Review article
Developing zebra
fi
sh models of autism spectrum disorder (ASD)
Adam Michael Stewart
a,b, Michael Nguyen
c, Keith Wong
d, Manoj K. Poudel
a, Allan V. Kalueff
a,⁎
aZENEREI Institute and Zebrafish Neuroscience Research Consortium (ZNRC), 309 Palmer Court, Slidell, LA 70458, USAbDepartment of Neuroscience, University of Pittsburgh, A210 Langley Hall, Pittsburgh, PA 15260, USA cDepartment of Biomedical Engineering, University of Virginia, 415 Lane Road, Charlottesville, VA 22908, USA dUniversity of California San Diego (UCSD) School of Medicine, 9500 Gilman Dr, La Jolla, CA 92093, USA
a b s t r a c t
a r t i c l e
i n f o
Article history:
Received 30 September 2013
Received in revised form 22 November 2013 Accepted 28 November 2013
Available online 6 December 2013
Keywords:
Autism spectrum disorder Behavioral tests Social deficits Translational research Zebrafish
Autism spectrum disorder (ASD) is a serious neurodevelopmental disorder with complex symptoms and unclear, multi-factorial pathogenesis. Animal (rodent) models of ASD-like behavior are extensively used to study genet-ics, circuitry and molecular mechanisms of ASD. The evolutionarily conserved nature of social behavior and its molecular pathways suggests that alternative experimental models can be developed to complement and enhance the existing rodent ASD paradigms. The zebrafish (Danio rerio) is rapidly becoming a popular model organism in neuroscience and biological psychiatry to study brain function, model human brain disorders and explore their genetic or pharmacological modulation. Representing highly social animals, zebrafish emerge as a strong potential model organism to study normal and pathological social phenotypes, as well as several other ASD-like symptoms. Here, we discuss the developing utility of zebrafish in modeling ASD as a new emerg-ingfield in translational neuroscience and drug discovery.
© 2013 Elsevier Inc. All rights reserved.
Contents
1. Introduction . . . 27
2. Traditional experimental models relevant to ASD . . . 28
2.1. Genetic rodent models relevant to ASD . . . 28
2.2. Pharmacological rodent models relevant to ASD . . . 29
3. Zebrafish models relevant to ASD . . . 30
3.1. Behavioral and pharmacological models . . . 30
3.2. Physiological correlates . . . 31
3.3. Genetic models relevant to ASD . . . 33
3.4. Environmental models potentially relevant to ASD . . . 33
4. Conclusion . . . 33
Acknowledgements . . . 34
References . . . 34
1. Introduction
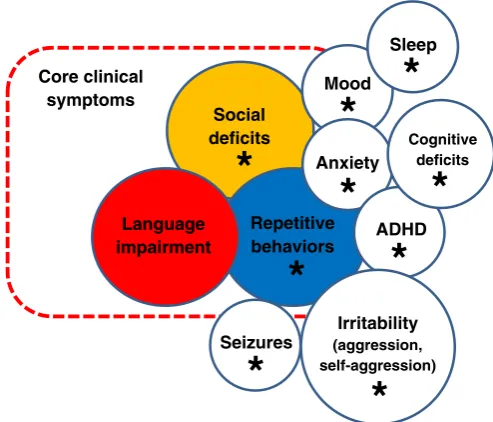
Autism spectrum disorder (ASD) is a serious debilitating mental ill-ness affecting approximately 1–2% of the general population (Evans, 2013; Mayes et al., 2011; Wing et al., 1967). Recently revisited by the American Psychiatric Association (2013), ASD represents a neurode-velopmental disorder characterized by impaired social communication, repetitive behavior and cognitive deficits (seeTable 1andFig. 1for details of clinical phenotypes associated with ASD). In addition to these core symptoms, ASD shows high (~90%) heritability, representing one of the most heritable brain disorders (Crawley, 2012; Edvardson et al., 2013). Notably, ASD is a polygenic disorder with multiple genetic determinants
–
Abbreviations:ADHD, attention deficit hyperactivity disorder; ASD, autism spectrum disorder; AVP, arginine vasopressin; DSM-5, Dianostic and Statistical Manual of mental disorders, 5th edition; FXS, fragile X syndrome; GABA, gamma-aminobutyric acid; MDMA, 3,4-methylenedioxy-N-methylamphetamine; NMDA, N-methyl-D-aspartate; PCP, phencyclidine; SERT, serotonin transporter; SFARI, Simons Foundation Autism Research Initiative; SSRIs, selective serotonin re-uptake inhibitors; V1aR, V1a receptor.
⁎Corresponding author at: ZENEREI Institute, 309 Palmer Court, Slidell, LA 70458, USA. Tel./fax: +1 240 328 2275.
E-mail address:[email protected](A.V. Kalueff).
0278-5846/$–see front matter © 2013 Elsevier Inc. All rights reserved.
http://dx.doi.org/10.1016/j.pnpbp.2013.11.014
Contents lists available atScienceDirect
Progress in Neuro-Psychopharmacology & Biological
Psychiatry
and candidate genes. For example, the SFARI Gene database (https:// gene.sfari.org) currently lists 546 genes associated with ASD. Thus, ASD has a complex, poorly understood pathogenesis and associated genetic/ environmental risk factors, aberrant brain circuits and disordered molec-ular pathways (Edvardson et al., 2013; Geschwind, 2008; Kesli et al., 2014; Matson et al., 2012).
Despite recent progress in dissecting the neural underpinnings of ASD (Kéïta et al., 2011; Kujala et al., 2013), its pharmacological therapy is complicated by the lack of efficient, disorder-specific and safe medica-tion (Benvenuto et al., 2013; Farmer et al., 2013). For example, the Food and Drug Administration approves two drugs (atypical neuroleptics ris-peridone and aripiprazole) for treating ASD-associated irritability (Crawley, 2012). Other agents, such as methylphenidate, selective serotonin re-uptake inhibitors (SSRIs), valproate, atomoxetine,α2 ad-renergic agonists and olanzapine, can also treat some ASD symptoms, but are not effective in alleviating others (Benvenuto et al., 2013; Myers, 2007).
Animal (experimental) models of brain disorders are an indispens-able tool for drug discovery and dissecting the pathogenic mechanisms of brain disorders (Kalueff et al., 2007; Kas et al., 2013; Silverman et al., 2012, 2013). Complementing traditional rodent models of brain disor-ders, new experimental approaches using zebrafish (Danio rerio) are rap-idly gaining popularity in neuroscience research (Gerlai, 2010a, 2011, Kalueff et al., in press). Can zebrafish be used to model complex brain dis-orders? For decades, zebrafish have been viewed as‘simple’organisms with relatively primitive, instinctively-driven behaviors suitable mainly for screening drugs, genetic mutations or developmental defects (see
Gaikwad et al., 2011; Kalueff et al., 2013;Stewart and Kalueff, 2012for discussion). However, mounting recent experimental evidence shows that zebrafish possess high genetic and physiological homology to mam-mals and display complex affective, social and cognitive responses which are similar to those observed in rodents and humans (Gerlai, 2010b, 2011; Kalueff et al., 2013 Pather and Gerlai, 2009;Stewart et al., 2013). Taken together, this suggests that zebrafish models can be used exten-sively in translational neuroscience research (Gerlai, 2010a; Kalueff et al., 2013, in press).
While various experimental (e.g., genetic or pharmacological) ma-nipulations model certain symptoms and/or disordered pathways of ASD, they do not reflect the entire disease state. However, several social, motor and cognitive phenotypes (Table 2) are commonly assessed in rodent models of ASD, providing important mechanistic insights into its neurobiology (Crawley, 2012; Kas et al., 2013); also see discussion of this further in the text. The evolutionarily conserved nature of social behavior and its molecular pathways suggests that novel experimental models can be developed to complement and enhance the existing rodent ASD paradigms. Can we use zebrafish to study ASD-related path-ogenesis? Addressing this important question, here we discuss recent advances and outline future promising directions of research in the
field of novel zebrafish models of ASD-like states.
2. Traditional experimental models relevant to ASD
2.1. Genetic rodent models relevant to ASD
Laboratory rodents are highly sociable animals, and therefore are useful to study normal and pathological social behaviors (Brodkin, 2007; Crawley, 2012; Fairless et al., 2013; Kas et al., 2013; McFarlane et al., 2008; Moy et al., 2008; Ryan et al., 2010; Silverman et al., 2012). With mouse and rat genomes now being fully characterized, various ge-netic models of ASD and related disorders have been developed. For ex-ample, the Fragile X Syndrome (FXS) is an inherited mental retardation disorder caused by a single mutation in the FMR1 gene in the X chromosome. FVB and C57BL/6 mice withFMR1genetic knockout dis-play some FXS social and behavioral symptoms (Bernardet and Crusio, 2006; Pietropaolo et al., 2011).
Loss-of-function mutations in theNlgn4 gene in mice encoding for Neuroligin-4 impair social behavior and vocal communication, establishingNlgn4mice as a genetic model of ASD-related pathogenesis (El-Kordi et al., 2012; Jamain et al., 2008) relevant to human genetic data linking this gene to ASD (Pampanos et al., 2009). Shank proteins are also involved in modulation of synaptic communication and
Table 1
Diagnostic criteria for autism spectrum disorder (ASD), according to the Diagnostic and Statistical Manual of Mental Disorders (DSM-5) of the American Psychiatric Association (2013).
A. Social deficits
Persistent deficits in social communication and interaction, including: i) deficits in social–emotional reciprocity and social approach, reduced sharing of interests, failure to initiate/respond to interactions; ii) deficits in nonverbal communication, poorly integrated verbal/nonverbal communication, poor eye contact and body language, deficits in understanding/use of gestures; a deficit in facial expressions and recognizing facial affect; iii) deficits in developing, maintaining and understanding relationships, problems with adjusting behavior to various social contexts, making friends and developing interest in peers.
B. Behavioral and cognitive perseverations
Restricted, repetitive patterns of behavior, interests or activities, manifested by: i) stereotyped/repetitive motor movements, use of objects or speech (e.g., simple motor stereotypies, idiosyncratic phrases); ii) insistence on sameness, routines, ritualized patterns in verbal/nonverbal behavior (e.g., distress at small changes, rigid thinking patterns, stable rituals); iii) highly restricted interests abnormal in intensity or focus (e.g., strong attachment to objects, excessively circumscribed or perseverative interest); iv) hyper/hyporeactivity to sensory input or sensory aspects of the environment (e.g., indifference to pain/temperature, adverse response to specific sounds or textures, excessive smelling/touching objects, visual fascination with lights or movement).
C. Symptoms' trajectory
Symptoms must be present in the early developmental period (but may not become fully manifest until social demands exceed limited capacities, or may be masked by learned strategies in later life); these symptoms cause clinically significant impairment in social, occupational, or other important areas of current functioning.
Mood
Sleep
Anxiety
ADHD
Seizures
Irritability
(aggression, self-aggression)Social
deficits
Repetitive
behaviors
Language
impairment
Core clinical
symptoms
Cognitive deficits
*
*
*
*
*
*
*
*
*
complex with Neuroligin. Loss-of-function mutation in theShank3gene, encoding for Shank protein 3, inhibits synaptic transmission and im-pairs mouse social behaviors, modeling the 22q13.3 deletion syndrome associated clinically with delayed development and speech impairment (Bozdagi et al., 2010).SHANK3is also of particular interest here because of its potential role in ASD (Gauthier et al., 2010) given recent human genetic studies linkingSHANK3to this disorder (Boccuto et al., 2013).
Serotonin transporter (SERT) gene knockout in C57BL/6J mice re-sults in increased grooming (Kyzar et al., 2012b) and causes social deficits in both SERT−/−mice and rats (Kalueff et al., 2010). SERT
variant Ala56 mice display differential serotonin neuronfiring patterns with altered social behavior, communication and repetitive behavior (Veenstra-VanderWeele et al., 2012). Integrinβ3 receptor interacts with SERT (Carneiro et al., 2008), and theITGB3knockout mice also show excessive grooming in novel environments (Carter et al., 2011).
In addition to mutant mice, several non-mutant mouse strains are relevant to ASD. For instance, the BTBR T+tf/J (BTBR) inbred mice
display core behavioral phenotypes of autism, including social deficits and stereotypic behavior (McFarlane et al., 2008; Meyza et al., 2012; Scattoni et al., 2012), which have been used in many studies relevant to ASD (McTighe et al., 2013; Schwartzer et al., 2013; Silverman et al., 2012). Systemic diazepam (Pobbe et al., 2011) or fluoxetine (Chadman, 2011) administration normalizes social activity in BTBR, but not in control C57BL/6 mice. Environmental factors, such as enriched housing in BTBR mice, reduce grooming duration, suggesting that enrichment normalizes certain repetitive behaviors (Reynolds et al., 2013). Social environmental factors also affect behavioral outcomes in mouse ASD models. For example, adolescent BTBR mice reared in the same housing as‘social’C57BL/6 strain exhibit markedly increased sociability, compared to BTBR adolescents reared with other BTBR mice. In contrast, adult BTBR mice do not show improvement in repetitive (self-grooming) behavior, suggesting that behavioral recov-ery differentially affects the social but not the motor ASD-like pheno-type in this model (Yang et al., 2011).
Likewise, the C58/J strain exhibits a behavioral phenotype re-sembling core ASD symptoms, including excess locomotor activity, social deficits, abnormal repetitive behaviors (Ryan et al., 2010), poor T-maze performance and motor stereotypies (Moy et al., 2008). BALB/c mice also exhibit low sociability, exaggerated aggres-sion, enlarged brain mass, low serotonin levels (Brodkin, 2007) and decreased passive social behavior (Fairless et al., 2013), suggesting them as a genetic model of ASD-related states. Systemic treatment of BALB/c mice with MK-801, an allosteric inhibitor of the gluta-matergic (NMDA) receptors, elicits stereotypic circling behavior (Burket et al., 2010), whereasD-serine, a NMDA receptor agonist,
in-creases sociability (Jacome et al., 2011). Together, this suggests the involvement of glutamatergic pathways in the pathogenesis of locomo-tor stereotypies and social impairment in the BALB/c mouse model of ASD-like behavior. Interestingly, acute administration of oxytocin does not alter social behavior, whereas its sub-chronic administration in-creases sociability 24 h post-treatment in BALC/cByJ mice and for 1–2 weeks in C58/J mice (Teng et al., 2013). Oxytocin receptorOxtr
knockout mice exhibit decreased social ability but maintain normal locomotor activity (Pobbe et al., 2012). Mice lacking theCd38gene (which encodes for CD38, an oxytocin secretion regulator;Bartz and McInnes, 2007) exhibit oxytocin signaling defects, behavioral impair-ments and deficits in social recognition (Higashida et al., 2011), similar to those exhibited by other mouse models with oxytocin deficits (Higashida et al., 2012; Higashida et al., 2011). Moreover, the arginine vasopressin (AVP) system also plays an important role in social behavior, and the V1a receptor (V1aR) gene has been linked to social deficits (Egashira et al., 2007). Subchronic phencyclidine (PCP) in rodents im-pairs social interaction and reduces density of brain V1aR (Egashira et al., 2007), whereas V1aR−/−mice display social deficits, and may
there-fore be a useful model of ASD-like behavior and other disorders associat-ed with social deficits (Egashira et al., 2007).
2.2. Pharmacological rodent models relevant to ASD
In addition to genetic models, rodents are valuable in studying drug-evoked responses relevant to ASD. For example, as social withdrawal is one of the symptoms of schizophrenia and ASD, the social approach test has long been used to study the psychopharmacology of social pheno-types (Hanks et al., 2013).D-Amphetamine disrupts rodent sociability without stimulating locomotor activity (this effect was not reversed by antipsychotics), whereas the gamma-aminobutyric acid (GABA)-A inverse agonist FG-7142 reduces sociability, reversed by the GABA-A antagonist,flumazenil (Hanks, Dlugolenski, 2013).
Antipsychotic drugs antagonize both dopaminergic-induced and glutamatergic N-methyl-D-aspartate (NMDA) antagonist-induced be-haviors (Corbett et al., 1995). A typical glutamatergic antagonist, PCP evokes social withdrawal in rats housed socially without affecting their locomotion (Corbett et al., 1995). SCH 23390, raclopride, haloperidol, chlorpromazine and risperidone fail to reduce PCP-evoked social withdrawal, whereas clozapine and olanzapine signifi -cantly reverse this deficit, demonstrating pharmacological sensitivity of rodent social phenotypes to various drug classes relevant to treating clinical ASD (Corbett et al., 1995). PCP also reduces social behavior in ro-dents following chronic treatment—an effect which lasted for several weeks after discontinuation, and is reversed by clozapine but not haloper-idol (Qiao et al., 2001). Ketamine, another glutamatergic antagonist, also causes similar social deficits as PCP (e.g., disrupting social recognition for several days), which were attenuated by chronic haloperidol or olanzapine (Gao et al., 2009).
While rodent models appear to be a valuable tool to study ASD genetics and pharmacology (Crawley, 2012, Kas et al., 2013), there is a growing need for alternative, time/cost-efficient and high-throughput models of this disorder. Furthermore, domain-oriented approaches to complex human brain disorders focus on various domains (e.g., cogni-tive, affeccogni-tive, motor or social) that represent clusters of various individ-ual behaviors, and overlap during brain pathogenesis (seeKalueff et al., 2008for details). Emphasizing the importance of cross-domain model-ing of CNS pathobiology, these‘integrative’approaches assess multiple disordered phenotypes as a system, and are particularly useful in study-ing diseases such as ASD (which is polygenic and displays a wide range of phenotypes with different severity) (Kalueff et al., 2008). In addition, this allows the researchers to analyze multiple co-expressed behaviors, simultaneously generating rich behavioral data in several distinct neurophenotypic domains (Warnick et al., 2011). Finally, inter-species trait genetics approaches (Kas et al., 2013) recognize the importance of cross-species modeling of complex human brain disorders. Given evolutionarily conserved mechanisms of social behavior, it is therefore
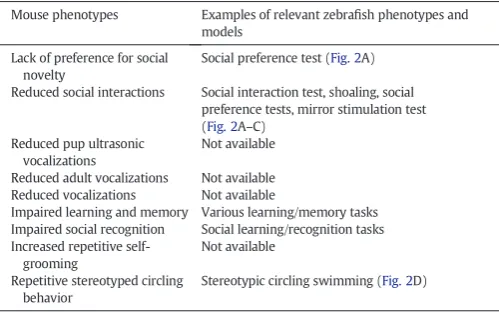
Table 2
Comparison of rodent phenotypes relevant to autism spectrum disorder (ASD; selected fromCrawley, 2012; Kas et al., 2013) and zebrafish behaviors (also seeTable 3–4and
Kalueff et al., 2013for a comprehensive zebrafish behavioral catalog).
Mouse phenotypes Examples of relevant zebrafish phenotypes and models
Lack of preference for social novelty
Social preference test (Fig. 2A)
Reduced social interactions Social interaction test, shoaling, social preference tests, mirror stimulation test (Fig. 2A–C)
Reduced pup ultrasonic vocalizations
Not available
Reduced adult vocalizations Not available Reduced vocalizations Not available
Impaired learning and memory Various learning/memory tasks Impaired social recognition Social learning/recognition tasks Increased repetitive
self-grooming
Not available
Repetitive stereotyped circling behavior
important to foster innovative modeling of ASD, expanding the spec-trum of model organisms for its translational research (Kalueff et al., 2007).
3. Zebrafish models relevant to ASD
3.1. Behavioral and pharmacological models
The zebrafish (Danio rerio) is a new popular model organism in bio-medical research. Thesefish are particularly useful for translational neu-roscience because of their high physiological and genetic homology to humans, ease of genetic manipulation, fully characterized genome, and rapid development (Gerlai, 2010a, 2010b, 2011; Kalueff et al., in press; Lopes et al., 2013; Suen et al., 2013). Zebrafish are also highly so-cial animals, spending the majority of their time in schools (Fig. 2–3) and staying very close to conspecifics. It has recently been suggested that the zebrafish system may contribute to understanding ASD patho-biology and genetics (Tropepe and Sive, 2003). Here, we discuss how zebrafish models can be applied practically for experimental studies of autism.
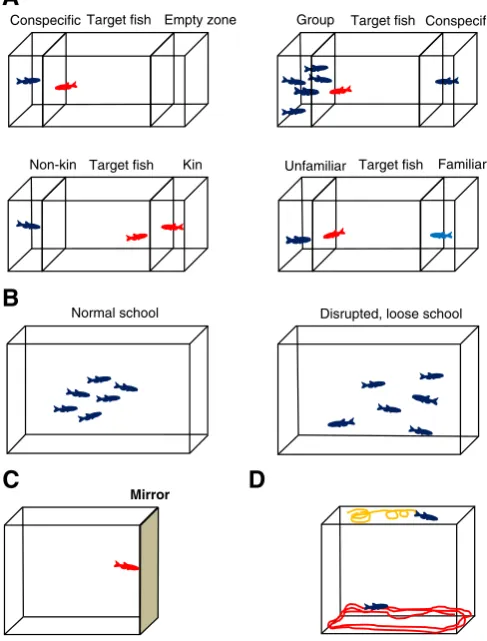
Tables 1–3show that many of the clinical and rodent ASD-like phenotypes can be successfully modeled in zebrafish. For example, sim-ilar to a mouse social interaction protocol (Kas et al., 2013), in the
zebrafish social interaction test two unfamiliar zebrafish introduced together explore each other, demonstrating a wide range of quantifiable social behaviors (Table 4) which can easily be assessed in terms of dura-tion and frequency of various types of social contacts and approach.
In the zebrafish social preference test, also adapted from rodent studies (Kas et al., 2013), a targetfish given a choice between staying close to the empty vs. conspecific zone, spends significantly more time near the conspecific area (Fig. 2A). In other modifications of this model, zebrafish typically spend more time near a group of zebrafish (vs. a singlefish), also showing kin recognition/preference (Table 3) and spending more time during social investigation of novel (unfamil-iar) zebrafish (Fig. 2A). Similarly, zebrafish models can also take advan-tage of the availability of various color variants, assessing the duration and frequency of social contacts with such phenotypically distinct strains offish. For example, while the wild type zebrafish typically avoid shoals of white-skin stripelessnacremutants, their social prefer-ence fornacrefish increases by systemic injections of oxytocin (and its
fish analog isotocin), paralleling its similar pro-social effects in rodents (Capurro, 2010); see discussion further.
In the shoaling test (Fig. 2B,Fig. 3), zebrafish spent most of their time swimming in dynamic groups (schools), characterized by short
inter-fish distance, smaller zebrafish group area size/diameter, as well as rel-ative polarizations. In contrast, disorganized social structure in zebrafish is characterized by reduced polarization offish shoals, looser and larger schools, and higher percentage offish leaving the group and spending time outside the shoal (Fig. 2B and3;Kalueff et al., in press).
The mirror stimulation test, quite specific for aquatic models, is based on mirror presentation and the fact thatfish perceive their own reflection as another zebrafish (Fig. 2C andTable 3; seeCachat et al., 2013for review). This situation can evoke both social behavior and aggression, which will be robustly disrupted if thefish display social deficits. For example, increased head-butting or mirror biting in this test may represent elevated aggression and/or sociability in zebrafish, whereas abnormally low responses can be used to quantify ASD-like symptoms (Cachat et al., 2013).
In addition to social deficits (Fig. 1), behavioral perseverations are also an important ASD-related phenotype (Table 1). Therefore, it may be critical to assess the availability of zebrafish behavioral stereotypies that can be measured in various tests. While zebrafish do not display patterned and complex grooming behavior, they do show several common behavioral stereotypies, including repetitive, stereotypic
‘thigmotaxic’swimming near the walls, or specific circling behavior, involving rotational swimming in tight circles (Fig. 2D,Table 4). Thus, zebrafish models are fully capable of displaying both hallmark behavior-al symptoms of ASD—social deficits and behavioral perseverations, indi-cating high translational potential of zebrafish models for ASD-related states.
Interestingly, zebrafish circling behavior can be induced by selected psychotropic drugs, such as glutamatergic antagonists MK-801, PCP or ketamine (Kyzar et al., 2012a), paralleling rodent drug-evoked circling locomotion already discussed above in relation to ASD. In line with this, various pharmacological agents have also been shown to potently modulate zebrafish social behaviors. For example, zebrafish shoaling is markedly disrupted by various psychoactive drugs (Fig. 3;Maaswinkel et al., 2013), resembling the effects on ASD-related social behavior de-scribed for these agents in rodents (see above). Collectively, this sup-ports the evolutionarily conserved nature of ASD-related social and motor phenotypes in various species, again emphasizing the transla-tional value of zebrafish models to study human ASD.
As mentioned earlier, there is a general lack of clinically efficient ASD-specific drugs, stressing the importance of developing novel com-pounds to treat various symptoms of ASD. For example, risperidone (clinically effective to treat irritability and motor/aggressive behaviors in ASD patients) does not reduce their social deficits and repetitive/ obsessive behaviors (Crawley, 2012). While risperidone was not tested infish social tests, it potently affects zebrafish motor and physiological
Target fish
Conspecific Empty zone Group Target fish Conspecific
Target fish Kin
Non-kin Unfamiliar Target fish Familiar
Normal school Disrupted, loose school
Mirror
C
B
A
D
activity (Prieto et al., 2012); also see similar results for aripiprazole (Lee et al., 2013). Fluoxetine, also commonly used in clinical and rodent ASD studies (Chadman, 2011), albeit not yet tested infish social tests, also strongly modulates zebrafish motor and emotional behavior (Egan et al., 2009). Collectively, this indicates a generally high sensitivity of zebrafish to compounds clinically relevant to correcting some ASD symp-toms, suggesting a potential predictive validity of zebrafish screens to anti-ASD drugs. Clearly, future studies examining in-depth this applica-tion of zebrafish models to ASD research merit further scrutiny.
Cognitive deficits are widely recognized as an important clinical aspect of ASD, and are often observed in rodent models of this disorder (Kas et al., 2013). Zebrafish possess excellent cognitive abili-ties, which can be comprehensively evaluated in various cognitive tasks (Oliveira, 2013; Stewart and Kalueff, 2012). In addition to zebrafish social cognitive models (e.g., social or kin recognition, Fig. 2a,Table 4), various other paradigms can be used to assess non-social cognitive deficits relevant to ASD in this organism. For example,
habituation to novelty (reflecting spatial working memory) can be reli-ably measured in zebrafish, and is affected by various pharmacological agents (Wong et al., 2010a). Memory performance in various mazes can be assessed in zebrafish in a manner similar to rodent studies (Gaikwad et al., 2011; Stewart and Kalueff, 2012). Since attention deficit hyperactivity disorder (ADHD) is frequently comorbid with ASD (Roy et al., 2013; Sprenger et al., 2013), and is related to both behavioral and cognitive clusters of ASD symptoms (Fig. 1), the possibility of modeling ADHD-like phenotypes in zebrafish (Norton, 2013) becomes important and translationally relevant to ASD (also seeParker et al., 2012a,2012b, 2013for details on recent progress in experimental models of attention in zebrafish).
3.2. Physiological correlates
In addition to behavioral biomarkers, ASD is often accompanied clini-cally by altered physiological (e.g., neuroendocrine) functions. For
MDMA 160 mg/L
Control
PCP 5 mg/L
Control
Reserpine 10 mg/L 7 days
Control
Ethanol 4% vol/vol
Control
Fig. 3.Examples of pharmacological modulation of zebrafish social (shoaling) phenotypes. Note that zebrafish shoaling responses are markedly impaired by acute 20-min exposure to phencyclidine (PCP, 5 mg/L), 3,4-methylenedioxymethamphetamine (MDMA, 160 mg/L), ethanol (4% vol/vol) and chronic reserpine (10 mg/L for 7 days) in representative 4-fish shoals (Kalueff et al., 2009–2013 unpublished studies; also seeGreen et al., 2012, vs. drug-free controls). PCP, a dissociative glutamatergic hallucinogen, is well-known to affect social behaviors in humans and rodents (see text for details) and evokes similar effects in zebrafish. A serotonergic psychedelic drug MDMA disrupts social behavior in rodents and zebrafish (Green et al., 2012). Alcohol at mild-to-high doses is commonly known to cause social withdrawal in human and rodents, also showing a similar profile in zebrafish. Reserpine, a plant alkaloid which depletes brain monoamines, is known to impair social behavior in rodents (Bisong et al., 2010, 2011), also causing social deficits in zebrafish in the shoaling test used here. Overall, these data support sensitivity and face validity of zebrafish autism spectrum disorder (ASD)-related social phenotypes to pharmacological modulation by drugs known to evoke ASD-like symp-toms clinically and in experimental (rodent) models. In addition to drugs evoking ASD-like behavior, it is logical to expect that zebrafish models can be used to study agents that modulate zebrafish behavior in the opposite direction, i.e., for screening for novel potential anti-ASD compounds (see discussion of zebrafish sensitivity to risperidone, aripiprazole andfluoxetine in the text).
example, a dysregulated oxytocinergic system has been strongly impli-cated in social deficits common for ASD (Carter, 2007; Insel et al., 1999; Jacob et al., 2007). Therefore, continued investigation into the develop-mental and functional role of oxytocin is critical. The zebrafish homologue
of oxytocin, isotocin, facilitates numerous facets of social behavior in tel-eosts, including social approach, fear, reproduction-related vocalizations and courtship behavior (Braida et al., 2012; Goodson and Bass, 2000; Thompson and Walton, 2004). Recently, peripheral administration of either oxytocin or isotocin (equally potent relative to each other), or an oxytocin antagonist, has been shown to modulate social preference and anxiety-related behavior in zebrafish (Braida et al., 2012), similar to humans (Ebstein et al., 2012; Kumsta and Heinrichs, 2013; Neumann and Landgraf, 2012) and rodents (Carter et al., 2008; Lim et al., 2005). Since the oxytocinergic system is increasingly recognized as a potential therapeutic target in the treatment of ASD (Harony and Wagner, 2010; Modi and Young, 2012; Striepens et al., 2011), zebrafish may provide use-ful tools to investigate the pharmacological profiles of oxytocin-related compounds, as well as to test the potential of novel anti-ASD drugs targeting the oxytocinergic neuroendocrine system.
Another potential physiological correlate of ASD may include altered cortisol levels, often reported in autistic patients. While some species differences may exist in mineralocorticoid vs. glucocorticoid action of this hormone (Pippal et al., 2011), zebrafish offer an excellent model for cortisol screening, as they not only possess a robust neuroendocrine stress axis, but also utilize cortisol (like humans), which can be easily and reliably quantified infish (Egan et al., 2009). Likewise, seizure be-havior is commonly observed in ASD in humans (Tuchman, 2013) and rodents (Gilby, 2008; Greco et al., 2013). Importantly, experimental sei-zures can be easily evoked in zebrafish (Hortopan et al., 2010; Stewart et al., 2012; Wong et al., 2010b) and analyzed (like other ASD-related symptoms,Fig. 1) via high-throughput screening, further supporting the use of zebrafish as an efficient model for ASD and drug discovery.
Table 4
Selected zebrafish behaviors potentially relevant to autism spectrum disorder (ASD; based onKalueff et al., 2013, also seeTable 3for a general framework).
Behaviors Brief description
Aggression Complex behaviors (including approach,fin raise, biting, charging, chasing and circling) directed at conspecifics in adult zebrafish; may appear in the context of establishing dominance (social interaction)
Charge Movement towards a secondfish with increasing acceleration; establishes social dominance and marks the resolution of a zebrafish‘fight’ (Oliveira et al., 2011).
Circling Repetitive swimming in a circular direction; can be defined by their diameter (e.g., 2 body lengths/~5 cm) and quantified by the number of complete circles per trial, the number (%) of animals showing circles and the direction of circling (left- or right-rotations); automated methods may also quantify turn angle and angular velocity (Kyzar et al., 2012a; Riehl et al., 2011)
Display Social behavior used to establish dominance/hierarchy (Spence et al., 2008). Lateral display involves twofish lined up parallel to each other head to tail, raising dorsalfins (fin raise), extending caudalfins and circling. Frontal display involves twofish approaching each other from the front with the attempt of biting (Schneider, 2011)
Fighting Agonistic confrontation between two individuals to establish social dominance (Oliveira et al., 2011). During thefirst phase, thefish assess each other by exhibiting display, biting/nipping, which continues until thefirst chase/flee occurs. Next, the‘winner’(chaser) initiates all agonistic behaviors, while the‘loser’displaysfleeing, submission behavior or freezing (Oliveira et al., 2011)
Habituation Tendency to show a decreased response upon repeated exposure to a novel stimulus/environment (reflecting cognitive ability of zebrafish). Includes inter-trial (inter-session) and
intra-trial (intra-session) habituation, quantified by calculating the ratios of behavioral activity during the initial vs. latest trials, or by assessing the behavioral profile offish across the trial(s) (Stewart et al., 2010; Stewart et al., 2013)
Kin preference and recognition
The preference for kin vs. unrelated zebrafish, particularly robust in juvenile zebrafish. Zebrafish are able to seek/recognize kin from unrelated zebrafish; this behavior involves approach/attraction and leads to increased time spent near kin (kin preference) (Spence et al., 2008) Mirror stimulation response Complex behaviors evoked infish by mirror exposure; most likely linked to aggression and social investigation; typically includes approach,
head-butting, biting the mirror and chasing own reflection (Cachat et al., 2013; Oliveira et al., 2011)
Place preference The tendency to establish a preferred location in which thefish spends more time; can be induced by social reward (e.g., sight of the conspecific) (Al-Imari and Gerlai, 2008)
Polarization Behavioral characteristic of adult zebrafish reflecting the degree to which members of the group are moving in the same direction; is high in established zebrafish groups (Miller and Gerlai, 2012b)
Shoaling Formation of a group (shoal) of adult zebrafish, held together by social pressures (Miller and Gerlai, 2011; Miller and Gerlai, 2012a,2012b). Zebrafish shoaling can be quantified manually or using automated video-tracking systems, assessing several endpoints, including the average inter-fish distance; shoal area size; proximity (time each member of the shoal spent within a specified distance from each other); nearest and farthest neighbor distances; time spent in shoal; time spent away from shoal; number of animals leaving the shoal and polarization (heading) Social interaction Normal social behavior of zebrafish, a reciprocal change in zebrafish behavior influenced by the presence or actions of other conspecifics
(Al-Imari and Gerlai, 2008; Miller and Gerlai, 2011; Oliveira et al., 2011). Some examples includefighting/aggression, shoaling, courtship, approach/boldness (social investigation), social recognition and social preference
Social preference and recognition
A natural tendency to spend time close to conspecifics; can be observed as part of shoaling behavior, kin recognition, social recognition, or preference of the‘conspecific’vs.‘empty’zones (Al-Imari and Gerlai, 2008; Spence et al., 2008). Zebrafish also recognize familiar from unfamiliar zebrafish (social recognition) and distinguish between own vs. phenotypically different other strain (Spence et al., 2008) Stereotypic locomotion A pattern of rigid, repetitive behaviors (e.g., swimming from corner to corner, stereotypic jaw movements) evoked in zebrafish under some
conditions (e.g., treatment with psychostimulants and PCP;Cachat et al., 2013; Kyzar et al., 2012a)
Submissive behavior A social behavior following aggressive confrontations. Submissivefish stays immobile (withfins retracted), typically near the bottom or near the surface, with the caudal part of the body oriented downward (Oliveira et al., 2011)
Table 3
Overview of zebrafish phenotypes relevant to autism spectrum disorder (ASD).
Clinical symptomsa Relevant zebrafish phenotypesb
Social deficits Reduced social interaction in the shoaling test (e.g., less time spent in shoals, increased shoal size, increased average inter-fish distance and farthest neighbor distance). Reduced social interaction and/or investigation time and frequency in the shoaling, mirror stimulation and the social preference tests (e.g., reduced time spent near conspecific, reduced time investigating new (vs. familiar) conspecific, reduced time and contacts with mirror). Repetitive behaviors and
cognitive inflexibility
Presence of specific behavioral stereotypies (e.g., cycling behavior,figure 8 swimming, stereotyped thigmotaxis-like swimming and other motor stereotypies, such as repetitive jaw movements) and/or cognitive deficits (e.g., detected in various motor and cognitive tasks, such as impaired habituation in the novelty tests or aberrant patterns of alternation in Y or/T-mazes)
Developmental nature of observed symptoms
Aberrant developmental trajectories of zebrafish social behaviors (e.g., in shoaling test across lifespan)
aBased on DSM-5; seeTable 1for details.
3.3. Genetic models relevant to ASD
Considering the high heritability of ASD and the ease of genetic manipulations in zebrafish, this species is also emerging as a useful tool to study ASD genetics. For example, the ASD-linked human 16p11.2 chromosomal area has a homologous region in zebrafish, consisting of several genes important for brain development (Blaker-Lee et al., 2012), including KCTD13, which is responsible for head size and neurogenesis in zebrafish, and was identified as a major contributor in some cases of autism in humans (Golzio et al., 2012). Similarly, the ASD susceptibility genemetregulates zebrafish cerebellar development and motor neuron migration.Metencodes a tyrosine kinase receptor, im-plicated in autism and brain development in mice and zebrafish (Elsen et al., 2009). Zebrafishmetis expressed in the cerebellar primordium (later localizing in the ventricular zone), and its morpholino knockdown reduces the size of the cerebellum and affects its cellular morphology, thereby paralleling the correlation between altered MET regulation and ASD (Elsen et al., 2009). Reflecting a neurodevelopmental aspect of ASD pathogenesis (Table 1), this phenotype illustrates the potential of zebrafish in modeling ASD (Blaker-Lee et al., 2012; also see Norton, 2013for a review).
The AUTS2 locus has also been strongly implicated in ASD and other human brain disorders, such as ADHD, epilepsy, dyslexia, motor delay and language delay (Oksenberg et al., 2013). The knock-down ofauts2
in zebrafish results in a smaller head size, neuronal reduction and de-creased mobility. Collectively, this suggests that AUTS2 plays an impor-tant role in neurodevelopment, consistent with both clinical and zebrafish phenotypes relevant to ASD (Oksenberg et al., 2013). Like-wise, mounting evidence has recently implicated neurexins and neuroligins in neurodevelopmental disorders, including ASD and
psychoses (Wright and Washbourne, 2011). These genes have been extensively characterized in zebrafish, demonstrating high sequence conservation with the human genes, therefore suggesting that zebrafish models (with their simpler circuitry and high-throughput capacity) may prove extremely useful in identifying therapeutic strategies to treat ASD (Wright and Washbourne, 2011).
The potential role ofSHANK3in human ASD (Boccuto et al., 2013) has recently been paralleled by zebrafish models, where the genetic knock-down of zebrafish zshank3orthologous genes (zs3.1and zs3.2) by morpholino results in a reduction in the head size and markedly impaired swim responses to touch (Gauthier et al., 2010). The fact that this gene is strongly implicated in CNS function in zebrafish, rodents and humans (Gauthier et al., 2010) strongly supports the value of zebrafish models to study CNS pathogenesis, including ASD.
In addition to genetically modifiedfish, recent studies on strain differences in shoaling have revealed important insights into the genetics of zebrafish social behavior. For example, based on strong differences between a wild-derived strain offish and the laboratory AB strain, quantitative trait loci (QTL) analysis identified a genomic region on chromosome 21 responsible for shoaling behavior (Wright et al., 2006;also seeWright et al., 2003). Overall, these results not only confirm the potential of zebrafish for genetic mapping of complex be-havioral traits, but also establish genetic determinants of social behavior in this species (Wright et al., 2003, 2006), generally consistent with the notion of high heritability of ASD.
3.4. Environmental models potentially relevant to ASD
In both humans and rodents, environmental factors play a signifi -cant role in the development of ASD (Kawicka and Regulska-Ilow, 2013; LaSalle, 2013; Thirtamara Rajamani et al., 2013). Recent studies suggest that ASD-like state may also be modulated by environmental stimuli in zebrafish. For example, it has been suggested that zebrafish also have gene-by-environment epigenetic regulation of complex be-haviors, such as ASD-like phenotypes (Norton, 2013). Consistent with this, exposure to a pesticide chlorpyrifos induces ASD-like behaviors in zebrafish (Richendrfer et al., 2012). Thus, while the environmental modulation of ASD-like behavior in zebrafish has not yet been thoroughly investigated, future studies of environmental effects on zebrafish phenotypes relevant to ASD may be important. Given the well-recognized history of zebrafish tests in developmental pharmacol-ogy and toxicolpharmacol-ogy, the possibility of developing ‘environmental’ zebrafish models of ASD-like pathogenesis can represent an interesting direction for future research in thisfield.
4. Conclusion
In summary, zebrafish emerge as an important model species in translational neuroscience and the neurogenetics of ASD. The presence of key behavioral phenotypes which parallel human and rodent ASD, as well as their sensitivity to major groups of pharmacological agents used clinically to treat (or evoke) ASD, also supports their role in experimen-tal modeling of autism. The availability of recently developed video-tracking tools enables efficient, objective and data-dense quantification of zebrafish social phenotypes (Green et al., 2012; Maaswinkel et al., 2013). A highly significant correlation for manual vs. automated analy-ses in such studies demonstrates the capacity of video-tracking technol-ogy to assess zebrafish social behaviors in a high-throughput and reliable manner, further fostering zebrafish models relevant to ASD (Green et al., 2012).
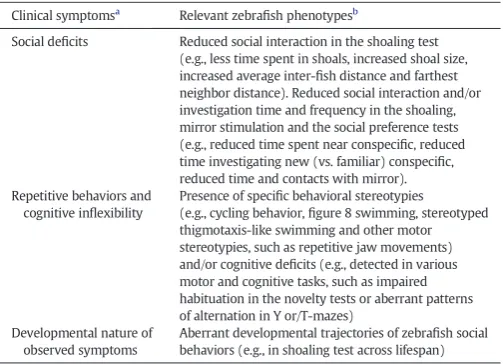
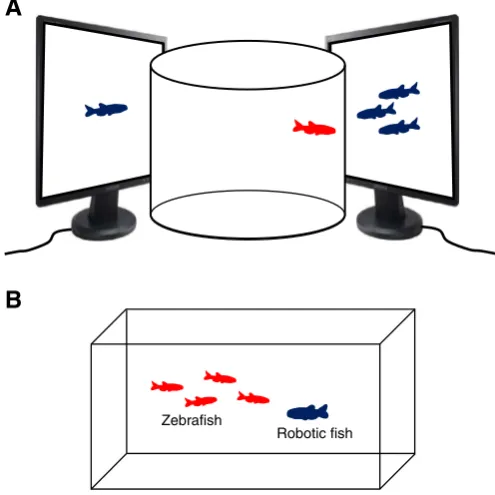
Notably, zebrafish behavior is strongly reliant on visual sensory infor-mation about their social environment, which offers a possibility of using standardized still images and‘virtual reality’videos of individual or group zebrafish rather than utilizing livefish as social stimuli (e.g.,Gerlai et al., 2009; Luca and Gerlai, 2012; Pather and Gerlai, 2009). Unlike rodent ASD models (which typically require presentation of live animals), zebrafish
Robotic fish Zebrafish
A
B
Fig. 4.Additional models for zebrafish social behaviors. A: An experimental setup utilizing standardized automated (vs. live) stimulusfish image presentation in zebrafish sociability models relevant to ASD. These approaches enable a better, standardized control of exper-imental conditions across all groups and eliminate potential confounding effects on the testedfish (e.g., resulting from aberrant stimulusfish behavior). In addition, they also en-able testing multiple additional experimental parameters (which can be easily adjusted using computers), such as size, shape, number of subjects, color, skin pattern, body orien-tation/angle, position in the tank, as well as swimming speed and trajectory of the present-ed stimulusfish. B: The potential application of roboticfish (e.g.,Polverino et al., 2012) for modeling zebrafish social behaviors, such as shoaling, relevant to ASD-like behavior (note that the same approaches can also be applied to zebrafish social preference tests shown in
Fig. 2).
social models based on automated stimulus presentation (Fig. 4A) can therefore be better standardized and controlled experimentally, leading to improved reproducibility and reduced variance of data. Moreover, recent evidence suggests the utility of automated roboticfish (Fig. 4B) to study social and other behavioral responses in zebrafish (Abaid et al., 2012; Polverino et al., 2012). For example, the ability of zebrafish to display robust social behaviors (e.g., shoaling or preference) when exposed to a robotic zebrafish provides an excellent opportunity to studyfish social behavior while fully controlling the stimulus presenta-tion by manipulating the robotic zebrafish‘behavior’(Fig. 4B).
Clearly, each experimental animal model of complex brain disorders has various limitations (Kalueff et al., 2007). For a complex multi-faceted brain disorder like ASD, it is impossible to develop an animal model that recapitulates all aspects of its pathogenesis. However, animal models are a valuable tool to dissect various aspects of brain pathobiology, explore the role of specific neural pathways or discover genetic determinants. Thus, the question of which models or model organisms are‘better’is misleading, since the answer will only be limited to specific research questions, phenotypes in question or putative mechanisms to study. Therefore, complementing the existing rodent models relevant to ASD (Crawley, 2012), zebrafish can be used complementarily, to advance our understanding of ASD and related behavioral deficits.
Moreover, ASD is a neurodevelopmental disorder (Kas et al., 2013), and this aspect is critical for experimental animal models of ASD-like states (Tropepe and Sive, 2003). It was already mentioned that zebrafish have a rapid development, which enables time- and cost-efficient modeling of this important pathogenetic aspect of ASD (note, however, that social behavior is not seen in larvalfish, and therefore larval models and screens may not be useful to study social phenotypes in zebrafish). Finally, the ease of genetic and pharmacological manipulations, as well as the low cost, high-throughput and potential for targeting develop-mental trajectories of ASD, make zebrafish a promising new model (Green et al., 2012; Kalueff et al., in press; Tropepe and Sive, 2003) to complement traditional rodent tests (El-Kordi et al., 2012; Kas et al., 2013). As noted recently, thefield of experimental modeling of ASD has taken thefirst step in a long and challenging journey (Crawley, 2012). So far, the mounting experimental evidence presented here indicates that zebrafish models relevant to ASD will provide an exciting new chap-ter for this endeavor.
Acknowledgements
The authors' research has been supported by the International Zebrafish Neuroscience Research Consortium (ZNRC) and the ZENEREI Institute.
References
Abaid N, Bartolini T, Macri S, Porfiri M.Zebrafish responds differentially to a roboticfish of varying aspect ratio, tail beat frequency, noise, and color. Behav Brain Res 2012;233: 545–53.
Al-Imari L, Gerlai R.Sight of conspecifics as reward in associative learning in zebrafish (Danio rerio). Behav Brain Res 2008;189:216–9.
Bartz JA, McInnes LA.CD38 regulates oxytocin secretion and complex social behavior. BioEssays News Rev Mol Cell and Dev Biol 2007;29:837–41.
Benvenuto A, Battan B, Porfirio MC, Curatolo P.Pharmacotherapy of autism spectrum dis-orders. Brain Dev 2013;35:119–27.
Bernardet M, Crusio WE.Fmr1 KO mice as a possible model of autistic features. Sci World J 2006;6:1164–76.
Bisong S, Brown R, Osim E.Comparative effects ofRauwolfia vomitoriaand chlorproma-zine on social behaviour and pain. N Am J Med Sci 2011;3:48–54.
Bisong SA, Brown R, Osim EE.Comparative effects ofRauwolfia vomitoriaand chlorprom-azine on locomotor behaviour and anxiety in mice. J Ethnopharmacol 2010;132: 334–9.
Blaker-Lee A, Gupta S, McCammon JM, De Rienzo G, Sive H.Zebrafish homologs of genes within 16p11.2, a genomic region associated with brain disorders, are active during brain development, and include two deletion dosage sensor genes. Dis Model Mech 2012;5:834–51.
Boccuto L, Lauri M, Sarasua SM, Skinner CD, Buccella D, Dwivedi A, et al.Prevalence of SHANK3 variants in patients with different subtypes of autism spectrum disorders. Eur J Human Genet EJHG 2013;21:310–6.
Bozdagi O, Sakurai T, Papapetrou D, Wang X, Dickstein DL, Takahashi N, et al.
Haploinsufficiency of the autism-associated Shank3 gene leads to deficits in synaptic function, social interaction, and social communication. Mol Autism 2010;1:15.
Braida D, Donzelli A, Martucci R, Capurro V, Busnelli M, Chini B, et al.Neurohypophyseal hormones manipulation modulate social and anxiety-related behavior in zebrafish. Psychopharmacology (Berl) 2012;220:319–30.
Brodkin ES.BALB/c mice: low sociability and other phenotypes that may be relevant to autism. Behav Brain Res 2007;176:53–65.
Burket JA, Cannon WR, Jacome LF, Deutsch SI.MK-801, a noncompetitive NMDA receptor antagonist, elicits circling behavior in the genetically inbred Balb/c mouse strain. Brain Res Bull 2010;83:337–9.
Cachat J, Kyzar EJ, Collins C, Gaikwad S, Green J, Roth A, et al.Unique and potent effects of acute ibogaine on zebrafish: the developing utility of novel aquatic models for hallu-cinogenic drug research. Behav Brain Res 2013;236:258–69.
Capurro V.New pharmacological tools for autism research: oxytocin receptor mutant mice and zebrafish as neurobehavioral models [Doctoral dissertation]. Milan: Univer-sity of Milan; 2010.
Carneiro AM, Cook EH, Murphy DL, Blakely RD. Interactions between integrin alphaIIbbeta3 and the serotonin transporter regulate serotonin transport and platelet aggregation in mice and humans. J Clin Invest 2008;118:1544–52.
Carter CS.Sex differences in oxytocin and vasopressin: implications for autism spectrum disorders? Behav Brain Res 2007;176:170–86.
Carter CS, Grippo AJ, Pournajafi-Nazarloo H, Ruscio MG, Porges SW.Oxytocin, vasopressin and sociality. Prog Brain Res 2008;170:331–6.
Carter MD, Shah CR, Muller CL, Crawley JN, Carneiro AM, Veenstra-VanderWeele J. Ab-sence of preference for social novelty and increased grooming in integrin beta3 knockout mice: initial studies and future directions. Autism Res Off J Int Soc Autism Res 2011;4:57–67.
Chadman KK.Fluoxetine but not risperidone increases sociability in the BTBR mouse model of autism. Pharmacol Biochem Behav 2011;97:586–94.
Corbett R, Camacho F, Woods AT, Kerman LL, Fishkin RJ, Brooks K, et al.Antipsychotic agents antagonize non-competitive N-methyl-D-aspartate antagonist-induced be-haviors. Psychopharmacology 1995;120:67–74.
Crawley JN.Translational animal models of autism and neurodevelopmental disorders. Dialogues Clin Neurosci 2012;14:293.
Ebstein RP, Knafo A, Mankuta D, Chew SH, Lai PS.The contributions of oxytocin and vaso-pressin pathway genes to human behavior. Horm Behav 2012;61:359–79.
Edvardson S, Ashikov A, Jalas C, Sturiale L, Shaag A, Fedick A, et al.Mutations in SLC35A3 cause autism spectrum disorder, epilepsy and arthrogryposis. J Med Genet 2013;50(11):733–9.
Egan RJ, Bergner CL, Hart PC, Cachat JM, Canavello PR, Elegante MF, et al.Understanding behavioral and physiological phenotypes of stress and anxiety in zebrafish. Behav Brain Res 2009;205:38–44.
Egashira N, Tanoue A, Matsuda T, Koushi E, Harada S, Takano Y, et al.Impaired social in-teraction and reduced anxiety-related behavior in vasopressin V1a receptor knockout mice. Behav Brain Res 2007;178:123–7.
El-Kordi A, Winkler D, Hammerschmidt K, Kastner A, Krueger D, Ronnenberg A, et al. De-velopment of an autism severity score for mice using Nlgn4 null mutants as a construct-valid model of heritable monogenic autism. Behav Brain Res 2012;251: 41–9.
Elsen GE, Choi LY, Prince VE, Ho RK.The autism susceptibility gene met regulates zebrafish cerebellar development and facial motor neuron migration. Dev Biol 2009;335:78–92.
Evans B.How autism became autism: the radical transformation of a central concept of child development in Britain. Hist Hum Sci 2013;26:3–31.
Fairless AH, Katz JM, Vijayvargiya N, Dow HC, Kreibich AS, Berrettini WH, et al. Develop-ment of home cage social behaviors in BALB/cJ vs. C57BL/6J mice. Behav Brain Res 2013;237:338–47.
Farmer C, Thurm A, Grant P.Pharmacotherapy for the core symptoms in autistic disorder: current status of the research. Drugs 2013;73:303–14.
Gaikwad S, Stewart A, Hart P, Wong K, Piet V, Cachat J, et al.Acute stress disrupts perfor-mance of zebrafish in the cued and spatial memory tests: the utility offish models to study stress–memory interplay. Behav Proc 2011;87:224–30.
Gao XM, Elmer GI, Adams-Huet B, Tamminga CA.Social memory in mice: disruption with an NMDA antagonist and attenuation with antipsychotic drugs. Pharmacol Biochem Behav 2009;92:236–42.
Gauthier J, Champagne N, Lafreniere RG, Xiong L, Spiegelman D, Brustein E, et al.De novo mutations in the gene encoding the synaptic scaffolding protein SHANK3 in patients ascertained for schizophrenia. Proc Natl Acad Sci U S A 2010;107:7863–8.
Gerlai R.High-throughput behavioral screens: thefirst step towardsfinding genes in-volved in vertebrate brain function using zebrafish. Molecules 2010a;15:2609–22.
Gerlai R.Zebrafish antipredatory responses: a future for translational research? Behav Brain Res 2010b;207:223–31.
Gerlai R.A smallfish with a big future: zebrafish in behavioral neuroscience. Rev Neurosci 2011;22:3–4.
Gerlai R, Fernandes Y, Pereira T.Zebrafish (Danio rerio) responds to the animated image of a predator: towards the development of an automated aversive task. Behav Brain Res 2009;201:318–24.
Geschwind DH.Autism: many genes, common pathways? Cell 2008;135:391–5.
Gilby KL.A new rat model for vulnerability to epilepsy and autism spectrum disorders. Epilepsia 2008;49(Suppl. 8):108–10.
Golzio C, Willer J, Talkowski ME, Oh EC, Taniguchi Y, Jacquemont S, et al.KCTD13 is a major driver of mirrored neuroanatomical phenotypes of the 16p11.2 copy number variant. Nature 2012;485:363–7.
Greco B, Manago F, Tucci V, Kao HT, Valtorta F, Benfenati F.Autism-related behavioral ab-normalities in synapsin knockout mice. Behav Brain Res 2013;251:65–74.
Green J, Collins C, Kyzar EJ, Pham M, Roth A, Gaikwad S, et al.Automated high-throughput neurophenotyping of zebrafish social behavior. J Neurosci Methods 2012;210: 266–71.
Hanks AN, Dlugolenski K, Hughes ZA, Seymour PA, Majchrzak MJ.Pharmacological dis-ruption of mouse social approach behavior: relevance to negative symptoms of schizophrenia. Behav Brain Res 2013;252:405–14.
Harony H, Wagner S.The contribution of oxytocin and vasopressin to mammalian social behavior: potential role in autism spectrum disorder. Neurosignals 2010;18:82–97.
Higashida H, Yokoyama S, Kikuchi M, Munesue T.CD38 and its role in oxytocin secretion and social behavior. Horm Behav 2012;61:351–8.
Higashida H, Yokoyama S, Munesue T, Kikuchi M, Minabe Y, Lopatina O.CD38 gene knockout juvenile mice: a model of oxytocin signal defects in autism. Biol Pharm Bull 2011;34:1369–72.
Hortopan GA, Dinday MT, Baraban SC.Spontaneous seizures and altered gene expression in GABA signaling pathways in a mind bomb mutant zebrafish. J Neurosci Off J Soc Neurosci 2010;30:13718–28.
Insel TR, O'Brien DJ, Leckman JF.Oxytocin, vasopressin, and autism: is there a connection? Biol Psychiatry 1999;45:145–57.
Jacob S, Brune CW, Carter CS, Leventhal BL, Lord C, Cook Jr EH.Association of the oxytocin receptor gene (OXTR) in Caucasian children and adolescents with autism. Neurosci Lett 2007;417:6–9.
Jacome LF, Burket JA, Herndon AL, Cannon WR, Deutsch SI.D-Serine improves dimensions of the sociability deficit of the genetically-inbred Balb/c mouse strain. Brain Res Bull 2011;84:12–6.
Jamain S, Radyushkin K, Hammerschmidt K, Granon S, Boretius S, Varoqueaux F, et al. Re-duced social interaction and ultrasonic communication in a mouse model of mono-genic heritable autism. Proc Natl Acad Sci 2008;105:1710–5.
Kalueff AV, Gebhardt M, Stewart AM, Cachat JM, Brimmer M, Chawla JS, et al.Towards a comprehensive catalog of zebrafish behavior 1.0 and beyond. Zebrafish 2013;10: 70–86.
Kalueff AV, Olivier JD, Nonkes LJ, Homberg JR.Conserved role for the serotonin transport-er gene in rat and mouse neurobehavioral endophenotypes. Neurosci Biobehav Rev 2010;34:373–86.
Kalueff AV, Ren-Patterson RF, LaPorte JL, Murphy DL.Domain interplay concept in animal models of neuropsychiatric disorders: a new strategy for high-throughput neurophenotyping research. Behav Brain Res 2008;188:243–9.
Kalueff AV, Wheaton M, Murphy DL.What's wrong with my mouse model? Advances and strategies in animal modeling of anxiety and depression. Behav Brain Res 2007;179: 1–18.
Kalueff AV, Stewart AM.Gerlai R. Trends Pharmacol Sci: Zebrafish as an emerging model for studying complex brain disorders; 2014 [in press].
Kas MJ, Glennon JC, Buitelaar J, Ey E, Biemans B, Crawley J, et al.Assessing behavioural and cognitive domains of autism spectrum disorders in rodents: current status and future perspectives. Psychopharmacology 2013. [in press].
Kawicka A, Regulska-Ilow B.How nutritional status, diet and dietary supplements can af-fect autism. A review. Rocz. Panstw. Zakl. Hig. 2013;64:1–12.
Kéïta L, Mottron L, Dawson M, Bertone A.Atypical lateral connectivity: a neural basis for altered visuospatial processing in autism. Biol Psychiatry 2011;70:806–11.
Kesli R, Gokcen C, Bulug U, Terzi Y.Investigation of the relation between anaerobic bacte-ria genus clostridium and late-onset autism etiology in children. J Immunoassay Immunochem 2014;35:101–9.
Kujala T, Lepistö T, Näätänen R.The neural basis of aberrant speech and audition in autism spectrum disorders. Neurosci Biobehav Rev 2013;37:697–704.
Kumsta R, Heinrichs M.Oxytocin, stress and social behavior: neurogenetics of the human oxytocin system. Curr Opin Neurobiol 2013;23:11–6.
Kyzar EJ, Collins C, Gaikwad S, Green J, Roth A, Monnig L, et al.Effects of hallucinogenic agents mescaline and phencyclidine on zebrafish behavior and physiology. Prog Neuropsychopharmacol Biol Psychiatry 2012a;37:194–202.
Kyzar EJ, Pham M, Roth A, Cachat J, Green J, Gaikwad S, et al.Alterations in grooming ac-tivity and syntax in heterozygous SERT and BDNF knockout mice: the utility of behavior-recognition tools to characterize mutant mouse phenotypes. Brain Res Bull 2012b;89:168–76.
LaSalle JM.Epigenomic strategies at the interface of genetic and environmental risk fac-tors for autism. J Hum Genet 2013;58:396–401.
Lee SH, Kim HR, Han RX, Oqani RK, Jin DI.Cardiovascular risk assessment of atypical an-tipsychotic drugs in a zebrafish model. J Appl Toxicol JAT 2013;33:466–70.
Lim MM, Bielsky IF, Young LJ.Neuropeptides and the social brain: potential rodent models of autism. Int J Dev Neurosci 2005;23:235–43.
Lopes da Fonseca T, Correia A, Hasselaar W, van der Linde HC, Willemsen R, Outeiro TF.
The zebrafish homologue of Parkinson's disease ATP13A2 is essential for embryonic survival. Brain Res Bull 2013;90:118–26.
Luca RM, Gerlai R.Animated bird silhouette above the tank: acute alcohol diminishes fear responses in zebrafish. Behav Brain Res 2012;229:194–201.
Maaswinkel H, Zhu L, Weng W.Assessing social engagement in heterogeneous groups of zebrafish: a new paradigm for autism-like behavioral responses. PloS One 2013;8: e75955.
Matson JL, Kozlowski AM, Matson MM.Speech deficits in persons with autism: etiology and symptom presentation. Res Autism Spectr Disord 2012;6:573–7.
Mayes SD, Calhoun SL, Murray MJ, Ahuja M, Smith LA.Anxiety, depression, and irritability in children with autism relative to other neuropsychiatric disorders and typical de-velopment. Res Autism Spectr Disord 2011;5:474–85.
McFarlane HG, Kusek GK, Yang M, Phoenix JL, Bolivar VJ, Crawley JN.Autism-like behav-ioral phenotypes in BTBR T+tf/J mice. Genes Brain Behav 2008;7:152–63.
McTighe SM, Neal SJ, Lin Q, Hughes ZA, Smith DG.The BTBR mouse model of autism spec-trum disorders has learning and attentional impairments and alterations in acetyl-choline and kynurenic acid in prefrontal cortex. PloS One 2013;8:e62189.
Meyza KZ, Defensor EB, Jensen AL, Corley MJ, Pearson BL, Pobbe RL, et al.The BTBR T(+) tf/J mouse model for autism spectrum disorders—in search of biomarkers. Behav Brain Res 2012;251:25–34.
Miller N, Gerlai R.Automated tracking of zebrafish shoals and the analysis of shoaling be-havior. T Zebrafish Protoc Neurobehav Res 2012a:217–30.
Miller N, Gerlai R.From schooling to shoaling: patterns of collective motion in zebrafish (Danio rerio). PLoS One 2012b;7:e48865.
Miller NY, Gerlai R.Shoaling in zebrafish: what we don't know. Rev Neurosci 2011;22: 17–25.
Modi ME, Young LJ.The oxytocin system in drug discovery for autism: animal models and novel therapeutic strategies. Horm Behav 2012;61:340–50.
Moy SS, Nadler JJ, Young NB, Nonneman RJ, Segall SK, Andrade GM, et al.Social approach and repetitive behavior in eleven inbred mouse strains. Behav Brain Res 2008;191: 118–29.
Myers SM.The status of pharmacotherapy for autism spectrum disorders. Expert Opin Pharmacother 2007;8:1579–603.
Neumann ID, Landgraf R.Balance of brain oxytocin and vasopressin: implications for anx-iety, depression, and social behaviors. Trends Neurosci 2012;35:649–59.
Norton WH.Toward developmental models of psychiatric disorders in zebrafish. Front Neural Circ 2013;7:79.
Oksenberg N, Stevison L, Wall JD, Ahituv N.Function and regulation of AUTS2, a gene im-plicated in autism and human evolution. PLoS Genet 2013;9:e1003221.
Oliveira RF.Mind thefish: zebrafish as a model in cognitive social neuroscience. Front Neural Circ 2013;7:131.
Oliveira RF, Silva JF, Simoes JM.Fighting zebrafish: characterization of aggressive behavior and winner–loser effects. Zebrafish 2011;8:73–81.
Pampanos A, Volaki K, Kanavakis E, Papandreou O, Youroukos S, Thomaidis L, et al.A sub-stitution involving the NLGN4 gene associated with autistic behavior in the Greek population. Genet Test Mol Biomark 2009;13:611–5.
Parker MO, Gaviria J, Haigh A, Millington ME, Brown VJ, Combe FJ, et al.Discrimination re-versal and attentional sets in zebrafish (Danio rerio). Behav Brain Res 2012a;232: 264–8.
Parker MO, Ife D, Ma J, Pancholi M, Smeraldi F, Straw C, et al.Development and automa-tion of a test of impulse control in zebrafish. Front Syst Neurosci 2013;7:65.
Parker MO, Millington ME, Combe FJ, Brennan CH.Development and implementation of a three-choice serial reaction time task for zebrafish (Danio rerio). Behav Brain Res 2012b;227:73–80.
Pather S, Gerlai R.Shuttle box learning in zebrafish (Danio rerio). Behav Brain Res 2009;196:323–7.
Pietropaolo S, Guilleminot A, Martin B, D'Amato FR, Crusio WE.Genetic-background mod-ulation of core and variable autistic-like symptoms in Fmr1 knock-out mice. PloS One 2011;6:e17073.
Pippal JB, Cheung CM, Yao YZ, Brennan FE, Fuller PJ.Characterization of the zebrafish (Danio rerio) mineralocorticoid receptor. Mol Cell Endocrinol 2011;332:58–66.
Pobbe RLH, Defensor EB, Pearson BL, Bolivar VJ, Blanchard DC, Blanchard RJ.General and social anxiety in the BTBR T+tf/J mouse strain. Behav Brain Res 2011;216:446–51.
Pobbe RLH, Pearson BL, Blanchard DC, Blanchard RJ.Oxytocin receptor and Mecp2308/Y knockout mice exhibit altered expression of autism-related social behaviors. Physiol Behav 2012;107:641–8.
Polverino G, Abaid N, Kopman V, Macri S, Porfiri M.Zebrafish response to roboticfish: preference experiments on isolated individuals and small shoals. Bioinspiration Bio-mimetics 2012;7:036019.
Prieto M, Gutierrez H, Arevalo R, Chiaramoni N, Alonso SdV.Effect of risperidone andfl u-oxetine on the movement and neurochemical changes of zebrafish. Open J Med Chem 2012;2:129–38.
Qiao H, Noda Y, Kamei H, Nagai T, Furukawa H, Miura H, et al.Clozapine, but not haloper-idol, reverses social behavior deficit in mice during withdrawal from chronic phency-clidine treatment. Neuroreport 2001;12:11–5.
Reynolds S, Urruela M, Devine DP.Effects of environmental enrichment on repetitive be-haviors in the BTBR T+tf/J mouse model of autism. Autism Res Off J Int Soc Autism Res 2013;6(5):337–43.
Richendrfer H, Pelkowski SD, Colwill RM, Creton R.Developmental sub-chronic exposure to chlorpyrifos reduces anxiety-related behavior in zebrafish larvae. Neurotoxicol Teratol 2012;34:458–65.
Riehl R, Kyzar E, Allain A, Green J, Hook M, Monnig L, et al.Behavioral and physiological effects of acute ketamine exposure in adult zebrafish. Neurotoxicol Teratol 2011;33:658–67.
Roy M, Ohlmeier MD, Osterhagen L, Prox-Vagedes V, Dillo W.Asperger syndrome: a fre-quent comorbidity infirst diagnosed adult ADHD patients? Psychiatr Danub 2013;25: 133–41.
Ryan BC, Young NB, Crawley JN, Bodfish JW, Moy SS.Social deficits, stereotypy and early emergence of repetitive behavior in the C58/J inbred mouse strain. Behav Brain Res 2010;208:178–88.
Scattoni ML, Martire A, Cartocci G, Ferrante A, Ricceri L.Reduced social interaction, behav-iouralflexibility and BDNF signalling in the BTBR T+tf/J strain, a mouse model of au-tism. Behav Brain Res 2012;251:35–40.
Schneider H.Measuring agonistic behavior in zebrafish. Neuromethods 2011;51:125–34.
Schwartzer JJ, Careaga M, Onore CE, Rushakoff JA, Berman RF, Ashwood P.Maternal im-mune activation and strain specific interactions in the development of autism-like behaviors in mice. Transl Psychiatry 2013;3:e240.
Silverman JL, Oliver CF, Karras MN, Gastrell PT, Crawley JN.AMPAKINE enhancement of social interaction in the BTBR mouse model of autism. Neuropharmacology 2013;64:268–82.
Silverman JL, Smith DG, Rizzo SJ, Karras MN, Turner SM, Tolu SS, et al.Negative allosteric modulation of the mGluR5 receptor reduces repetitive behaviors and rescues social deficits in mouse models of autism. Sci Transl Med 2012;4:131ra51.
Spence R, Gerlach G, Lawrence C, Smith C.The behaviour and ecology of the zebrafish,
Danio rerio. Biol Rev 2008;83:13–34.
Sprenger L, Buhler E, Poustka L, Bach C, Heinzel-Gutenbrunner M, Kamp-Becker I, et al.
Impact of ADHD symptoms on autism spectrum disorder symptom severity. Res Dev Disabil 2013;34:3545–52.
Stewart A, Kadri F, DiLeo J, Chung K, Cachat J, Goodspeed J, et al.The developing utility of zebrafish in modeling neurobehavioral disorders. Int J Comp Psychol 2010;23: 104–21.
Stewart AM, Cachat J, Green J, Gaikwad S, Kyzar E, Roth A, et al.Constructing the habituome for phenotype-driven zebrafish research. Behav Brain Res 2013;236: 110–7.
Stewart AM, Desmond D, Kyzar E, Gaikwad S, Roth A, Riehl R, et al.Perspectives of zebrafish models of epilepsy: what, how and where next? Brain Res Bull 2012;87: 135–43.
Stewart AM, Kalueff AV.The developing utility of zebrafish models for cognitive en-hancers research. Curr Neuropharmacol 2012;10:263–71.
Striepens N, Kendrick KM, Maier W, Hurlemann R.Prosocial effects of oxytocin and clin-ical evidence for its therapeutic potential. Front Neuroendocrinol 2011;32:426–50.
Suen MF, Chan WS, Hung KW, Chen YF, Mo ZX, Yung KK.Assessments of the effects of nic-otine and ketamine using tyrosine hydroxylase-greenfluorescent protein transgenic zebrafish as biosensors. Biosens Bioelectron 2013;42:177–85.
Teng BL, Nonneman RJ, Agster KL, Nikolova VD, Davis TT, Riddick NV, et al.Prosocial ef-fects of oxytocin in two mouse models of autism spectrum disorders. Neuropharma-cology 2013;72:187–96.
Thirtamara Rajamani K, Doherty-Lyons S, Bolden C, Willis D, Hoffman C, Zelikoff J, et al.
Prenatal and early-life exposure to high-level diesel exhaust particles leads to in-creased locomotor activity and repetitive behaviors in mice. Autism Res Off J Int Soc Autism Res 2013;6:248–57.
Thompson RR, Walton JC.Peptide effects on social behavior: effects of vasotocin and isotocin on social approach behavior in male goldfish (Carassius auratus). Behav Neurosci 2004;118:620–6.
Tropepe V, Sive HL.Can zebrafish be used as a model to study the neurodevelopmental causes of autism? Genes Brain Behav 2003;2:268–81.
Tuchman R.Autism and social cognition in epilepsy: implications for comprehensive ep-ilepsy care. Curr Opin Neurol 2013;26:214–8.
Veenstra-VanderWeele J, Muller CL, Iwamoto H, Sauer JE, Owens WA, Shah CR, et al.
Autism gene variant causes hyperserotonemia, serotonin receptor hypersensitivity, social impairment and repetitive behavior. Proc Natl Acad Sci U S A 2012;109: 5469–74.
Warnick JE, LaPorte JL, Kalueff AV.Domain interplay in mice and men: new possibilities for the“natural kinds”theory of emotion. New Ideas Psychol 2011;29:49–56.
Wing J, O'Connor N, Lotter V.Autistic conditions in early childhood: a survey in Middle-sex. Br Med J 1967;3:389.
Wong K, Elegante M, Bartels B, Elkhayat S, Tien D, Roy S, et al.Analyzing habituation re-sponses to novelty in zebrafish (Danio rerio). Behav Brain Res 2010a;208:450–7.
Wong K, Stewart A, Gilder T, Wu N, Frank K, Gaikwad S, et al.Modeling seizure-related behavioral and endocrine phenotypes in adult zebrafish. Brain Res 2010b;1348: 209–15.
Wright D, Rimmer LB, Pritchard VL, Krause J, Butlin RK.Inter and intra-population varia-tion in shoaling and boldness in the zebrafish (Danio rerio). Naturwissenschaften 2003;90:374–7.
Wright D, Nakamichi R, Krause J, Butlin RK.QTL analysis of behavioral and morphological differentiation between wild and laboratory zebrafish (Danio rerio). Behav Genet 2006;36:271–84.
Wright GJ, Washbourne P.Neurexins, neuroligins and LRRTMs: synaptic adhesion getting
fishy. J Neurochem 2011;117:765–78.