Analisis Molekuler dan Keragaan Fenotipe Galur
Padi Gogo BC
3
F
2
yang Membawa Lokus Alt dan Pup1
(Molecular Analysis and Phenotypic Performances of BC
3F
2Upland
Rice Lines Containing Alt and Pup1 Loci)
Muhammad Rauful Mizan1*, Desta Wirnas2, Tasliah3, Nurul Hidayatun3, dan Joko Prasetiyono3 1Program Studi Pemuliaan dan Bioteknologi Tanaman, Sekolah Pascasarjana Institut Pertanian Bogor,
Jl. Meranti, Kampus IPB Dramaga, Bogor 16680 Indonesia
Telp. (0251) 8629347; Faks. (0251) 8629347; *E-mail: [email protected]
2Departemen Agronomi dan Hortikultura, Fakultas Pertanian, Institut Pertanian Bogor, Jl. Meranti, Kampus IPB Dramaga, Bogor 16680 Indonesia 3Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumber Daya Genetik Pertanian, Jl. Tentara Pelajar 3A, Bogor 16111 Indonesia
Diajukan: 25 Juni 2019; Direvisi: 25 November 2019; Diterima: 6 Desember 2019
ABSTRACT
The challenges in upland rice cultivation are phosphorous (P) deficiency and aluminum (Al) toxicity, each controlled by Pup1
and Alt loci, respectively. Pyramiding the two genes into Indonesian rice varieties were previously done through Marker-Assisted Backcrossing method to obtain BC3F2 populations. The aims of this study were to analyze the BC3F2 upland rice lines
containing the Alt and Pup1 loci molecularly (foreground and background analyses) and to test their phenotypic performances. Genetic materials tested included Dupa variety (donor of Alt) and three improved Indonesian genotypes (Dodokan-Pup1, Situ Bagendit-Pup1, and Batur-Pup1) as recurrent parents, Kasalath (donor of Pup1), and 300 BC3F2 lines from
Dodokan-Pup1+Alt, Situ Bagendit-Pup1+Alt, and Batur-Pup1+Alt, respectively. The rice genotypes were selected individually using modified Yoshida nutrient solution, followed by foreground and background analyses. 150 out of 300 seedlings were selected and maintained until harvest in the greenhouse. Foreground analysis using markers (RM1361, RM12031, and Kas46-2) and tiller number performances resulted in 18 plants from BC3F2 Dodokan-Pup1+Alt, 30 plants from BC3F2 Situ
Bagendit-Pup1+Alt, and 25 plants from BC3F2 Batur-Pup1+Alt still carrying Alt and Pup1 loci. Background analysis using molecular
markers showed that the best individual lines of BC3F2 were number 56 for BC3F2 Dodokan-Pup1+Alt, number 32 or 70 for
BC3F2 Situ Bagendit-Pup1+Alt, and number 20 for BC3F2 Batur-Pup1+Alt. The selected lines having both both Alt and Pup1 loci
in homozygote condition with highest number of tiller per plant which are useful genetic materials for developing upland rice variety tolerance to low P and Al toxicity.
Keywords: Upland rice, Pup1, Alt, phosphorus deficiency, aluminum toxicity.
ABSTRAK
Lahan marginal Indonesia umumnya berupa lahan kering masam dengan kadar fosfor (P) rendah dan aluminium (Al) tinggi. Penggabungan (pyramiding) dua gen (Pup1 dan Alt) ke dalam tiga varietas padi Indonesia telah dilakukan melalui silang balik dengan bantuan marka dan telah mencapai generasi BC3F2.Penelitian ini bertujuan menganalisis galur BC3F2 padi gogo yang
membawa lokus gen Alt and Pup1 secara molekuler (analisis foreground dan background) dan keragaaan fenotipe toleransi terhadap keracunan Al dan komponen hasil berdasarkan jumlah anakan terbanyak per tanaman. Kegiatan ini menggunakan varietas Dupa (donor Alt) dan tiga genotipe padi Indonesia yang telah diperbaiki (Dodokan-Pup1, Situ Bagendit-Pup1, dan Batur-Pup1) sebagai tetua penerima, Kasalath (donor Pup1), dan 300 individu dari setiap populasi BC3F2 Dodokan-Pup1+Alt,
Situ Bagendit-Pup1+Alt, dan Batur-Pup1+Alt. Genotipe padi diseleksi dengan menggunakan larutan hara Yoshida yang di-modifikasi, dilanjutkan dengan analisis foreground dan background. Sebanyak 150 genotipe dari 300 genotipe terpilih dan ditanam di rumah kaca hingga panen. Berdasarkan seleksi marka foreground (RM1361, RM12031, dan Kas46-2) dan jumlah anakan terbanyak, terseleksi 18 individu BC3F2 Dodokan-Pup1+Alt, 30 individu BC3F2 Situ Bagendit-Pup1+Alt, dan 25 individu
BC3F2 Batur-Pup1+Alt yang masih memiliki lokus Alt dan Pup1. Seleksi berdasarkan marka background menunjukkan bahwa
individu terbaik dari populasi BC3F2 tersebut yaitu nomor 56 untuk BC3F2 Dodokan-Pup1+Alt, nomor 32 atau 70 untuk BC3F2
Situ Bagendit-Pup1+Alt, dan nomor 20 untuk BC3F2 Batur-Pup1+Alt. Individu terbaik tersebut telah memiliki alel dari lokus Alt
dan Pup1 dalam kondisi homozigot dan memiliki jumlah anakan terbanyak. Data-data penelitian ini sangat penting untuk memperoleh informasi galur-galur yang akan dipilih untuk ditanam pada periode berikutnya.
Kata kunci: Padi gogo, Pup1, Alt, kahat fosfor, keracunan aluminium.
PENDAHULUAN
Makanan pokok penduduk dunia saat ini dido-minasi oleh beras. Kebutuhan beras pada tahun 2050 nanti diperkirakan akan menjadi dua kali lipat daripada kebutuhan sekarang sehingga peningkatan produksi yang lebih signifikan diperlukan untuk mengimbangi peningkatan jumlah penduduk (Alexandratos dan Bruinsma 2012). Di Indonesia, untuk mencukupi kebutuhan beras nasional peme-rintah mengandalkan pertanaman padi di sawah dan lahan kering. Tercatat luas panen padi tahun 2017 sebesar 15.712.015 hektar, dengan luas panen padi sawah sebesar 14.555.996 hektar dan padi gogo sebesar 1.156.019 hektar. Hal ini menunjukkan luas panen padi gogo hanya sebesar 7,36% dari luas panen padi nasional. Total panen padi sawah pada tahun 2017 sebesar 77.366.049 ton gabah kering giling (GKG) dan padi gogo sebesar 3.782.542 ton GKG (Kementan 2018).
Potensi lahan kering masam di Indonesia sangat besar. Berdasarkan identifikasi lahan suboptimal, dari 189,2 juta hektar daratan Indonesia, sekitar 108,8 juta hektar termasuk lahan kering masam yang tersebar di pulau Sumatra, Kalimantan, dan Papua (Mulyani dan Sarwani 2013). Pada umumnya, budi daya padi gogo tidak seintensif padi sawah. Bahkan, penanaman padi gogo kebanyakan dilakukan di lahan-lahan ku-rang subur yang lebih dikenal dengan lahan marginal dengan ketersediaan hara yang sangat minim. Selain itu, tingkat keasaman tanah yang tinggi menyebabkan kelarutan Al menjadi tinggi sehingga konsentrasi Al3+ dalam tanah semakin tinggi. Selain meracuni tanam-an padi melalui penghambattanam-an pertumbuhtanam-an akar, Al3+ juga akan mengikat unsur-unsur lain dalam tanah yang berbentuk ion negatif seperti fosfor. Ion fosfor yang seharusnya tersedia banyak, dijerap dalam tanah dan ketersediaannya untuk tanaman menjadi rendah (Hanafiah 2010). Efek keracunan Al membuat akar tidak berkembang sehingga menyebabkan ta-naman kesulitan menyerap air dan unsur hara dari dalam tanah. Akibatnya, produktivitas tanaman padi di lahan masam menjadi rendah. Salah satu cara mengatasi hal ini ialah dengan menanam padi yang toleran terhadap keracunan Al.
Eksplorasi sumber daya genetik padi yang mem-bawa lokus toleran terhadap keracunan Al telah di-lakukan dan ditemukan genotipe padi lokal Indonesia yang toleran keracunan Al, yakni Dupa. Dupa merupa-kan padi lokal Indonesia yang berasal dari Kalimantan Timur dan toleran terhadap keracunan Al. Berdasarkan bentuk bulirnya, padi ini digolongkan ke dalam padi tipe tropical japonica (Hairmansis et al.
2005). Genotipe Dupa akhir-akhir ini digunakan sebagai varietas cek toleran keracunan Al termasuk pada penelitian rumah kaca dengan menggunakan larutan hara Yoshida (Prasetiyono et al. 2012). Hasil eksplorasi lokus-lokus terkait toleransi terhadap ke-racunan Al pada padi Dupa berupa QTL yang berkait-an dengberkait-an karakter pertumbuhberkait-an akar. Karakter ini diketahui terletak pada kromosom 1 (2 lokasi QTL), kromosom 3 (1 lokasi QTL), kromosom 6 (1 lokasi QTL), dan kromosom 7 (1 lokasi QTL). Nilai logarithm of the odds (LOD) tertinggi terdapat pada QTL yang dideteksi pada kromosom 1, yaitu pada posisi 156,00– 158,19 cM (Hidayatun 2014). Sementara, toleransi terhadap ke-racunan Al pada padi Cabacu, QTL yang berkaitan dengan perkembangan akar terletak pada kromosom 1, 9, dan 12, dengan marka molekuler yang terpaut erat dengan karakter toleran keracunan Al terletak pada kromosom 1 (Yohana et al. 2018).
Untuk meningkatkan toleransi tanaman padi ter-hadap kondisi tanah yang kahat P, telah dilakukan eksplorasi lokus-lokus yang bertanggung jawab terha-dap sistem perakaran yang berkembang bagus dengan jumlah akar lebih banyak pada kondisi tanah kahat P dan tingkat keasaman tanah rendah. Lokus tersebut ialah Phosphorus uptake 1 (Pup1) yang di dalam lokus tersebut terdapat gen Phosphorus-Starvation Tolerance (PSTOL1) (Wissuwa et al. 1998; Heuer et al. 2009; Gamuyao et al. 2012). Analisis berbagai genotipe padi menunjukkan bahwa gen PSTOL juga terdapat pada genotipe lain, seperti padi liar Oryza rufipogon (Neelam et al. 2017) dan varietas unggul Indonesia (Tasliah et al. 2016). Bahkan, gen PSTOL1 diketahui berkaitan dengan peningkatan toleransi tanaman padi terhadap penyakit blas dan hawar daun bakteri (Chithrameenal et al. 2018). Gen PSTOL1 juga terdeteksi pada tanaman jagung (Azevedo et al. 2015).
Penggabungan (pyramiding) kedua lokus Pup1 dan Aluminum tolerance (Alt) ke dalam satu genotipe padi Indonesia menjadi salah satu alternatif untuk meningkatkan toleransi tanaman padi terhadap ke-racunan Al ketika ditanam di lahan Ultisol dengan pH rendah. Peran lokus Alt akan mengeluarkan asam organik untuk melepaskan P yang terikat oleh Al (Hidayatun 2014) sehingga meningkatkan ketersedian P di dalam tanah. Lokus Pup1 merangsang pertum-buhan akar yang lebih panjang dan lebih banyak, dan dengan sistem perakaran yang demikian akan me-nangkap P tersedia di dalam tanah yang telah dilepas oleh lokus Alt. Kegiatan penelitian pyramiding kedua lokus (Pup1 dan Alt) ini telah dilakukan sejak tahun 2014 dan sampai saat ini telah diperoleh populasi BC3F2 (Prasetiyono et al. 2015; Amdela 2016; Mizan et
al. 2019). Pada kegiatan penelitian sebelumnya telah diperoleh tiga genotipe padi yang memiliki lokus Pup1, yaitu Dodokan-Pup1, Situ Bagendit-Pup1, dan Batur-Pup1. Genotipe padi ini telah digunakan se-bagai tetua betina pada program pemuliaan dengan teknik Marker-Assisted Backcrossing (MAB). Varietas lokal Dupa telah digunakan sebagai tetua jantan yang merupakan donor lokus Alt. Sampai saat ini, kegiatan penelitian pyramiding dua gen ini telah menghasilkan tiga galur BC3F2, yaitu Dodokan-Pup1+Alt, Situ Bagendit-Pup1+Alt, dan Batur-Pup1+Alt. Tujuan penelitian ini ialah menganalisis galur BC3F2 padi gogo yang membawa lokus gen Alt and Pup1 secara molekuler (analisis foreground dan background) dan keragaaan fenotipe toleransi terhadap keracunan Al dan komponen hasil berdasarkan jumlah anakan ter-banyak per tanaman.
BAHAN DAN METODE Waktu dan Tempat Penelitian
Penelitian dilaksanakan di Laboratorium Biologi Molekuler dan Rumah Kaca Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumber Daya Genetik Pertanian (BB Biogen), Bogor, Jawa Barat. Waktu pelaksanaan dimulai pada bulan Januari sampai dengan Juni 2016.
Materi Penelitian
Materi genetik yang digunakan dalam penelitian ini ialah 300 individu BC3F2 Dodokan-Pup1+Alt, 300 individu BC3F2 Situ Bagendit-Pup1+Alt, 300 individu BC3F2 Batur-Pup1+Alt), dan tetua-tetuanya, yaitu Dupa (donor lokus Alt, toleran terhadap keracunan Al), Kasalath (donor lokus Pup1, toleran terhadap ka-hat P). Dodokan, Situ Bagendit, dan Batur merupakan varietas unggul yang telah dilepas di Indonesia yang keunggulannya telah dideskripsikan di dalam buku deskripsi padi (Suprihatno et al. 2010). Sementara, Dodokan-Pup1, Situ Bagendit-Pup1, dan Batur-Pup1 merupakan varietas unggul yang telah memiliki lokus Pup1. Marka molekuler untuk seleksi foreground yang digunakan disajikan pada Tabel 1.
Uji Toleransi Populasi BC3F2 terhadap Keracunan
Al pada Larutan Hara Yoshida
Tiga ratus benih dari setiap populasi BC3F2 Dodokan-Pup1+Alt, BC3F2 Situ Bagendit-Pup1+Alt, dan BC3F2 Batur-Pup1+Alt ditumbuhkan di dalam cawan petri selama 3 hari, lalu diletakkan ke dalam lubang styrofoam yang bagian bawahnya telah diberi kain kasa nyamuk. Styrofoam tersebut diapungkan di dalam wadah berisi larutan hara Yoshida (Yoshida et al. 1976) sebanyak 10 liter yang dimodifikasi dengan konsentrasi P sebanyak 0,5 ppm dan Al sebanyak 60 ppm selama 2 minggu. Setiap 2 hari sekali, keasaman (pH) larutan diamati dan dijaga agar tetap pada pH 4, serta dilakukan penambahan air (H2O) apabila terjadi penurunan air di dalam wadah akibat penguapan. pH diatur dengan menambahkan NaOH atau HCl sesuai kebutuhan. Wadah yang berisi larutan hara Yoshida dilengkapi dengan aerator untuk menjaga homogeni-tas larutan. Perlakuan tanaman dalam larutan hara Yoshida dilakukan sampai tanaman berumur 2 minggu. Sebanyak 150 genotipe dipilih dari tiap popu-lasi BC3F2 yang memiliki keragaan akar terpanjang dibanding dengan tetua kontrol.
Analisis Foreground (Lokus Alt dan Pup1) dan Pengamatan Jumlah Anakan per Tanaman
Individu-individu terpilih hasil uji dengan larutan hara Yoshida dipindahkan ke dalam pot dengan media tanah yang telah dilumpurkan dan tanaman di-pelihara dengan seksama hingga panen. Tanaman terpilih berdasarkan uji hara Yoshida dianalisis foreground dengan marka pengapit SSR lokus Alt dan marka Kas46-2 untuk verifikasi keberadaan lokus Pup1. Karakteristik marka molekuler yang digunakan untuk seleksi foreground pada penelitian ini disajikan pada Tabel 1.
Analisis foreground untuk kedua lokus target pe-muliaan dilakukan melalui beberapa tahapan, yaitu isolasi DNA, amplifikasi DNA, elektroforesis gel, dan dokumentasi hasil elektroforesis dengan ChemiDoc EQ™ System (Bio-Rad). Isolasi DNA mengacu pada metode Dellaporta (Dellaporta et al. 1983) yang di-modifikasi. Proses PCR dilakukan pada volume 20 µl yang mengandung bufer 1×, dNTPs 100 µM, marka
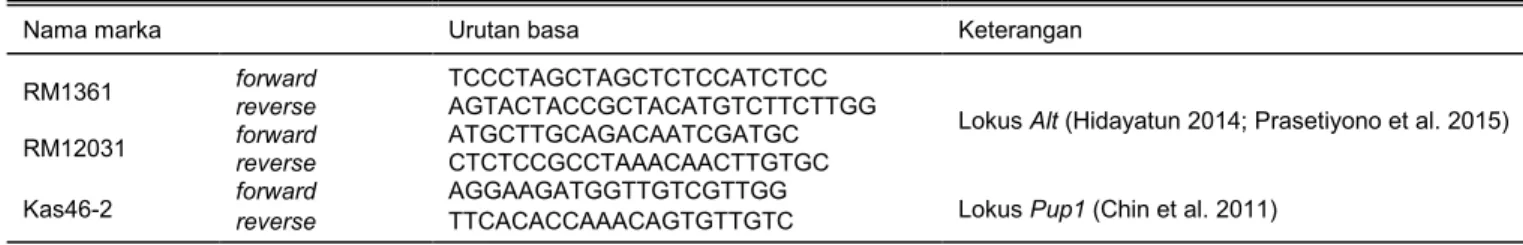
Tabel 1. Marka foreground yang digunakan untuk verifikasi lokus Alt dan Pup1.
Nama marka Urutan basa Keterangan
RM1361 forward TCCCTAGCTAGCTCTCCATCTCC
Lokus Alt (Hidayatun 2014; Prasetiyono et al. 2015)
reverse AGTACTACCGCTACATGTCTTCTTGG
RM12031 forward ATGCTTGCAGACAATCGATGC
reverse CTCTCCGCCTAAACAACTTGTGC
(forward dan reverse) 0,5 µM, 5 ng DNA, dan 1 unit Taq polimerase DNA. Program PCR yang digunakan, yaitu 5 menit pada 94°C sebagai denaturasi permula-an, dilanjutkan dengan 35 siklus yang terbagi menjadi 60 detik pada 94°C untuk denaturasi DNA cetakan, 60 detik pada 55°C untuk penempelan marka, dan 2 menit pada 72°C untuk perpanjangan marka. Perpan-jangan akhir dilakukan pada 72°C selama 7 menit. Produk PCR dipisahkan pada gel poliakrilamida 8% selama 2 jam dengan tegangan 80 volt. Pewarnaan DNA dilakukan dengan merendam gel dalam larutan etidium bromida dan didokumentasi dengan ChemiDoc EQ™ System (Bio-Rad).
Alel-alel galur BC3F2 dan jumlah anakan total di-amati untuk setiap galur BC3F2 pada persilangan masing-masing. Galur yang membawa alel-alel lokus Alt dan Pup1 dalam kondisi homozigot dan memiliki jumlah anakan terbanyak dipilih. Sebanyak 30 galur per populasi dari tiga populasi yang diuji kemudian dipilih dan digunakan pada analisis marka background.
Analisis Background Genom Tetua Pemulih Menggunakan Marka SSR
Tiga puluh genotipe BC3F2 hasil seleksi foreground dari tiap populasi yang memiliki alel Dupa dengan jumlah anakan terbanyak dianalisis kembali dengan marka SSR yang lokasinya tersebar pada seluruh kromosom padi (background). Marka mole-kuler yang digunakan untuk seleksi background adalah marka-marka yang polimorfik antara tetua pemulih dan tetua Dupa serta tersebar di seluruh kro-mosom (Prasetiyono et al. 2016). Hasil analisis marka backgroud tersebut merupakan gabungan dari anali-sis marka background pada generasi BC1F1 (Prasetiyono et al. 2016), BC2F1 (Amdela 2016), BC3F1 (Mizan et al. 2019), dan BC3F2.
Proses skoring dan visualisasi proporsi gen tetua pemulih dari hasil seleksi background BC3F2 akumulasikan dengan jumlah alel homozigot yang di-peroleh pada penelitian generasi BC2F1 (Amdela 2016) dan BC3F1 sebelumnya. Visualisasi dilakukan dengan perangkat lunak Graphical Genotypes (GGT) 2.0 (van Berloo 1999, 2008).
HASIL DAN PEMBAHASAN Keragaan Galur BC3F2 beserta Tetuanya
Berdasarkan Hasil Uji pada Seleksi Larutan Hara Yoshida
Hasil uji dalam larutan hara Yoshida menunjuk-kan bahwa varietas Dupa memiliki akar yang lebih
panjang dibanding dengan Kasalath, baik tetua pe-mulih maupun seluruh generasi BC3F2 (Tabel 2). Tetua Dupa merupakan tetua yang memiliki toleransi terhadap cekaman Al karena terbukti masih memiliki akar terpanjang dan berdasarkan penelitian sebelum-nya diketahui bahwa varietas Dupa merupakan jenis padi yang toleran terhadap keracunan Al karena menghasilkan senyawa malondialdehyde (MDA) yang lebih sedikit saat keracunan Al yang mengindikasikan bahwa varietas ini menyerap Al dalam jumlah rendah (Hidayatun 2014).
Gambar 1 adalah foto keragaan tetua Situ Bagendit-Pup1 beserta galur BC3F2 pada kondisi cekaman Al 60 ppm selama 2 minggu. Dari Gambar 1 terlihat bahwa mayoritas individu BC3F2 Situ Bagendit-Pup1+Alt memiliki keragaan akar yang lebih baik di-banding dengan tetuanya, termasuk pada populasi Dodokan-Pup1+Alt dan Batur-Pup1+Alt (data tidak ditampilkan). BC3F2 Situ Bagendit-Pup1+Alt akarnya lebih panjang dibanding dengan BC3F2 Dodokan-Pup1+Alt dan BC3F2 Batur-Pup1+Alt (Tabel 2). Pan-jang akar merupakan peubah yang paling banyak di-gunakan dalam pengamatan keracunan Al pada per-cobaan menggunakan larutan hara Yoshida (Wu et al. 2000; Nguyen et al. 2001; Famoso et al. 2010). Tanaman yang toleran akan memiliki pertumbuhan yang normal sehingga akarnya lebih panjang diban-ding dengan tanaman yang tidak toleran.
Hal ini diduga bahwa individu-individu populasi BC3F2 dari setiap persilangan telah membawa lokus Alt yang didapat dari varietas Dupa dan lokus Pup1 dari varietas Kasalath sebelumnya. Galur yang me-miliki lokus Pup1 biasanya menunjukkan penampilan akar yang bagus sehingga diharapkan efisien dalam penyerapan unsur P pada kondisi kahat (Kottearachchi dan Wijesekara 2013). Hal ini sejalan dengan percobaan yang dilakukan oleh Gamuyao et al. (2012) yang membandingkan Nipponbare (wild type) dengan Nipponbare yang ditambahi gen PSTOL1. Tanaman padi yang toleran terhadap kera-cunan Al umumnya memiliki akar yang panjang dan tidak ada pertumbuhan akar yang menggerombol di pangkal batang (Prasetiyono et al. 2012).
Analisis Foreground (Lokus Alt dan Pup1) dan Keragaan Jumlah Anakan per
Tanaman Galur BC3F2
Dari hasil analisis foreground galur BC3F2 Dodokan-Pup1+Alt menggunakan marka lokus Alt (RM12031 dan RM1361), diperoleh 34 individu dengan marka RM12031 dan 26 individu dengan marka RM1361 yang sama-sama memiliki alel tetua donor, dan diperoleh 25 individu memiliki alel pita Dupa
hasil uji foreground menggunakan kedua marka (Gambar 2). Pada BC3F2 Situ Bagendit-Pup1+Alt, dari 140 individu yang dianalisis, 49 individu di antaranya memiliki alel pita Dupa dengan marka RM12031 dan 47 individu berpita yang sama dengan marka RM1361. Pada kedua marka tersebut diketahui bahwa 47 individu yang sama di antaranya sama-sama memiliki pita Dupa. Pada BC3F2 Batur-Pup1+Alt terdapat 20 individu yang memiliki pita seperti Dupa, baik dengan marka RM12031 maupun RM1361, dan 18 individu di antaranya memiliki kedua pola pita seperti Dupa dengan kedua marka (Tabel 3).
Hasil amplifikasi PCR marka lokus Pup1 untuk ketiga populasi BC3F2 menggunakan marka verifikasi lokus Pup1 (Kas46-2) disajikan pada Gambar 3. Terlihat bahwa semua individu galur BC3F2, baik pada
populasi BC3F2 Dodokan-Pup1+Alt, BC3F2 Situ Bagendit-Pup1+Alt, maupun BC3F2 Batur-Pup1+Alt, pita lokusnya teramplifikasi dengan baik oleh marka Pup1, walaupun ada beberapa individu yang kurang sejajar dengan amplikon marka Kas46-2 sebesar 227 bp (Chin et al. 2011). Namun, kemungkinan besar lokus tersebut akan tetap ada dikarenakan tetua yang digunakan pada tiap persilangan balik merupakan tetua yang memiliki alel Pup1 (Prasetiyono 2010).
Proporsi tetua pemulih terbanyak merupakan kriteria utama dalam seleksi marka background un-tuk memilih individu yang akan digunakan pada ta-hapan seleksi selanjutnya. Setelah dilakukan serang-kaian tahapan seleksi, individu terpilih pada populasi BC3F2 Dodokan-Pup1+Alt ialah nomor 56 yang me-miliki jumlah anakan 13 dengan kumulatif marka
Tabel 2. Panjang akar rata-rata hasil seleksi dalam larutan hara Yoshida (Al 60 ppm dan P 0,5 ppm) pada populasi BC3F2.
Genotipe Panjang akar rata-rata (cm)
Dupa 12,2
Kasalath 0,83
Dodokan-Pup1 3,1
BC3F2 Dodokan-Pup1+Alt 3,4
Situ Bagendit-Pup1 2,5
BC3F2 Situ Bagendit-Pup1+Alt 3,7
Batur-Pup1 1,5
BC3F2 Batur-Pup1+Alt 2,4
Gambar 1. Perbandingan panjang akar Situ Bagendit-Pup1 beserta BC3F2 Situ Bagendit-Pup1+Alt pada kondisi Al 60 ppm.
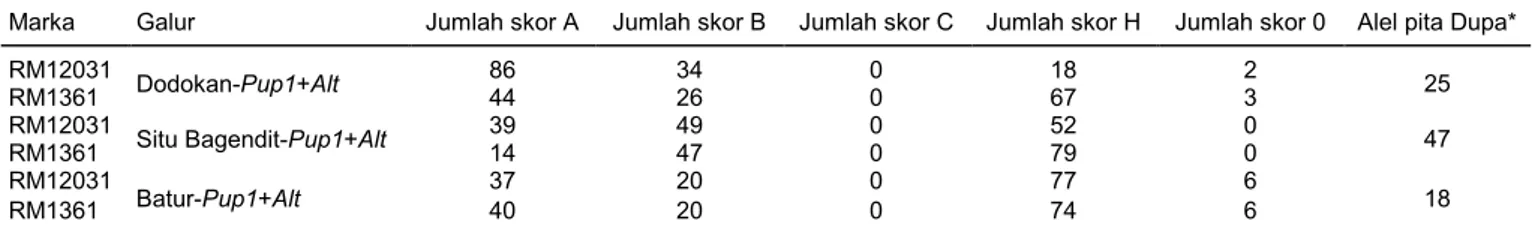
Tabel 1. Skoring pola pita marka foreground lokus Alt pada individu galur uji BC3F2.
Marka Galur Jumlah skor A Jumlah skor B Jumlah skor C Jumlah skor H Jumlah skor 0 Alel pita Dupa*
RM12031
Dodokan-Pup1+Alt 86 34 0 18 2 25
RM1361 44 26 0 67 3
RM12031
Situ Bagendit-Pup1+Alt 39 49 0 52 0 47
RM1361 14 47 0 79 0
RM12031
Batur-Pup1+Alt 37 20 0 77 6 18
RM1361 40 20 0 74 6
A = pita DNA mengikuti pita Dodokan-Pup1, Situ Bagendit-Pup1, atau Batur-Pup1; B = pita DNA mengikuti pita Dupa; C = pita DNA tidak mengikuti pola pita tetua pemulih dan tetua donor; H = heterozigot, apabila individu BC2F1 memiliki pita tetua pemulih dan tetua donor; 0 =
tidak ada pita DNA yang muncul; * = alel pita Dupa (B) pada kedua marka RM12031 dan RM1361 pada individu yang sama.
D u p a Ka s a la th S it u B d g t-P u p 1 BC3F2 Situ Bagendit-Pup1+Alt
homozigot sebanyak 64 marka (Gambar 4A). Pada BC3F2 Situ Bagendit-Pup1+Alt telah terpilih individu nomor 32 atau 70 yang sama-sama memiliki jumlah anakan 13 dengan kumulatif marka homozigot se-banyak 59 marka (Gambar 4B). Pada populasi BC3F2 Batur-Pup1+Alt terpilih individu nomor 20 yang me-miliki jumlah anakan 9 dengan kumulatif marka homozigot sebanyak 62 marka (Gambar 4C).
Analisis Background Genom Tetua Pemulih Menggunakan Marka SSR
Analisis background dilakukan pada semua nomor individu terpilih hasil seleksi marka foreground dan jumlah anakan terbanyak yang disajikan pada Tabel 4. Banyaknya marka yang digunakan pada setiap kromosom bergantung pada jumlah marka polimorfik yang dihasilkan pada seluruh marka yang diujikan sehingga banyaknya marka yang digunakan pada setiap persilangan dapat berbeda.
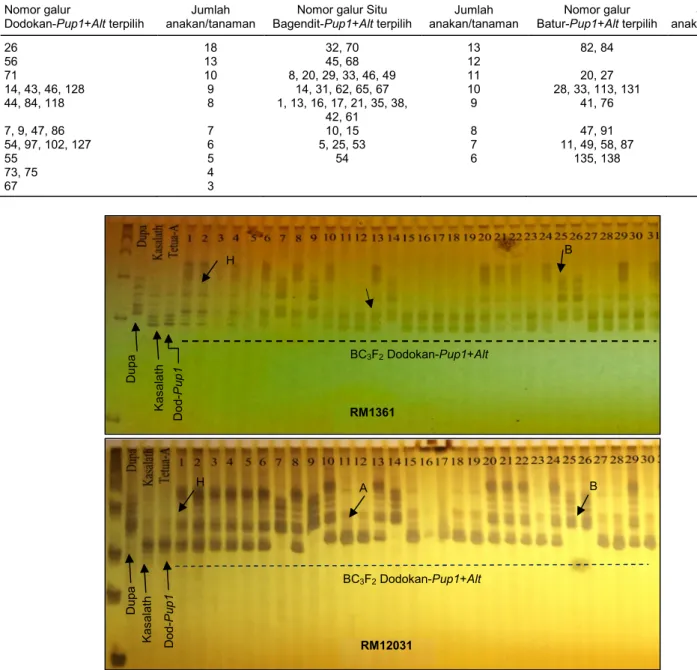
Tabel 4. Individu galur BC3F2 terpilih hasil analisis foreground menggunakan marka lokus Alt dan Pup1 dan jumlah anakan per tanaman
terbanyak galur BC3F2 terpilih.
Nomor galur
Dodokan-Pup1+Alt terpilih
Jumlah anakan/tanaman
Nomor galur Situ Bagendit-Pup1+Alt terpilih
Jumlah anakan/tanaman
Nomor galur Batur-Pup1+Alt terpilih
Jumlah anakan/tanaman 26 18 32, 70 13 82, 84 10 56 13 45, 68 12 71 10 8, 20, 29, 33, 46, 49 11 20, 27 9 14, 43, 46, 128 9 14, 31, 62, 65, 67 10 28, 33, 113, 131 8 44, 84, 118 8 1, 13, 16, 17, 21, 35, 38, 42, 61 9 41, 76 7 7, 9, 47, 86 7 10, 15 8 47, 91 6 54, 97, 102, 127 6 5, 25, 53 7 11, 49, 58, 87 5 55 5 54 6 135, 138 4 73, 75 4 67 3
Gambar 2. Pola pita marka pengapit lokus Alt (RM1361 dan RM12031) hasil analisis foreground individu galur populasi BC3F2
Dodokan-Pup1+Alt. A = alel Dodokan-Pup1, B = alel Dupa, H = heterozigot.
D u p a K a s a la th D o d -P u p 1
BC3F2 Dodokan-Pup1+Alt
H B D u p a K a s a la th D o d -P u p 1
BC3F2 Dodokan-Pup1+Alt
H A B
RM1361
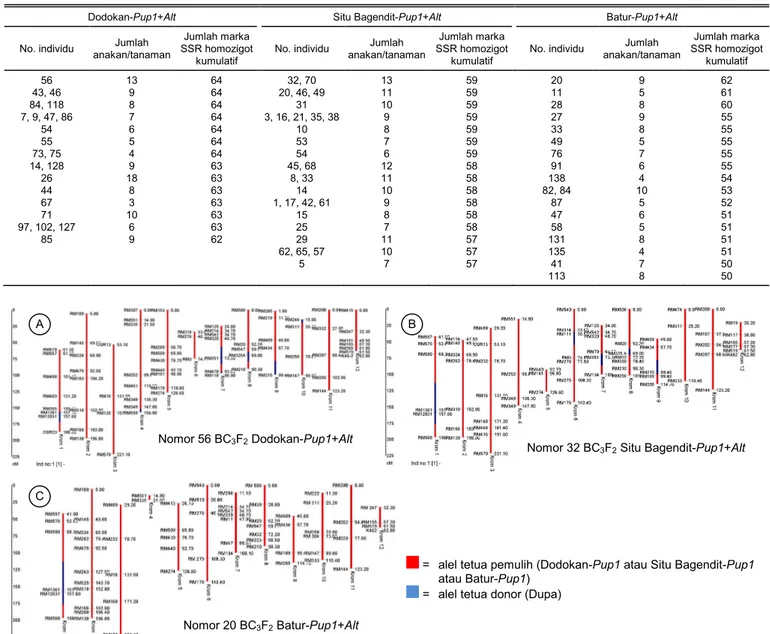
Jumlah marka yang digunakan pada analisis background pada setiap populasi dari tiga populasi BC3F2 yaitu 41 marka SSR polimorfik. Penggabungan jumlah alel marka SSR homozigot hasil penelitian ini dengan hasil penelitian pada populasi sebelumnya (BC2F1 dan BC3F1) memperoleh jumlah total marka SSR homozigot pada populasi BC3F2 Dodokan-Pup1+Alt mencapai lebih dari 60 marka, dengan marka homozigot tertinggi ditunjukkan oleh individu nomor 7, 9, 43, 46, 47, 54, 55, 56, 73, 75, 84, 86, dan 118 dengan 64 marka (Tabel 5). Pada populasi BC3F2 Situ Bagendit-Pup1+Alt jumlah rata-rata marka homozigot yang didapat lebih banyak dibanding dengan populasi BC3F2 Dodokan-Pup1+Alt. Jumlah rata-rata marka yang diperoleh sebanyak 21 marka tiap individu BC3F2, dengan individu nomor 3, 10, 16, 20, 21, 31, 32, 35, 38, 46, 49, 53, 54, dan 70 memiliki perolehan marka homozigot (22 marka) serta me-miliki jumlah kumulatif perolehan marka homozigot terbanyak (59 marka) hingga saat ini. Pada BC3F2 Batur-Pup1+Alt individu nomor 20 merupakan indivi-du yang paling banyak memperoleh marka homo-zigot, yaitu sebanyak 14 marka dengan perolehan marka homozigot kumulatif sebanyak 62 marka.
Secara teoritis, semakin banyak marka yang me-nunjukkan homozigot seperti alel tetua pemulih
pe-nampilan galur tersebut, semakin mirip dengan tetua-nya sehingga proses silang balik tidak perlu sampai BC5 atau BC6 (Hasan et al. 2015). Idealnya, untuk me-lakukan analisis background menggunakan marka SSR diperlukan marka polimorfik dengan jarak antar-marka sekitar 510 cM sehingga satu kromosom padi memerlukan marka polimorfik >10 marka tiap kromosom. Artinya, total marka yang diperlukan yaitu >120 marka polimorfik. Oleh karena analisis background dengan marka SSR ini memerlukan banyak tenaga (selain bahan kimia), marka-marka lain seperti marka SNP sekarang digunakan pula untuk kegiatan sejenis. Walaupun memerlukan biaya yang mahal, jumlah marka yang digunakan dapat mencapai ribuan untuk sekali running sehingga data yang diperoleh lebih cepat dan lebih banyak (Yu et al. 2014; Nadeem et al. 2018). Namun, masih banyak permasalahan dalam penelitian pemuliaan marka molekuler ini, seperti identifikasi marka QTL yang harus benar-benar tepat, desain marka untuk seleksi yang benar, dan bagaimana mentransfer QTL ke dalam genom varietas elit tanpa membawa sifat buruk tetua donor (Cobb et al. 2019).
Ketiga individu galur BC3F2 terpilih berdasarkan hasil analisis foreground dengan marka gen Pup1 dan
Gambar 3. Pola pita marka Kas46-2 untuk verifikasi lokus Pup1 hasil analisis foreground pada 25 individu masing-masing galur BC3F2. A = Dupa, B = Kasalath, C = Dodokan-Pup1 atau Situ Bagendit-Pup1
atau Batur-Pup1. Nomor pada gambar menunjukkan nomor galur uji.
A B C 7 9 14 25 26 35 43 44 46 47 54 55 56 67 71 73 75 84 85 86 97 101 118 127 128
A B C 11 20 27 28 33 41 47 49 58 76 82 84 87 91 113 131 135 138
BC3F2 Dodokan-Pup1+Alt
BC3F2 Situ Bagendit-Pup1+Alt
BC3F2 Batur-Pup1+Alt
Alt1, analisis background marka SSR, dan keragaan jumlah anakan terbanyak akan dilanjutkan pada program pemuliaan selanjutnya untuk merakit varie-tas unggul padi gogo produktivivarie-tas tinggi dan toleran terhadap keracunan Al dan kahat unsur P.
KESIMPULAN
Analisis molekuler foreground, background, dan analisis keragaan fenotipe toleransi terhadap kera-cunan Al dan komponen hasil jumlah anakan per tanaman menghasilkan satu galur masing-masing dari tiga populasi BC3F2 yang diuji. Individu galur ter-baik dari setiap populasi BC3F2 adalah nomor 56
untuk populasi BC3F2 Dodokan-Pup1+Alt, nomor 32 atau 70 untuk populasi BC3F2 Situ Bagendit-Pup1+Alt, dan nomor 20 untuk populasi BC3F2 Batur-Pup1+Alt. Seluruh individu galur terpilih tersebut memiliki akar terpanjang, jumlah anakan terbanyak, dan proporsi tetua pemulih terbesar berdasarkan analisis background. Semua galur terpilih telah terkonfirmasi membawa lokus Alt dan Pup1 berdasarkan analisis foreground menggunakan marka kedua lokus terse-but. Galur-galur tersebut potensial untuk dikembang-kan pada perakitan varietas padi gogo toleran ter-hadap keracunan Al dan kahat P dengan komponen hasil baik yang ditunjukkan oleh jumlah anakan yang banyak.
Tabel 5. Rekapitulasi jumlah anakan dan kumulatif marka homozigot hasil analisis background menggunakan marka SSR polimorfik pada populasi B2F1, BC3F1,
dan BC3F2 pada populasi BC3F2.
Dodokan-Pup1+Alt Situ Bagendit-Pup1+Alt Batur-Pup1+Alt
No. individu Jumlah anakan/tanaman
Jumlah marka SSR homozigot
kumulatif
No. individu Jumlah anakan/tanaman
Jumlah marka SSR homozigot
kumulatif
No. individu Jumlah anakan/tanaman Jumlah marka SSR homozigot kumulatif 56 13 64 32, 70 13 59 20 9 62 43, 46 9 64 20, 46, 49 11 59 11 5 61 84, 118 8 64 31 10 59 28 8 60 7, 9, 47, 86 7 64 3, 16, 21, 35, 38 9 59 27 9 55 54 6 64 10 8 59 33 8 55 55 5 64 53 7 59 49 5 55 73, 75 4 64 54 6 59 76 7 55 14, 128 9 63 45, 68 12 58 91 6 55 26 18 63 8, 33 11 58 138 4 54 44 8 63 14 10 58 82, 84 10 53 67 3 63 1, 17, 42, 61 9 58 87 5 52 71 10 63 15 8 58 47 6 51 97, 102, 127 6 63 25 7 58 58 5 51 85 9 62 29 11 57 131 8 51 62, 65, 57 10 57 135 4 51 5 7 57 41 7 50 113 8 50
Gambar 4. Hasil analisis proporsi genetik individu terbaik galur BC3F2.
Nomor 56 BC3F2 Dodokan-Pup1+Alt
Nomor 32 BC3F2 Situ Bagendit-Pup1+Alt
= alel tetua pemulih (Dodokan-Pup1 atau Situ Bagendit-Pup1 atau Batur-Pup1)
= alel tetua donor (Dupa) Nomor 20 BC3F2 Batur-Pup1+Alt
A B
UCAPAN TERIMA KASIH
Penelitian ini dibiayai melalui DIPA BB Biogen TA 2016 dengan judul ROPP “MAS Padi Efisien Fosfor (Pup1) dan Toleran Aluminium (Alt): Seleksi Rumah Kaca dan Lapang Galur-galur Hasil Persilangan”.
KONTRIBUTOR PENULISAN
MRM: kontributor utama, pelaksana kegiatan dan penulisan. DW: kontributor anggota, analisis data dan penulisan. TAS: kontributor anggota, analisis mo-lekuler dan kegiatan rumah kaca. NH: kontributor anggota, analisis molekuler dan kegiatan rumah kaca. JP: kontributor utama, penanggung jawab kegiatan dan penulisan.
DAFTAR PUSTAKA
Alexandratos, N. & Bruinsma, J. (2012) World agriculture
towards 2030–2050: The 2012 revision. ESA Working Paper No. 12-03. Rome, FAO.
Amdela, E.P. (2016) Deteksi introgresi lokus Aluminium
Tolerance (Alt) dan Phosphorus Uptake 1 (Pup1) pada tiga populasi BC2F1 tanaman padi (Oryza sativa L.)
dengan bantuan marka molekuler. Skripsi S1. Universitas Gadjah Mada.
Azevedo, G.C., Cheavegatti-Gianotto, A., Negri, B.F., Hufnagel, B., de Silva, L.D., Magalhaes, J.V., Garcia, A.A.F., Lana, U.G.P., de Sousa, S.M. & Guimaraes, C.T. (2015) Multiple interval QTL mapping and
searching for PSTOL1 homologs associated with root
morphology, biomass accumulation and phosphorus
content in maize seedlings under low-P. BMC Plant
Biology. [Online] 15, 172. Available from: https://doi.org/10.1186/s12870-015-0561-y [Accessed 2 May 2019].
Badan Pusat Statistik (2014) Rata-rata konsumsi per kapita
seminggu beberapa macam bahan makanan penting, 20072015. [Online] Tersedia pada: https:// www.bps.go.id/-linkTabelStatis/view/id/950 [Diakses 29 Mei 2017].
Chin, J.H., Gamuyao, R., Dalid, C., Bustamam, M., Prasetiyono, J., Moeljopawiro, S., Wissuwa, M. & Heuer, S. (2011) Developing rice with high yield under
phosphorus deficiency: Pup1 sequence to application.
Plant Physiology, 156, 12021216.
Chithrameenal, K., Alagarasan, G., Raveendran, M., Robin, S., Meena, S., Ramanathan, A. & Ramalingam, J. (2018) Genetic enhancement of phosphorus starvation tolerance through marker assisted introgression of
OsPSTOL1 gene in rice genotypes harbouring
bacterial blight and blast resistance. PLoS ONE.
[Online] 13 (9), e0204144. Available from: https:// doi.org/10.1371/journal.pone.0204144 [Accessed 2 May 2019].
Cobb, J.N., Biswas, P.S. & Platten, J.D. (2019) Back to the future: Revisiting MAS as a tool for modern plant
breeding. Theoretical and Applied Genetics, 132, 647–
667.
Dellaporta, S.L., Wood, J. & Hicks, J.B. (1983) A plant DNA
minipreparation: Version II. Plant Molecular Biology,
1 (4), 19–21.
Famoso, A.N., Clark, R.T., Shaff, J.E., Craft, E., McCouch, S.R. & Kochian, L.V. (2010) Development of a novel aluminum tolerance phenotyping platform used for comparisons of cereal Al tolerance and investigations
into rice Al tolerance mechanisms. Plant Physiology,
153, 1678–1691.
Gamuyao, R., Chin, J.H., Tanaka, J.P., Pesaresi, P., Catausan, S., Dalid, C., Loedin, I.S., Mendoza, E.M.T., Wissuwa, M. & Heuer, S. (2012) The protein kinase
PSTOL1 from traditional rice confers tolerance of
phosphorus deficiency. Nature, 488, 535–539.
Hairmansis, A., Aswidinnoor, H., Trikoesoemaningtyas & Suwarno (2005) Evaluasi daya pemulih kesuburan
padi lokal dari kelompok tropical japonica. Buletin
Agronomi, 33 (3), 1–6.
Hanafiah, K.A. (2010) Dasar-dasar ilmu tanah. Jakarta,
Raja Grafindo Persada.
Hasan, M.M., Rafii, M.Y., Ismail, M.R., Mahmood, M., Rahim, H.A., Alam, Md.A., Ashkani, S., Malek, Md.A. & Latif, M.A. (2015) Marker-assisted backcrossing: A
useful method for rice improvement. Biotechnology &
Biotechnological Equipment, 29 (2), 237254.
Heuer, S., Lu, X., Chin, J.H., Pariasca-Tanaka, J., Kanamon, H., Matsumoto, T., Leon, T.D., Ulat, V.J., Ismail, A.M., Yano, M. & Wissuwa, M. (2009) Comparative sequence analyses of the major
quantitative trait locus Phosphorus uptake 1 (Pup1)
reveal a complex genetic structure. Plant
Biotechnology Journal, 7, 456–471.
Hidayatun, N. (2014) Genetics and physiological responses
of selected Indonesian rice (Oryza sativa Linn.) genotypes of aluminium toxicity and phosphorus deficiency experienced in acid soil condition. Ph.D. Dissertation. University of the Philippines Los Baños.
Kementerian Pertanian (2018) Statistik pertanian 2018.
Jakarta, Pusat Data dan Sistem Informasi Pertanian Kementerian Pertanian Republik Indonesia.
Kottearachchi, N.S. & Wijesekara, U.A.D.S.L. (2013)
Implementation of Pup1 gene based markers for
screening of donor varieties for phosphorus deficiency
tolerance in rice. Indian Journal of Plant Sciences,
2 (4), 7683.
Mizan, M.R., Wirnas, D. & Prasetiyono, J. (2019) Verifikasi
lokus Aluminum tolerance (Alt) pada tiga populasi
BC3F1 padi. Jurnal Agronomi Indonesia, 47 (1), 917.
Mulyani, A. & Sarwani, M. (2013) Karakteristik dan potensi lahan suboptimal untuk pengembangan pertanian di
Nadeem, M.A., Nawaz, M.A., Shahid, M.Q., Doğan, Y., Comertpay, G., Yıldız, M., Hatipoğlu, R., Ahmad, F., Alsaleh, A., Labhane, N., Özkan, H., Chung, G. & Baloch, F.S. (2018) DNA molecular markers in plant breeding: Current status and recent advancements in
genomic selection and genome editing. Biotechnology
& Biotechnological Equipment, 32 (2), 261285. Neelam, K., Thakur, S., Neha, Yadav, I.S., Kumar, K.,
Dhaliwal, S.S. & Singh, K. (2017) Novel alleles of
Phosphorus-Starvation Tolerance 1 gene (PSTOL1)
from Oryza rufipogon confers high phosphorus uptake
efficiency. Frontiers in Plant Science, 8 (509), 112.
Nguyen, V.T., Burow, M.D., Nguyen, H.T., Le, B.T., Le, T.D. & Paterson, A.H. (2001) Molecular mapping of genes
conferring aluminum tolerance in rice (Oryza sativa L.).
Theoretical and Applied Genetics, 102, 1002–1010.
Prasetiyono, J. (2010) Studi efek introgresi Pup1 (P uptake
1) untuk meningkatkan toleransi padi terhadap defisiensi P. Disertasi S3. Institut Pertanian Bogor. Prasetiyono J, Suhartini, T., Soemantri, I.H., Tasliah,
Moeljopawiro, S., Aswidinnoor, H., Sopandie, D. &
Bustaman, M. (2012) Evaluasi beberapa galur-Pup1
tanaman padi (Oryza sativa L.) pada larutan hara dan
lapangan. Jurnal Agronomi Indonesia, 40 (2), 83–90.
Prasetiyono, J., Somatri, I.H., Suhartini, T., Tasliah &
Ma’sumah (2015) Drought from a different perspective:
Improved tolerance through phosphorus acquisition dan developing rice with dual tolerance of phosphorus deficiency and aluminum toxicity. Laporan Akhir Proyek Generation Challenge Programme Tahun Anggaran 20132014. Bogor, Balai Besar Bioteknologi dan Sumber Daya Genetik Pertanian.
Prasetiyono, J., Somatri, I.H., Suhartini, T., Tasliah &
Ma’sumah (2016) MAS padi efisien fosfor (Pup1) dan
toleran aluminium (Alt). Laporan Kegiatan ROPP TA 2015, dengan RPTP Berjudul: Penciptaan Galur Unggul melalui Aplikasi Marka Molekuler. Bogor, Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumber Daya Genetik Pertanian.
Suprihatno, B., Daradjat, A.A., Satoto, Baehaki, Suprihanto,
Setyono, A., Indrasari, S.D., Wardana, I.P. &
Sembiring, H. (2010) Deskripsi varietas padi. Subang,
Balai Besar Penelitian Tanaman Padi.
Tasliah, Ma'sumah & Prasetiyono, J. (2016) Eksplorasi
lokus Pup1 pada 55 genotipe padi berdasarkan
analisis marka molekuler dan sekuensing. Jurnal
AgroBiogen, 12 (2), 6372.
van Berloo, R. (1999) GGT: Software for the display of
graphical genotypes. Journal of Heredity, 90 (2),
328329.
van Berloo, R. (2008) GGT 2.0: Versatile software for
visualization and analysis of genetic data. Journal of
Heredity, 99 (2), 232236.
Wissuwa, M., Yano, M. & Ae, N. (1998) Mapping of QTLs
for phosphorus-deficiency tolerance in rice (Oryza
sativa L.). Theoretical and Applied Genetics, 97, 777– 783.
Wu, P., Liao, C.Y., Hu, B., Yi, K.K., Jin, W.Z., Ni, J.J. & He, C. (2000) QTLs and epistasis for aluminum tolerance
in rice (Oryza sativa L.) at different seedling stages.
Theoretical and Applied Genetics, 100, 1295–1303. Yohana, K., Adisyahputra & Trijamiko, K.R. (2018) Validasi
QTL dan aplikasinya untuk perbaikan sifat toleran
keracunan Al pada padi (Oryza sativa L.). Bioma,
14 (1), 3748.
Yoshida, S., Forno, D.A., Cock, J. & Gomez, K.A. (1976)
Laboratory manual for physiological studies of rice. Los Baños, IRRI.
Yu, H., Xie, W., Li, J., Zhou, F. & Zhang, Q. (2014) A whole-genome SNP array (RICE 6K) for genomic breeding in