yang terinvasi oleh sporozoit. Sedangkan untuk E. tenella dan E. praecox untuk perkembangannya mempunyai lokasi yang telah tertentu, yaitu sekum untuk E. tenella d a n duodenum untuk E.
praecox (Sharma d a n Reid, 1962). Long (1978), juga menyatakan bahwa kecepatan proses ekskistasi akan menentukan bagian mana dari usus yang akan diinvasi oleh sporozoit. Meskipun demikian, kadang-kadang siklus hidup
menjadi lengkap bukan pada bagian yang seharusnya, seperti E. bvunefti yang kadang-kadang mengalami perkembangan siklus hidup d i sekum. Hal ini dapat terjadi karena misalnya material dari kloaka kembali ke dalarn sekum akibat adanya proses anti peristaitik (Akester, Anderson, Hill d a n Osbaldiston, 1967), sehingga keadaan ini menyebabkan transfer merozoit yang berlanjut ke perkembangan stadium selanjutnya.
Faktor yang juga berpengaruh terhadap pe rjalanan penyakit adaIah jenis sel yang terinfeksi beserta kelainan-kelainan yang te j a d i . Kebanyakan coccidia terutama menginfeksi sel-sel jauh ke dalam kripta. Hanya coccidia yang memproduksi meron dalam jumlah sedikit yang d a p a t menginfeksi sel-sel yang telah ada pada villi, misalnya E. acervulina. Jenis semacam ini menyebabkan lesio yang dengan cepat sembuh kembali, karena sel stem pada kripta tidak mengalami kerusakan. Rusaknya kripta akibat coccidia tertentu dapat berlanjut dengan rusaknya sel stem. Apabila seluruh sel stem dalam suatu daerah rusak d a n mati, epitel tidak dapat digantikan kecuali melalui "pelanggaran batas" dari tempat yang rusak (Gambar 4). Kadang-kadang beberapa stadium dari coccidia menghindar dari masalah ini dengan menyebabkan berpindahnya seI induk semang dari epitelium ke lamina propria (Fernando, Lawn, Rose d a n Al-Attar,
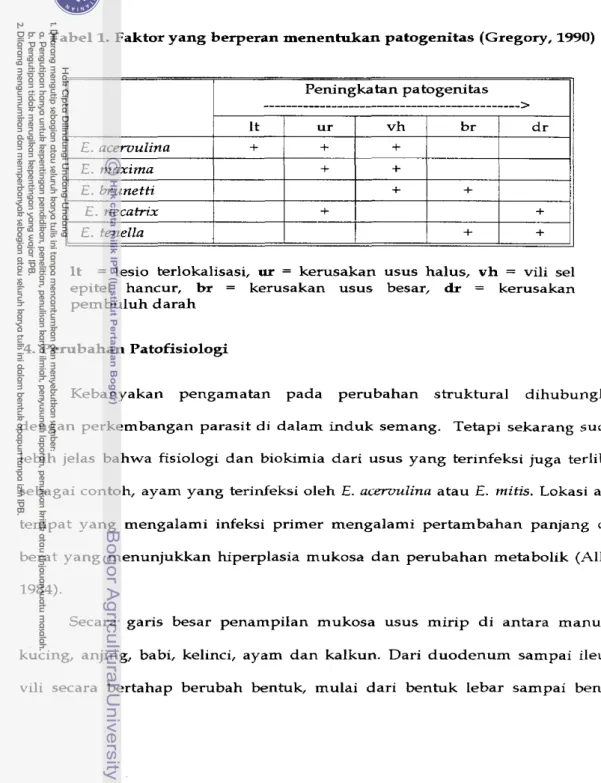
1983). Tabel I menyajikan faktor yang berperan pada patogenitas untuk beberapa jenis Eimeria.
INFEKSI KOKSIDIA sel hilang hiperplasia kemam uan memperbaiki rerusakan berkurang imaturitas permukaan sel epitel keluruhan
ukaan &pice1 menurun
\
a, ,- infasi oleh
/
oportunis\
I
ma la bsorpsi\
//
nekrosa r ~ > ~ j a k n v a4
diare dehidrasi \ absorpsi toksrn . - - -. . . pembuGi darah-
dara h/
kehi'anganGambar 4. Patogenesis koksidiosis, jalur kemungkinan penyebab te jadinya perubahan. yang kurang patogen rnengikuti jalur sebelah kiri, yang lebih patogen biasanya mengikuti keempatjalur (Gregory, 1990)
23
Tabel 1. Faktor yang berperan menentukan patogenitas (Gregory, 1990)
It = lesio terlokalisasi, ur = kerusakan usus halus, v h = vili sel epitel hancur, br = kerusakan usus besar, dr = kerusakan pembuluh darah
4. Perubahan Patofisiologi
Kebanyakan pengamatan pada perubahan struktural dihubungkan dengan perkembangan parasit di dalam induk semang. Tetapi sekarang eudah lebih jelas bahwa fisiologi dan biokimia dari usus yang terinfeksi juga terfibat, sebagai contoh, ayam yang terinfeksi oleh E. acervulina atau E. mifis. Lokasi atau tempat yang mengalami infeksi primer mengalami pertambahan panjang dan berat yang menunjukkan hiperplasia mukosa d a n perubahan metabolik (Allen, 1984).
Secara garis besar penampiIan mukosa usus mirip di antara manusia, kucing, anjing, babi, kelinci, ayam dan kalkun. Dari duodenum sampai ileum, vili secara bertahap berubah bentuk, mulai dari bentuk lebar sampai bentuk
d a u n d i duodenum; dari bentuk jari sampai bentuk lidah pada ileum. Vili-vili itu juga memendek dan sempit. Umumnya ha1 ini diterima sebagai gradasi dari proses yang berhubungan dengan laju pergantian sel, d a n berpengaruh pada derajat infeksi terhadap kerusakan fisik d a n kimia isi lumen usus.
Peningkatan pergantian sel merupakan ha1 yang biasa pada infeksi coccidia. Diferensiasi sel juga meningkat s e h n g g a terjadi hiperplasia sel goblet. Infeksi tidak hanya menyebabkan kehiIangan darah, tetapi populasi sel-sel absorptif pada mukosa juga berkurang.
Coccidia yang menginfeksi dan berkembang pada sel stem kripta, umumnya pada usus kecil bagian bawah, sekum d a n kolon bersifat sangat patogen karena cenderung menghancurkan sel stern kripta yang diinvasi. Kerusakan ini mencegah pembaharuan epitel vili sehingga te q a d i peluruhan dari vili yang berakibat hilangnya cairan, hemoragi, kerentanan terhadap invasi bakteri (terutama coliform), dan kemudian te rjadi lesio nekrotik. Infeksi berat kadang-kadang disertai diare yang diakhiri dengan kematian.
Efek dari coccidia yang secara primer mendiami epitel vili lebih bervariasi. Infeksi E. rnitis misalnya, yang tidak memperlihatkan lesio, bahkan pada infeksi berat sekalipun, tetapi menyebabkan depigmentasi d a n gangguan pertambahan berat badan (Ruff d a n Edgar, 1982). Sedangkan pada infeksi E. necatrix secara hebat merubah sel kripta induk semang yang diinvasi. Perubahan-perubahan yang te rjadi antara Iain perluasan sel, sehingga sel induk
semang menjadi lebih peka untuk melisis terhadap tripsin walaupun dalam konsentrasi rendah. Mayoritas patologi dari infeksi E. necatrix berhubungan dengan kerusakan sel-sel d i atas.
Dalam mempelajari infeksi dengan E. a c m u l i n a juga terlihat perubahan metabolistik pada daerah usus yang tidak terinfeksi secara primer (Allen, 1984). Malabsorpsi nutrisi pada tempat infeksi primer tampak berhubungan dengan derajat kerusakan mukosa yang dipengaruhi oleh dosis infeksi (Allen, 1984). Pada infeksi ini juga ditemukan kehilangan pigmen plasma (Ruff dan Fuller, 1975) d a n pergantian epitel pada seluruh vili meningkat (Long, 1978). Meskipun mukosa usus halus bagian distal mengalami hiperplasia selama infeksi E. a c m u l i n a , tetapi kemampuan menyerap karotenoid berkurang (Allen, 1987). Pada unggas yang terinfeksi E. acervulina juga terlihat laju oksidasi glukosa dan asam oktanoat. Effek ini kemungkinan disebabkan oleh penurunan fungsi pada daerah lapisan mukosa duodenum yang terinfeksi atau menurunnya fungsi mitokondria, atau keduanya. Oleh karena itu perlu dipejelas apakah mitokondria dari epitel yang terinfeksi mempunyai fungsi yang sama dengan jaringan yang tidak terinfeksi. Pada kenyataannya mitokondria dari duodenum yang terinfeksi mengoksidasi asam oktanoat d a n alfa keto glutarat lebih rendah dibanding kontrol. Pada jaringan normal yang memetabolis glukosa melalui jalur glikolitik dan asam trikarboksilat, karbon C, dan C, dari glukosa dapat dibebaskan sebagai CO, dengan laju yang sama. Infeksi oleh jenis ini
menyebabkan rasio C1 : C6 meningkat menjadi 4 kali lipat. h i merupakan indikasi bahwa metabolisme dari oksidasi mitokondria beralih ke oksidasi melalui jalur pentosa fosfat. Jenis Eimeria yang lain seperti E. miuati dan E.
brunetti juga menyebabkan malabsorpsi nutrisi pada induk semang (Ruff dan Edgar, 1982).
E.mitis selama bertahun-tahun dikabkan relatif tidak patogen, tetapi Joyner (1958) dan Fitz-Coy dan Edgar (1981), melaporkan adanya sedikit gangguan pertumbuhan, kadang-kadang juga dipengaruhi oleh jenis ini. E. mitis
menyebabkan malabsorpsi glukosa dan methionin sampai 52 98, sehingga tidak mengherankan apabila terjadi penurunan berat badan d a n efek patologik laimya. Malabsorpsi ini terjadi tanpa adanya lesio yang cukup hebat. Biasanya malabsorpsi selalu berhubungan dengan adanya parasit, lesio dan kerusakan pada usus. Tidak adanya lesio pada infeksi E. mitis situasinya hampir sama dengan koksidiosis pada kalkun dimana malabsorpsi te rjadi d a n parasitpun ada tanpa adanya kerusakan usus (Ruff, Augustine dan Madden, 1981), sehingga masalah muncul apabila te qadi malabsorpsi nutrisi dan penurunan berat badan tanpa adanya lesio yang biasa digunakan untuk mendiagnosis koksidiosis (Ruff dan Edgar, 1982). Perubahan ultrastruktur pada sel yang bukan terinfeksi secara primer, diduga bukan disebabkan langsung oleh parasit, tetapi oleh beberapa produk hasil interaksi parasit dan induk semang. Penemuan ini memberi dugaan bahwa infeksi coccidia menyebabkan penurunan output energi pada
jaringan terinfeksi yang menyebabkan penurunan absorpsi zat-zat nutrisi yang aktif ditranspor dan lebih lanjut malabsorpsi disebabkan pula oleh karena hilangnya permukaan mukosa yang bersifat absorptif (Allen d a n Danforth, 1984). Berbagai efek fisik, fisiologi d a n metabolik berinteraksi menghasilkan efek akhir dari infeksi coccidia (Ruff dan Allen, 1990).
Dengan demikian di antarst jenis-jenis Eimeria yang menginfeksi ayam menimbulkan tanda-tanda penyakit yang hampir sama meskipun melalui jalan yang agak berbeda karena terdapat faktor-faktor tertentu yang ikut berperan. Malabsorbsi sebenarnya berhubungan dengan situasi dimana terdapat abnormalisasi dari struktur mukosa usus yang menyebabkan penurunan kapasitas absorptif dari usus (Robinson d a n Huxtable, 1988).
Menurut Ruff d a n Edgar (1982), untuk j e ~ s - j e n i s E. acervulina, E. miuati E . brunetti d a n E . necatrix, malabsorbsi zat nutrisi berhubungan dengan adanya parasit d a n keadaan lesio usus secara menyeluruh. Secara umum koksidiosis pada ayam menimbulkan malabsorbsi zat nutrisi pada daerah usus halus yang diinfeksi oleh parasit, akan tetapi infeksi ganda yang melibatkan "seluruh" usus dapat menyebabkan malabsorbsi d i seluruh bagian dan kemudian menghasilkan penurunan absorpsi total yang nyata secara fisiologis (Ruff d a n Wilkins, 1980).
Sedangkan Michael (1973) dalam Allen dan Danforth (1984) menyatakan bahwa malabsorbsi berhubungan dengan penurunan aktifitas metabolik dari sel epitel duodenum.
Berbagai zat nutrisi mengalami transportasi menyeberangi epitel usus halus melalui proses-proses yang membutuhkan energi metabolis (Lehninger, 1975, dalam Allen dan Danforth, 1984). Apabila terjadi gangguan efisiensi katabolisme karbohdrat pada jaringan sedemikian rupa sebagai pengurangan kemampuan energi metabolis (terutama dalam bentuk ATP), maka absorbsi zat nutrisi bisa berkurang.
Pada pemeriksaan pasca mati, terdapat lesio-lesio terutama akibat radang, hiperplasia epitel dan luruhnya epitel (Long, 1990). Pada infeksi E. tenella,
hemoragi atau lesio muncul pada sekum pada hari ke 5 (Gregory, 1990). Semua pemeriksaan pasca mati pada ayam-ayam yang pada tinjanya terdapat darah, sekumnya dijumpai berdilatasi d a n berisi campuran darah beku dan darah cair (Urquhart e t al., 1987).
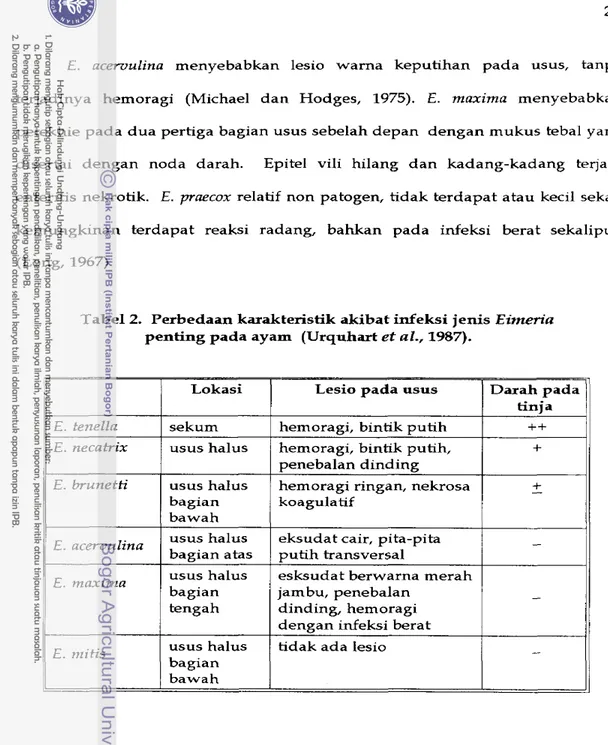
Pada koksidiosis usus halus, lokasi dan macam lesio bervariasi tergantung jenisnya (Urquhart e t al., 1987), seperti yang disajikan pada Tabel 2.
Pada infeksi E. necatrix, arsitektur kripta seringkali tetap terpelihara dan lapisan dari set yang tidak terinfeksi acapkali melapisi lubang-lubang pada sel-sel terinfeksi, menyebabkan te rjadinya regenerasi pada kripta (Stockdale dan Fernando, 1975).
E . acervulina menyebabkan lesio warna keputihan pada usus, tanpa tejadinya hemoragi (Michael dan Hodges, 1975). E. maxima menyebabkan petekhie pada dua pertiga bagian usus sebelah depan dengan mukus tebal vang disertai dengan noda darah. Epitel vili hilang dan kadang-kadang terjadi enteritis nekrotik. E. praecox relatif non patogen, tidak terdapat atau kecil sekali kemungkinan terdapat reaksi radang, bahkan pada infeksi berat sekalipun (Long, 1967).
Tabel 2. Perbedaan karakteristik akibat infeksi jellis Eimeria penting pada ayam (Urquhart et al., 1987).
11
E . tenellaI
sekum1
hernoragi, bintik putihI
++11
Lokasi Lesio pada ususE . necatrix E. brunetti
E . acervulina
usus halus tidak ada lesio E . mitis bagian Darah pada
/
tinia I E . maximaI
bawah usus halus usus haIus bagian bawah usus halus bagian atas usus halus bagian tengahhemoragi, bintik putih, penebalan dinding hemoragi ringan, nekrosa koagulatif
eksudat cair, pita-pita putih transversal
+
-
+
- esksudat berwarna merah
jambu, penebalan dinding, hemoragi dengan infeksi berat
-
6. Aspek Kekebalan
Respon kekebalan yang mempakan interaksi antara induk semang dengan E i m e r i a s p p sudah banyak dibicarakan dan yang menarik dalam ha1 ini adalah adanya perbedaan diantara jenis E i m w i a . Semuanya memang menginduksi kekebalan dengan derajat yang sangat tinggi, tetapi usaha untuk mencapainya berbeda-beda.
Kekebalan terhadap koksidiosis pada unggas sudah banyak dipelajari oleh beberapa peneliti. Kekebalan ini sangat jenis spesifik, bahkan beberapa memiliki galur spesifik (Rose, 1978; Lillehoj, 1987), tergantung dari jenis Ei-meria
yang menginfeksi. Hal ini karena tidak adanya kekebalan silang antar jenis
Eimeria. Akibatnya ayarn yang terifeksi oleh satu jenis E i m w i a akan tetap rentan terhadap infeksi dari jenis Eimeria yang lain ( Danforth, 1997; Mc Dougald dan Reid, 1997; Shirley, 1993 ; Watkins, 1997). Namun demikian mekanisme dari kekebalan masih belum jelas, terutama peranan antibodi serum terhadap kekebalan yang protektif (Lillehoj, 1987; Jenkins dan Dame, 1987). Antibodi yang bersirkulasi, umumnya terdeteksi dalam satu minggu pasca infeksi, mencapai puncak satu sampai dua bulan kemudian, dan kemudian menurun, tetapi masih tetap ada di dalam sirkulasi (Lillehoj, 1991). Lamanya dan tingkatan reaksi antibodi tergantung dari umur induk semang, dosis inokulasi dan jenis dari Eimeria. Antibodi ini dapat menjadi efektif apabila te rjadi kontak
dengan parasit (Barriga, 1981; Rose, Long d a n Bradley, 1975). Oleh karena itu seharusnya ada m e k a ~ s m e yang dapat menyebabkan terjadinya kontak tersebut. Mekanisme semacam itu dapat diidentifikasi pada peningkatan permeabilitas vaskuler, yang dihasilkan dari tantangan parasit pada usus yang kebal (Rose ef a1
.,
1975). Kenyataannya telah ditemukan antibodi dari kelas IgG dan IgM dalam serum, serta IgG dan IgA dijumpai dalam sekresi usus ayam terinfeksi (Barriga, 1981).IgG merupakan imunoglobulin terbanyak (Goding, 1987; Roitt, 1994), khususnya di dalam cairan ekstravaskuler. IgG mengikat komplemen, melekat pada sel fagosit dan menembus plasenta (Roitt, 1994), serta memiliki dua rantai panjang dengan berat molekul mencapai 160000 (Goding, 1987).
IgG terdiri atas empat sub kelas yaitu IgG1, IgG2, IgG3 dan IgG4 pada manusia (Roitt, 1994; Goding 1987 ; Clark, 1991), sedangkan pada mencit terdiri atas IgG1, IgG2a, IgG2b dan IgG3, serta pada tikus IgGl, IgGZa, IgG2b dan IgG2c pada tikus (Goding, 1987). Nomenklatur dari sub kelas IgG pada suatu jenis tidak berhubungan dengan jenis lain (Goding, 1987). Perbedaan terdapat pada rantai panjang yang disebut sebagai8yl,y2,y3,y4 (Roitt, 1994; Clark, 1991; Goding, 1987). Rantai-rantai panjang ini mirip satu sama lain, tetapi tidak identik dalam ha1 komposisi asam amino dan ikatan disulfida (Roitt, 1994), sehingga menyebabkan adanya perbedaan dalam sifat-sifat biologik. Semua subkelas IgG relatif stabil dalam larutan, . meskipun ikatan disulfida dapat
dipenggal oleh enzim proteolitik dalam kondisi yang tepat (Goding, 1987). Terdapat korelasi antara ikatan disulfida dengan kemampuan mengikat komplemen. IgG3 merupakan pengikat komplemen yang paling efektif, diikuti oleh IgGl dan IgG2. Umumnya IgG4 gaga1 mengikat komplemen (Clark, 1991). Menurut Goding (1987), IgGZb mempunyai ikatan yang sangat fleksibel dan i m ~ n o ~ l o b u l i n ini sangat efisien dalam mengikat komplemen. Menurut Lillehoj (1993; 1996), infeksi oleh Eimeria spp. menggertak timbulnya antibodi , baik yang bersirkulasi (IgM, IgG dan IgA) maupun yang disekresikan oleh mukosa usus (IgA).
Pemberian ookista peroral dari berbagai j e ~ s Eimeria pada ayam diikuti dengan tanda yang khas dalam beberapa jam, berupa reaksi hipersensitivitas lokal. Permeabilitas vaskuler terhadap makromolekul meningkat. Perubahan histologik juga muncul, terutama pengalihan dari jaringan epitel ke dalam lumen. Reaksi ini meskipun terlihat pada ayam rentan, umumnya terdapat pada ayam yang kebal terhadap infeksi dan dapat disimpulkan bahwa pergantian sel epitel merupakan mekanisme yang penting bagi terekspresinya kekebalan terhadap koksidiosis (Long dan Speer, 1977).
Berkembang dua teori yang menerangkan reaksi ini dan kemungkinan jalannya kekebalan. Rose dan Long (1969 dalam Long dan Speer ,1977, menyatakan bahwa masuknya sporozoit kedalam sel epitel meningkatkan permeabilitas usus dan menyebabkan terlepasnya makromolekul. Pada saat ini
antibodi kemungkinan keluar sehingga memungkinkan te jadinya kontak antara parasit dan antibodi yang bersifat parasitisid.
Di sisi lain Kouwenhoven dan Horst (1973 dalam Long d a n Speer, 2977) menganggap reaksi yang terjadi adalah dengan mengaiirnya cairan ookista ke dalam lumen selama ekskistasi sporozoit. Enzim yang terutama terdapat pada cairan ookista dikatakan mengkonversikan glukosa pada sisa makanan menjadi asam laktat dan asam lain sehingga m e n u ~ n k a n p H usus, dan ini menyebabkan kerusakan epitelium dengan akibat adanya kebocoran protein serum.
Pada individu yang kebal, secara histologi terlihat bahwa sporozoit dari E . fenella menginfeksi epitel sekum akan tetapi tidak te qadi perkembangan.
Studi histopatologi menunjukkan reaksi seluler relatif ringan, akan tetapi kemudian sel polimofornuklear muncul diantara sel epitel. Selama skizogoni te rjadi peningkatan jumlah sel polimorfonuklear, limfosit dan sel pironinofilik pada submukosa dan lamina propria (Long dan Speer, 1977).
Pada dasarnya dalam menginduksi reaksi kekebalan tergantung kepada jumlah inokulum dan rute pemberian tanpa mengabaikan respon induk semang
(Rose, 1978). Apabila kekebalan te rjadi dengan lengkap, maka tidak akan ada efek patogenik yang muncul. Reaksi selulerpun muncul dalam waktu singkat dan menunjukkan bahwa reaksi yang te j a d i dapat bersirkulasi. Pembuktian mobilisasi sel dan sirkulasi diperoleh dari fluktuasi leukosit yang bersirkulasi, jumlah heterofil dan limfosit yang meningkat. Lebih lanjut lagi dengan antigen,
dimana kadang-kadang timbul reaksi hipersensitifitas yang menghasilkan peningkatan permeabilitas. Mekanismenya tidak diketahui dengan pasti, tetapi situasinya menunjukkan bahwa disini melibatkan kompleks antigen dan antibodi. Bagaimana antigen muncul, cara sehingga mereka sesuai dengan induk semang, dan bagaimana reaksi induk semang terhadap parasit tergantung pada stadium di dalam siklus hidup dan keterkaitannya dengan sel dan jaringan induk semang. Perbedaan antigen pada stadium dalam siklus hidup merupakan penentu perbedaan imunogenitas (Rose, 1978).
Hanya sedikit penelitian yang telah dilakukan untuk mempelajari lamanya reaksi kekebalan terhadap Eimeria spp., karena kesulitan dalam memelihara ayam-ayam bebas dari infeksi lain untuk waktu yang lama (Long dan Rose, 1982). Data yang ada menunjukkan bahwa kekebalan menurun sejalan dengan waktu dan tergantung pada umur saat induk semang diimunisasi, serta cara pemberian (Long, 1967). Akan tetapi meskipun timbul reaksi kekebalan, ayam masih tetap dapat terinfeksi karena lingkungan yang terkontaminasi yang kerapkali te rjadi di alam. Menurut Long dan Millard (1969 dalarn Long dan Rose ,1982), ayam-ayam yang dipelihara daIam kandang litter dan terinfeksi pada umur satu atau dua minggu, ternyata sangat tahan terhadap tantangan setelah diuji pada umur 18 minggu. Hal ini diduga karena ayam-ayam tersebut
Stadium mana dari coccidia yang mengandung imunogen yang merangsang respon kekebalan protektif juga masih diperdebatkan. Namun diduga stadium perkembangan aseksual yang merupakan sumber dari imunogen terkait
.
7. Diagnosis
Diagnosis yang paling baik adalah melalui pemeriksaan pasca mati dari beberapa unggas yang terinfeksi. Walaupun ookista dapat dideteksi melalui pemeriksaan tinja, namun ha1 ini dapat menimbulkan kekeliruan karena 1) efek patogenik umumnya muncul sebelum produksi ookista dan 2) jumlah ookista tidak selalu berkorelasi linier dengan keparahan perubahan patologi di usus.
Pada nekropsi, lokasi d a n tipe dari perlukaan yang timbul merupakan petunjuk yang baik terhadap jenis coccidia yang menginfeksi , namun perlu dikonfirmasikan melalui pemeriksaan ookista pada tinja dan skizon serta ookista pada kerokan usus (Urquhart et al., 1987 ; Conway dan Mc Kenzi, 1991).
Kesulitan dalam mendiagnosis koksidiosis pada unggas secara akurat dikarenakan adanya tumpang tindih antara daerah usus yang diinfeksi oleh jenis yang berbeda, kemudian perubahan perlukaan selama siklus hidup berlangsung, dan perbedaan status dari ayam yang berbeda terhadap tahapan
pe jalanan koksidiosis d i dalam satu flok , karena sebagian menampakkan lesio akut, sebagian dalam tahap persembuhan dan yang lainnya tidak terinfeksi (Conway d a n Mc Kenzie, 1991).
Diagnosis terhadap koksidiosis yang akut tidaklah sulit uoyner, 1978), tetapi wabah penyakit ini tidak selalu terlihat meskipun pemberian obat anti coccidia dalam makanan telah dihentikan. Pada ayam pedaging penyebab enteritis yang lain dapat mengacaukan diagnosis seperti infeksi Clostridia atau diare yang berhubungan dengan penyakit Gumboro (Macpherson, 1978).
8. Pengendalian
8.1. Kemoterapi
Metode intensif yang mungkin dilakukan untuk menanggulangi koksidiosis adalah perbaikan sistem perkandangan, penggunaan kemoterapi d a n secara imunologis (Long, 1978). Selain penggunaan obat anti coccidia, juga dengan manajemen lingkungan dan kontrol kualitas makanan (Conway dan Mc Kenzie, 1991). Akan tetapi sterilisasi kandang secara menyeluruh dan lingkungan kandang yang bebas dari ookista, dapat menghambat terbentuknya respon kekebalan ayam secara dini (Mc Dougald dan Reid, 1997).
Pencegahan koksidiosis pada ayam didasarkan pada kombinasi dari manajemen yang baik dan penggunaan anti coccidia dalam air minum dan
pakan (Urquhart et al., 1987). Pengobatan kadang-kadang kurang perlu bagi ayam-ayam yang dipelihara di lantai (Long
,
1978), karena dengan sistem pemeliharaan ini reinfeksi dapat terjadi berulang kali. Penggunaan disinfektan tampaknya juga kurang efektif untuk memusnahkan ookista, karena ookista sebagai sumber infeksi, terbukti tahan terhadap berbagai macam disinfektan (Janssen Pharmaceutica, 1992; Mc Dougald dan Reid, 1997; Weber, 19971, seperti formalin, campuran quatenary ammonium, copper sulfat, potassium dihidroksida dan potassium permanganat (Reid, 1989 dalam Van der Sluis, 1993) belum ada disinfektan yang efektif untuk membunuh coccidia. Fumigasi dengan menggunakan methyl bromide atau ammonia diharapkan lebih efektif, tetapi masih diragukan bahwa penggunaan disinfektan saja tanpa pengobatan dapat melindungi koksidiosis subakut (Long , 1978).Selama 25 tahun terakhir sekitar 20 jenis obat-obatan yang bebeda digunakan untuk mengendalikan koksidiosis pada ayam. Pada umumnya koksidiostat bekerja menghambat perkembangan dari stadium cocc~dia dalam jaringan, hanya sedikit yang dapat membunuh. Aktifitas kimiawi dari obat terhadap parasit perlu diketahui. Ionophor, quinolon dan clopidol bekerja melawan sporozoit dan tropozoit (Mc Dougald , 1990), sementara clopidol mempengaruhi sporulasi (Arakawa, Nakai, Baba d a n Fukata, 1991) demikian juga amprolium dan dinitolmid (Joyner dan Norton, 1977 ; Mathis dan
Mc Dougald, 1981) maupun aprinocid (Ruff, Anderson d a n Reid , 1978; Tamas, Olson, Smyth dan Miller, 1978).
M e k a ~ s m e ke rja yang pasti dari berbagai obat-obatan tidak terlalu dipahami. Salah satu yang banyak diketahui adalah quinolon. Obat ini mengikat sistem transpor elektron dalam mitokondria pada sporozoit, s e h n g g a menghambat reaksi pemenuhan energi (Wang , 1976).
Amprolium secara kimiawi mirip dengan thiamin, dan diduga menghambat penggunaan thiamin oleh parasit (Rogers , 1962). Sporozoit tidak dapat bertahan hidup d i dalam sel setelah pengobatan dengan ionophor, yang diduga karena tidak adanya mekanisme yang efektif untuk mempertahankan keseimbangan osmosa (Smith d a n Strout, 1979 ; Long, 1978). Pada dekade 70-an, ionophor monensin , merupakan anti coccidia pilihan dari hampir 80% peternakan ayam broiler d i Amerika Serikat dan Inggris (Long
,
1978; Watkins, 1997). Beberapa keunggulan dari anti coccidia ionophor antara lain aktifitasnya yang bersprektum luas, merniliki sifat sebagai pemacu pertumbuhan d a n memiliki aktifitas anti jamur, anti viral serta dapat berperan sebagai antibiotik (Watkins, 1997). Pada dasarnya coccidia dapat menjadi resisten terhadap semua obat, apabila di berikan secara terus menerus. Dugaan ini telah dibuktikan oleh Gilbert, Mc Dougald dan Mc Murray (1992),
yang menemukan bahwa coccidia yang diisolasi daripeternakan broiler komersial sudah menjadi resisten terhadap semua obat komersial.
Menurut Mc. Dougald (1992), dibutuhkan suatu strategi untuk menghindari timbulnya resistensi terhadap obat anti coccidia. Dianjurkan d u a macam program yang disebut sebagai shuttle d a n rotation. Program rotation merupakan program yang sederhana dengan menggunakan dua atau lebih obat-obatan secara bergilir pada berbagai intervaI waktu. Sedangkan program shuttle menggunakan satu produk untuk periode starter d a n produk lain untuk fase graving (Conway, 1996; Mc Douga1;d d a n Reid, 1997; Weber, 1997).
8.2. Imunisasi
Sejak diketahui timbulnya kekebalan setelah infeksi koksidiosis (Smith, Long d a n Pierce, 1963), maka para ahli mulai tertarik untuk menanggulangi koksidiosis secara imunologis. Berbagai cara diusahakan seperti pengujian antigenesitas dari parasit mati, d a n juga penggunaan parasit hidup sebagai bahan imunisasi, yang diolah sedemikian rupa supaya tidak merusak induk semang.
8.2.1. Imunisasi dengan Parasit Mati
Imunisasi dengan cara menyuntikkan antigen parasit secara parenteral sudah banyak dilakukan, tetapi masih banyak yang belum dipublikasikan. Antibodi dalam serum memang terbentuk akibat infeksi, tetapi tidak efektif di
dalam usus (Long dan Rose, 1982).
Kemampuan menurnbuhkan beberapa jenis Eimeria dalam biakan sel (Patton, 1965; Doran, 1970; Strout dan Ouellete, 1970) atau pada embrio ayam (Long, 1965; 1966) telah dapat dilakukan. Akan tetapi penyuntikan material ~ n i secara parenteral temyata tidak mampu melindungi terhadap infeksi, meskipun antibodi d a n reaksi perantaraan sel terinduksi (Long dan Rose, 1982).
8.2.2. Imunisasi dengan Parasit Hidup
Penggunaan parasit hidup sebagai bahan imunisasi juga telah banyak diteliti. Dengan mengatenuasi parasit ini diharapkan dapat merangsang kekebalan yang protektif, tapi tidak terlalu patogen terhadap induk semang.
Atenuasi yang dilakukan dengan menggunakan agen fisika seperti pendinginan, pemanasan atau ultrasonik (Conder dan Duszyuki, 1977) dari ookista menunjukkan bahwa penurunan efek patogenitas yang disebabkan oleh inokulasi ookista yang mendapat perlakuan sebanding dengan pengurangan jumlah ookista hidup dalam inokulum.
Menurut Rose (1973), pemberian dengan ookista yang diradiasi, masih meragukan karena kondisi irradiasi belum distandardisasi dan perbandingan yang cermat antar hasil dari dosis tinggi dari ookista dengan perlakuan dari dosis yang lebih rendah dari ookista tanpa perlakuan belum dilakukan. Menurut Jang e f al. (1992), imunogenisitas dari E. tenella yang diradiasi dengan Cobalt 60, akan bertambah sejalan dengan bertambahnya jumlah pasase pada ayam.
Di Indonesia, mulai dikembangkan radio vaksin untuk coccidia seperti vaksin koksivet dan koksivet supra 95 polivalen akan tetapi hasil penggunaan di lapangan belum banyak dilaporkan. Atenuasi dengan adaptasi atau seleksi dimulai setelah berhasilnya menumbuhkan E. tenella pada embrio ayam (Long, 19651, yang menimbulkan gagasan untuk melemahkan patogenitas daiam embrio ayam. Penurunan patogenitas ini berhasil dilakukan setelah 62 pasase pada embrio (Long, 1974), dan ha1 ini disebabkan oleh karena ukuran dari skizon generasi kedua yang mengecil dan letaknya di mukosa lebih superfisial.
Galur E. fenella yang menurun virulensinya juga berhasil ditemukan dengan seleksi yang terus menerus terhadap perkembangan awal yang disebut sebagai galur precocious yang disebabkan oleh terganggunya pembentukan skizon generasi kedua (Jeffers, 1986; Shirley dan Long, 1990).
9. Antibodi monoklonal
Antibodi merupakan suatu protein yang dibentuk oleh berbagai jenis hewan sebagai bagian dari reaksi kekebalan spesifik terhadap substansi asing (Zola, 1987), dan spesifikasi merupakan ciri utama dari reaksi kekebalan. Metode konvensional untuk memproduksi antibodi adaiah dengan menginfeksikan antigen ke dalam tubuh hewan percobaan untuk merangsang timbulnya tanggap kebal. Antibodi (imunoglobulin) dapat dimurnikan secara biokimiawi (Zola, 1987; Lopes dan Alves, 1984). Hasil akhirnya bukan merupakan antibodi yang murni terhadap antigen yang diberikan, tetapi terdiri dari berbagai antibodi dan jumiahnya terbatas (Sardjoko, 1991). Menurut Lopes dan Alves (1984) reaksi yang heterogen ini mempunyai keterbatasan apabila dibutuhkan spesifisitas antibodi yang lebih besar. Pada hewan, anhbodi dihasilkan oleh sel plasma yang nerupakan diferensiasi dari limfosit B. Sel plasma tidak dapat dibiakkan dalam biakan jaringan, sehingga tidak dapat dipakai sebagai sumber antibodi secara in vitro (Harlow dan David, 1988).
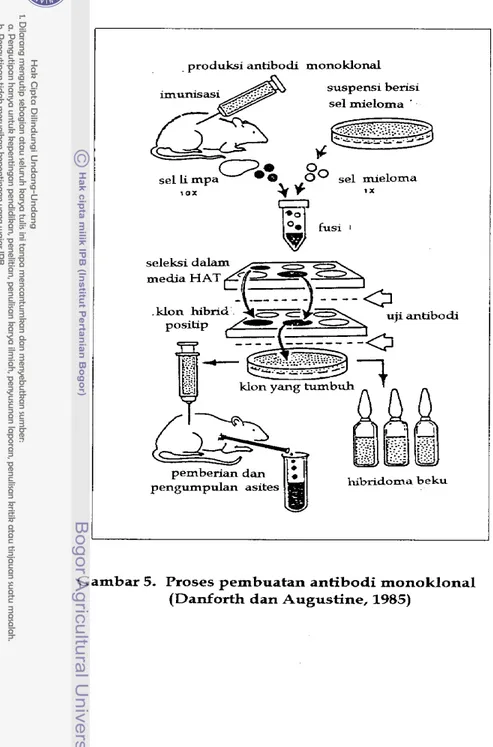
Pada tahun 1975, George Kohler dan Cesar Milstein berhasil menemukan penggabungan dua tipe sel yaitu sel limfosit B dan sel kanker (Smith, 1988). Gabungan tersebut, yang kemudian disebut sebagai hibridoma, yang mempunyai kemampuan untuk tumbuh dalam biakan sel dan dapat mensekresikan antibodi dalam jumlah besar dengan spesifisitas tertentu (Lopes dan Alves, 1984). Penggabungan (fusi) dua macam sel ini merupakan dasar
untuk pembuatan antibodi monoklonal (NRC, 1987), yaitu antibodi yang diproduksi oleh hibridoma (Harlow d a n David, 1988). Masing-masing hibridoma mempunyai kemampuan untuk tumbuh yang tidak terbatas dalam biakan jaringan, sehingga dapat memproduksi antibodi monoklonal dalam jumlah yang hampir tidak terbatas (Sardjoko, 1991). Dengan demikian antibodi monoklonal merupakan reagen m u r N (Zola, 1987). Penggunaannya antara lain untuk mengidentifikasi dan menggandakan molekul solubel pada campuran, purifikasi atau untuk aplikasi k l i ~ s (Zola, 1987). Menurut Lopes dan Alves (1984), imunoparasitologi merupakan contoh yang baik untuk menggunakan tehnik ini.
Pusi antara sel mieloma dengan sel pengekskresi antibodi dapat berhasil dengan adanya agen fusi (Harlow dan David, 1988; Lopes d a n Alves, 1984). Zat kimia seperti ion kalsium, lisolesitin d a n glikol polietilen dapat digunakan sebagai agen fusi (Sardjoko, 1991). Sekarang yang paling banyak digunakan adalah glikol polietilen (Harlow d a n David, 1988; Lopes d a n Alves, 2984; Sardjoko, 1991), karena bereaksi cepat dan penanganannya mudah (Harlow dan David, 1988).
Pemilihan jenis hewan terutama bergantung kepada tersedianya galur biakan jaringan yang sesuai untuk fusi (Sardjoko, 1991). Sampai saat ini hanya tersedia galur mieloma tikus dan mencit. Mieloma d a p a t diinduksi pada mencit dengan menyuntikkan minyak mineral ke dalam peritoneum (Harlow dan
David, 1988; Sardjoko, 1991). Fusi silang (antar jenis yang berbeda) dapat dilakukan, tetapi hasilnya tidak efisien. Untuk mencegah terbentuknya hibridoma yang mensekresikan lebih dari satu tipe antibodi, mieloma yang digunakan telah diseleksi. MieIoma ini tidak mengandung antibodi fungsional (Harlow d a n David, 1988).
Organ yang dipilih adalah limpa, karena organ ini mengandung limfosit B dalam jumlah besar (Lopes dan Alves, 1984). Pada fusi juga dihasilkan produk-produk lain yang tidak diperlukan (Drlica, 1984). Menurut Harlow dan David (1988), pada fusi yang efisien sekalipun, kurang lebih hanya 1% sel-sel permulaan yang bergabung. Sel-sel limpa tidak berkembang dalam biakan jaringan, dengan demikian tidak mengganggu perkembangan hibridoma. Akan tetapi sel mieloma berkembang terus dalam biakan jaringan (Harlow dan David, 1988; Lopes dan Alves, 1984; Drlica, 1984), sehingga dibutuhkan suatu cara untuk seleksi. Untuk menseleksi h n g g a hanya sel hibridoma yang tumbuh, digunakan metode Littlefield (Lopes dan Alves, 1984; Zola, 1987; Goding, 1986). Jalur utama sintesis nukleotida dihambat oleh aminopterin sebagai antagonis dari asam folat, yang terdapat dalam medium HAT (Hypoxantin- aminopterin-Ttumidin). Sintesis dilanjutkan dengan menggunakan jalur "salvage" dengan adanya thimidin dan hypoxantin. Keadaan ini dipengaruhi oIeh enzim hypoxathin-guanine-fosforibosil-transfers (HGPRT) dan thymidin kinase (TK). Apabila tidak terdapat salah satu dari enzim-enzim ini, maka sel
akan mati dalam media HAT. Hanya hibrid antara mieloma dan sel limpa yang dapat bertahan hidup dalam media HAT. Sel limpa memiliki enzim HGPRT
dan TK, sedangkan sel mieloma memiliki kemampuan untuk tumbuh (Lopes dan Alves, 1984).
Metode ini sudah banyak dipelajari dalam kaitannya dengan penyakit koksidiosis. Penelitian lebih banyak menggunakan stadium sporozoit mengingat stadium ini yang melakukan penetrasi pada sel induk semang setelah sel ookista terekskistasi (Danforth dan Augustine, 1985). Antibodi yang diperoleh digunakan antara lain untuk menentukan spesifisitas stadium (Danforth, 1983), menentukan lokasi antigen sporozoit (Danforth, Barta dan Augustine, 1992), untuk mempelajari pengaruhnya terhadap penetrasi sporozoit pada jaringan secara in vitro (Augustine, 1986; Crane, Norman, Gnozzio, Tate, Gammon dan Murray, 1986). Gambar 5 menunjukkan skema dari proses pembuatan antibodi monoklonal.
produksi antibodi monoklonal
suspensi berisi sel rnieloma '
sel Li mpa
%
SO
sel mieloma, o x
% *
1 X'r?-al'
hibridoma b eku
Gambar 5. Proses pembuatan antibodi monoklonal (Danforth dan Augustine, 1985)