Synthesis of acarbose transfer products by
Bacillus
stearothermophilus
maltogenic amylase with simmondsin
Jin Sook Baek
a, Hye Young Kim
b, Seung Seok Yoo
a, Tae Kyou Cheong
a,
Myo Jeong Kim
a, Soo Bok Lee
c, Thomas P. Abbott
d, Hyun Joo Song
b,
Mee Ra Rhyu
b, Byung Ha Oh
e, Kwan Hwa Park
a,*
aDepartment of Food Science & Technology and Research Center for New Bio-Materials in Agriculture,Seoul National Uni6ersity, Suwon441-744,South Korea
bFood Chemistry & Biotechnology Di6ision,Korea Food Research Institute,Sungnam463-420,South Korea cCollege of Animal Husbandry,Konkuk Uni6ersity,Seoul143-701,South Korea
dNational Center for Agricultural Utilization Research,USDA,Peoria,IL61604,USA eDepartment of Life Science,Pohang Uni6ersity of Science & Technology,Pohang790-784,South Korea
Accepted 12 May 2000
Abstract
Simmondsin, a material related to food intake inhibition from jojoba (Simmondsia chinensis), was transglycosylated by Bacillus stearothermophilus maltogenic amylase (BSMA) reaction with acarbose to synthesize an antiobese compound with hypoglycemic activity. Ten percent each of acarbose and simmondsin were mixed and incubated with BSMA at 55°C. Glycosylation products of simmondsin were observed by thin layer chromatography (TLC) and high performance ion chromatography (HPIC). The major transfer product was purified by using Biogel P-2 column. The structure was determined by matrix-assisted laser desorption ionization with time of flight (MALDI-TOF)/mass spectrometry (MS) and13C-NMR. The major transglycosylation product was pseudotrisaccharide (PTS)-simmondsin,
in which PTS was attached by ana-(16) glycosidic linkage to simmondsin. The administration of transglycosylated simmondsin with acarbose (200 mg/kg per day for 6 days) significantly reduced the food intake by 74%, comparable to 62% of simmondsin versus control in ob/ob mice. The transfer product (10 mg/kg) significantly suppressed the postprandial blood glucose response to starch (2 g/kg) by 68%, comparable to 60% of acarbose in Zucker fa/fa rats. The results indicated that the transfer products would be effective agents in lowering both food intake and blood glucose. © 2000 Elsevier Science B.V. All rights reserved.
Keywords: Bacillus stearothermophilus maltogenic amylase (BSMA); Transglycosylation; Acarbose; Simmondsin; Food intake inhibition; Blood glucose
www.elsevier.com/locate/indcrop
* Corresponding author. Tel.: +82-31-2902582; fax: +82-31-2934789. E-mail address:[email protected] (K.H. Park).
J.S.Baek et al./Industrial Crops and Products12 (2000) 173 – 182 174
1. Introduction
Simmondsin, a jojoba constituent glycoside, is known to be a food intake inhibitor which can be obtained from deoiled seed. A number of studies on simmondsin dealt with the food intake reduc-ing effect (Cokelaere et al., 1992), physiological mechanism and its corresponding structure (Boven et al., 1994). The mechanism of food intake inhibition, however, is still unknown.
Obese people frequently are presented with or prone to associated dysregulation of glucose metabolism, while people with hyperglycemia are presented with or prone to associated dysregula-tion of body weight (Arnaud and Nathan, 1990). The features of these syndromes have generally
been neglected in the development of new preven-tive or treatment agents. Agents with a more holistic approach need to be developed. It would be more beneficial for human health, if the agents were developed from natural products or could be ingested as food rather than as a drug.
The transglycosylation reaction has been known to modify certain characteristics of sub-stances including glycosides. By the use of transfer reactions using distinct enzymes, a wide variety of transfer products have been obtained between a segment of a donor and various kinds of accep-tors. In general, the transglycosylation reaction takes place from a specific donor molecule to a relatively large number of different acceptors in structure. The specificity of the transfer seems to depend on the type of the enzyme, determining the configuration of the glycosidic bond formed between donor and acceptor molecule (Fu et al., 1990; Robyt, 1995; Park et al., 1998).
Enzymatic transglycosylation has many advan-tages over chemical methods for synthesizing many substances. These include simpler reaction steps, efficient purification procedures, and rela-tively high yields. A new type of amylase named
Bacillus stearothermophilus maltogenic amylase (BSMA) was obtained fromB.stearothermophilus
in the group, which has transglycosylation activity as well as hydrolytic activity (Cha et al., 1998). The maltogenic amylase can transfer mono- or disaccharide to the acceptor molecule by forming either a-(1,3)-, a-(1,4)- or a-(1,6)-glycosidic link-ages. The mechanism of transglycosylation was first illustrated by observation of an extra sugar binding space in the crystal structure of the en-zyme (Kim et al., 1999a).
Acarbose has been reported as a potent in-hibitor for several carbohydrases such as a -glu-cosidase, glucoamylase, a-amylase, and/or cyclodextrin glucanosyltransferase (CGTase). It is a pseudotetrasaccharide that has a pseudo sugar ring at the non-reducing end linked to the nitro-gen of 4-amino-4,6-dideoxy-D-glucopyranose, which is linked a-(14) to maltose (Schmidt et al., 1982; Aleshin et al., 1994; Brzozowski and Davies, 1997). Acarbose binds the active sites of a-amylase and CGTase in such a manner as to prevent cleavage of a-(1,4)-glycosidic linkages. Fig. 1. Thin layer chromatography (TLC) analysis of transfer
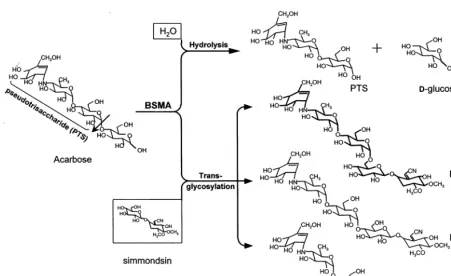
Fig. 2. Schematic summary of the reaction byBacillus stearothermophilusmaltogenic amylase (BSMA) with acarbose.
J.S.Baek et al./Industrial Crops and Products12 (2000) 173 – 182 176
Neither hydrolysis nor transglycosylation could occur. On the other hand, the maltogenic amylase from B. stearothermophilus (BSMA) binds to acarbose to cleave the first glycosidic linkage that gives transglycosylation of a pseudotrisaccharide (PTS) (Kang et al., 1997).
In the presence of various acceptor molecules, BSMA could transfer PTS to the acceptor molecule by forming a-(13), a-(14), or a -(16)-glycosidic linkages (Park et al., 1998). In this study, simmondsin was modified with acar-bose using the transglycosylation activity of BSMA. The transfer products were analyzed by thin layer chromatography (TLC) and purified using gel permeation chromatography. Also, the structure of the major transglycosylated product was determined by matrix-assisted laser desorp-tion ionizadesorp-tion with time of flight (MALDI-TOF)/ mass spectrometry (MS) and 1H- and 13C-NMR.
Enzyme inhibition of the transfer products, PTS-simmondsin, produced by BSMA, were evaluated ona-glucosidase anda-amylase. The in vivo effect of the transfer product was tested using obese and diabetic animal models.
2. Materials and methods
2.1. Materials
BSMA was obtained from recombinant Es
-cherichia coli DH5athat harbors plasmid pSG18, which originated in B. stearothermophilus ET1 (Cha et al., 1998). a-Glucosidase was prepared from rat intestine (Kim, 1997), and porcine pan-creatic a-amylase (No. A3176) was purchased from Sigma (St. Louis, MO). Acarbose was sup-plied by Bayer Korea (Seoul, South Korea) and simmondsin was purified in USDA, Peoria (IL, USA) as previously described (Abbott et al., 1999).
2.2. Enzyme assay
The activity of BSMA was assayed using 1% b-CD (b-cyclodextrin) in 50 mM Na-citrate buffer (pH 6.0). BSMA (100 ml) diluted in the same buffer were reacted with 500 ml of 1%b-CD and 400 ml of the buffer at 55°C for 30 min. The reaction was stopped by adding 1 ml of DNS
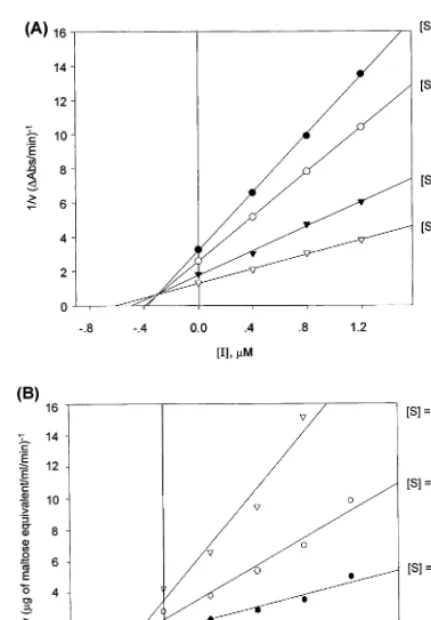
Fig. 5. Dixon plot of the reaction of (A) intestinala -glucosi-dase and (B) porcine pancreatica-amylase in the presence of the pseudotrisaccharide (PTS)-simmondsin.
For the kinetics of the enzyme inhibition, the activity of a-glucosidase from rat intestine was measured by a glucose oxidase/peroxidase method with a slight modification (Kim et al., 1999b). a-Amylase activity was assayed using the copper-bicinchoninate reducing-value method with a mi-crosample plate reader (Fox and Robyt, 1991). The kinetic parameters were calculated using a Dixon plot as well as the software for general enzyme inhibition by Duggleby (Duggleby and Leonard, 1992).
2.3. Transglycosylation of simmondsin with acarbose by BSMA
The reaction mixture of 10% (w/v) acarbose as a donor and 10% (w/v) simmondsin as an accep-tor was preincubated for 1 h at 55°C with 50 mM sodium citrate buffer (pH 6.0). One unit of BSMA per mg of acarbose was added to the mixture and the transglycosylation reaction was carried out for 48 h at 55°C. After boiling for 5 min to stop the reaction, it was centrifuged at 12 000×g for 5 min to remove the insoluble simmondsin. The reaction products were analyzed by TLC and high performance ion chromatography (HPIC) as follows.
The TLC analysis was performed using K5F Silica gel 150A TLC plate (Whatman, UK) with acetonitrile and water (8:2, v/v). After developing for two ascents, the plate was dried and visualized by dipping in 0.3% (w/v) N -(1-naphtyl)ethylene-diamine, 5% (v/v) H2SO4 in methanol, followed
by heating at 110°C for 10 min (Bounias, 1980). The transfer products were confirmed using HPIC after filtration through a 0.2 mm pore size membrane filter (Gelman Sciences). Twenty mi-solution, and colorized by heating at 100°C for 5
min. The absorbance of the mixture was measured at 575 nm using spectrophotometer (Ultrospec III, Pharmacia LKB, Uppsala, Sweden). One unit of the enzyme activity was defined as the amount of enzyme that produced 1 mmol of maltose at 55°C and pH 6 for 1 min (Miller, 1959; Cha et al., 1998).
Table 1
Acarbose and pseudotrisaccharide (PTS)-simmondsin inhibitor constants for rat intestinala-glucosidase and porcine pancreatic
a-amylase
Inhibitors
Enzymes Type of inhibition Ki(mM) Ki%(mM) Inhibitor potency
Acarbose
a-Glucosidase (rat intestinal) Competitive 0.05990.0059 1 PTS-simmondsin Competitive 0.30690.0244 0.2
0.64090.06
J.S.Baek et al./Industrial Crops and Products12 (2000) 173 – 182 178
Table 2
Effect of pseudotrisaccharide (PTS)-simmondsin, simmondsin and acarbose on the food intake inhibition and body weight change in lean and ob/ob mice for 6 daysa
Type of mice Groups Initial body weight Final body weight Body weight change Food intake (g/6 days) (g/6 days)
(g) (g)
Lean Control 25.6 26.6 1.0090.29 26.991.58
25.8 25.6
PTS-simmondsin 0.2290.29ab 23.690.74
26.5 26.6 0.0890.40ab 23.992.88
Acarbose
25.8 26.0 0.2090.24ab
simmondsin 27.292.07
Obese Control 51.0 53.4 2.3490.18a 51.090.79a
51.7 52.3
PTS-simmondsin 0.6090.31b 41.391.95b
52.4 53.3 0.9490.23b 55.993.15a
Acarbose
52.2 51.3 0.8890.31b 38.591.35b
Simmondsin
aValues are mean9S.E.M. (n=5). Different superscripts indicate statistically significant differences (PB0.05) within columns.
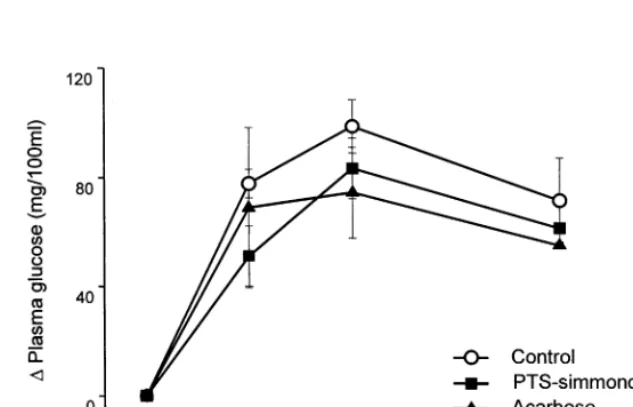
Fig. 6. Effect of pseudotrisaccharide (PTS)-simmondsin on glycemic response to starch in Zucker lean rats. (Rats received single oral dose of 2 g starch plus 10 mg/kg PTS-simmondsin or acarbose. Data are presented with S.E.M. (n=5). Superscript letters mean significant differences among groups.)
crolitres of the sample were applied to a Dionex Carbopak PA1 column (0.4×25 cm). The sample was eluted with a linear gradient from 100% A (150 mM NaOH in water) to 30% B (600 mM sodium acetate in A) over 30 min. The flow rate of mobile phase was maintained at 1.0 ml/ min.
2.4. Purification of the major transfer products
ma-jor transfer product was purified by preparative TLC. The purity of the major transfer product was verified by TLC and HPIC. The amount of PTS-simmondsin was estimated by UV ab-sorbance at 215 nm.
2.5. Determination of the structure of the transfer product
2.5.1. MALDI-TOF/MS analyses
MALDI-TOF/MS was used to measure the molecular weight of the major transfer product. A Voyager™-DE (PerSeptive Biosystem, Framing-ham, USA) system was adopted usinga -cyano-4-hydroxycinnamic acid as a matrix and des-Arg1-bradykinin for calibration. The purified
sample (1 ml) with the same aliquot of 2,5-dihy-drobenzoic acid was used for the determination of molecular weight of the transfer product.
2.5.2. 1H- and 13C-NMR analyses
The 1H- and 13C-NMR spectra of the major
transfer product were recorded with a JEOL LA-400 MHz NMR instrument. The sample was dis-solved in DMSO-d6 at 24.9°C with tetramethyl
silane (TMS) as the internal reference.
2.6. Kinetics of enzyme inhibition
The transfer products were preincubated with substrate for 20 min at 55°C prior to enzyme reaction. The enzyme reaction was performed ac-cording to the above reaction conditions with the inhibitors, acarbose and PTS-simmondsin, of var-ious concentrations. The types of inhibition were determined by Dixon plot, and its replot of slopes versus reciprocals of substrate concentrations. Ki-netic parameters including Vm, Km, and Ki were
evaluated by the non-linear regression method based on the following inhibition equation;
6i=Vm·S/[Km(1+I/Ki)+S(1+I/Ki%)]
where 6i is the initial velocity in the absence or presence of the inhibitor. SandI are the concen-tration of substrate and inhibitor, respectively.Vm
is the maximum velocity, Km is the Michaelis –
Menten constant, Ki is the inhibition constant,
which defined as [E][I]/[EI], and Ki% corresponds
to Kidefined as [ES][I]/[ESI] (Kim et al., 1999a).
2.7. Food intake inhibition in 6i6o
Eight-week-old male ob/ob (C57BL/6J-Lep; ge-netically obese and diabetic) and lean (ob/ + or
J.S.Baek et al./Industrial Crops and Products12 (2000) 173 – 182 180
?/ +) mice were obtained from Jackson Lab. (Bar Harbor, Maine, USA). The mice were housed individually in a temperature-controlled room at 24°C with a 12:12 h light – dark cycle, lights on at 07:00 h. PTS-simmondsin, simmondsin and acar-bose, prepared as a 20 mg/ml solution in water, were orally dosed into the mice. The dosage was 100 mg/kg two times per day (09:00 and 15:00 h), and fed for 6 days. Water and food were offered ad libitum. Food intake and body weight were measured at 09:00 h daily. Statistical significance of the four groups was analyzed by Duncan’s multiple range test (a=0.05).
2.8. Hypoglycemic effect in6i6o
Eight-week-old male Zucker fa/fa rats (obese and diabetic) and lean rats were provided from Harlan Sprague Dawley (Indianapolis, IN) to evaluate the postprandial glycemic effect of PTS-simmondsin. The housing was designed as previ-ously described. The overnight-fasted rats received soluble starch suspended in water (2 g starch/kg of body weight) by gastral intubation with or without PTS-simmondsin or acarbose (10 mg/kg BW). Blood samples were taken at 0, 0.5, 1 and 2 h after starch intubation. The glucose level in the plasma was measured by the glucose assay kit (Sigma Diagnostics, St. Louis, MO). The area under the glycemic response curve (AUC) was determined by the difference between the intu-bated glucose and the remaining glucose value in plasma. Total AUC was obtained by the zoidal rule. AUC represents the sum of the trape-zoidal areas and is expressed as mg×h/100 ml. Statistical significance of three groups was ana-lyzed by the Duncan’s test (a=0.05) in fa/fa and lean rats, respectively.
3. Results and discussion
3.1. Transglycosylation of simmondsin with acarbose by BSMA
Simmondsin was transglycosylated by BSMA, which has hydrolysis and transglycosylation activ-ities (Cha et al., 1998). In the absence of any
acceptor molecule, acarbose was hydrolyzed pri-marily to acarviosine-glucose (pseudotrisaccha-ride, PTS) and D-glucose. In the presence of D-glucose released from the hydrolysis, isoacar-bose was produced by transfer of PTS to D-glu-cose, resulting in formation of an a-(16) glycosidic bond (lane B in Fig. 1).
In the presence of simmondsin as an acceptor, however, one major product and one minor product were formed respectively (designated as 1 and 2) as shown in lane C of Fig. 1. After confirming the transfer products by TLC, the transfer products were purified using Bio-Gel P-2 gel filtration chromatography (lane D) and preparative TLC (lane E). The overall transglyco-sylation reaction of simmondsin with acarbose by BSMA is illustrated in Fig. 2.
3.2. Structure of the transfer product
The molecular weight of the purified transfer product was determined by MALDI-TOF/MS as shown in Fig. 3. It was estimated to be 840 Da from the two molecular ion peaks which appeared at m/z841 ([M+H]+) and 863 ([M+Na]+). The values corresponded to the calculated molecular mass of PTS-simmondsin hydrogen and sodium adducts, respectively. The results implied that the major transfer product by BSMA was PTS-sim-mondsin. The minor product was also purified and analyzed by HPIC and MALDI-TOF/MS. It was separated into two peaks by HPIC and gave identical molecular masses of 840 Da, which indi-cated that these two transferred products were also PTS-simmondsin with different linkages (data not shown).
The 1H- and 13C-NMR analyses were carried
out to determine the configuration of the glyco-sidic linkage between PTS and simmondsin. The chemical shifts in the 13
simmondsin, described as PTS-a -(1,6)-sim-mondsin.
3.3. Kinetics of enzyme inhibition
Enzyme inhibition by the transfer product, PTS-simmondsin, was evaluated against a -glu-cosidase anda-amylase by comparison with acar-bose (PTS generating donor). The inhibition plot for rat intestinal a-glucosidase by PTS-sim-mondsin is shown in Fig. 5A. The inhibition of PTS-simmondsin on a-glucosidase was competi-tive, and the Ki value was in the range of 0.28 –
0.33 mM. In the case of a-amylase, inhibition of the transferred product was competitive; how-ever, the Ki value decreased dramatically in the
range of 0.027 – 0.037mM (Fig. 5B).
The inhibitor potency compared to acarbose was calculated from the ratio of the inhibitor constant of acarbose to those of PTS-sim-mondsin. Table 1 summarizes the potency of each inhibitor as well as the respective dissocia-tion constants of the enzyme-inhibitor complex (EI) and the enzyme-substrate-inhibitor complex (ESI) designated as Ki and Ki%, respectively. The
transfer product, PTS-simmondsin, was 20 times more effective an inhibitor for a-amylase than was acarbose.
3.4. Food intake inhibition in 6i6o
In lean mice, PTS-simmondsin, simmondsin or acarbose did not reduce the food intake, but they suppressed the body weight gain to 0.22, 0.20 and 0.08 g, respectively shown in Table 2. In ob/ob mice, however, PTS-simmondsin and simmondsin significantly decreased the food in-take to 41.3 (19% reduction) and 38.5 g (25% reduction), respectively. Also body weight gain in PTS-simmondsin-, simmondsin-dosed groups was significantly depressed as 74 and 62% reduction. The result showed that ob/ob mice were more strongly affected by the administration of PTS-simmondsin than lean mice. This is fairly consis-tent with the report by Cokelaere et al. (1992).
Although the administration of acarbose did not reduce food intake, it inhibited the body weight gain of ob/ob mice by 60%.
William-Olsson (1985) also found that acarbose reduced body weight, and suggested it is associated with total food intake. More than 500 mg/kg could greatly affect the reduction of body weight in
animals and humans (Vasselli and
Haraczkiewicz, 1982). The result implies food in-take inhibition is not the only mechanism to reduce the body weight. There was no difference in body weight gain either by PTS-simmondsin or simmondsin.
3.5. Hypoglycemic effect in 6i6o
In Zucker lean rats, both PTS-simmondsin and acarbose (10 mg/kg) resulted in the same AUC reduction (20%), which represents glycemic re-sponse, as shown in Fig. 6. There was no signifi-cant difference among the change of plasma glucose from the 0 h value. However, in Zucker fa/fa rats, each PTS-simmondsin and acarbose significantly reduced AUC to 32 and 40%, re-spectively (Fig. 7). By PTS-simmondsin, each glucose level in plasma at 0.5, 1 and 2 h was significantly depressed as 41, 47 and 39%, respec-tively, while acarbose inhibited as 34, 46 and 37%. The result suggested that the hypoglycemic activity of PTS-simmondsin was as effective as that of acarbose.
J.S.Baek et al./Industrial Crops and Products12 (2000) 173 – 182 182
Acknowledgements
This work was supported in part by the HAN project grant from Ministry of Science and Tech-nology through Dae Sang, Ltd., and the Good Health 21 project grant from Ministry of Health and Welfare in Korea.
References
Abbott, T.P., Holser, R.A., Plattner, B.J., Plattner, R.D., Purcell, H.C., 1999. Pilot-scale isolation of simmondsin and related jojoba constituents. Ind. Crops Prod. 10, 65 – 72.
Aleshin, A.E., Firsov, L.M., Honzatko, R.B., 1994. Refined structure for the complex acarbose with glucoamylase from Aspergillus awamorivar. X100 to 2.4-angstroms resolution. J. Biol. Chem. 269, 15631 – 15639.
Arnaud, O., Nathan, C., 1990. Antiobesity and lipid-lowering agents with antidiabetic activity. In: Bailey, C.J., Flatt, P.R. (Eds.), New Antidiabetic Drugs. Smith-Gordon, Lon-don, UK, p. 133.
Bounias, M., 1980. N-(1-naphtyl)ethylenediamine dihy-drochloride as a new reagent for nanomole quantification of sugars on thin-layer plates by a mathematical calibra-tion process. Anal. Biochem. 106, 291 – 295.
Boven, M.V., Daenens, P., Cokelaere, M.M., Janssen, G., 1994. Isolation and structure elucidation of the major simmondsin analogues in jojoba meal by two-dimensional NMR spectroscopy. J. Agric. Food Chem. 42, 2684 – 2687. Brzozowski, B., Davies, M.J., 1997. Structure of the Asper -gillus oryzae alpha-amylase complexed with the inhibitor acarbose at 2.0 angstrom resolution. Biochemistry 36, 10837 – 10845.
Cha, H.J., Yoon, H.G., Kim, Y.W., Lee, H.S., Kim, J.W., Kweon, K.S., Oh, B.H., Park, K.H., 1998. Molecular and enzymatic characterization of a maltogenic amylase that hydrolyzes and transglycosylates acarbose. Eur. J. Biochem. 253, 251 – 262.
Cokelaere, M.M., Dangreau, H.D., Arnouts, S., Kuhn, E.R., Decuyere, E.M.P., 1992. Influence of pure simmondsin on the food intake in rats. J. Agric. Food Chem. 40, 1839 – 1842.
Duggleby, R.G., Leonard, D.R., 1992. DNRPEASY, a data analysis computer program.
Fox, J.D., Robyt, J.F., 1991. Modification of starch granules by hydrolysis with hydrochloric acid in various alcohols, and the formation of new kinds of limit dextrins. Anal.
Biochem. 195, 93 – 96.
Fu, D., Slodki, M.E., Robyt, J.F., 1990. Specificity of acceptor binding to Leuconostoc mesenteroides B-512F dextransu-crase: binding and acceptor-product structure of alpha-methyl-D-glucopyranoside analogs modified at C-2, C-3, and C-4 by inversion of the hydroxyl and by replacement of the hydroxyl with hydrogen. Arch. Biochem. Biophys. 276, 460 – 465.
Kang, G.J., Kim, M.J., Kim, J.W., Park, K.H., 1997. Immobi-lization of thermostable maltogenic amylase fromBacillus stearothermiphilusfor continuous production of branched oligosaccharides. J. Agric. Food Chem. 45, 4168 – 4172. Kim, H.Y., 1997. In vitro inhibitory activity on rat intestinal
mucosa a-glucosidase by rice hull extract. Korean J. Food Sci. Technol. 29, 601 – 608.
Kim, J.S., Cha, S.S., Kim, H.J., Kim, T.J., Ha, N.C., Oh, S.T., Cho, H.S., Cho, M.J., Kim, M.J., Lee, H.S., Kim, J.W., Choi, K.Y., Park, K.H., Oh, B.H., 1999a. Crystal structure of a maltogenic amylase provides insights into a catalytic versatility. J. Biol. Chem. 274, 26279 – 26286.
Kim, M.J., Lee, S.B., Lee, H.S., Lee, S.Y., Baek, J.S., Kim, D., Moon, T.W., Robyt, J.F., 1999b. Comparative study of the inhibition ofa-glucosidase,a-amylase, and cycloma-ltodextrin glucanosyltransferase by acarbose, isoacarbose, and acarviosin-glucose. Arch. Biochem. Biophys. 371, 277 – 283.
Miller, G.L., 1959. Use of dinitrosalycylic acid reagent for determination of reducing sugar. Anal. Chem. 31, 426 – 428.
Park, K.H., Kim, M.J., Lee, H.B., Han, N.S., Kim, D., Robyt, J.F., 1998. Transglycosylation reaction of Bacillus stearothermophilus maltogenic amylase with acarbose and various acceptors. Carbohydr. Res. 313, 235 – 246. Robinson, K.M., Rhinehart, B.L., Ducep, J.B., Danzin, C.,
1992. Intestinal disaccharidase inhibitors. Drugs Future 17 (8), 705 – 720.
Robyt, J.F., 1995. Mechanisms in the glucansucrase synthesis of polysaccharides and oligosaccharides from sucrose. Adv. Carbohydr. Chem. Biochem. 51, 151 – 157.
Schmidt, D.D., Frommer, W., Junge, B., Iler, L.M., Wingen-der, W., Truscheit, E., 1982. In: Creutzfeldt, W. (Ed.), First International Symposium on Acarbose. Excerpta Medica, Amsterdam, pp. 5 – 15.
Vasselli, J.R., Haraczkiewicz, E., 1982. Intestinal glucosidase inhibition: effects upon food intake and the development of obesity in 2 rat models. In: Creutzfeldt, W. (Ed.), Proceedings of First International Symposium on Acar-bose, October, 1981, Montreux. Excerpta Medica, Amster-dam, pp. 161 – 166.
William-Olsson, T., 1985. Glucosidase inhibition in obesity. Acta Med. Scand. 706, 1 – 39.